

Abstract
Episodic memory retrieval can be influenced by individuals’ current goals, including those that are emotional in nature. Participants underwent an fMRI scan while reappraising, or changing the way they thought about aversive images they had previously encoded, to down-regulate (i.e., decrease), up-regulate (i.e., increase), or maintain the emotional intensity associated with their recall. A conjunction analysis between down- and up-regulation during the entire 12-sec recall period revealed that both commonly activated reappraisal-related regions, particularly in the lateral and medial prefrontal cortex (PFC). However, when we analyzed a reappraisal instruction phase prior to recall and then divided the recall phase into the time when individuals were first searching for their memories and later elaborating on their details, we found that down- and up-regulation engaged greater neural activity at different time points. Up-regulation engaged greater PFC activity than down-regulation or maintenance during the reappraisal instruction phase. In contrast, down-regulation engaged greater lateral PFC activity as images were being searched for and retrieved. Maintaining the emotional intensity associated with the aversive images engaged similar regions to a greater extent than either reappraisal condition as participants elaborated on the details of the images they were holding in mind. Our findings suggest that down- and up-regulation engage similar neural regions during memory retrieval, but differ in the timing of this engagement.
Keywords: episodic memory, retrieval, emotion, emotion regulation, cognitive reappraisal
1. Introduction
Behavioral and neural investigations of emotion regulation have traditionally focused on individuals’ attempts and abilities to modify their emotional reactions about information in their external environments, such as a frightening image (see Ochsner & Gross 2008; 2005, for reviews). However, emotions also can be generated in response to internal cognitions, such as when we imagine a confrontation or recall a disturbing news story; just as we may be motivated to modify an emotional response to an external stimulus, we may also desire to change how we feel about these cognitions. Though there has been work demonstrating that the different emotional qualities of memories can influence the neural regions engaged during recall (e.g., LaBar & Cabeza, 2006), there has been a relative lack of work examining how one’s current emotional goals might guide the construction of past episodes. Therefore, the present study sought to examine the neural correlates of the emotion regulation of recalled information. Because it is widely studied and thought to be one of the most effective emotion regulation strategies (e.g., Gross & Thompson, 2007), we focus specifically on cognitive reappraisal, which involves the reinterpretation of emotional information in such a way as to enhance or reduce its impact (Gross, 1998).
When the target of regulation is external information, cognitive reappraisal is supported by cortical-subcortical interactions. Regions of the prefrontal cortex (PFC) and dorsal anterior cingulate cortex (dACC), thought to support the cognitive control processes necessary for both down- and up-regulation (Ochsner & Gross 2008; 2005), interact with emotion appraisal regions like the amygdala, insula, and orbitofrontal cortex (OFC; see Denny et al., in press, for review). The specific locations of PFC involvement appear to vary somewhat depending on the nature of reappraisal instructions (Ochsner & Gross, 2005; see also Kalisch, 2009); in general, dorsolateral areas may support selective attention toward less emotional aspects of information and the maintenance and manipulation of the information in working memory (Ochsner & Gross, 2008), ventrolateral regions are thought to underlie response inhibition and the selection of contextually-appropriate reappraisals (Ochsner & Gross, 2008; Denny et al., in press), and medial regions are engaged as individuals reflect upon their own or others’ emotional states (Ochsner & Gross, 2005).
There is less known about the processes that support the cognitive reappraisal of internally-generated memories, because the bulk of the work examining the relation between memory and emotion regulation has focused on regulation that takes place at the time of encoding (i.e., as an emotional stimulus is being presented). This work has demonstrated that cognitive reappraisal at the time of encoding can lead to mnemonic benefits (Richards & Gross, 2000; Richards et al., 2003; Dillon et al., 2007; Steinberger, Payne, & Kensinger, 2011), perhaps because common reappraisal techniques (e.g., making an emotional scene more or less self-relevant as a way to increase or decrease intensity, respectively) require deeper and more elaborative information processing. In line with this hypothesis, an fMRI investigation found evidence for enhanced co-activations in regions associated with elaborative encoding (inferior frontal gyrus, the hippocampus, and the amygdala) for subsequently remembered images whose intensity had been reappraised during encoding (Hayes et al., 2010). Similarly, an fMRI study that scanned participants during a recognition memory test for negative objects they had viewed 1-year prior found greater activity in cognitive control regions like the dorsolateral PFC for images that had been previously regulated vs. passively viewed (Erk et al., 2010). The authors hypothesized that during down-regulation, the non-emotional features of the objects received greater attention—perhaps as a regulation strategy—and were consequently more deeply encoded than the emotional content of the images. Those item features then provided a memory cue at recognition, perhaps explaining why the regulated images were associated with PFC activity (Erk et al., 2010).
Thus far, the work that has examined the reappraisal of retrieved memories has focused on how individuals regulate the emotions associated with autobiographical memory (AM) recall (e.g., Kross et al., 2009; Fabiansson et al., 2012; Holland & Kensinger, 2013; see also Cooney et al., 2007). Much like work with controlled laboratory stimuli, reappraisal appears to be an effective regulation technique in modifying the emotional intensity experienced upon AM recall (e.g., Denson, Moulds, & Grisham, in press). In addition, reappraisal that occurs during AM recall engages many of the same neural regions previously implicated in the reappraisal of emotional images and film clips (reviewed by Ochsner & Gross, 2005; 2008). Activation is evident in cognitive control-related regions in lateral PFC (Kross et al., 2009; Holland & Kensinger, 2013), as well as medial PFC and dACC (Holland & Kensinger, 2013), and in emotion appraisal-related regions like the amygdala and insula (Holland & Kensinger, 2013).
In the present experiment we sought to extend these prior findings by examining whether reappraisal influences neural activity in a similar manner during retrieval when the memories being recalled are not from a person’s autobiography but instead are memories of emotional images studied at an earlier point in time. In addition to having the benefit of using stimuli commonly used in studies of cognitive reappraisal (e.g., Ochsner et al., 2002; Bebko et al., 2011; McRae et al., 2009; Erk et al., 2010; see Kalisch, 2009, for meta-analysis), the present experiment also afforded us more precise control over encoding conditions and memorial content than is possible within autobiographical memory studies (e.g., all participants encoded the same images and all encoding occurred on the same day and for the same amount of time for each image). To examine the effect of reappraisal on the retrieval of these memories, we adapted a cognitive reappraisal task traditionally used during the presentation of emotional images (e.g., Ochsner et al., 2002) for use during the recall of those images.
An important point to keep in mind when considering the relation between regulation and memory recall is that both processes are dynamic and therefore not uniform across time. Indeed, one of the most striking outcomes of a recent investigation of the reappraisal of AM-generated emotions (Holland & Kensinger, 2013) was a divergence in the time course over which the down- and up-regulation of emotional intensity occurred. In that prior study, we took advantage of the extended time course over which recall occurs (e.g., Steinvorth et al., 2006; Addis et al., 2007) and examined the neural activity that was present during three separate phases: (a) an instruction phase during which time participants saw only a regulation cue (Increase, Decrease, or Maintain), (b) a memory search phase during which time participants retrieved an AM referenced by a self-generated memory cue created during a pre-scan interview, and (c) a memory elaboration phase during which time participants expounded upon the details of the AM they were holding in mind in accordance with the regulation instructions for that condition. Our analyses revealed that down-regulation engaged cognitive control and emotion processing regions previously associated with reappraisal primarily during the memory search phase. In contrast, up-regulation engaged similar regions during the instruction phase and once again during the elaboration phase.
The results of this prior investigation, taken together with two theoretical models of emotion regulation, highlight that timing matters in regulation and deserves further attention. The process model of emotion regulation (Sheppes & Gross, 2011) considers the effectiveness over time of various emotion regulation strategies like distraction, expressive suppression, and reappraisal. This model predicts that down-regulation via reappraisal is most effective early after an emotional stimulus is encountered and therefore before emotional intensity has had time to build and pass a “point of no return” after which point reappraisal is no longer a viable strategy (Sheppes & Gross, 2011; Sheppes & Meiran, 2007). If the process model is accurate, then it may explain why neural activity was greatest for down-regulation during the earlier memory onset phase in our prior study (Holland & Kensinger, 2013).
Though the process model (Sheppes & Gross, 2011) proposes timing differences for the effectiveness of distinct regulatory strategies, it is also important to note that reappraisal itself is not a unitary process and may be divided into distinct early and late phases (Kalisch, 2009). The implementation-maintenance model of reappraisal (IMMO; Kalisch, 2009) posits that down-regulation via reappraisal begins with an early phase during which time a reappraisal strategy is selected and implemented; this is followed by a later maintenance period during which time the outcome of the strategy implementation is monitored for its effectiveness. In the present study as well as in our prior study on autobiographical memory, we focused on separating the phases of memory recall rather than reappraisal in our study design. However, the IMMO also would predict that down-regulation recruits the greatest amount of neural activity early on, presumably during the memory onset phase, the time when a reappraisal strategy is being selected and implemented. Similarly, the IMMO predicts that once down-regulation is deemed successful, there is likely no need to continue maintaining and monitoring the selected strategy (Kalisch, 2009), which might translate to little neural activity during the later memory elaboration phase for down-regulation relative to either up-regulation or a maintenance condition.
