

Abstract
Only a sub-group of human drug users progress from initial drug-taking to drug addiction. An important aspect of this progression is the influence of the learned association between the effects of the drug and the environment in which it is experienced on continued drug-taking and-seeking. These associations can be modeled using the conditioned place preference (CPP) paradigm, although no current method of CPP analysis allows for identification of within-group variability among subjects. In the present study, we adapted a “criterion” method of analysis to separate “CPP expressing” from “non-CPP expressing” rats, in order to study more directly within-group variability in the CPP paradigm. Male Sprague-Dawley rats were conditioned with cocaine (5, 10, 20 mg/kg) or saline in an unbiased three-chamber CPP apparatus in either a single- or four-trial CPP procedure. A classification and regression tree analysis of time spent in the cocaine-paired chamber established a time of 324 s spent in the cocaine-paired chamber as the criterion for cocaine CPP expression. This criterion effectively discriminated control from cocaine-conditioned rats and was reliable for rats trained in both a single- and four-trial CPP procedure. The criterion method showed an enhanced ability to detect effective doses of cocaine in the single trial procedure and a blockade of CPP expression by MK 212 (0.125 mg/kg) treatment in a sub-group of rats. These data support the utility of the criterion analysis as an adjunct to traditional methods that compare group averages in CPP.
Keywords: classical conditioning, statistical analysis, classification and regression tree analysis, serotonin 2C receptor, animal model, rat
Introduction
Variability in the response to cocaine plays a major role in the development of cocaine abuse and addiction. One estimate predicts that five to six percent of cocaine users will become addicted within the first two years of cocaine use (O'Brien & Anthony 2005). The biological substrates that allow a sub-group of cocaine users to remain in control of their drug use while a different sub-group becomes addicted are largely unknown. Achievement of sustained abstinence in the sub-group of users who do become addicted to cocaine is challenging, as stimuli commonly associated with cocaine use (i.e., drug paraphernalia) elicit craving and relapse to drug-taking (Childress et al. 1999), a process likely supported by the development of learned, classically-conditioned associations between the effects of cocaine (unconditioned response) and the environment in which cocaine is experienced (conditioned stimulus; Childress et al. 1992). These observations suggest a critical need for a method to analyze differences between population sub-groups in the response to cocaine-associated cue environments.
The conditioned place preference (CPP) paradigm is a very attractive animal model of these conditioned cocaine-environment associations. In the acquisition phase of this paradigm, cocaine administration is temporally paired with placement in a distinct environment, and treatment with saline is paired with an alternate environment. Although the exact properties of cocaine that become associated with the environment during the conditioning sessions are unknown, both the rewarding and discriminative stimulus properties of cocaine are thought to comprise (in part) the unconditioned stimulus and thus are important for the formation of the CPP (Carr et al. 1989). Following conditioning sessions, subjects are allowed to freely roam both environments and the time spent in each environment is assessed. During this expression test, increased time spent in the environment formerly paired with cocaine is defined as a CPP and suggests that an association between cocaine and the environment has been learned (Bevins & Cunningham 2006).
Determination of CPP expression typically involves computation of the average performance on a specific metric (i.e., time in the cocaine-paired environment; Cunningham et al. 2003). Previous reports suggest, however, that calculating a group average may obscure differences between population sub-groups in the acquisition or expression of CPP. For example, Adams and colleagues (Adams et al. 2001) observed variability in the ability of the dopamine D1 receptor antagonist SCH 23390 (0.03 mg/kg) to block the expression of cocaine CPP; specifically, rats treated with SCH 23390 before the test session “could be divided into three separate and equal-sized groups” based upon differences in the amount of time spent in the chamber during the test session that had been paired with cocaine compared to that paired with saline. Population sub-groups have also been observed in CPP when 3,4-methylenedioxymethamphetamine (MDMA) was used as the conditioning drug; in this research, some animals showed a strong CPP, while others showed an aversion to the MDMA-paired chamber (Daza-Losada et al. 2007). These reports suggest the need for an additional method for the analysis of CPP data that allows for separation of subjects within a treatment group. A method that allows for the dichotomous categorization of individual subjects within a treatment group as “CPP expressing” or “non-CPP expressing” would provide a complementary approach to consider differences among subjects within a treatment population.
