

Abstract
Preterm infants are especially susceptible to late-onset sepsis that is often due to Gram-positive bacterial infections resulting in substantial morbidity and mortality. Herein, we will describe neonatal innate immunity to Staphylococcus spp. comparing differences between preterm and full-term newborns with adults. Newborn innate immunity is distinct demonstrating diminished skin integrity, impaired Th1-polarizing responses, low complement levels, and diminished expression of plasma antimicrobial proteins and peptides, especially in preterm newborns. Characterization of distinct aspects of the neonatal immune response is defining novel approaches to enhance host defense to prevent and/or treat staphylococcal infection in this vulnerable population.
1. Introduction
Over 30% of deaths in children under the age of 5 occur within the first 4 weeks of life [1]. In this context, understanding the immunologic mechanisms underlying neonatal susceptibility is essential for the development of novel approaches to prevent and/or treat bacterial infection. Newborns in neonatal intensive care units (NICUs) are at risk of infection. An improvement in practices regarding hand washing, nutrition, skin, and respiratory care decreases Staphylococcus spp. infections [2, 3]. Such NICU quality improvements also significantly decrease costs per patient [4]. Antibiotics are the primary treatment for staphylococcal infections, but the use of these agents is also associated with resistance and alteration of the host microbial flora. Herein, we review innate host defense against Staphylococcus spp. with an emphasis on S. epidermidis (SE) and S. aureus (SA). Accordingly, we searched PubMed, a computer-based literature search engine, using the following terms: “newborn” OR “neonate” OR “neonatal” AND “Staphylococcus” AND/OR “sepsis” OR “innate” OR “cytokine” OR “Toll-like receptor” OR “pattern recognition receptor” OR “antimicrobial peptide” OR “neutrophil” OR “monocyte.” We then organized the resulting references grouping them into detector function, effector function, and translational efforts.
2. Neonatal Staphylococci Infections
SE colonizes newborns [5] and remains a part of the human normal flora [6, 7]. SE-induced sepsis in preterm newborns has been associated with an increased risk of adverse common outcomes, prolonged hospital stay, and increased costs [8–17]. SE is the most frequently isolated strain of coagulase-negative staphylococci (CoNS) and is identified diagnostically from SA because of its inability to produce coagulase [18, 19]. SE forms a biofilm on catheters and commonly infects immunocompromised patients [6, 20–22].
Invasive infections due to extracellular pathogens such as CoNS are largely restricted to premature infants. At the University Hospital of Patras in Greece, between 2006 and 2007, 8.5% of all NICU admissions had late-onset CoNS sepsis. SE was the most prevalent organism found, and biofilm production was identified as a determinant for persistent infection [23]. The majority of late-onset sepses (defined as 1 or more positive blood cultures obtained after 72 h of life) in very low birth weight (VLBW) newborns were caused by Gram-positive organisms [19, 24–26]. CoNS were responsible for 48% of infections in a study that examined over 6000 NICU infants in the USA [24]. Risk factors identified included low gestational age, low birth weight, an increased duration of ventilator support, central vascular catheter, and prolonged parenteral nutrition [24]. Close to half of the newborns that were <25 weeks of gestation developed late-onset sepsis and had a longer length of hospital stay [24]. Although CoNS infections often rapidly resolve with a few days of intravenous antibiotics, they are responsible for significant healthcare costs and also induce inflammatory responses that can sometimes result in long-term harm to the newborn, including potential cerebral injury [8–13, 27].
SA is the second leading cause of late onset sepsis in neonates [24]. SA leads to more prolonged bacteremia, dissemination to additional anatomic sites (e.g., osteomyelitis), and substantially more sepsis-related deaths than CoNS infections [28, 29]. SA-associated neonatal sepsis is associated with increased antibiotic resistance [28, 30, 31]. Newborns are often colonized with SA from their mothers via horizontal rather than a vertical transfer [32]. Accordingly, a study examining over 400 mothers admitted for preterm labor and the newborns subsequently admitted to the NICU in the USA found that vertical transmission of methicillin-resistant SA (MRSA) at the time of delivery was unlikely [33]. These findings suggested that there was a horizontal transfer of MRSA from health care workers or from parents while taking care of their infants [33]. Indeed, community-based MRSA strains have been identified in some NICU infections in the USA [28].
3. Innate Immune System in Neonates
Given the “in-born” nature of the innate immune response, it has been surprising that the innate immune response actually develops with age [34]. As has been recently reviewed, the innate immune response in neonates is distinct from that of older individuals [35, 36]. Multiple cells mediate innate immune responses, including skin and mucosal epithelia, neutrophils, monocytes/macrophages, and dendritic cells [35, 36]. The innate immune system also influences the adaptive immune response, and therefore understanding neonatal innate immunity may also inform development of age-specific vaccines.
3.1. Soluble Factors That Modulate Neonatal Immune Responses
Newborn plasma contains multiple factors that modulate the immune response [37]. Neonatal cord blood plasma has significantly more adenosine, an endogenous purine metabolite that inhibits Toll-like receptor (TLR)-mediated Th1 responses, than adult plasma [38]. The neonatal adenosine system inhibits TLR2-induced tumor necrosis factor (TNF) production but not interleukin (IL)-6 [38]. Serum of human newborns in the first week of life demonstrates a higher basal IL-6/TNF ratio than that of adults [39]. Moreover, when compared to monocytes of adults, neonatal cord blood monocytes produce a high ratio of IL-6 to TNF in response to TLR stimulation [39]. IL-6 can impair neutrophil production, migration, and function during sepsis [39–42] possibly contributing to the susceptibility of newborns to bacterial infection.
3.2. Antimicrobial Proteins and Peptides
A key mechanism by which the innate immune systems kill microbes and neutralize microbial toxins is via expression and mobilization of antimicrobial proteins and peptides (APPs) [43–45]. APPs are typically cationic molecules that have membrane-active effects on bacteria. Some APPs have additional function such as lactoferrin, which binds iron, a key nutrient for many bacteria [46], and lysozyme, which has enzymatic activity by muramidase that damages bacterial cell walls [47]. Defensins are small cationic antimicrobial peptides produced by leukocytes and epithelial cells in humans [48, 49]. Of note, preterm human neonates demonstrate deficient expression of plasma APPs that may contribute to the ability of bacteria to proliferate rapidly in preterm bloodstream. Moreover, newborn neutrophils demonstrate impairment in production of nucleic acid-based neutrophil extracellular traps (NETs) that serve as scaffolds for APPs and are important for host defense [50]. Overall, reduced plasma levels of complement and APPs as well as impaired deployment of APPs on NETs may, in part, explain why neonates are more susceptible to infection [51, 52].