Both the process model and the IMMO focus on down-regulation; neither addresses the timing of up-regulation via reappraisal. Based on the results of our prior experiment (Holland & Kensinger, 2013), we hypothesized that anticipatory neural activity during the up-regulation instruction phase may have facilitated access to negative memory details during the memory onset phase, thereby leading to relatively less memory search-related lateral PFC activity when compared to a down-regulation or maintenance condition. Subsequently, increased neural activity during the later memory elaboration phase may have reflected the use of expounding upon vivid sensory and perceptual details of an event as a strategy for increasing negative emotions. We sought to address whether these differences between the timing of down- and up-regulation would hold true when aversive images, rather than AMs, were recalled. We focus specifically on regions known to contribute to both episodic memory and emotion regulation processes (e.g., PFC, dACC, insula, and MTL).
2. Methods
2.1. Participants
Eighteen young adults (9 female; M = 22.11 years, SD = 3.68 years) who had no history of psychiatric, neurological, or learning disorders, or any history or current use of psychiatric medication participated in this study. Informed consent was obtained from all participants in accordance with the Boston College Institutional Review Board. Individuals who participated in this study did so immediately following the completion of a study examining the effects of emotion regulation on autobiographical memory recall reported elsewhere (Holland & Kensinger, 2013).
2.2. Pre-Scan Encoding Session
Approximately 1 week (range = 4–10 days; M = 6.61 days; SD = 1.51 days) prior to the scanning portion of the study, participants completed a pre-scan session in which they encoded a set of negative and neutral images that they were later asked to recall during the scan session. Participants viewed 36 aversive and 7 neutral images from the International Affective Picture System database (Lang et al., 1999) in the center of a computer screen in a randomized order. Participants were instructed to carefully study each image for a total of 10 sec, after which time they were prompted to write down a title that was just a few words but specific enough that if they were to see the title at the subsequent scan session they would be able to recall its associated image. After writing each title, participants made a button press to advance to the next image.
2.3. Scan Session
Of the aversive IAPS images encoded at the pre-scan session, 12 images were subsequently assigned to a Decrease condition, 12 to an Increase condition, and 12 to a Maintain condition. The images assigned to each condition were counterbalanced across participants and matched on their normative values of valence and arousal. Participants also viewed their titles for the 7 neutral images with a Maintain instruction and completed 8 trials of a sentence generation task adapted from Addis et al. (2009). These 15 filler trials were divided among 3 functional runs and interspersed with the negative image trials to prevent the propagation of a negative mood state that might influence regulation attempts or emotion ratings.
2.3.1. Emotion Regulation Task
Participants received instructions for the emotion regulation task immediately prior to the scanning portion of the study. They were instructed that upon seeing a “Decrease” or “Increase” instruction, they should attempt to cognitively reappraise the emotional intensity associated with the memory of the image referenced by their self-generated titles in such a way to feel a lesser or greater emotional reaction, respectively. Upon seeing a “Maintain” instruction, participants were instructed to recall the images referenced by their titles without attempting to change their emotional reaction. Example decrease and increase reappraisal strategies were given for images not used during the scan. For instance, when recalling the image of a man being held up at gunpoint, participants were instructed that they might increase their emotional reaction by imagining that they were in that situation and being held at gunpoint. If they had to decrease their emotional reaction, they might imagine that the image they recalled was simply a screenshot from a movie.
The 36 negative image titles (12 each of the Decrease, Increase, and Maintain) were pseudo-randomized such that no instruction appeared more than twice consecutively to avoid any global effects of a particular regulation or emotional state. These trials were divided among 3 functional runs along with the 15 filler trials (7 of the neutral image trials and 8 of the sentence generation trials); the structure for each trial is summarized in Fig. 1. Each trial began with a 3 sec fixation cross (jittered between 1–7 sec) during which time participants were instructed to relax. The fixation cross was followed by a reappraisal or maintain instruction (i.e., Decrease, Increase, or Maintain) for an average of 6 sec (jittered between 3–9 sec), during which time participants were instructed to prepare to regulate or maintain their emotions toward the image referenced by the subsequent memory title (corresponding to the “Instruction Phase”). A self-generated memory title then appeared below the regulation or maintain instructions for a maximum of 12 sec. Once participants felt they had the fully-formed image referenced by the memory cue in mind, they were instructed to make a button press to indicate so (corresponding to the “Memory Onset Phase”). Following the button press, participants continued to elaborate upon the details of the image they were holding in mind for the remainder of the 12 sec trial (corresponding to the “Memory Elaboration Phase”). Each trial ended with two 4 sec rating scales that asked participants to rate how emotionally intense and vivid their memory of the image was on a 7-point scale [1 (low) – 7 (high)].
Figure 1.
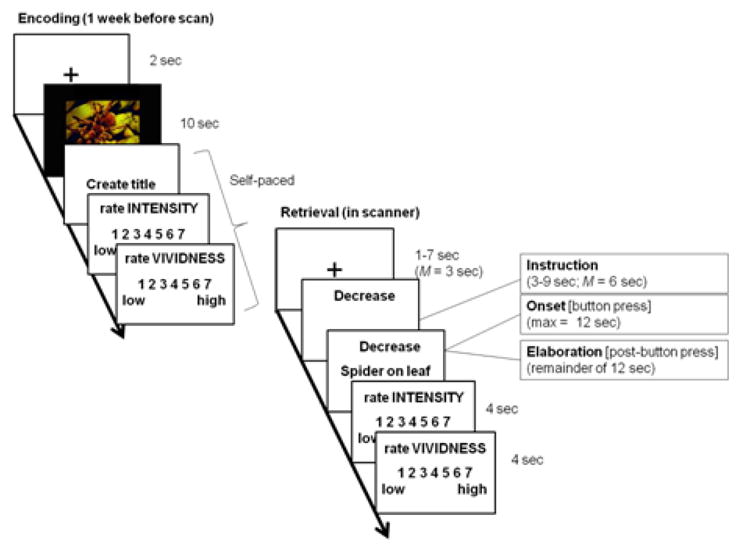
(Left) Example trial for a task in which individuals encoded aversive images at a prescan session and generated brief titles for those images. (Right) Example trial for a recall task in which individuals decreased, increased, or maintained the emotional intensity associated with aversive images while undergoing an fMRI scan.
2.4. Post-Scan Task
Following the scanning portion of the study, after an approximately 15-minute delay, participants completed a self-paced forced-choice recognition task. The IAPS images were presented with 3 thematically and visually similar foil images. Participants were given the title they had created for the image and asked to choose which, if any, of the images they had recalled in the scanner. For each image they reported recognizing, participants also rated the following on a 7-point scale [1 (low) – 7 (high)]: (a) emotional intensity, (b) positivity, and (c) negativity.
2.5. Scanning Parameters
Images were acquired on a 3 Tesla Siemens Tim Trio MRI scanner using a 32-channel head coil. Stimuli were presented using the MacStim presentation software in white text (Arial 36-point font) on a black background. Participants viewed the back-projected stimuli using a mirror attached to the head coil.
T1-weighted localizer images and a T1-weighted inversion recovery echo planar image required for auto-alignment were collected. Anatomic data were collected with a multi-echo multi-planar rapidly acquired gradient-echo (MEMPRAGE) sequence (TR = 2200 ms; TE = 1.64, 3.5, 5.46, 7.22 ms; flip angle = 7°; field of view = 256 X 256 mm; slice thickness = 1 mm, no gap; 1 X 1 X 1 mm resolution). Functional images were collected using a T2*-weighted echo-planar imaging (EPI) sequence with the following parameters: TR = 3000 ms, TE = 30 ms, FOV = 216 mm, flip angle = 85°. Forty-seven interleaved coronal-oblique slices aligned perpendicular to the long axis of the hippocampus were collected in a 3 mm3 matrix (slice thickness = 3 mm).
Preprocessing and data analysis were conducted in SPM8 (Wellcome Department of Cognitive Neurology, London). Preprocessing steps were as follows: (1) slice timing correction, (2) motion correction using a six parameter, rigid body transformation algorithm, (3) normalization to the Montreal Neurological Institute (MNI) template (resampling at 3 mm isotropic voxels), and (4) spatial smoothing using a 3 mm full-width half maximum isotropic Gaussian kernel.
2.5.1. Imaging data analysis
We modeled and analyzed event types in two separate ways. The first method treated the following events as 12-sec blocks (i.e., collapsing across the memory onset and elaboration phases), consistent with block analyses employed in previous reappraisal investigations (e.g., Ochsner et al., 2004): (a) Decrease, (b) Increase, and (c) Maintain Negative. We then conducted a conjunction analysis to establish that being instructed to reappraise the emotional intensity associated with recalled images activated regions previously implicated in the reappraisal of images as they were being viewed (e.g., Ochsner et al., 2004). For this conjunction analysis, we first computed first-level contrasts between the down-regulation trials and a random selection of one-half of the maintain negative trials (i.e., decrease > ½ of maintain negative trials) as well as between the up-regulation trials and the remaining one-half of the maintain negative trials (i.e., increase > ½ of maintain negative trials). The resulting contrast images were entered into a second-level random-effects one-sample t-test. A mask was created from the activated voxels in the first t-test (decrease > ½ maintain negative trials); this mask was then applied to the increase > ½ maintain negative trials t-test to reveal those regions that were commonly activated by the reappraisal trials. Each individual one-sample t-test used a threshold of p < .01 to ensure that the joint probability of the conjunction analysis was p < .001 using Fisher’s estimate (Fisher, 1950).