We thus sought to develop a criterion for CPP analysis that would allow individual subjects to be categorized into two sub-groups: “CPP expressing” or “non-CPP expressing.” To establish this criterion, data from rats conditioned with saline in both environments (control) were compared with data from rats conditioned with cocaine to identify a numerical cut-off, based on the number of seconds spent in the cocaine-paired chamber that appropriately separated control from cocaine groups. The criterion was confirmed by comparing the dose-effect relationship for cocaine to elicit a CPP generated by the criterion analysis to that generated by a traditional analysis of time spent in the cocaine-paired chamber (Martin-Iverson & Reimer 1996; Belzung & Barreau 2000). Following establishment, this criterion method was used to analyze the effects of several doses of MK 212, a preferential agonist at the serotonin 2C receptor (5-HT2CR), on the expression of single-trial CPP. MK 212 was investigated given previous evidence suggesting involvement of the 5-HT2CR in the behavioral response to cocaine-associated cues in rats (Liu & Cunningham 2006). Employing the criterion analysis, we demonstrate an enhanced ability to detect a cocaine CPP in the single-trial procedure, as well as pharmacological blockade of CPP in a sub-group of rats, suggesting this new method of analysis for CPP data may be useful as an adjunct to traditional methods for the study of differences between sub-groups in the CPP paradigm.
Methods
Subjects
Male Sprague-Dawley rats (Harlan Sprague-Dawley, Inc., Indianapolis, IN) weighed 225–275 g at the beginning of the study. The rats were housed four per cage in standard, clear plastic rodent cages with food and water freely available, in a temperature (21–23°C) and humidity (55–65%) controlled environment under a 12-h light/dark cycle (lights on 0700 h). Animals were acclimated to the colony for at least one week and were handled prior to the start of experimental sessions. All experiments were conducted during the light phase of the light-dark cycle (0800–1800 h) and were carried out in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals and with approval from the Institutional Animal Care and Use Committee.
Conditioned place preference apparatus
The CPP apparatuses were housed within sound- and light-attenuating cabinets and contained three sensory environments distinguished by wall colors and floor textures (n=8, ENV-013, Med Associates, Georgia, VT USA). The two side conditioning chambers (interior dimensions: 25.5 cm L x 21.0 cm W x 20.9 cm H) were separated by a smaller chamber (13.2 cm L X 21.0 cm W x 20.9 cm H). One conditioning chamber had white walls and a stainless steel mesh (1.3 × 1.3 cm) floor and the other conditioning chamber had black walls and a floor of stainless steel rods (4.8 mm placed on 1.6 cm centers). The center chamber had gray walls and a floor of sheet metal. Guillotine doors separated each conditioning compartment from the center compartment. The intensity of ambient illumination was adjusted to 7 lux in each conditioning chamber and 30 lux in the center chamber, so as to balance side preference for each conditioning chamber (Roma & Riley 2005). Automated data collection was accomplished through photobeam detectors. There were 15 infrared photobeam detectors that were 4.5 cm above the chamber floor. Of these, six photobeams were arrayed along the length of each conditioning compartment 1.25 cm from the end wall with 5 cm between beams. Three photobeams were arrayed along the length of the central compartment spaced 4.75 cm apart. The photobeams were connected to a computer interface, and MED-PC software (Med Associates) recorded the time spent in each chamber based on the recorded activity counts (any beam break within the current chamber) between disruption of (1) the entrance beam of that chamber (beam break beyond the first beam of a chamber) and (2) the entrance beam of a different chamber.
Acquisition and expression of cocaine conditioned place preference
Each experiment used separate groups of naïve animals. The CPP training consisted of three phases (preconditioning, conditioning, and test) which occurred over consecutive days. In all phases, the home cages of the animals were transferred from the animal colony to the test room 5 min before the animals were placed in the CPP apparatus. To determine baseline preference (preconditioning), rats were placed in the center chamber with the guillotine doors raised and allowed to freely roam the apparatus for 30 min. The time spent in each conditioning chamber was recorded, and the conditioning chamber in which an animal spent the least amount of time was designated the initially least-preferred chamber for that animal. Conditioning was performed using a biased protocol in which cocaine was paired with the initially least-preferred chamber (Spyraki et al. 1982; Blander et al. 1984; Isaac et al. 1989).
Cocaine CPP conditioning sessions consisted of alternating sessions of cocaine and saline injections with the guillotine doors in place. Two sessions were conducted each day separated by at least six hours (Shippenberg & Heidbreder 1995), thus rats experienced both a morning and an afternoon conditioning session. For cocaine CPP training, one daily conditioning session began with an injection of cocaine and immediate confinement to the initially least-preferred chamber for 45 min. The other daily conditioning session began with a saline (1 mL/kg, IP) injection and immediate confinement to the alternate conditioning chamber for 45 min. Thus, some rats received cocaine during the morning session, and others received cocaine during the afternoon session. Control rats were injected with saline prior to placement into each conditioning chamber for both sessions on each day. At the termination of the session, rats were returned to their home cages. Rats experienced one conditioning session with cocaine and one with saline for studies in the single-trial conditioning procedure. In the four-trial conditioning procedure, rats experienced eight alternating sessions of cocaine and saline injections over four consecutive days.