3.3. Quantitative Differences in Phagocytes
Premature neonates admitted to the NICU have a relatively high frequency of neutropenia that can reach up to 8% [53]. In full-term newborns, impaired function of phagocytes has been described at birth [54]. Newborn neutrophils demonstrate impaired chemotaxis, phagocytosis, and impaired respiratory burst [54–57] and an impaired ability to form extracellular traps important for capture and killing extracellular bacteria [50].
3.4. Qualitative Differences in Leukocytes
The neonatal immune response is skewed towards Th2 and anti-inflammatory cytokine production. This may be important for protection of the fetus in utero and to avoid excessive inflammation during colonization with normal flora during the first days of life. Preterm newborns demonstrate mostly an anti-inflammatory response characterized by high IL-10 production while production of other cytokines is relatively low [58].
Inhibitory immune receptors antagonize cell-activating signals. Several of these inhibitory immune receptors function through immunoreceptor tyrosine-based inhibitory motifs (ITIMs). Newborn immune cells express a distinct pattern of inhibitory receptors compared to adult immune cells. Cord blood and 1-month-old newborn neutrophils express higher levels of the inhibitory receptors, leukocyte-associated immunoglobulin- (Ig-) like receptor-1 (LAIR-1), and siglec-9 than adults [59]. However, cord blood monocytes exhibited decreased expression of the immune receptor expressed on myeloid cells (IREM)-1, and 1-month-old newborn monocytes expressed lower levels of LAIR-1 compared to adults [59]. These observations suggest that neonatal neutrophils and monocytes are at a different basal set point from adult leukocytes.
4. Toll-Like Receptors (TLRs)
TLRs are pattern-recognition receptors (PRRs) of the innate immune system essential for early recognition of pathogen and also guide the adaptive immune response. There have been 10 TLRs identified in humans that signal through adaptor molecules such as myeloid differentiation factor-88 (MyD88) to activate transcription of immune mediators such as cytokines that direct the response to infection [60, 61]. While basal expression of TLRs is similar on full-term human newborn and adult monocytes [62–64], it can change with gestational age. Extremely low birth weight newborns (ELBW), <28 weeks of gestation, demonstrated lower expression of innate immune receptors TLR2, TLR4, CD14, and MD-2 on neutrophils [65]. Monocyte TLR4 mRNA and protein expression increase with gestational age [66]. In contrast, TLR2 expression is constitutively expressed on monocytes across gestational age and is therefore at similar levels in monocytes of preterms, full-term neonates, and adult monocytes [64]. Interestingly, Gram-positive bacteremia apparently induces increases in neonatal peripheral blood monocyte and granulocyte TLR2 expression in infected human newborns [67, 68].
Protein expression of MyD88, a cytosolic adaptor molecule essential TLR signaling, was decreased in newborn cord blood neutrophils [69] and monocytes [70] compared to those of adults. MyD88 mRNA levels increase in preterm infants cord blood mononuclear cells along gestational age. Preterm infants demonstrate lower MyD88 mRNA levels, but term infants are comparable to adults [64]; see Figure 1. Thus, there may be an inherent defect in newborns ability to make cytokine in response to infection due to a deficiency in this important signaling molecule.
Figure 1.
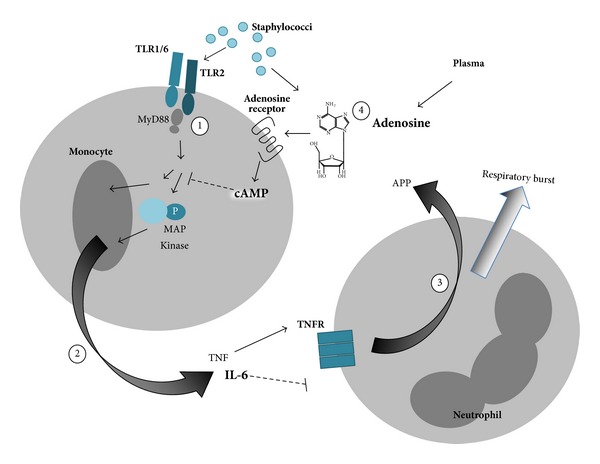
Sensor and effector function of neonatal innate immune system to Staphylococci spp. (1) Monocytes detect Staphylococci through TLR2 and (2) result in TLR-mediated production of proinflammatory cytokines such as TNF, and (3) TNF activates neutrophils to produce oxygen radicals and release APPs. (4) Both endogenous plasma and Staphylococcus-derived adenosine inhibit pro-inflammatory innate immune responses. Newborn monocytes are deficient in MyD88, activation of MAP kinases, and in TLR-mediated TNF but do produce robust amounts of IL-6, a proresolution cytokine that inhibits neutrophil migration. Overall, this pattern of response impairs neutrophil activation and migration and secretion of APPs. Deficiencies in neonatal responses to staphylococci are depicted by smaller font size, whereas agents that are elevated in newborns are indicated with a larger and bolded font.
A longitudinal study that examined TLR responses of individuals from birth to 2 years of age suggests that there is not a linear progression from an “immature” to “mature” innate immune response from newborns to adults [71]. The percentage of blood monocytes was higher in adults and newborns than 1- and 2-year-olds [71]. 2-year-olds demonstrated greater PAM3CSK4-(TLR2/1 agonist-) induced levels of intracellular cytokines than adults [71]. There was a higher percentage of 1- and 2-year-old classical (c)DCs making cytokine than adult cDCs [71]. Cytokines secreted from monocytes increased from birth to 2 years old for TNF and IL-1β; however, IL-6, IL-23, and IL-10 secretion decreased [71]. Preterm infants cord blood mononuclear cells have a significant defect in IL-12/IL-23p40 production in comparison to term infants after stimulation with TLR2/1 agonist PAM3CSK4, TLR2/6 agonist Fibroblast-stimulating lipopeptide (FSL), and TLR4 agonist LPS [58].
Micro- (mi-)RNAs involved in inhibiting the TLR4 signaling pathway are increased in newborn monocytes compared to adults and may contribute to decreased cytokine production [72]. Further investigation into the role miRNAs play into TLR2-signaling is warranted to gain further understanding of the potential role of miRNA in the neonatal innate immune response. Further research into other pattern-recognition receptors such as the NOD-like receptors and regulation of those receptors is warranted in the newborn to further understand neonatal staphylococcal-induced sepsis.
5. Staphylococcal Infections and Neonatal Host Immune Responses
TLR2 mediates innate immune responses to SE and is essential for clearance of SE in mice [73]. TLR2 also mediates the innate immune response to SA infection [74, 75]. Pretreatment of microglial cells with a TLR2 agonist decreased the inflammatory response to S. aureus but enhanced the microglial phagocytosis of this bacterium. Thus, TLR-modulation may be a useful treatment strategy to minimize inflammation in the eye [76].
When interpreting the literature of in vitro responses to staphylococci it is important to note that the immune response varies accordingly to whether the bacteria are heat-killed, ethanol-killed, or live [77]. Live SE induced significantly higher levels of cytokines compared to killed SE, including robust activation of the inflammasome for IL-1β production, induction of type I interferon production, nuclear factor (NF)κB, and signal transducers and activators of transcription (STAT)1 activation. In contrast, killed SE activated NFκB but did not activate the other innate immune pathways [77].