The second set of analyses used an event-related approach and segmented each trial into its Instruction, Memory Onset, and Memory Elaboration phases. Therefore, for each individual, the onsets of the following events were modeled and analyzed using the general linear model approach on a voxel-by-voxel basis: (a) Decrease Instruction, (b) Increase Instruction, (c) Maintain Negative Instruction, (d) Maintain Neutral Instruction, (e) Sentence Instruction, (f) Decrease Onset, (g) Increase Onset, (h) Maintain Negative Onset, (i) Maintain Neutral Onset, (j) Sentence Onset, (k) Decrease Elaboration, (l) Increase Elaboration, (m) Maintain Negative Elaboration, (n) Maintain Neutral Elaboration, (o) Sentence Elaboration. Contrasts between the various trial types were computed as described below, and the resulting contrast images were entered into second-level, random-effects analyses. To account for differences in recall latencies from trial to trial, we included RT as a parametric regressor for the onset phase on a trial-by-trial basis in the first-level analysis for each participant. In addition, to account for differences in neural activity related to variability in the delay between the pre-scan and scan sessions, we including the number of days between encoding and retrieval as a covariate in each second-level analysis. In all of our analyses, unless otherwise noted, we employed a statistical threshold of p < .001, uncorrected, and a 5-voxel threshold extent, shown by Monte Carlo simulations (Slotnick, Moo, Segal, & Hart, 2003) to be equivalent to a corrected p <.05. Because of our a priori hypotheses about how reappraisal instructions would modulate activity in the amygdala, we applied a small volume correction using an anatomically-defined mask of the bilateral amygdala from the MARINA toolbox (Walter, Blecker, Kirsch, Sammer, Schienle, et al., 2003). Any regions of the amygdala that resulted from this small volume correction are noted. Although in the text we focus specifically on regions of PFC, dACC, insula, and MTL, we report all regions in the Tables and Figures.
3. Results
3.1. Behavioral Results
Behavioral results are summarized in Table 1.
Table 1.
Behavioral characteristics of negative recalled images that participants were instructed to decrease, increase, or maintain during the scan session. Standard deviations are presented in parentheses.
| Scan Behavioral Ratings | |||
|---|---|---|---|
|
| |||
| Trial Type | RT (sec) | Intensity | Vividness |
| Decrease Neg | 3.32 (1.19) | 3.12 (0.82) | 4.62 (0.66) |
| Increase Neg | 3.46 (1.42) | 5.07 (0.84) | 5.33 (0.79) |
| Maintain Neg | 3.17 (1.16) | 3.99 (0.79) | 4.83 (0.72) |
3.1.1. Recognition
Performance on the post-scan memory task revealed that participants successfully recognized the correct negative images on 90% of trials. A within-subjects ANOVA confirmed that recognition performance did not depend on regulation condition, F(2, 34) = 0.98, p = .39, partial-χ2 = .06.
3.1.2. Reaction time
A within-subjects ANOVA examining the effect of emotion regulation condition on recall latencies was not significant, suggesting equivalent recall reaction times across conditions, F(2, 34) = 1.84, p = .17, partial-χ2 < .10.
3.1.3. Memory qualities
As anticipated, there was a main effect of emotion regulation condition on emotional intensity ratings made during the scan, F(2, 34) = 32.87, p < .001, partial-χ2 = .66. Recalled images in the decrease condition were rated as lowest in intensity, followed by those in the maintain condition, and then the increase condition, all ps < .003.
There was also a main effect of emotion regulation condition on vividness ratings made during the scan, F(2, 34) = 11.98, p < .001, partial-χ2 = .41. Recalled images in the increase condition were recalled with the greatest subjective vividness, all ps ≤ .005; there was also a trend for negative images in the maintain condition to be recalled with greater vividness than those in the decrease condition, p = .06.
3.1.4. Post-Scan Behavioral Ratings
Our final set of behavioral analyses examined the effect of emotion regulation condition on ratings made about the images during the recognition task following the scan (see Table 2). There was a main effect of condition on post-scan emotional intensity ratings, F(2, 34) = 7.10, p < .005, partial-χ2 = .29. Negative images whose cues had previously appeared with an up-regulation instruction continued to be rated as higher in emotional intensity than negative images that had appeared with either down-regulation or maintain instructions, ps < .02, though images previously associated with the decrease and maintain instructions did not differ from one another in emotional intensity following the scan, p = .90. The same main effect of condition was evident for post-scan negativity ratings, F(2,34) = 6.54, p < .005, partial-χ2 = .28. As with intensity ratings, image cues that had previously appeared with an up-regulation instruction continued to be rated as more negative than images whose cues had been associated with decrease or maintain instructions, ps < .03, though images whose cues appeared with the latter two instructions did not differ from one another in negativity ratings, p = .67. There was no effect of condition for post-scan positivity ratings, F(2,34) = 0.58, p = .57, partial-χ2 = .03.
Table 2.
Post-scan mean behavioral characteristics of negative images that had appeared with decrease, increase, or maintain instructions during the scan. Standard deviations are presented in parentheses.
| Post-Scan Behavioral Ratings | |||
|---|---|---|---|
|
| |||
| Trial Type | Intensity | Negative | Positive |
| Decrease Neg | 4.02 (0.78) | 4.55 (0.74) | 1.23 (0.40) |
| Increase Neg | 4.66 (1.05) | 5.08 (0.90) | 1.16 (0.25) |
| Maintain Neg | 4.00 (0.77) | 4.47 (0.87) | 1.19 (0.27) |
3.2. Imaging Results
3.2.1. Identifying Reappraisal-Related Regions: Blocked Design Analyses
We employed a conjunction analysis to examine which regions were commonly activated as individuals decreased and increased the emotional intensity associated with images they were cued to recall (Table 3). Our conjunction analysis confirmed that down- and up-regulation recruited common neural regions when the entire 12-sec recall trial was considered as a block. Both directions of reappraisal recruited a number of left-lateralized regions in the PFC, including ventrolateral (BA 47) and ventromedial (BA 10) PFC as well as superior and middle frontal gyri (BAs 6 and 8).
Table 3.
Group activations for the decrease and increase conjunction treating memory recall as a single 12-sec block of time.
| Lobe/Region | BA | H | Talairach | t | k | ||
|---|---|---|---|---|---|---|---|
|
| |||||||
| x | y | z | |||||
| Increase and Decrease Conjunction
| |||||||
| Frontal | |||||||
| Medial frontal gyrus | 10 | L | −10 | 44 | −11 | 3.86 | 11 |
| Superior frontal gyrus | 6 | L | −4 | 5 | 64 | 3.35 | 6 |
| 9/10 | L | −2 | 59 | 23 | 3.24 | 11 | |
| Middle frontal gyrus | 8 | L | −22 | 37 | 39 | 3.03 | 6 |
| Inferior frontal gyrus | 47 | L | −38 | 25 | −15 | 3.37 | 9 |
| 47 | R | 40 | 33 | −10 | 3.13 | 11 | |
| Temporal | |||||||
| Middle temporal gyrus | 21 | L | −55 | −24 | −9 | 4.46 | 7 |
| 39 | L | −46 | −61 | 27 | 3.71 | 7 | |
3.2.2. Emotion Regulation During Memory Recall: Event-Related Analyses
We next turned our attention to the neural activity present during each individual phase of our trials (i.e., the Instruction Phase, the Memory Onset Phase, and the Memory Elaboration Phase), using an event-related design to model these phases as individuals down- and up-regulated (vs. maintained) the negative emotions associated with their memories of the negative images.
3.2.2.1. Regulation During Recall: Instruction Phase
Reappraising vs. Maintaining Emotions
A conjunction analysis for the instruction phase did not reveal any common regions of activation between down- and up-regulation. When we examined the regions engaged as individuals prepared to decrease or increase vs. maintain the emotional intensity associated with a subsequent memory cue (Fig. 2a and 2b), the overall pattern of results revealed that viewing up-regulation instructions engaged the most extensive neural activity. In particular, when compared to the maintain instruction, up-regulation instructions engaged regions of right ventrolateral PFC (BA 47). The reverse contrast (maintain > increase instruction) did not reveal any activity above our statistical threshold. Similarly, viewing down-regulation (vs. maintain) instructions did not engage any regions above our statistical threshold, and viewing maintain (vs. decrease) instructions engaged regions of the cerebellum.
Figure 2.