The expression test for CPP was conducted 16–24 h after the final conditioning session. The time of the test session (morning vs. afternoon) was counterbalanced across rats relative to the time of cocaine conditioning. Rats were placed in the center chamber with the guillotine doors raised and allowed to freely roam the apparatus for 15 min while the time spent in each chamber was recorded. Data are presented as mean time (sec ± S.E.M.) spent in the initially least-preferred chamber.
Establishment of criterion for CPP expression
To develop a criterion that would allow the identification of a sub-group of rats that express a CPP versus a sub-group that does not express a cocaine CPP, we rationalized that control rats (conditioned with saline) should not express a CPP, while animals conditioned with cocaine should express a CPP. We thus set out the goal of identifying a numerical cut-off (i.e., number of seconds spent during the test session in the initially least-preferred chamber) that would separate the control from cocaine groups. To establish this value, we compiled CPP data from the expression test in animals (n=279) conditioned with saline (control) or cocaine (one pairing with 20 mg/kg of cocaine or four pairings with 10 mg/kg of cocaine, without any other drug treatments); these data were collected in the course of several studies run in our laboratory over a 3-year period (dela Cruz and Cunningham, in preparation). The amount of time spent by each rat in the initially least-preferred chamber during the expression test session was organized into 25 sec time bins (i.e., 200–224 sec, 225–249 sec, etc). The resulting curves for control and cocaine groups were compared, and exact P values for the chi square test for independence were estimated using Cytel Studio Software (version 6.3, Cambridge, MA; Kupperman 1960) using the Monte Carlo option. The mean and median for each group (control and cocaine) were calculated and a t-test was used to compare the group means.
To establish the numerical cut-off that could serve as a criterion for CPP expression, we subjected the compiled CPP data to classification and regression tree analysis (CART; Salford Systems, San Diego, CA). The CART analysis is a form of binary partitioning (Lewis 2000) used to identify population subgroups as well as the factor (i.e., number of seconds spent during the test session in the initially least-preferred chamber) upon which the groups differ (Lemon et al. 2003). The data from all subjects (for our analysis, the amount of time each rat spent during the test session in the initially least-preferred chamber) were input into a “parent node” which was then split into two “child nodes” based on a “splitting” criterion. This splitting criterion seeks to minimize the average impurity in each child node by achieving the largest difference between the impurity of the parent node and the weighted average impurity of the two child nodes (Lemon et al. 2003). The minimum value for impurity is zero, which implies no variability in the dependent variable; the maximum value for impurity is 0.5 when subjects from the underlying subpopulations are equally distributed in the child nodes (Zhang et al. 1996; Lemon et al. 2003). Thus, the splitting criterion established by the CART analysis was deemed the criterion used to categorically identify rats that express or do not express a cocaine CPP.
Assessment of cocaine CPP criterion reliability
To assess the ability of the criterion to identify rats that express cocaine CPP, we first analyzed the expression of CPP following conditioning with different doses of cocaine (0, 5, 10, or 20 mg/kg) in separate groups of rats. Preconditioning was conducted as described, and rats were then randomly assigned to one of eight groups: single trial control (0 mg/kg), single trial 5 mg/kg, single trial 10 mg/kg, single trial 20 mg/kg, four trial control, four trial 5 mg/kg, four trial 10 mg/kg, and four trial 20 mg/kg. Conditioning was conducted as described. Testing was conducted 16–24 h after the final conditioning session, as described. The percentage of subjects that met the criterion for positive expression of CPP (and thus categorized as “CPP expressing”) was compared in the control and cocaine groups using χ2 analysis, and simple comparisons between the control and each cocaine group were conducted using Fisher’s exact test (Sheskin 2004). For purposes of comparison, one-way ANOVA followed by Student-Newman-Keuls’ test was used to compare the amount of time spent during the test session in the initially least-preferred chamber between each conditioning group.
Application of cocaine CPP criterion
Following assessment of the reliability of the criterion, the cutoff was applied to analyze a data set in which rats showed variability within a given conditioning group in the amount of time spent in the initially least-preferred chamber during the CPP expression test. The effects of the preferential 5-HT2CR agonist MK 212 (0, 0.125, 0.25 mg/kg) on the expression of a single-trial cocaine (20 mg/kg) CPP were determined. Preconditioning was conducted as described above and rats were randomly divided into six groups. The next day, two 45 min conditioning sessions took place 6 hours apart. In one session, cocaine rats received a cocaine (20 mg/kg) injection immediately prior to confinement to the chamber determined to be least-preferred during preconditioning; in the other conditioning session, cocaine animals received saline (1 ml/kg) immediately prior to confinement to the opposite conditioning chamber. Control animals received saline immediately prior to both conditioning sessions. The expression test for CPP was conducted 16–24 h following the final conditioning session. Rats (n=21–23/group) received an injection of MK 212 or saline 10 min before the expression test session.