In a novel model of intrajugular infection in mice less than 24 hours of life, newborn mice demonstrate impaired weight gain when injected intravenously with SE compared to saline-injected controls [78]. Similar to the pattern noted in the peripheral blood mononuclear cells of preterm human newborns during Gram-positive bacteremia [67, 68], TLR2 and MyD88 mRNA levels in the liver were significantly increased by injection of SE that induced inocula-dependent serum IL-6 and TNF concentrations [78].
SE-induced cytokine production from human neonatal mononuclear cells (MCs) in vitro is dependent on gestational age [79–82]. Monocytes of preterm newborns demonstrate reduced IL-1β, IL-6, IL-8, and TNF production in response to SE despite adult-level TLR2 expression [83]. Impaired TNF production may contribute to impaired neutrophil responses to Staphylococcus spp. as TNF activates neutrophils. SE-induced phosphorylation of cell-signaling molecules (e.g., phospho-p65, phospho-p38 and phospho-JNK) was similar between newborns and adults [83]. In contrast, treatment of preterm neonatal monocytes demonstrated decreased SA lipoteichoic-acid- (LTA-) induced/TLR-mediated phosphorylation of p38 and ERK in whole blood [64]. LTA-induced production of IL-1β, IL-6, and IL-8 increased with gestational age [64].
Staphylococci spp. evade clearance by the immune system in part by generating adenosine, an endogenous purine metabolite that acts via cognate seven-transmembrane receptors to induce immunomodulatory intracellular cyclic adenosine monophosphate (cAMP; Figure 1), and therefore modulate the immune response [84]. Among the effects of adenosine is to boost production of IL-6, which can inhibit neutrophil migration [41, 42, 85–88] while inhibiting production of TNF important to neutrophil activation [38, 89–91]. Neonatal mononuclear cells are particularly sensitive to the effects of adenosine [38]. Accordingly, this adenosine generating effect of Staphylococci spp. may be particularly effective at disarming neonatal innate defense.
Opsonophagocytic Mechanisms. Human newborn and adult monocytes demonstrate similar phagocytic capacity and intracellular killing of SE [83]. However, preterm neonatal neutrophils demonstrate impaired SE-induced neutrophil oxidative burst compared to term newborns [57]. The plasma of premature neonates, especially extremely low birth weight (ELBW) newborns, had a lower opsonophagocytic capacity than term neonates and adults for SA [65].
The impact of these differences on the innate immune responses depending on age to SE and SA is that lower gestational age has a significant impact on the susceptibility of the individual to infection (Table 1). Since neonates have impaired sepsis-induced cytokine production, replenishing cytokines or APPs in neonates may be particularly helpful in the treatment of the preterm newborn. Knowing the deficiencies in the innate immune response may provide specific avenues for developing new treatments.
Table 1.
Differences in the innate immune response between preterm newborns, full-term newborns, infants, and adults in response to SE and SA.
| Preterm newborns | Full-term newborns | Adults | |
|---|---|---|---|
| Monocyte TLR2 expression | + | + | + |
| Monocyte MyD88 expression | ? | + | ++ |
| Phosphorylation of signaling molecules in response to G+ | + | ++ | ++ |
| Th1 cytokine expression | + | ++ | +++ |
| Neutrophil oxidative burst | + | ++ | ++ |
| Plasma opsonophagocytic capacity | + | ++ | ++ |
| Plasma antimicrobial proteins and peptides | + | ++ | +++ |
6. Potential Therapeutics
Although SE infections are often cleared from the newborn bloodstream within a few days of intravenous antibiotics (e.g., vancomycin), these infections can recur and are associated with substantial morbidity and healthcare costs [92–94]. Moreover, vancomycin resistance may be emerging [95]. Accordingly, additional preventative and therapeutic strategies are needed.
Injection of the S. simulans-derived metalloendopeptidase lysostaphin that cleaves crosslinking pentaglycine bridges in staphylococcal cell walls to MRSA-infected 2-day-old mice reduced bacterial load, improved neonatal weight gain, and enhanced survival similarly to vancomycin [96].
Another approach to addressing staphylococcal infection is to boost host defense by enhancing the quality of phagocytic responses in early life. In a study examining leukocytes from extremely premature infants (24–32 weeks of gestation), treating their leukocytes ex vivo with interferon (IFN)-γ reversed their innate immune deficiency [65]. Plasma from whole blood of ELBW newborns treated with IFN-γ significantly increased the phagocytosis of SA and SE by HL-60 cells [65]. This suggests that further studies are warranted to explore any potential therapeutic benefits for newborns. Administration of granulocyte-macrophage colony-stimulating factor (GM-CSF) to human newborns increased neutrophil production but had no impact on sepsis [97]. Treating septic very low birth weight infants with granulocyte (G)-CSF increased neutrophil phagocytic activity and oxidative burst but had no reported impact on sepsis due to the low number of sepsis patients in the study [98].
Since newborns have an increased susceptibility to sepsis, treating newborns with antibodies specific for SE and/or SA was examined. However, giving immunoglobulin intravenously from donors that had high titers of antibodies to SE and/or SA failed to significantly impact sepsis in preterm newborns [99–101]. However, the authors report a trend towards a decreased incidence ratio for SA infection in patients treated with antistaphylococcal antibodies suggesting that a higher-powered study would be required to examine efficacy more accurately [101].
7. Future Directions/Prospects
Many studies have documented late-onset sepsis in neonates due to staphylococcal infection. Current knowledge of the distinct immune system of preterm newborns provides at least three approaches to prevent and/or treat Staphylococcus spp. infections.
PRR Activation to Enhance Innate Antibacterial Defense. Activation of PRRs can change the set point of the innate immune system resulting in enhanced host defense in response to subsequent challenge with a range of pathogens. This phenomenon is a form of innate memory, that is, demonstrable in many life forms, including plants and insects and has been called “trained immunity” [102]. For example, intraperitoneal administration of a TLR agonist 24 hours prior to a polymicrobial peritonitis challenge markedly enhances neonatal defense and survival after subsequent polymicrobial sepsis by boosting bacteria-induced cytokine production and phagocytic function [103].
Use of TLR Antagonists as Adjunctive Anti-infective Therapy. In contrast to preexposure to TLR agonists to boost innate defense prior to an infection, a different strategy may be beneficial during an established infection. Antibiotic-killed bacteria are no longer viable but do continue to activate PRRs thereby inducing inflammation that can be harmful to multiple organ systems, including the brain [27]. Accordingly, adjunctive treatment with a TLR antagonist together with conventional antibiotics may help resolve infection-associated inflammation and reduce consequent morbidity of infection as has been demonstrated in vivo in other models and clinical settings [104, 105].