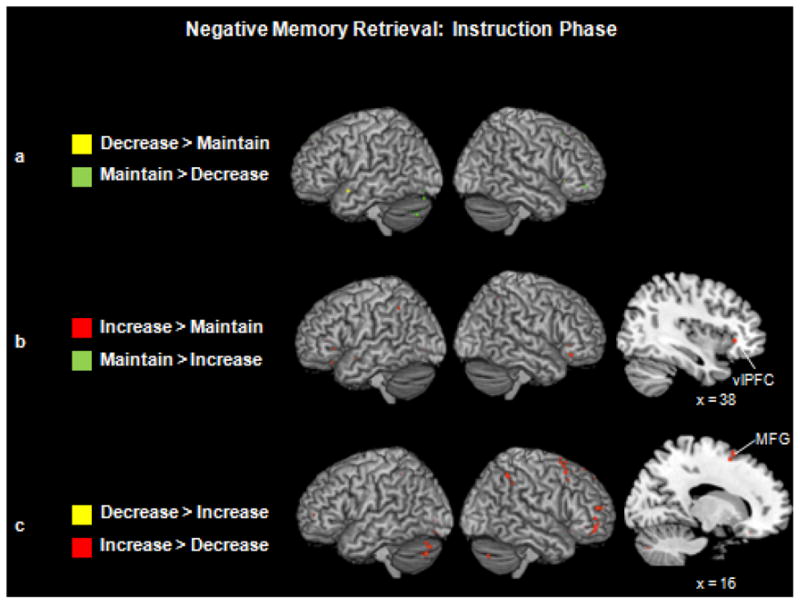
(a) Neural activity for the decrease and maintain trials during the reappraisal instruction phase. (b) Neural activity for the increase and maintain trials during the reappraisal instruction phase. Saggital slice shows a region of left left ventrolateral PFC (x = 38, y = 27, z = −3) that was more active as individuals were being instructed to increase vs. maintain their emotions. (c) Neural activity for the decrease and increase trials during the reappraisal instruction phase. Saggital slice shows a region of middle frontal gyrus (x = 16, y = 13, z = 58) that was more active when individuals were instructed to increase vs. decrease their emotions.
Down-regulation vs. Up-regulation
Directly comparing the decrease and increase conditions during the instruction phase confirmed that neural activity was greater during the increase than decrease instructions (Table 4; Fig. 2c). The increase > decrease contrast revealed several regions of right-lateralized PFC, including ventrolateral (BA 47) and medial (BA 8) PFC, frontal pole (BA 10), and middle (BA 6) and superior (BA 8) frontal gyri. In contrast, no regions survived our statistical threshold when examining the decrease > increase contrast during the instruction phase.
Table 4.
Group activations for the increase > decrease instruction contrast. Regions that were present in the Condition X Phase Interaction examining the onset and elaboration phases are noted in the last column with the distance between the peak evident in the t-test and the interaction.
| Lobe/Region | BA | H | Talairach | t | k | Condition X Phase Interaction | ||
|---|---|---|---|---|---|---|---|---|
|
| ||||||||
| x | y | z | ||||||
| Decrease > Increase Instruction | ||||||||
| No voxels survived even a liberal threshold of p < .005 and k > 5 | ||||||||
|
| ||||||||
| Increase > Decrease Instruction | ||||||||
| Frontal | ||||||||
| Inferior frontal gyrus | 10 | R | 48 | 49 | −1 | 4.32 | 5 | |
| Middle frontal gyrus | 11 | R | 48 | 46 | −14 | 4.72 | 16 | |
| 6 | R | 16 | 13 | 58 | 5.87 | 19 | ||
| 10 | R | 34 | 50 | −8 | 4.58 | 13 | ||
| 10 | L | −34 | 47 | 0 | 4.46 | 13 | ||
| 47/10 | R | 40 | 51 | 10 | 4.43 | 19 | ||
| Medial frontal gyrus | 8 | R | 2 | 49 | 42 | 4.45 | 5 | |
| Superior frontal gyrus | 8 | R | 30 | 16 | 49 | 4.43 | 10 | |
| Occipital | ||||||||
| Middle occipital gyrus | 19/18 | L | −30 | −79 | 6 | 6.73 | 16 | |
| 19 | L | −22 | −85 | 8 | 4.85 | 7 | ||
| Parietal | ||||||||
| Inferior parietal lobe | 40 | R | 46 | −48 | 48 | 4.66 | 17 | 10 mm* |
| Other | ||||||||
| Cerebellum | L | −24 | −69 | −30 | 6.27 | 154 | ||
| L | −4 | −77 | −21 | 5.05 | 69 | |||
| R | 10 | −77 | −23 | 5.66 | 22 | |||
| R | 36 | −76 | −32 | 4.40 | 10 | |||
| R | 18 | −69 | −22 | 4.40 | 6 | |||
| R | 2 | −39 | −6 | 3.85 | 7 | |||
|
| ||||||||
| R | 28 | −67 | −24 | 4.28 | 12 | |||
Present in the (decrease instruction > decrease onset) > (increase instruction > increase onset) interaction contrast.
3.2.2.2. Regulation During Recall: Memory Onset Phase
Reappraising vs. Maintaining Emotions
Despite the common activations evident when considering the memory onset and elaboration phases as a single block of time (section 3.2.1), a conjunction analysis revealed no statistically significant commonalities between down- and up-regulation when the memory onset phase was isolated. When we examined which regions were engaged by down- and up-regulation compared to the maintenance condition during the memory onset phase (Fig. 3a and 3b), the pattern was distinct from that seen during the instruction phase, in that the greatest activity occurred for the down-regulation condition. When comparing the decrease > maintain conditions, activity was evident in a number of primarily left-lateralized PFC regions, including ventral (BAs 11 and 47) and dorsal (BAs 44/45, 46 and 9) lateral areas as well as medial PFC (BAs 6 and 8). Recalling negative images associated with the decrease instruction also engaged a region of left dorsal anterior cingulate cortex (dACC; BA 32/24) to a greater extent than the maintain memory onset phase. No regions survived our statistical threshold when examining neural activity in the maintain > decrease contrast.
Figure 3.
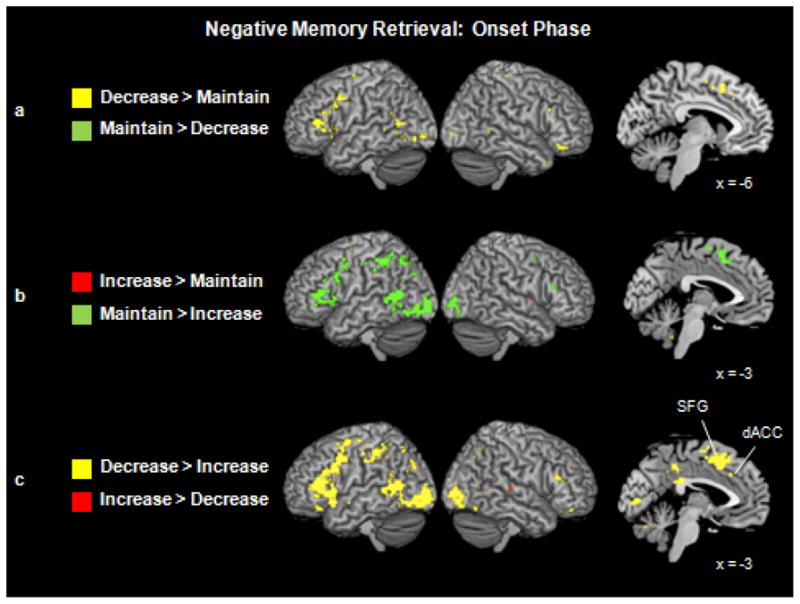
(a) Neural activity for the decrease and maintain trials during the memory onset phase. Saggital slice highlights mPFC activity that was greater as individuals decreased (vs. maintained) the negative emotions associated with their memories. (b) Neural activity for the increase and maintain trials during the memory onset phase. Saggital slice displays greater mPFC activity for the maintain than increase trials. (c) Neural activity for the decrease and increase trials during the memory onset phase. Saggital slice shows regions engaged to a greater extent by decrease than increase trials, including a region of dACC (x = −4, y = 25, z = 32) and superior frontal gyrus (x = −6, y = 14, z = 49).
In contrast to the widespread pattern of activity evident when comparing the decrease and maintain conditions, the increase condition did not engage any regions of the PFC, dACC, insula or MTL to a greater extent than the maintain condition during the memory onset phase. The pattern of activity evident in the maintain > increase condition was similar to the decrease > maintain contrast. In particular, the maintain condition engaged primarily left-lateralized regions of ventral (BAs 11 and 47) and dorsal (BAs 9, 45, and 46) lateral PFC, as well as medial PFC (BA 6).
Down-regulation vs. Up-regulation
Directly comparing the decrease and increase conditions confirmed that decreasing the emotional intensity associated with recalled negative images engaged the most widespread neural activity during the memory onset phase (Table 5; Fig. 3c). These regions included bilateral dorsal (BAs 9 and 46) and ventral (BAs 11 and 47) lateral PFC, bilateral medial PFC, and left middle (BA 6) and bilateral superior (BAs 6 and 8) frontal gyri. Activity was also evident in left dACC (BA 32) and right parahippocampal gyrus. In contrast, increasing the emotional intensity associated with recalled images did not engage any PFC, dACC, insula, or MTL regions to a greater extent than decreasing intensity during the memory onset phase.
Table 5.