Two analyses were performed to assess the ability of MK 212 to block the expression of cocaine CPP. The binary outcome of “CPP expression” (yes ≥ 324 s versus no < 324 s) was used in a logistic regression with predictor variables conditioning (saline versus cocaine) and MK 212 treatment (0, 0.125, 0.25 mg/kg), and on the conditioning x treatment interaction. A priori individual comparisons were performed using categorical modeling via the CATMOD procedure in SAS. For comparison, a traditional analysis using the continuous variable mean time spent in the initially least-preferred chamber during the test session was analyzed using a two-way ANOVA with factors conditioning (saline versus cocaine) and MK 212 treatment (0, 0.125, 0.25 mg/kg) and the conditioning x treatment interaction. A priori individual contrasts were conducted for pairwise comparisons. For all comparisons, α was set at 0.05.
Drugs
Cocaine HCl salt (National Institute on Drug Abuse, Research Triangle, NC) and MK 212 [6-chloro-2-(1-piperazinyl)pyrazine hydrocholoride; Tocris, Ellisville, MO] were dissolved in 0.9% NaCl. All injections were given IP in a volume of 1 ml/kg. Doses of all drugs refer to the weight of the salt. Doses of MK 212 utilized in this study were based on previous work from our laboratory (Liu & Cunningham 2006).
Results
Establishment of criterion for CPP expression
Data from preconditioning sessions were compiled for all rats (n=342) in this study. Out of the 1800 sec session, rats spent a mean (± SEM) of 700.06 sec (± 8.68) on the black side (39.0% of total time) and 680.01 sec (± 8.95) on the white side (38.0% of total time); the remaining time (420.00 ± 7.17 sec) was logged in the center grey chamber. The white chamber was the initially least preferred chamber for 51% of rats, and the black chamber was initially least preferred for 49% of rats. A χ2 analysis indicated that these percentages did not differ significantly from those due to chance: thus the CPP apparatus with the conditions employed is considered unbiased.
The time spent during the test session in the initially least-preferred chamber for rats in either the control (n=121) or cocaine (n=158) groups was divided into 25 sec bins and plotted. We observed a clear distinction between the amount of time spent in the initially least-preferred chamber during the test session in cocaine versus control groups, as expected, and a χ2 analysis revealed a significant difference between the distribution of control and cocaine group members across time bins (p<0.001, Fig. 1). For the control group, the bin that contained the most rats (n=23) was 275–299 sec, with an average time (±SEM) spent in the initially least-preferred chamber equal to 275 ± 8 sec (Fig. 1A). Rats in the control group spent a median of 276 sec (interquartile range 225–319 sec, Fig. 1B) in the initially least-preferred chamber during the test session. The control group spent 338 ± 8 sec in the initially more-preferred chamber during the test session. To further demonstrate the unbiased nature of the apparatus, the amount of time that control (n=121) animals spent in each chamber during the 15 min test session was analyzed. On test day, control animals spent an average (±SEM) of 306.84 sec (±7.17) in the black chamber (34.1% of total time) and 305.89 sec (±7.889) in the white chamber (34.0% of total time); the remaining time (287.43 ± 7.45 sec, 31.9% of total time) was logged in the center grey chamber. Thus, non-drug conditioned animals do not develop a natural preference for one conditioning chamber, and the apparatus conditions employed are considered unbiased.
Figure 1. Distribution of time spent in the initially least-preferred chamber demonstrates a significant difference between saline and cocaine conditioning.