Use of APPs as Novel Anti-infective Agents. APPs with activity against Gram-positive bacteria include defensins, cathelicidins, lactoferrin and secretory phospholipase A2 [106, 107]. Biopharmaceutical development of APPs as novel anti-infective agents is proceeding, and replenishing deficient levels in preterm newborns either by direct infusion of APPs or by administration of agents that induce their expression may represent a promising approach to reduce infection.
Overall, further research on unique aspects of the neonatal host/staphylococcal pathogen interaction is warranted to assess the safety and efficacy of the aforementioned approaches and to identify new ones.
8. Discussion
This review has summarized recent studies of the innate immune response in preterm and full-term neonates compared to adults in response to SE or SA infection. We highlight important progress in defining the distinct innate immune response of newborns to Staphylococci spp. As there are currently limited strategies to address disease caused by these pathogens, it is hoped that recent progress in defining relevant host defense and pathogenic factors [108, 109] will inform new approaches to prevent and treat late onset sepsis due to Staphylococci spp.
Acknowledgments
OL's Laboratory is funded by the National Institutes of Health R01-AI100135-01 and by Bill & Melinda Gates Foundation Global Health Grants OPPGH5284 and OPP1035192 and has also received sponsored and reagent support from VentiRx Pharmaceuticals and 3 M Drug Delivery Systems. Melanie R. Power Coombs is currently funded by the Mitacs-Accelerate Internship Program.
References
- 1.Lawn JE, Cousens S, Zupan J. 4 million neonatal deaths: when? where? why? The Lancet. 2005;365(9462):891–900. doi: 10.1016/S0140-6736(05)71048-5. [DOI] [PubMed] [Google Scholar]
- 2.Horbar JD, Rogowski J, Plsek PE, et al. Collaborative quality improvement for neonatal intensive care. Pediatrics. 2001;107(1):14–22. doi: 10.1542/peds.107.1.14. [DOI] [PubMed] [Google Scholar]
- 3.Bersani I, Speer CP. Nosocomial sepsis in neonatal intensive care: inevitable or preventable? Zeitschrift für Geburtshilfe und Neonatologie. 2012;216(4):186–190. doi: 10.1055/s-0032-1321837. [DOI] [PubMed] [Google Scholar]
- 4.Rogowski JA, Horbar JD, Plsek PE, et al. Economic implications of neonatal intensive care unit collaborative quality improvement. Pediatrics. 2001;107(1):23–29. doi: 10.1542/peds.107.1.23. [DOI] [PubMed] [Google Scholar]
- 5.Keyworth N, Millar MR, Holland KT. Development of cutaneous microflora in premature neonates. Archives of Disease in Childhood. 1992;67(7):797–801. doi: 10.1136/adc.67.7_spec_no.797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Cheung GYC, Otto M. Understanding the significance of Staphylococcus epidermidis bacteremia in babies and children. Current Opinion in Infectious Diseases. 2010;23(3):208–216. doi: 10.1097/QCO.0b013e328337fecb. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Kloos WE, Musselwhite MS. Distribution and persistence of Staphylococcus and Micrococcus species and other aerobic bacteria on human skin. Journal of Applied Microbiology. 1975;30(3):381–395. doi: 10.1128/am.30.3.381-395.1975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Gray JE, Richardson DK, McCormick MC, Goldmann DA. Coagulase-negative staphylococcal bacteremia among very low birth weight infants: relation to admission illness severity, resource use, and outcome. Pediatrics. 1995;95(2):225–230. [PubMed] [Google Scholar]
- 9.Payne NR, Carpenter JH, Badger GJ, Horbar JD, Rogowski J. Marginal increase in cost and excess length of stay associated with nosocomial bloodstream infections in surviving very low birth weight infants. Pediatrics. 2004;114(2):348–355. doi: 10.1542/peds.114.2.348. [DOI] [PubMed] [Google Scholar]
- 10.Gonzalez A, Sosenko IRS, Chandar J, Hummler H, Claure N, Bancalari E. Influence of infection on patent ductus arteriosus and chronic lung disease in premature infants weighing 1000 grams or less. The Journal of Pediatrics. 1996;128(4):470–478. doi: 10.1016/s0022-3476(96)70356-6. [DOI] [PubMed] [Google Scholar]
- 11.Liljedahl M, Bodin L, Schollin J. Coagulase-negative staphylococcal sepsis as a predictor of bronchopulmonary dysplasia. Acta Paediatrica. 2004;93(2):211–215. doi: 10.1080/08035250310008168. [DOI] [PubMed] [Google Scholar]
- 12.Stoll BJ, Hansen NI, Adams-Chapman I, et al. Neurodevelopmental and growth impairment among extremely low-birth-weight infants with neonatal infection. Journal of the American Medical Association. 2004;292(19):2357–2365. doi: 10.1001/jama.292.19.2357. [DOI] [PubMed] [Google Scholar]
- 13.Martin CR, Dammann O, Allred EN, et al. Neurodevelopment of extremely preterm infants who had necrotizing enterocolitis with or without late bacteremia. The Journal of Pediatrics. 2010;157(5):751.e1–756.e1. doi: 10.1016/j.jpeds.2010.05.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Bizzarro MJ, Raskind C, Baltimore RS, Gallagher PG. Seventy-five years of neonatal sepsis at Yale: 1928–2003. Pediatrics. 2005;116(3):595–602. doi: 10.1542/peds.2005-0552. [DOI] [PubMed] [Google Scholar]
- 15.Isaacs D. A ten year, multicentre study of coagulase negative staphylococcal infections in Australasian neonatal units. Archives of Disease in Childhood. 2003;88(2):F89–F93. doi: 10.1136/fn.88.2.F89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Shane AL, Stoll BJ. Recent developments and current issues in the epidemiology, diagnosis, and management of bacterial and fungal neonatal sepsis. American Journal of Perinatology. 2013;30(2):131–141. doi: 10.1055/s-0032-1333413. [DOI] [PubMed] [Google Scholar]
- 17.Isayama T, Lee SK, Mori R, et al. Comparison of mortality and morbidity of very low birth weight infants between Canada and Japan. Pediatrics. 2012;130(4):e957–e965. doi: 10.1542/peds.2012-0336. [DOI] [PubMed] [Google Scholar]
- 18.Vuong C, Otto M. Staphylococcus epidermidis infections. Microbes and Infection. 2002;4(4):481–489. doi: 10.1016/s1286-4579(02)01563-0. [DOI] [PubMed] [Google Scholar]