Group activations for the decrease > increase onset contrast. Regions that were present in the Condition X Phase Interaction examining the onset and elaboration phases are noted in the last column with the distance between the peak evident in the t-test and the interaction.
| Lobe/Region | BA | H | Talairach | t | k | Condition X Phase Interaction | ||
|---|---|---|---|---|---|---|---|---|
|
| ||||||||
| x | y | z | ||||||
| Decrease > Increase Onset
| ||||||||
| Frontal | ||||||||
| Inferior frontal gyrus | 9 | L | −44 | 15 | 23 | 9.08 | 1812 | |
| 47 | L | −34 | 28 | −15 | 6.96 | 77 | 5 mm** | |
| 9 | R | 38 | 7 | 29 | 5.20 | 16 | 5 mm* and 3 mm** | |
| 46 | R | 51 | 28 | 15 | 5.09 | 26 | 6 mm* | |
| Middle frontal gyrus | 11 | R | 30 | 34 | −19 | 7.57 | 64 | 5 mm* |
| 6 | L | −34 | 12 | 53 | 4.87 | 6 | ||
| Medial frontal gyrus | 6 | L | −18 | 10 | 53 | 4.42 | 5 | |
| 6 | R | 10 | 5 | 64 | 4.29 | 6 | ||
| Superior frontal gyrus | 8 | L | −6 | 14 | 49 | 9.78 | 477 | 6 mm* |
| 6 | L | −12 | 22 | 52 | 5.16 | 6 | ||
| 6 | R | 12 | 8 | 53 | 4.82 | 30 | ||
| Precentral gyrus | 4 | L | −58 | −19 | 43 | 4.46 | 11 | |
| 6 | L | −40 | −13 | 58 | 4.35 | 9 | ||
| Temporal | ||||||||
| Middle temporal gyrus | 39 | L | −57 | −62 | 12 | 8.41 | 337 | 11 mm* and 12 mm** |
| 39 | L | −42 | −72 | 29 | 4.69 | 7 | ||
| Fusiform gyrus | 37 | L | −32 | −45 | −15 | 7.41 | 2197 | |
| 37 | R | 38 | −61 | −9 | 7.64 | 56 | ||
| 37 | R | 44 | −45 | −10 | 4.21 | 5 | ||
| Parahippocampal gyrus | 30 | R | 24 | −52 | 3 | 4.29 | 7 | |
| Occipital | ||||||||
| Cuneus | 18 | R | 14 | −71 | 15 | 6.87 | 70 | |
| 30 | L | −14 | −66 | 9 | 6.33 | 127 | 8 mm* | |
| Superior occipital gyrus | 39 | R | 32 | −68 | 29 | 5.52 | 46 | 3 mm* |
| Parietal | ||||||||
| Inferior parietal lobe | 40 | R | 36 | −56 | 45 | 6.81 | 93 | 4 mm* and 10 mm** |
| 40 | R | 42 | −43 | 41 | 4.15 | 5 | ||
| Superior parietal lobe | 7 | L | −24 | −52 | 41 | 6.23 | 388 | |
| 7 | L | −24 | −61 | 53 | 5.28 | 35 | ||
| Precuneus | 19 | L | −34 | −76 | 35 | 5.21 | 67 | |
| 7 | L | −10 | −70 | 40 | 4.31 | 7 | ||
| Limbic | ||||||||
| Cingulate gyrus | 31 | L | −4 | −39 | 42 | 5.44 | 41 | |
| 23 | R | 4 | −33 | 29 | 5.58 | 71 | 7 mm* | |
| 32 | L | −4 | 25 | 32 | 4.52 | 21 | ||
| 30 | R | 16 | −56 | 8 | 5.47 | 60 | ||
| Other | ||||||||
| Cerebellum | L | −8 | −79 | −25 | 6.34 | 83 | 9 mm** | |
| Thalamus | L | −6 | −15 | 6 | 4.98 | 28 | ||
| R | 8 | −13 | 8 | 4.15 | 6 | |||
|
| ||||||||
| Increase > Decrease Onset | ||||||||
|
| ||||||||
| Temporal | ||||||||
| Superior temporal gyrus | 22 | R | 63 | −25 | 7 | 5.81 | 11 | |
Present in the (decrease onset > decrease elaboration) > (increase onset > increase elaboration) interaction contrast.
Present in the (decrease onset > decrease instruction) > (increase onset > increase instruction) interaction contrast.
3.2.2.3. Regulation During Recall: Memory Elaboration Phase
Reappraising vs. Maintaining Negative Emotions
As with the instruction and memory onset phases, a conjunction analysis did not reveal any statistically significant commonalities between down- and up-regulation when the memory elaboration phase was considered in isolation from the other phases. When comparing the reappraisal and maintenance conditions during the memory elaboration phase (Fig. 4a and 4b), the most widespread neural activity was engaged by the maintain condition. The down-regulation > maintain contrast did not reveal any regions above our statistical threshold. In contrast, the maintain > down-regulation contrast revealed activity in right-lateralized ventrolateral PFC (BA 47) and dorsolateral PFC (BA 9) as well as left middle frontal gyrus (BA 6). Consistent with our predictions, the maintain condition also engaged a region of right insula to a greater extent than the decrease condition.
Figure 4.
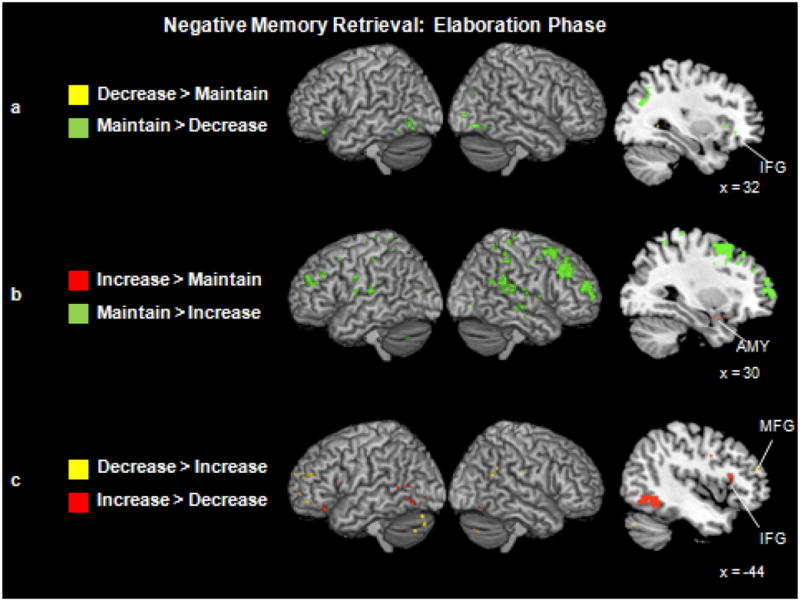
(a) Neural activity for the decrease and maintain trials during the memory elaboration phase. Saggital slice shows a region of right inferior frontal gyrus (x = 36, y = 7, z = 22) engaged more by the maintain than decrease trials. (b) Neural activity for the increase and maintain trials during the memory elaboration phase. Saggital slice shows a region of right amygdala (x = 30, y = −1, z = −15) engaged more by the increase than maintain trials. (c) Neural activity for the decrease and increase trials during the memory elaboration phase. Saggital slice highlights greater anterior PFC activity for the decrease condition (e.g., in middle frontal gyrus; x = −44, y = 44, z = 16) but greater ventrolateral PFC activity for the increase condition (e.g., in inferior frontal gyrus; x = −46, y = 18, z = 12).
Also in accordance with our predictions, a small volume correction revealed a cluster in the right amygdala that was more active as individuals increased than maintained their emotions. As with down-regulation, however, the maintain condition engaged a more widespread pattern of activity than up-regulation during this elaboration phase. Regions more active in the maintain than the up-regulation condition included the dorsolateral PFC (BA 9), frontal pole (BA 10) and medial PFC (BA 6) bilaterally, and the left dACC (BA 32). Contrary to our expectations, the maintain condition also engaged regions of insula bilaterally (BA 13) to a greater extent than the increase condition during the elaboration phase.
Down-regulation vs. Up-regulation
A direct comparison of the down- and up-regulation conditions during the elaboration phase revealed separate regions of the PFC that were engaged more by one condition or the other (Table 6; Fig. 4c). Down-regulation recruited more anterior regions of left PFC, including dorsolateral PFC (BA 46) and frontal pole (BA 10). On the other hand, up-regulation recruited more ventrolateral portions PFC bilaterally (BAs 11, 45, and 47).
Table 6.