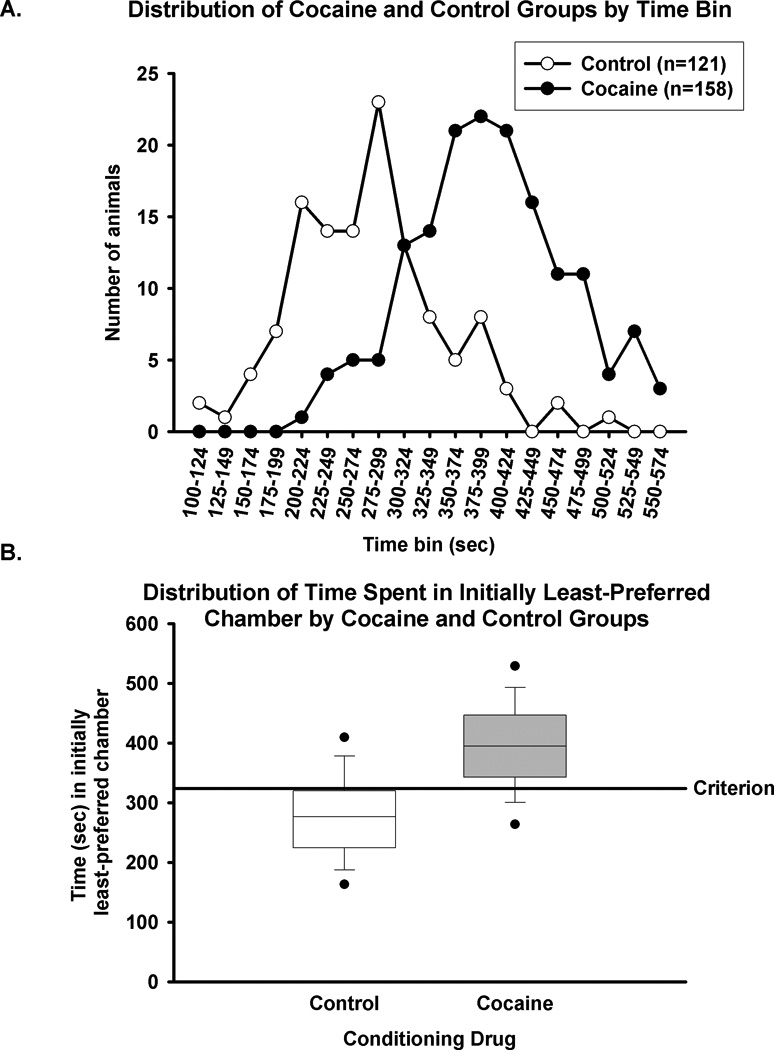
Fig. 1A, Rats in control (n=121, open circles) and cocaine (n=158, black circles) groups were tested for the expression of CPP in a 15 min session. The amount of time spent by each rat during the test session in the initially least-preferred chamber was categorized according to 25 sec bins. χ2 analysis revealed a significant difference between the two distributions (p<0.001). Fig. 1B, Rats in control (open box) and cocaine groups (grey box) were tested for the expression of CPP in a 15 min session. The line in the center of the box represents the median time, and the top and bottom of the box represent the 75th and 25th percentiles, respectively. The black circles represent the 95th and 5th percentiles. The solid line (“criterion”) represents the 324 s criterion for the expression of cocaine CPP.
For the cocaine group, the bin that contained the most rats (n=22) was 375–399 sec, with an average time (±SEM) spent in the initially least-preferred chamber equal to 394 ± 6 sec (Fig. 1A). In comparison, the cocaine group spent an average time (±SEM) of 273 ± 5 sec in the initially more-preferred (saline paired) chamber, significantly less time than was spent in the initially least-preferred chamber (p<0.001). This group spent 234 ± 5 sec in the center grey chamber. Rats in the cocaine group spent a median of 394 sec (interquartile range 343–446 sec, Fig. 1B) in the initially least-preferred chamber during the test session. A significant difference between the average amount of time spent in the initially least-preferred chamber during the CPP expression test was observed between the control and cocaine groups (Fig. 1A, T=13.05, p<0.001).
To develop a criterion for the expression of CPP, an assessment of the amount of time spent in the initially least-preferred chamber during the expression test by rats in the cocaine group compared to control group was performed using CART analysis (see Methods). This analysis revealed that a criterion of 324 s yielded the greatest separation between cocaine and control groups. Thus, any rat that spent at least 324 s in the initially least-preferred chamber during the test session was categorized as a “CPP expressing” subject.