- 19.Boghossian NS, Page GP, Bell EF, et al. Late-onset sepsis in very low birth weight infants from singleton and multiple-gestation births. The Journal of Pediatrics. 2013 doi: 10.1016/j.jpeds.2012.11.089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Mueller-Premru M, Gubina M, Kaufmann ME, et al. Use of semi-quantitative and quantitative culture methods and typing for studying the epidemiology of central venous catheter-related infections in neonates on parenteral nutrition. Journal of Medical Microbiology. 1999;48(5):451–460. doi: 10.1099/00222615-48-5-451. [DOI] [PubMed] [Google Scholar]
- 21.O’Gara JP, Humphreys H. Staphylococcus epidermidis biofilms: importance and implications. Journal of Medical Microbiology. 2001;50(7):582–587. doi: 10.1099/0022-1317-50-7-582. [DOI] [PubMed] [Google Scholar]
- 22.Okada Y, Klein NJ, van Saene HKF, Webb G, Holzel H, Pierro A. Bactericidal activity against coagulase-negative staphylococci is impaired in infants receiving long-term parenteral nutrition. Annals of Surgery. 2000;231(2):276–281. doi: 10.1097/00000658-200002000-00018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Dimitriou G, Fouzas S, Giormezis N, et al. Clinical and microbiological profile of persistent coagulase-negative staphylococcal bacteraemia in neonates. Clinical Microbiology and Infection. 2011;17(11):1684–1690. doi: 10.1111/j.1469-0691.2011.03489.x. [DOI] [PubMed] [Google Scholar]
- 24.Stoll BJ, Hansen N, Fanaroff AA, et al. Late-onset sepsis in very low birth weight neonates: the experience of the NICHD Neonatal Research Network. Pediatrics. 2002;110(2):285–291. doi: 10.1542/peds.110.2.285. [DOI] [PubMed] [Google Scholar]
- 25.Orsi GB, d’Ettorre G, Panero A, Chiarini F, Vullo V, Venditti M. Hospital-acquired infection surveillance in a neonatal intensive care unit. American Journal of Infection Control. 2009;37(3):201–203. doi: 10.1016/j.ajic.2008.05.009. [DOI] [PubMed] [Google Scholar]
- 26.Lim WH, Lien R, Huang YC, et al. Prevalence and pathogen distribution of neonatal sepsis among very-low-birth-weight infants. Pediatrics & Neonatology. 2012;53(4):228–234. doi: 10.1016/j.pedneo.2012.06.003. [DOI] [PubMed] [Google Scholar]
- 27.Wang X, Mallard C, Levy O. Potential role of coagulase-negative Staphylococcus infection in preterm brain injury. Advances in Neuroimmune Biology. 2012;3(1):41–48. [Google Scholar]
- 28.Healy CM, Hulten KG, Palazzi DL, Campbell JR, Baker CJ. Emergence of new strains of methicillin-resistant Staphylococcus aureus in a neonatal intensive care unit. Clinical Infectious Diseases. 2004;39(10):1460–1466. doi: 10.1086/425321. [DOI] [PubMed] [Google Scholar]
- 29.Mhada TV, Fredrick F, Matee MI, Massawe A. Neonatal sepsis at Muhimbili National Hospital, Dar es Salaam, Tanzania; aetiology, antimicrobial sensitivity pattern and clinical outcome. BMC Public Health. 2012;12, article 904 doi: 10.1186/1471-2458-12-904. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Ho TS, Wang SM, Wu YH, et al. Long-term characteristics of healthcare-associated infections in a neonatal intensive care unit. Journal of Microbiology, Immunology and Infection. 2010;43(5):407–415. doi: 10.1016/S1684-1182(10)60064-3. [DOI] [PubMed] [Google Scholar]
- 31.Kamath S, Mallaya S, Shenoy S. Nosocomial infections in neonatal intensive care units: profile, risk factor assessment and antibiogram. Indian Journal of Pediatrics. 2010;77(1):37–39. doi: 10.1007/s12098-010-0005-5. [DOI] [PubMed] [Google Scholar]
- 32.Jimenez-Truque N, Tedeschi S, Saye EJ, et al. Relationship between maternal and neonatal Staphylococcus aureus colonization. Pediatrics. 2012;129(5):e1252–e1259. doi: 10.1542/peds.2011-2308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lazenby GB, Soper DE, Beardsley W, Salgado CD. Methicillin-resistant Staphylococcus aureus colonization among women admitted for preterm delivery. American Journal of Obstetrics & Gynecology. 2012;206(4):329.e1–329.e5. doi: 10.1016/j.ajog.2012.01.038. [DOI] [PubMed] [Google Scholar]
- 34.Kollmann TR, Levy O, Montgomery RR, Goriely S. Innate immune function by Toll-like receptors: distinct responses in newborns and the elderly. Immunity. 2012;37(5):771–783. doi: 10.1016/j.immuni.2012.10.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Cuenca AG, Wynn JL, Moldawer LL, Levy O. Role of innate immunity in neonatal infection. American Journal of Perinatology. 2013;30(2):105–112. doi: 10.1055/s-0032-1333412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Levy O. Innate immunity of the newborn: basic mechanisms and clinical correlates. Nature Reviews Immunology. 2007;7(5):379–390. doi: 10.1038/nri2075. [DOI] [PubMed] [Google Scholar]
- 37.Belderbos ME, Levy O, Meyaard L, Bont L. Plasma-mediated immune suppression: a neonatal perspective. Pediatric Allergy and Immunology. 2012;24(2):102–113. doi: 10.1111/pai.12023. [DOI] [PubMed] [Google Scholar]
- 38.Levy O, Coughlin M, Cronstein BN, Roy RM, Desai A, Wessels MR. The adenosine system selectively inhibits TLR-mediated TNF-α production in the human newborn. The Journal of Immunology. 2006;177(3):1956–1966. doi: 10.4049/jimmunol.177.3.1956. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Angelone DF, Wessels MR, Coughlin M, et al. Innate immunity of the human newborn is polarized toward a high ratio of IL-6/TNF-α production in vitro and in vivo. Pediatric Research. 2006;60(2):205–209. doi: 10.1203/01.pdr.0000228319.10481.ea. [DOI] [PubMed] [Google Scholar]
- 40.Xing Z, Gauldie J, Cox G, et al. IL-6 is an antiinflammatory cytokine required for controlling local or systemic acute inflammatory responses. The Journal of Clinical Investigation. 1998;101(2):311–320. doi: 10.1172/JCI1368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Fielding CA, McLoughlin RM, McLeod L, et al. IL-6 regulates neutrophil trafficking during acute inflammation via STAT3. The Journal of Immunology. 2008;181(3):2189–2195. doi: 10.4049/jimmunol.181.3.2189. [DOI] [PubMed] [Google Scholar]