Group activations for the decrease > increase elaboration contrast. Regions that were present in the Condition X Phase Interaction examining the onset and elaboration phases are noted in the last column with the distance between the peak evident in the t-test and the interaction.
| Lobe/Region | BA | H | Talairach | t | k | Condition X Phase Interaction | ||
|---|---|---|---|---|---|---|---|---|
|
| ||||||||
| x | y | z | ||||||
| Decrease > Increase Elaboration
| ||||||||
| Frontal | ||||||||
| Middle frontal gyrus | 46 | L | −44 | 44 | 16 | 4.83 | 7 | 4 mm* |
| Superior frontal gyrus | 10 | L | −28 | 54 | −3 | 5.69 | 24 | |
| 10 | L | −36 | 46 | −11 | 4.60 | 7 | ||
| Other | ||||||||
| Cerebellum | L | −32 | −81 | −25 | 5.13 | 27 | ||
| L | −32 | −79 | −16 | 4.77 | 10 | |||
| R | 30 | −77 | −28 | 4.71 | 15 | |||
|
| ||||||||
| Basal ganglia | R | 10 | −3 | 17 | 4.01 | 11 | ||
|
| ||||||||
| Increase > Decrease Elaboration | ||||||||
|
| ||||||||
| Frontal | ||||||||
| Inferior frontal gyrus | 47 | L | −38 | 28 | −15 | 4.93 | 9 | |
| 45 | L | −46 | 18 | 12 | 4.65 | 12 | ||
| Middle frontal gyrus | 11 | R | 28 | 34 | −13 | 4.33 | 5 | 5 mm** |
| Temporal | ||||||||
| Fusiform gyrus | 19 | R | 38 | −64 | −10 | 5.72 | 93 | |
| Inferior temporal gyrus | 19 | L | −44 | −55 | −6 | 5.49 | 121 | |
| Superior temporal gyrus | 22 | L | −55 | −50 | 8 | 4.33 | 5 | 5 mm** |
Present in the (decrease elaboration > decrease onset) > (increase elaboration > increase onset) interaction contrast.
Present in the (decrease onset > decrease elaboration) > (increase onset > increase elaboration) interaction contrast.
3.2.3. Condition x Phase Interaction
To further examine the possibility that down- and up-regulation engaged neural activity over different time courses during memory recall, we submitted the reappraisal > maintain first-level contrasts images to a second-level 2 (Condition: Increase, Decrease) x 3 (Phase: Instruction, Memory Onset, Memory Elaboration) repeated-measures analysis of variance (ANOVA). We then further explored the interaction results using contrasts that compared the instruction phase to the onset phase and the onset to the elaboration phase. Each contrast used an inclusive mask derived from the 2 x 3 ANOVA (mask p < .001) and a statistical threshold of p < .001 and a 5-voxel threshold extent.
The first contrast examined which regions were more active for the decrease condition in the memory instruction phase and the increase condition in the memory onset phase [i.e., (decrease instruction > decrease onset) > (increase instruction > increase onset)]. Consistent with the t-test results, no regions survived our statistical threshold. However, when the contrast was reversed to examine which regions were more active for the decrease condition in the memory onset phase and the increase condition in the memory instruction phase [i.e., (decrease onset > decrease instruction) > (increase onset > increase instruction)], a number of regions were revealed. In particular, activity was evident throughout primarily left-lateralized regions of dorsal (BAs 9/44 and 44/45) and ventral (BAs 11 and 47) lateral PFC and medial PFC (BA 6).
We also examined which regions were more active for the decrease condition in the memory onset phase and the increase condition in the memory elaboration phase [i.e., (decrease onset > decrease elaboration) > (increase onset > increase elaboration)]. Consistent with the t-test results reported above, this interaction contrast revealed activity throughout bilateral PFC, in dorsolateral (BAs 9/44 and 46), ventrolateral (BAs 11 and 47), and medial (BA 6) regions. However, when the interaction contrast was reversed to investigate which regions were more active for the increase condition in the memory onset phase and the decrease condition in the elaboration phase [i.e., (decrease elaboration > decrease onset) > (increase elaboration > increase onset)], only a region of right cerebellum was revealed.
4. Discussion
The present fMRI study examined the neural correlates of reappraisal that took place as individuals recalled aversive images that they had encoded a week prior to the scan. Emotional intensity ratings made as individuals recalled the images in the scanner confirmed that participants successfully reappraised their emotional responses when instructed to do so, and the post-scan ratings confirmed a lasting impact of reappraisal on the qualities of the memories. Although block design analyses revealed that there was overlap in the neural processes that underlie the down- and up-regulation of episodic memories, our findings also point to time course differences in neural activation depending on the direction of regulation. We expand on these points below.
4.1. Processes shared by down- and up-regulation
When we analyzed the entire 12-sec block of time that participants had to recall an image in response to a self-generated memory cue and to regulate their emotional intensity (i.e., using a conjunction analysis), we found that recalling images associated with both instructions recruited activity in PFC regions previously implicated in both the down- and up-regulation of images as they were being viewed (Ochsner et al., 2004). In particular, common activity was evident in several left-lateralized regions of lateral PFC that are thought to support the cognitive control processes necessary for reappraisal (Ochsner et al., 2004), dorsomedial PFC regions that are engaged when individuals monitor and reflect upon their emotional states during reappraisal (see review by Denny et al., in press), and ventromedial regions associated with emotional appraisal (Ochsner & Gross, 2005).
4.2. Down- and up-regulation of episodic memories differ in their time course
Though these regions previously associated with reappraisal were commonly activated by down- and up-regulation when the 12-sec recall period was considered as a block of time, considering a reappraisal instruction period prior to recall and then dividing recall into its onset (i.e., when individuals were searching for and retrieving the image from memory) and elaboration (i.e., when individuals were expounding upon the details of the image they had recalled) phases revealed that down- and up-regulation engaged neural activity at somewhat different time points.
4.2.1. Reappraisal Instruction Phase
Consistent with our prior findings in an AM investigation (Holland & Kensinger, 2013), the up-regulation condition recruited the greatest neural activity compared to the down-regulation and maintain conditions during the instruction phase. Interestingly, even though this was the phase before a memory cue was presented, much of the activation was evident in ventrolateral, orbitofrontal, and medial PFC regions that have previously been associated with the up-regulation of aversive images as they were being viewed (Ochsner et al., 2004) as well as with episodic memory retrieval (see review by Cabeza & Nyberg, 2000). As we discuss elsewhere (Holland & Kensinger, 2013), one possibility is that this anticipatory neural activity in the instruction phase for the up-regulation condition facilitated the subsequent retrieval and reappraisal of negative details about the images during the memory onset phase. This finding is also in accordance with research on pain perception demonstrating that employing a regulatory strategy when expecting an aversive event is associated with increased dorsolateral and medial PFC activity before that event is experienced (Herwig et al., 2007).
4.2.2. Memory Onset Phase
Although the instruction phase revealed the greatest neural activity during up-regulation, the increase condition recruited almost no neural activity to a greater extent than the decrease or maintain conditions during the memory onset phase, perhaps keeping in line with our hypothesis that the anticipatory activity during the instruction phase facilitated a relatively less demanding search for the images. Instead, the memory onset phase was associated with the greatest neural activity for the down-regulation condition, followed by the maintain condition, and finally by the up-regulation condition. The regions activated here during down-regulation are generally consistent with those recruited during reappraisal of aversive images or films as they are being viewed (Goldin et al., 2008; Ochsner et al., 2002; Phan et al., 2005; Ochsner et al., 2004) or during the down-regulation of negative, personally-relevant AMs (Holland & Kensinger, 2013). Specifically, relative to the other conditions, the decrease condition engaged several ventral and dorsolateral PFC and dACC regions implicated in the cognitive control necessary for the retrieval of episodic details (e.g., Cabeza & Nyberg, 2000) as well as the reappraisal of emotional information (see Denny et al., in press, for review). In addition, regions of dorsomedial PFC previously associated with self-reflection and the attention to and monitoring of emotional states (see Denny et al., in press, for review) were engaged during the down-regulation of negative memories; as we elaborate below, this activity may reflect emotional appraisal processes that occur as negative images are first being recalled. Finally, the decrease condition preferentially engaged MTL activity (in parahippocampal gyrus) previously associated with the recollection of emotional images (Dolcos, LaBar, & Cabeza, 2005).
The disproportionate PFC and dACC activity when down-regulating vs. up-regulating during the memory onset phase is particularly interesting. Although this disproportionate engagement seems to contrast with prior studies that have noted similar engagement of cognitive control regions activated for both down- and up-regulation of aversive images as they are being viewed (Ochsner et al., 2004), it is important to note that even in the present study, the conjunction analysis revealed extensive overlap within PFC regions when analyses collapsed across the memory onset and elaboration phases. Thus, this enhanced recruitment for down-regulation is specific to this onset phase of episodic memory retrieval, in keeping with our prior work demonstrating increased activity in these regions during the memory onset phase for the down-regulation of negative AMs (Holland & Kensinger, 2013).
It is possible that the enhanced neural activity during the memory onset phase for down-regulation trials fits with the process model of emotion regulation (Sheppes & Gross, 2011). If down-regulation is most effective soon after a stimulus is appraised as emotional, as the process model predicts (Sheppes & Gross, 2011), then it may follow that the bulk of the work in appraising and down-regulating the emotions associated with a memory occurs in the earlier memory onset phase, as the details of the memory are first being recalled.
Our participants were instructed to indicate with a button press when they had the recalled image in mind rather than when they had selected a reappraisal for the image, so we can only speculate as to how our findings fit with the IMMO’s predictions that the early phase of reappraisal involves selecting and implementing a reappraisal strategy (Kalisch, 2009). It is possible, however, that the differences for down- and up-regulation are related to timing differences in when a reappraisal strategy is initially selected and implemented. For down-regulation, the strategy may be selected as the content of a memory is first being retrieved.