Binary classification based upon a specific criterion provides a means to discriminate subjects within a population as either “CPP expressing” or “non-CPP expressing.” In a perfect separation, all control rats would be classified as “non-CPP expressing” and all cocaine rats would be classified as “CPP expressing.” However, a perfect separation based upon a binary classification is rarely seen, and, indeed, the distributions of data collected in control and cocaine groups during the test session overlap (Fig. 1A). Binary classification schemes are commonly used in the clinical literature to assess the likelihood of disease presence given a positive test result, and this field has developed language to describe the four possible outcomes from this type of categorization: true positive, true negative, false positive, or false negative. “Positive” describes a positive test result, while “negative” describes a negative test result. “True” describes a test result consistent with disease state (“true positive” is a positive test result when disease is present while a “true negative” is a negative test result when disease is absent) and “false” describes a test result inconsistent with disease state (“false positive” is a positive test result in the absence of disease while “false negative” is a negative test result in the presence of disease). We have utilized this language to assess how well a classification provided by the criterion (“expressing” or “not expressing”) matched the actual conditioning protocol (cocaine vs control). Based on the CPP criterion (324 s), rats defined as not expressing (<324 s) are classified as “negative,” and those defined as expressing (≥ 324 s) are classified as “positive.” Rats in the cocaine group correctly classified as “CPP expressing” are classified as “true positives,” and rats in the cocaine group defined as “non-CPP expressing” are “false negatives.” Rats in the control group correctly classified as “non-CPP expressing” are “true negatives,” while those defined as “CPP expressing” are considered “false positives.” With these designations in place, we then determined the true positive and true negative probabilities of the 324 s criterion. The true positive probability (Sheskin 2004)--equal to the number of true positives divided by the total number of rats in the cocaine group--is a measure of the ability of the criterion to identify animals that express a CPP. A criterion of 324 s gives a true positive probability equal to 0.83. The true negative probability, which is equal to the number of true negatives divided by the total number of rats in the control group, is a measure of the ability of the criterion to exclude animals that do not express a CPP (Sheskin 2004). The 324 s criterion gives a true negative probability equal to 0.78. True positive and true negative probabilities are inversely related; thus, choosing a criterion requires balancing these probabilities. The 324 s criterion classifies 83% of animals conditioned with cocaine as expressing a CPP and classifies 78% of animals in the control group as not expressing a CPP. As this criterion gives appropriately high values for both the true positive and true negative probabilities, this analysis supports the decision to use a criterion of 324 s.
Examination of the time spent during the test session in each chamber by the subjects classified as “CPP expressing” compared to those classified as “non-CPP expressing” provides additional verification of the criterion. During the test session, the “CPP expressing” subjects (n=131) spent an average (±SEM) of 417 ± 5 sec in the initially least-preferred chamber and 259 ± 5 in the initially more-preferred chamber, clearly demonstrating a cocaine CPP. For comparison, the “non-CPP expressing” subjects (n=27) spent an average (±SEM) of 283 ± 6 sec in the initially least-preferred chamber and average (±SEM) of 337 ± 14 sec in the initially more-preferred chamber; thus, these animals do not express a CPP. Based on this comparison, the criterion can be said to appropriately separate animals that express a CPP from those that do not.
Two groups of subjects trained on a cocaine CPP were utilized to generate the criterion: one group of rats (n=71) experienced a single pairing of the conditioning environment and cocaine (20 mg/kg), while a second, separate group of rats experienced four pairings of the conditioning environment and cocaine (10 mg/kg). Our next analysis tested the hypothesis that the criterion of 324 s for the expression of CPP was applicable to both cocaine groups by separately analyzing the data from rats that experienced one cocaine conditioning session with 20 mg/kg from those that experienced four conditioning sessions with 10 mg/kg of cocaine (Fig. 2). The time each animal spent in the initially least-preferred chamber during the test session was grouped into 25 sec bins and plotted (Fig. 2A). The distributions of time spent in the initially least-preferred chamber during the test session for the rats that experienced one (20 mg/kg) or four (10 mg/kg) cocaine conditioning sessions overlapped neatly. A χ2 analysis revealed no difference between the frequency distributions of single- and four-trial group members across time bins. The average time (±SEM) in the initially least-preferred chamber spent during the test session for the single (397 ± 9 sec) and four trial (392 ± 8 sec) groups did not differ (Fig. 2A, T=154, p = 0.51). The 324 s criterion classified 82% of rats conditioned with the single-trial procedure and 84% of rats conditioned with the four-trial procedure as expressing a CPP. Rats in the single-trial conditioning group spent a median of 397 sec (interquartile range 343–448 sec, Fig. 2B) in the initially least-preferred chamber during the test session. Rats in the four-trial conditioning group spent a median of 392 sec (interquartile range 341–435 sec, Fig. 2B) in the initially least preferred chamber during the test session. These overlapping distributions emphasize a similarity in CPP expression between the group of rats conditioned with a single pairing of 20 mg/kg of cocaine and environment and those rats conditioned with four pairings of 10 mg/kg of cocaine and environment. We thus conclude that the criterion of 324 s for the expression of CPP is reliable regardless of the two protocols chosen for conditioning.
Figure 2. The distribution of time spent in the initially least-preferred chamber does not depend on number of conditioning sessions.