- 42.Jones SA. Directing transition from innate to acquired immunity: defining a role for IL-6. The Journal of Immunology. 2005;175(6):3463–3468. doi: 10.4049/jimmunol.175.6.3463. [DOI] [PubMed] [Google Scholar]
- 43.Levy O. Antimicrobial proteins and peptides of blood: templates for novel antimicrobial agents. Blood. 2000;96(8):2664–2672. [PubMed] [Google Scholar]
- 44.Levy O. Antimicrobial proteins and peptides: anti-infective molecules of mammalian leukocytes. Journal of Leukocyte Biology. 2004;76(5):909–925. doi: 10.1189/jlb.0604320. [DOI] [PubMed] [Google Scholar]
- 45.Levy O. A pediatric perspective on antimicrobial proteins and peptides: expression, function, and clinical relevance. In: Gallo RL, editor. Antimicrobial Peptides in Human Health and Disease. Norfolk, UK: Horizon Bioscience; 2005. pp. 305–329. [Google Scholar]
- 46.Vogel HJ. Lactoferrin, a bird’s eye view. Biochemistry and Cell Biology. 2012;90(3):233–244. doi: 10.1139/o2012-016. [DOI] [PubMed] [Google Scholar]
- 47.Callewaert L, Michiels CW. Lysozymes in the animal kingdom. Journal of Biosciences. 2010;35(1):127–160. doi: 10.1007/s12038-010-0015-5. [DOI] [PubMed] [Google Scholar]
- 48.Pinheiro da Silva F, Machado MC. Antimicrobial peptides: clinical relevance and therapeutic implications. Peptides. 2012;36(2):308–314. doi: 10.1016/j.peptides.2012.05.014. [DOI] [PubMed] [Google Scholar]
- 49.Lehrer RI, Ganz T. Defensins of vertebrate animals. Current Opinion in Immunology. 2002;14(1):96–102. doi: 10.1016/s0952-7915(01)00303-x. [DOI] [PubMed] [Google Scholar]
- 50.Yost CC, Cody MJ, Harris ES, et al. Impaired neutrophil extracellular trap (NET) formation: a novel innate immune deficiency of human neonates. Blood. 2009;113(25):6419–6427. doi: 10.1182/blood-2008-07-171629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Strunk T, Doherty D, Richmond P, et al. Reduced levels of antimicrobial proteins and peptides in human cord blood plasma. Archives of Disease in Childhood. 2009;94(3):F230–F231. doi: 10.1136/adc.2008.143438. [DOI] [PubMed] [Google Scholar]
- 52.Davis CA, Vallota EH, Forristal J. Serum complement levels in infancy: age related changes. Pediatric Research. 1979;13(9):1043–1046. doi: 10.1203/00006450-197909000-00019. [DOI] [PubMed] [Google Scholar]
- 53.Nittala S, Subbarao GC, Maheshwari A. Evaluation of neutropenia and neutrophilia in preterm infants. The Journal of Maternal-Fetal & Neonatal Medicine. 2012;25(supplement 5):100–103. doi: 10.3109/14767058.2012.715468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Filias A, Theodorou GL, Mouzopoulou S, Varvarigou AA, Mantagos S, Karakantza M. Phagocytic ability of neutrophils and monocytes in neonates. BMC Pediatrics. 2011;11, article 29 doi: 10.1186/1471-2431-11-29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Gessler P, Nebe T, Birle A, Haas N, Kachel W. Neutrophil respiratory burst in term and preterm neonates without signs of infection and in those with increased levels of C-reactive protein. Pediatric Research. 1996;39(5):843–848. doi: 10.1203/00006450-199605000-00017. [DOI] [PubMed] [Google Scholar]
- 56.Bektas S, Goetze B, Speer CP. Decreased adherence, chemotaxis and phagocytic activities of neutrophils from preterm neonates. Acta Paediatrica Scandinavica. 1990;79(11):1031–1038. doi: 10.1111/j.1651-2227.1990.tb11379.x. [DOI] [PubMed] [Google Scholar]
- 57.Björkqvist M, Jurstrand M, Bodin L, Fredlund H, Schollin J. Defective neutrophil oxidative burst in preterm newborns on exposure to coagulase-negative staphylococci. Pediatric Research. 2004;55(6):966–971. doi: 10.1203/01.pdr.0000127018.44938.89. [DOI] [PubMed] [Google Scholar]
- 58.Lavoie PM, Huang Q, Jolette E, et al. Profound lack of interleukin (IL)-12/IL-23p40 in neonates born early in gestation is associated with an increased risk of sepsis. Journal of Infectious Diseases. 2010;202(11):1754–1763. doi: 10.1086/657143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Walk J, Westerlaken GH, van Uden NO, Belderbos ME, Meyaard L, Bont LJ. Inhibitory receptor expression on neonatal immune cells. Clinical & Experimental Immunology. 2012;169(2):164–171. doi: 10.1111/j.1365-2249.2012.04599.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Kondo T, Kawai T, Akira S. Dissecting negative regulation of Toll-like receptor signaling. Trends in Immunology. 2012;33(9):449–458. doi: 10.1016/j.it.2012.05.002. [DOI] [PubMed] [Google Scholar]
- 61.Kawai T, Akira S. Toll-like receptors and their crosstalk with other innate receptors in infection and immunity. Immunity. 2011;34(5):637–650. doi: 10.1016/j.immuni.2011.05.006. [DOI] [PubMed] [Google Scholar]
- 62.Dasari P, Zola H, Nicholson IC. Expression of Toll-like receptors by neonatal leukocytes. Pediatric Allergy and Immunology. 2011;22(2):221–228. doi: 10.1111/j.1399-3038.2010.01091.x. [DOI] [PubMed] [Google Scholar]
- 63.Levy O, Zarember KA, Roy RM, Cywes C, Godowski PJ, Wessels MR. Selective impairment of TLR-mediated innate immunity in human newborns: neonatal blood plasma reduces monocyte TNF-α induction by bacterial lipopeptides, lipopolysaccharide, and imiquimod, but preserves the response to R-848. The Journal of Immunology. 2004;173(7):4627–4634. doi: 10.4049/jimmunol.173.7.4627. [DOI] [PubMed] [Google Scholar]
- 64.Sadeghi K, Berger A, Langgartner M, et al. Immaturity of infection control in preterm and term newborns is associated with impaired Toll-like receptor signaling. Journal of Infectious Diseases. 2007;195(2):296–302. doi: 10.1086/509892. [DOI] [PubMed] [Google Scholar]
- 65.Tissieres P, Ochoda A, Dunn-Siegrist I, et al. Innate immune deficiency of extremely premature neonates can be reversed by interferon-gamma. PLoS ONE. 2012;7(3) doi: 10.1371/journal.pone.0032863.e32863 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Förster-Waldl E, Sadeghi K, Tamandl D, et al. Monocyte Toll-like receptor 4 expression and LPS-induced cytokine production increase during gestational aging. Pediatric Research. 2005;58(1):121–124. doi: 10.1203/01.PDR.0000163397.53466.0F. [DOI] [PubMed] [Google Scholar]