4.2.3. Memory Elaboration Phase
Though the results from the instruction and memory onset phases were generally similar to our findings from a study using AM recall (Holland & Kensinger, 2013), the results from the elaboration condition were only partially so. Some of our findings were consistent with prior work: First, as would be expected based on its role in emotional appraisal (see meta-analysis by Sergerie, Chochol, & Armony, 2008), the amygdala was more active when participants were elaborating on the details of recalled images in the up-regulate condition than it was when they elaborated on images in the maintain condition. Second, in the line with our working hypothesis that down-regulation preferentially engages cognitive control regions during the memory onset phase, there was little neural activity that was greater for the decrease than the maintain condition during the elaboration phase. As mentioned in the Introduction, the IMMO predicts that once a reappraisal strategy is implemented and deemed to be appropriate, there may be little necessity for continued maintenance (Kalisch, 2009), possibly explaining why there was little neural activity in PFC or dACC evident for the down-regulation condition compared to the maintain condition during the later memory elaboration phase. Third, the differential recruitment of PFC activity revealed by the direct down- versus up-regulation contrast was partially in line with a prior investigation comparing down- and up-regulation of aversive images as they were being viewed (Ochsner et al., 2004). More specifically, down-regulation was associated with more anterior PFC activity than up-regulation, consistent with the OFC’s purported association with changing the affective value of information (e.g., generating a positive appraisal about a negative stimulus) and with its role in behavioral inhibition (Ochsner et al., 2004).
However, there were also key differences between the present results and prior work: During an AM elaboration phase, up-regulation revealed not only greater activity in emotion appraisal regions, but also enhanced activity throughout the PFC (Holland & Kensinger, 2013). In contrast, during the elaboration phase of memories for aversive images, the maintain condition showed the most extensive activity compared to both the down- or up-regulation conditions, particularly in cognitive control regions in lateral PFC and dACC. One possibility is that, similar to the down-regulation condition, much of the “work” in up-regulating the recalled images occurred before the elaboration phase, especially if the anticipatory activity in the instruction phase allowed for more efficient recall and reappraisal of emotional information. This suggestion might be reconciled with the regulation of AMs if we consider some of the behavioral differences between traditional laboratory memory stimuli and AM stimuli: AMs are typically more effortful to produce, more emotionally intense, and contain a greater quantity of vivid sensory details (Cabeza et al., 2004). Regulation during AM recall might therefore be more effortful and rely on the later elaboration phase to a greater extent than regulation during the recall of images encoded in the laboratory.
It is also possible that being instructed to maintain the emotions associated with aversive images led individuals to continue recalling and expounding upon details about the images throughout the elaboration phase, thereby engaging lateral PFC regions important for the retrieval and monitoring of episodic details (e.g., (Henson, Rugg, Shallice, & Dolan, 2000; Henson, Shallice, et al., 1999) to a greater extent than the regulation conditions. Indeed, one possible limitation of the present experiment is the use of the word “Maintain” in the non-reappraisal condition. This condition was meant to be analogous to a “View” condition in experiments where participants regulate their emotions to images that are currently being presented in that our participants were instructed to simply recall the details of the images in that condition as they normally would, without trying to alter their emotional reaction. It is plausible, however, that the “Maintain” instruction became a regulatory instruction unto itself by motivating participants to actively sustain the emotional intensity associated with recall throughout the onset and elaboration phases. If this were the case, it might shed light on why the maintain condition engaged PFC regions to a greater extent than the reappraisal conditions during the elaboration phase, though it is also notable that the use of the same instruction during AM recall (in the same participants) did not result in this pattern. However, in subsequent investigations it may be desirable to find an alternative instruction to use for this control condition (e.g., such as simply using a “Recall” instruction).
4.3. Future Directions
An open avenue for future work is to examine first how long-lasting any reappraisal-related changes in self-reported affect ratings and/or neural activity are when reappraised memories are retrieved in the future, and second what mechanism might underlie any durable changes. The post-scan recognition memory data suggest that reappraisal-related changes can be somewhat durable (see also Holland & Kensinger, 2013; MacNamara, Ochsner, & Hacjak, 2011; Shurick et al., 2012, for related evidence) but it is not clear whether such changes persist over time frames of days or weeks. One possibility is that when emotion regulation is employed as memories are reactivated and presumably in a labile state (Hardt, Einarsson, & Nader, 2010; McKenzie & Eichenbaum, 2011, for reviews), changes in emotional experience become associated and stored with the reconsolidated memory, leading to long-lasting changes. Conversely, it is possible that the reappraisals used during initial regulation attempts are associated with the memories – either explicitly or implicitly. Although the memory may cue the associated reappraisal, the strength of the association may dissipate over time, leading to shorter-acting effects of cognitive reappraisal.
Another open question concerns the timing of the reappraisal instruction given during recall. In the present study, the instruction was given prior to the memory cue, but the instruction could alternatively be given in between the memory onset and elaboration phases (i.e., once the memory has already been recalled and is being held in mind), or even after the elaboration phase. These types of design would better elucidate the effectiveness of using reappraisal as an emotion regulation strategy at different time points during and/or after memory retrieval. For example, it is possible that down-regulation is less effective if the reappraisal instruction is given after emotional details have already been recalled or elaborated upon, similar to the way that it is a less efficient regulation technique if implemented at later time points while viewing an emotional film clip (Sheppes & Gross, 2007). On the other hand, up-regulation might be even more effective if implemented after the emotional memory is already in mind.
4.4. Conclusion
The present study reveals that emotional goals are associated with differences in the time course over which neural regions are engaged as individuals recall aversive images. For example, having the goal to increase the emotional intensity associated with a past event recruits (a) greater neural activity in cognitive control regions before that specific event is even cued and (b) greater neural activity in emotional appraisal regions like the amygdala once the event is being held in mind. On the other hand, decreasing the emotional intensity associated with the recall of aversive information recruits greater neural activity in both cognitive control and emotional appraisal regions as that information is first being retrieved. Taken together, these results suggest that adopting a particular mindset prior to recall may facilitate the up-regulation of emotions, whereas down-regulation relies on processes during the early retrieval and appraisal of event details. More broadly, our results demonstrate that employing emotion regulation strategies at the time of memory retrieval can modulate the subjective experiences and neural activity associated with recall and provide an example of at least one type of goal that can influence recall at both a behavioral and neural level. Indeed, this ability to flexibly reappraise and update our evaluations of past emotional episodes may be an important function of the constructive nature of memory.
Highlights.
Individuals reappraised the emotions associated with the recall of aversive images
Down- and up-regulation commonly engaged lateral and medial PFC during recall
Down-regulation was linked to the greatest neural activity during memory search
Up-regulation was linked to anticipatory activity prior to a memory cue
Acknowledgments
This research was supported by NIH grant MH080833 to EAK and a National Defense Science and Engineering Graduate Fellowship to ACH. We thank Tammy Moran and Ross Mair at the Harvard Center for Brain Science for their assistance with MR data collection and Daniel Schacter, Scott Slotnick, and Ehri Ryu for helpful discussion.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Addis DR, Wong AT, Schacter DL. Remembering the past and imagining the future: Common and distinct neural substrates during event construction and elaboration. Neuropsychologia. 2007;45:1363–1377. doi: 10.1016/j.neuropsychologia.2006.10.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bebko GM, Franconeri SL, Ochsner KN, Chiao JY. Look before you regulate: Differential perceptual strategies underlying expressive suppression and cognitive reappraisal. Emotion. 2011;11:732–742. doi: 10.1037/a0024009. [DOI] [PubMed] [Google Scholar]
- Cabeza R, Nyberg L. Imaging cognition II: An empirical review of 275 PET and fMRI studies. Journal of Cognitive Neuroscience. 2000;12:1–47. doi: 10.1162/08989290051137585. [DOI] [PubMed] [Google Scholar]