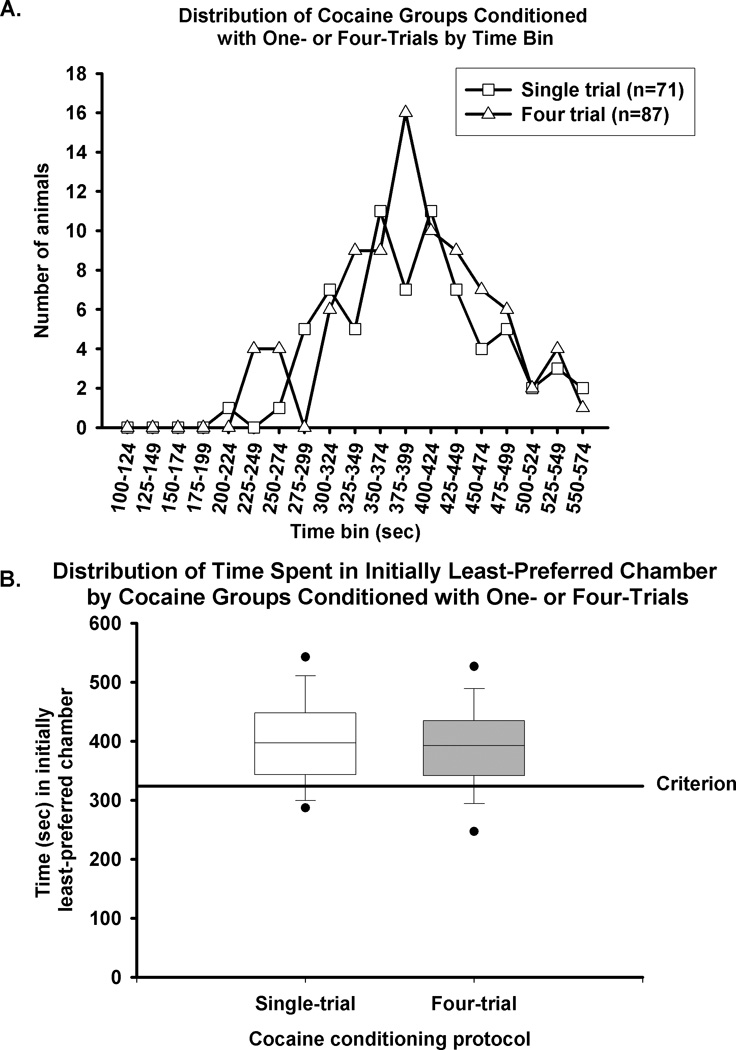
Fig. 2A, Following conditioning with either a single pairing with the environment of 20 mg/kg of cocaine (open square, n=71) or four pairings of 10 mg/kg of cocaine (open triangle, n=87), animals were tested for the expression of CPP in a 15 min session. The amount of time spent by each rat in the initially least-preferred chamber during the test session was categorized according to 25 sec bins. No differences in the percentage of animals meeting the CPP expression criterion were observed between the two conditioning procedures. Fig. 2B, Rats in single- (open box) and four-trial groups (grey box) were tested for the expression of CPP in a 15 min session. The line in the center of the box represents the median, and the top and bottom of the box represent the 75th and 25th percentiles, respectively. The filled circles represent the 95th and 5th percentiles. The solid line represents the 324 s criterion for the expression of cocaine CPP.
Assessment of cocaine CPP criterion reliability
To assess the reliability of the CPP criterion, we compared the dose-effect function for cocaine CPP acquisition generated by the criterion analysis (which compares the percentage of group members that meet or exceed the CPP criterion) to that generated by a traditional analysis (which compares the average group time spent during the test session in the initially least-preferred chamber). The dose effect function was examined for three doses of cocaine (5, 10, 20 mg/kg) in both the single- and four-pairing paradigms. An analysis of the proportion of rats in each group that spent at least 324 s in the initially least-preferred chamber during the test session (and thus met the criterion for CPP expression) revealed a significant effect of a single cocaine conditioning trial on the percentage of rats meeting the CPP criterion (Fig. 3A, left; Fisher’s exact test, p<0.05). A priori comparisons revealed that both 10 and 20 mg/kg evoked a single-trial CPP upon expression test (Fisher’s exact test, p<0.05). Eighty-three percent of rats conditioned with either 10 or 20 mg/kg of cocaine, respectively, met the criterion compared to 33% of rats in the control group (0 dose; Fig. 3A). These data were separately examined using a more-widely employed (“traditional”) analysis that compared the mean time spent in the initially least least-preferred chamber during the test session between control and cocaine groups. In this analysis, an ANOVA revealed a significant main effect of cocaine conditioning on the mean time spent in the initially least-preferred chamber during the test session (Fig. 3A, right; F3,47=3.75, p<0.05). A priori comparisons revealed that a single conditioning session with 20 mg/kg of cocaine significantly increased the time spent in the initially least-preferred chamber compared to control (p<0.05).