- 67.Zhang JP, Yang Y, Levy O, Chen C. Human neonatal peripheral blood leukocytes demonstrate pathogen-specific coordinate expression of TLR2, TLR4/MD2, and MyD88 during bacterial infection in vivo. Pediatric Research. 2010;68(6):479–483. doi: 10.1203/PDR.0b013e3181f90810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Zhang JP, Chen C, Yang Y. Changes and clinical significance of Toll-like receptor 2 and 4 expression in neonatal infections. Zhonghua Er Ke Za Zhi. 2007;45(2):130–133. [PubMed] [Google Scholar]
- 69.Al-Hertani W, Yan SR, Byers DM, Bortolussi R. Human newborn polymorphonuclear neutrophils exhibit decreased levels of MyD88 and attenuated p38 phosphorylation in response to lipopolysaccharide. Clinical and Investigative Medicine. 2007;30(2):E44–E53. doi: 10.25011/cim.v30i2.979. [DOI] [PubMed] [Google Scholar]
- 70.Yan SR, Qing G, Byers DM, Stadnyk AW, Al-Hertani W, Bortolussi R. Role of MyD88 in diminished tumor necrosis factor alpha production by newborn mononuclear cells in response to lipopolysaccharide. Infection and Immunity. 2004;72(3):1223–1229. doi: 10.1128/IAI.72.3.1223-1229.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Corbett NP, Blimkie D, Ho KC, et al. Ontogeny of Toll-like receptor mediated cytokine responses of human blood mononuclear cells. PLoS ONE. 2010;5(11) doi: 10.1371/journal.pone.0015041.e15041 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Lederhuber H, Baer K, Altiok I, Sadeghi K, Herkner KR, Kasper DC. MicroRNA-146: tiny player in neonatal innate immunity? Neonatology. 2011;99(1):51–56. doi: 10.1159/000301938. [DOI] [PubMed] [Google Scholar]
- 73.Strunk T, Power Coombs MR, Currie AJ, et al. TLR2 mediates recognition of live Staphylococcus epidermidis and clearance of bacteremia. PLoS ONE. 2010;5(4) doi: 10.1371/journal.pone.0010111.e10111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Yoshimura A, Lien E, Ingalls RR, Tuomanen E, Dziarski R, Golenbock D. Cutting edge: recognition of Gram-positive bacterial cell wall components by the innate immune system occurs via Toll-like receptor 2. The Journal of Immunology. 1999;163(1):1–5. [PubMed] [Google Scholar]
- 75.Takeuchi O, Hoshino K, Akira S. Cutting edge: TLR2-deficient and MyD88-deficient mice are highly susceptible to Staphylococcus aureus infection. The Journal of Immunology. 2000;165(10):5392–5396. doi: 10.4049/jimmunol.165.10.5392. [DOI] [PubMed] [Google Scholar]
- 76.Kochan T, Singla A, Tosi J, Kumar A. Toll-like receptor 2 ligand pretreatment attenuates retinal microglial inflammatory response but enhances phagocytic activity toward Staphylococcus aureus . Infection and Immunity. 2012;80(6):2076–2088. doi: 10.1128/IAI.00149-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Strunk T, Richmond P, Prosser A, et al. Method of bacterial killing differentially affects the human innate immune response to Staphylococcus epidermidis . Innate Immunity. 2011;17(6):508–516. doi: 10.1177/1753425910379840. [DOI] [PubMed] [Google Scholar]
- 78.Kronforst KD, Mancuso CJ, Pettengill M, et al. A neonatal model of intravenous Staphylococcus epidermidis infection in mice LTHEXA24 h old enables characterization of early innate immune responses. PLoS ONE. 2012;7(9) doi: 10.1371/journal.pone.0043897.e43897 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Härtel C, Osthues I, Rupp J, et al. Characterisation of the host inflammatory response to Staphylococcus epidermidis in neonatal whole blood. Archives of Disease in Childhood. 2008;93(2):F140–F145. doi: 10.1136/adc.2007.124685. [DOI] [PubMed] [Google Scholar]
- 80.Peoples JD, Cheung S, Nesin M, et al. Neonatal cord blood subsets and cytokine response to bacterial antigens. American Journal of Perinatology. 2009;26(9):647–657. doi: 10.1055/s-0029-1220788. [DOI] [PubMed] [Google Scholar]
- 81.Tatad AMF, Nesin M, Peoples J, et al. Cytokine expression in response to bacterial antigens in preterm and term infant cord blood monocytes. Neonatology. 2008;94(1):8–15. doi: 10.1159/000112541. [DOI] [PubMed] [Google Scholar]
- 82.Mohamed MA, Cunningham-Rundles S, Dean CR, Hammad TA, Nesin M. Levels of pro-inflammatory cytokines produced from cord blood in-vitro are pathogen dependent and increased in comparison to adult controls. Cytokine. 2007;39(3):171–177. doi: 10.1016/j.cyto.2007.07.004. [DOI] [PubMed] [Google Scholar]
- 83.Strunk T, Prosser A, Levy O, et al. Responsiveness of human monocytes to the commensal bacterium Staphylococcus epidermidis develops late in gestation. Pediatric Research. 2012;72:10–18. doi: 10.1038/pr.2012.48. [DOI] [PubMed] [Google Scholar]
- 84.Thammavongsa V, Schneewind O, Missiakas DM. Enzymatic properties of Staphylococcus aureus adenosine synthase (AdsA) BMC Biochemistry. 2011;12, article 56 doi: 10.1186/1471-2091-12-56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Schwaninger M, Neher M, Viegas E, Schneider A, Spranger M. Stimulation of interleukin-6 secretion and gene transcription in primary astrocytes by adenosine. Journal of Neurochemistry. 1997;69(3):1145–1150. doi: 10.1046/j.1471-4159.1997.69031145.x. [DOI] [PubMed] [Google Scholar]
- 86.Xing Z, Gauldie J, Cox G, et al. IL-6 is an antiinflammatory cytokine required for controlling local or systemic acute inflammatory responses. The Journal of Clinical Investigation. 1998;101(2):311–320. doi: 10.1172/JCI1368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Fiebich BL, Biber K, Gyufko K, Berger M, Bauer J, van Calker D. Adenosine A2b receptors mediate an increase in interleukin (IL)-6 mRNA and IL-6 protein synthesis in human astroglioma cells. Journal of Neurochemistry. 1996;66(4):1426–1431. doi: 10.1046/j.1471-4159.1996.66041426.x. [DOI] [PubMed] [Google Scholar]
- 88.Ritchie PK, Spangelo BL, Krzymowski DK, Rossiter TB, Kurth E, Judd AM. Adenosine increases interleukin 6 release and decreases tumour necrosis factor release from rat adrenal zona glomerulosa cells, ovarian cells, anterior pituitary cells, and peritoneal macrophages. Cytokine. 1997;9(3):187–198. doi: 10.1006/cyto.1996.0153. [DOI] [PubMed] [Google Scholar]