- Cabeza R, Prince SE, Daselaar SM, Greenberg D, Budde M, Dolcos F, LaBar KS, Rubin DC. Brain activity during episodic retrieval of autobiographical and laboratory events: An fMRI study using a novel photo paradigm. Journal of Cognitive Neuroscience. 2004;9:1533–1594. doi: 10.1162/0898929042568578. [DOI] [PubMed] [Google Scholar]
- Cooney RE, Joormann J, Atlas JY, Eugène F, Gotlib IH. Remembering the good times: neural correlates of affect regulation. Neuroreport. 2007;18:1771–1774. doi: 10.1097/WNR.0b013e3282f16db4. [DOI] [PubMed] [Google Scholar]
- Denny B, Silvers J, Ochsner KN. How we heal what we don't want to feel: The functional neural architecture of emotion regulation. In: Kring AM, Sloan DM, editors. Emotion regulation and psychopathology: A transdiagnostic approach to etiology and treatment. New York: Guilford Press; pp. 59–87. in press. [Google Scholar]
- Denson TF, Moulds ML, Grisham JR. The effects of rumination, reappraisal, and distraction on anger experience. Behavioral Therapy. doi: 10.1016/j.beth.2011.08.001. In press. [DOI] [PubMed] [Google Scholar]
- Dillon DG, Ritchey M, Johnson BD, LaBar KS. Dissociable effects of conscious emotion regulation strategies on explicit and implicit memory. Emotion. 2007;7:354–365. doi: 10.1037/1528-3542.7.2.354. [DOI] [PubMed] [Google Scholar]
- Dolcos F, LaBar KS, Cabeza R. Remembering one year later: Role of the amygdala and the medial temporal lobe memory system in retrieving emotional memories. Proceedings of the National Academy of Sciences of the United States of America. 2005;102:2626–2631. doi: 10.1073/pnas.0409848102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Erk S, von Kalckreuth A, Walter H. Neural long-term effects of emotion regulation on episodic memory processes. Neuropsychologia. 2010;48:989–996. doi: 10.1016/j.neuropsychologia.2009.11.022. [DOI] [PubMed] [Google Scholar]
- Fabiansson EC, Denson TF, Moulds ML, Grisham JR, Schira MM. Don’t look back in anger: Neural correlates of reappraisal, analytical rumination, and angry rumination during recall of an anger-inducing autobiographical memory. NeuroImage. 2012;59:2974–2981. doi: 10.1016/j.neuroimage.2011.09.078. [DOI] [PubMed] [Google Scholar]
- Fisher RA. Statistical methods for research workers. London: Oliver & Boyd; 1950. [Google Scholar]
- Goldin PR, McRae K, Ramel W, Gross JJ. The neural bases of emotion regulation: Reappraisal and suppression of negative emotion. Biological Psychiatry. 2008;63:577–586. doi: 10.1016/j.biopsych.2007.05.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gross JJ. The emerging field of emotion regulation: An integrative review. Review of General Psychology. 1998;2:271–299. [Google Scholar]
- Gross JJ, Thompson RA. Emotion regulation: Conceptual foundations. In: Gross JJ, editor. Handbook of emotion regulation. New York: Guilford Press; 2007. pp. 3–24. [Google Scholar]
- Hardt O, Einarsson EO, Nader K. A bridge over troubled water: Reconsolidation as a link between cognitive and neuroscientific memory research traditions. Annual Reviews in Psychology. 2010;61:141–167. doi: 10.1146/annurev.psych.093008.100455. [DOI] [PubMed] [Google Scholar]
- Hayes JP, Morey RA, Petty C, Seth S, Smoski M, McCarthy G, LaBar KS. Staying cool when things get hot: emotion regulation modulates neural mechanisms of memory encoding. Frontiers in Neuroscience. 2010;4:1–10. doi: 10.3389/fnhum.2010.00230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herwig U, Baumgartner T, Kaffenberger T, Bruhl A, Kottlow M, et al. Modulation of anticipatory emotion and perception processing by cognitive control. NeuroImage. 2007;37:652–662. doi: 10.1016/j.neuroimage.2007.05.023. [DOI] [PubMed] [Google Scholar]
- Holland AC, Kensinger EA. The neural correlates of cognitive reappraisal during emotional autobiographical memory recall. Journal of Cognitive Neuroscience. 2013;25:87–108. doi: 10.1162/jocn_a_00289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kalisch R. The functional neuroanatomy of reappraisal: time matters. Neuroscience and Biobehavavioral Reviews. 2009;33:1215–1226. doi: 10.1016/j.neubiorev.2009.06.003. [DOI] [PubMed] [Google Scholar]
- Kensinger EA, Garoff-Eaton RJ, Schacter DL. Memory for specific visual details can be enhanced by negative arousing content. Journal of Memory and Language. 2006;54:99–112. [Google Scholar]
- Kross E, Davidson M, Weber J, Ochsner K. Coping with emotions past: The neural bases of regulating affect associated with negative autobiographical memories. Biological Psychiatry. 2009;65:361–366. doi: 10.1016/j.biopsych.2008.10.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LaBar KS, Cabeza R. Cognitive neuroscience of emotional memory. Nature Reviews Neuroscience. 2006;7:54–64. doi: 10.1038/nrn1825. [DOI] [PubMed] [Google Scholar]
- Lang PJ, Bradley MM, Cuthbert BN. International affective picture system (IAPS): Technical manual and affective ratings. Gainesville, FL: The Center for Research in Psychophysiology; 1999. [Google Scholar]
- MacNamara A, Ochsner KN, Hajcak G. Previously reappraised: The lasting effects of description type on picture-elicited electrocortical activity. Social Cognitive and Affective Neuroscience. 2011;6:348–358. doi: 10.1093/scan/nsq053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McKenzie S, Eichenbaum H. Consolidation and reconsolidation: Two lives of memories? Neuron. 2011;71:224–233. doi: 10.1016/j.neuron.2011.06.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McRae K, Hughes B, Chopra S, Gabrieli JD, Gross JJ, Ochsner KN. The neural bases of distraction and reappraisal. Journal of Cognitive Neuroscience. 2009;22:248–262. doi: 10.1162/jocn.2009.21243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mickley KR, Kensinger EA. Neural processes supporting subsequent recollection and familiarity of emotional items. Cognitive, Affective, and Behavioral Neuroscience. 2008;8:143–152. doi: 10.3758/cabn.8.2.143. [DOI] [PubMed] [Google Scholar]
- Mickley KR, Kensinger EA. Phenomenological characteristics of emotional memories in younger and older adults. Memory. 2009;17:528–543. doi: 10.1080/09658210902939363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ochsner KN, Bunge SA, Gross JJ, Gabrieli JD. Rethinking feelings: an FMRI study of the cognitive regulation of emotion. Journal of Cognitive Neuroscience. 2002;14:1215–1229. doi: 10.1162/089892902760807212. [DOI] [PubMed] [Google Scholar]
- Ochsner KN, Gross JJ. The cognitive control of emotion. Trends in Cognitive Sciences. 2005;9:242–249. doi: 10.1016/j.tics.2005.03.010. [DOI] [PubMed] [Google Scholar]
- Ochsner K, Gross JJ. Cognitive emotion regulation: Insights from social cognitive and affective neuroscience. Current Directions in Psychological Science. 2008;17:153–158. doi: 10.1111/j.1467-8721.2008.00566.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ochsner KN, Ray RD, Cooper JC, Robertson ER, Chopra S, Gabrieli JDE, Gross JJ. For better or for worse: Neural systems supporting the cognitive down- and up-regulation of negative emotion. NeuroImage. 2004;23:483–499. doi: 10.1016/j.neuroimage.2004.06.030. [DOI] [PubMed] [Google Scholar]
- Phan KL, Fitzgerald DA, Nathan PJ, Moore GJ, Uhde TW, Tancer ME. Neural substrates for voluntary suppression of negative affect: a functional magnetic resonance imaging study. Biological Psychiatry. 2005;57:210–219. doi: 10.1016/j.biopsych.2004.10.030. [DOI] [PubMed] [Google Scholar]
- Phan KL, Wager TD, Taylor SF, Liberzon I. Functional neuroimaging studies of human emotions. CNS Spectrums. 2004;9:258–266. doi: 10.1017/s1092852900009196. [DOI] [PubMed] [Google Scholar]
- Richards JM, Butler EA, Gross JJ. Emotion regulationin romantic relationships: The cognitive consequences of concealing feelings. Journal of Social and Personal Relationships. 2003;20:599–620. [Google Scholar]
- Richards JM, Gross JJ. Emotion regulation and memory: The cognitive costs of keeping one’s cool. Personality Processes and Individual Differences. 2000;79:410–424. doi: 10.1037//0022-3514.79.3.410. [DOI] [PubMed] [Google Scholar]
- Sergerie K, Chochol C, Armony JL. The role of the amygdala in emotional processing: A quantitative meta-analysis of functional neuroimaging studies. Neuroscience and Biobehavioral Reviews. 2008;32:811–830. doi: 10.1016/j.neubiorev.2007.12.002. [DOI] [PubMed] [Google Scholar]
- Sheppes G, Gross JJ. Is timing everything? Temporal considerations in emotion regulation. Personality and Social Psychology Review. 2011;15:319–331. doi: 10.1177/1088868310395778. [DOI] [PubMed] [Google Scholar]
- Sheppes G, Meiran N. Better late than never? On the dynamics of online regulation of sadness using distraction and cognitive reappraisal. Personality and Social Psychology Bulletin. 2007;33:1518–1532. doi: 10.1177/0146167207305537. [DOI] [PubMed] [Google Scholar]
- Shurick AA, Hamilton JR, Harris LT, Roy AK, Gross JJ, Phelps EA. Durable effects of cognitive restructuring on conditioned fear. Emotion. 2012;12:1393–1397. doi: 10.1037/a0029143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Slotnick SD, Moo LR, Segal JB, Hart J. Distinct prefrontal cortex activity associated with item memory and source memory for visual shapes. Cognitive Brain Research. 2003;17:75–82. doi: 10.1016/s0926-6410(03)00082-x. [DOI] [PubMed] [Google Scholar]
- Steinberger A, Payne JD, Kensinger EA. The effect of cognitive reappraisal on the emotional memory trade-off. Cognition and Emotion. 2011;25:1237–1245. doi: 10.1080/02699931.2010.538373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steinvorth S, Corkin S, Halgren E. Ecphory of autobiographical memories: an fMRI study of recent and remote memory retrieval. NeuroImage. 2006;30:285–298. doi: 10.1016/j.neuroimage.2005.09.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wheeler ME, Petersen SE, Buckner RL. Memory’s echo: Vivid remembering reactivates sensory-specific cortex. Proceedings of the National Academy of Sciences of the United States of America. 2000;97:11125–11129. doi: 10.1073/pnas.97.20.11125. [DOI] [PMC free article] [PubMed] [Google Scholar]