- 89.Le Vraux V, Chen YL, Masson I, et al. Inhibition of human monocyte TNF production by adenosine receptor agonists. Life Sciences. 1993;52(24):1917–1924. doi: 10.1016/0024-3205(93)90632-d. [DOI] [PubMed] [Google Scholar]
- 90.Prabhakar U, Brooks DP, Lipshlitz D, Esser KM. Inhibition of LPS-induced TNFα production in human monocytes byadenosine (A2) receptor selective agonists. International Journal of Immunopharmacology. 1995;17(3):221–224. doi: 10.1016/0192-0561(94)00096-7. [DOI] [PubMed] [Google Scholar]
- 91.Ferrante A, Nandoskar M, Walz A, Goh DHB, Kowanko IC. Effects of tumour necrosis factor alpha and interleukin-1 alpha and beta on human neutrophil migration, respiratory burst and degranulation. International Archives of Allergy and Applied Immunology. 1988;86(1):82–91. doi: 10.1159/000234610. [DOI] [PubMed] [Google Scholar]
- 92.Hemels MAC, Verboon-Maciolek MA, Gerards LJ, Krediet TG, Fleer A. Rapid clearance from the bloodstream of coagulase-negative staphylococci in infants with late-onset sepsis. Pediatric Infectious Disease Journal. 2009;28(9):853–854. doi: 10.1097/INF.0b013e3181b20ea9. [DOI] [PubMed] [Google Scholar]
- 93.Wiederhold NP, Coyle EA, Raad II, Prince RA, Lewis RE. Antibacterial activity of linezolid and vancomycin in an vitro pharmacodynamic model of Gram-positive catheter-related bacteraemia. Journal of Antimicrobial Chemotherapy. 2005;55(5):792–795. doi: 10.1093/jac/dki106. [DOI] [PubMed] [Google Scholar]
- 94.Raad I, Hanna H, Jiang Y, et al. Comparative activities of daptomycin, linezolid, and tigecycline against catheter-related methicillin-resistant Staphylococcus bacteremic isolates embedded in biofilm. Antimicrobial Agents and Chemotherapy. 2007;51(5):1656–1660. doi: 10.1128/AAC.00350-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Rasigade JP, Raulin O, Picaud JC, et al. Methicillin-resistant Staphylococcus capitis with reduced vancomycin susceptibility causes late-onset sepsis in intensive care neonates. PLoS ONE. 2012;7(2) doi: 10.1371/journal.pone.0031548.e31548 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Placencia FX, Kong L, Weisman LE. Treatment of methicillin-resistant Staphylococcus aureus in neonatal mice: lysostaphin versus vancomycin. Pediatric Research. 2009;65(4):420–424. doi: 10.1203/PDR.0b013e3181994a53. [DOI] [PubMed] [Google Scholar]
- 97.Carr R, Brocklehurst P, Doré CJ, Modi N. Granulocyte-macrophage colony stimulating factor administered as prophylaxis for reduction of sepsis in extremely preterm, small for gestational age neonates (the PROGRAMS trial): a single-blind, multicentre, randomised controlled trial. The Lancet. 2009;373(9659):226–233. doi: 10.1016/S0140-6736(09)60071-4. [DOI] [PubMed] [Google Scholar]
- 98.Ahmad M, Fleit HB, Golightly MG, La Gamma EF. In vivo effect of recombinant human granulocyte colony-stimulating factor on phagocytic function and oxidative burst activity in septic neutropenic neonates. Biology of the Neonate. 2004;86(1):48–54. doi: 10.1159/000077585. [DOI] [PubMed] [Google Scholar]
- 99.DeJonge M, Burchfield D, Bloom B, et al. Clinical trial of safety and efficacy of IHN-A21 for the prevention of nosocomial staphylococcal bloodstream infection in premature infants. The Journal of Pediatrics. 2007;151(3):260.e1–265.e1. doi: 10.1016/j.jpeds.2007.04.060. [DOI] [PubMed] [Google Scholar]
- 100.Benjamin DK, Schelonka R, White R, et al. A blinded, randomized, multicenter study of an intravenous Staphylococcus aureus immune globulin. Journal of Perinatology. 2006;26(5):290–295. doi: 10.1038/sj.jp.7211496. [DOI] [PubMed] [Google Scholar]
- 101.Bloom B, Schelonka R, Kueser T, et al. Multicenter study to assess safety and efficacy of INH-A21, a donor-selected human staphylococcal immunoglobulin, for prevention of nosocomial infections in very low birth weight infants. Pediatric Infectious Disease Journal. 2005;24(10):858–866. doi: 10.1097/01.inf.0000180504.66437.1f. [DOI] [PubMed] [Google Scholar]
- 102.Netea MG, Quintin J, van der Meer JWM. Trained immunity: a memory for innate host defense. Cell Host and Microbe. 2011;9(5):355–361. doi: 10.1016/j.chom.2011.04.006. [DOI] [PubMed] [Google Scholar]
- 103.Wynn JL, Scumpia PO, Winfield RD, et al. Defective innate immunity predisposes murine neonates to poor sepsis outcome but is reversed by TLR agonists. Blood. 2008;112(5):1750–1758. doi: 10.1182/blood-2008-01-130500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Levin M, Quint PA, Goldstein B, et al. Recombinant bactericidal/permeability-increasing protein (rBPI21) as adjunctive treatment for children with severe meningococcal sepsis: a randomised trial. The Lancet. 2000;356(9234):961–967. doi: 10.1016/s0140-6736(00)02712-4. [DOI] [PubMed] [Google Scholar]
- 105.Guinan EC, Barbon CM, Kalish LA, et al. Bactericidal/permeability-increasing protein (rBPI21) and fluoroquinolone mitigate radiation-induced bone marrow aplasia and death. Science Translational Medicine. 2011;3(110) doi: 10.1126/scitranslmed.3003126.110ra118 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Weinrauch Y, Elsbach P, Madsen LM, Foreman A, Weiss J. The potent anti-Staphylococcus aureus activity of a sterile rabbit inflammatory fluid is due to a 14-kD phospholipase A2. The Journal of Clinical Investigation. 1996;97(1):250–257. doi: 10.1172/JCI118399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Yen CC, Lin CY, Chong KY, et al. Lactoferrin as a natural regimen for selective decontamination of the digestive tract: recombinant porcine lactoferrin expressed in the milk of transgenic mice protects neonates from pathogenic challenge in the gastrointestinal tract. Journal of Infectious Diseases. 2009;199(4):590–598. doi: 10.1086/596212. [DOI] [PubMed] [Google Scholar]
- 108.Otto M. MRSA virulence and spread. Cellular Microbiology. 2012;14(10):1513–1521. doi: 10.1111/j.1462-5822.2012.01832.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Otto M. Molecular basis of Staphylococcus epidermidis infections. Seminars in Immunopathology. 2012;34(2):201–214. doi: 10.1007/s00281-011-0296-2. [DOI] [PMC free article] [PubMed] [Google Scholar]