Figure 3. Cocaine produces a CPP following a single or four pairings of cocaine and environment.
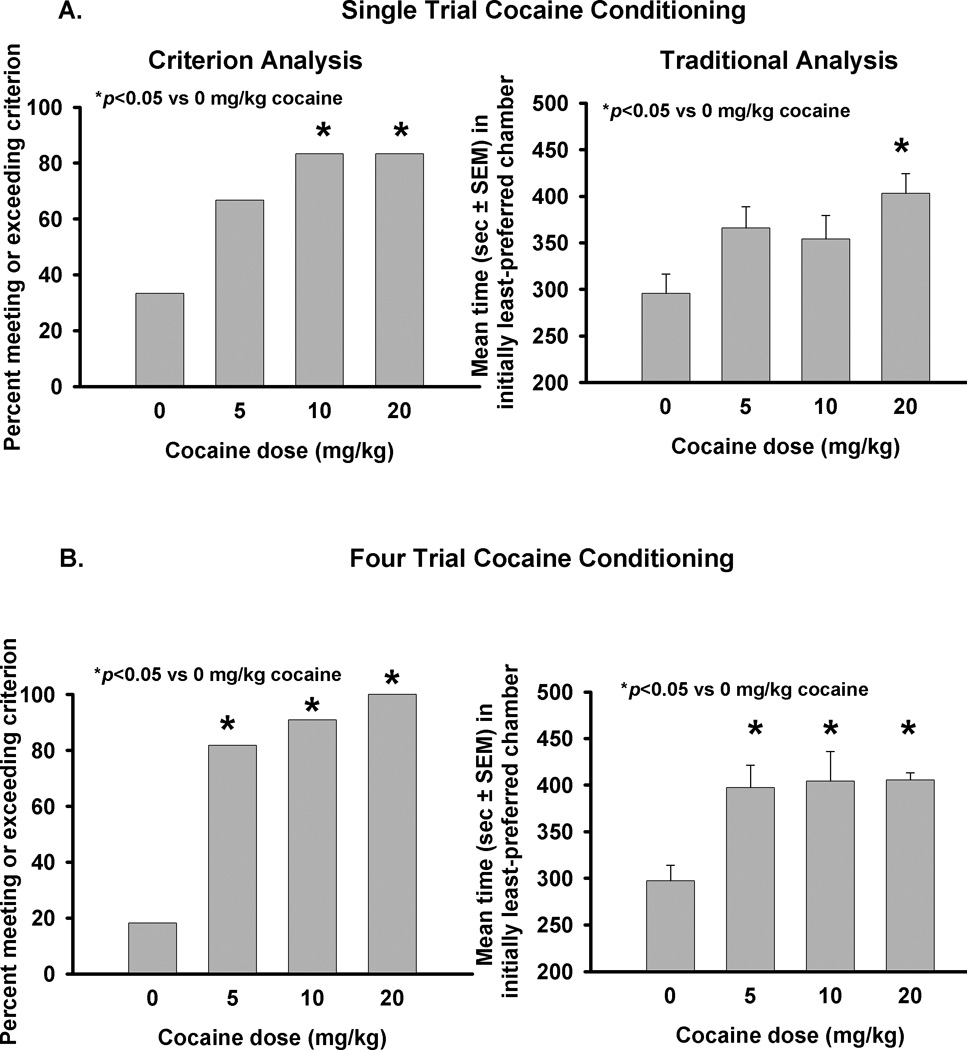
Following either a single (Fig. 3A) or four (Fig. 3B) conditioning sessions in which the environment was paired with cocaine (0, 5, 10, 20 mg/kg), the amount of time rats (n=11–12/group) spent during a 15 minute drug-free test session in the initially least-preferred chamber was observed. Fig. 3A, left, Criterion analysis comparing the proportion of animals spending at least 324 s during the test session in the initially least-preferred chamber revealed a significant effect of cocaine (10 or 20 mg/kg) conditioning. Fig. 3A, right, Traditional analysis comparing the mean time spent in the initially least-preferred chamber during the test session by each conditioning group revealed a significant effect of cocaine (20 mg/kg) conditioning. Fig. 3B, left, Criterion analysis demonstrated a significant effect of cocaine (5, 10, 20 mg/kg) conditioning following four pairings. Fig 3B, right, Traditional analysis also demonstrated a significant effect of cocaine (5, 10, 20 mg/kg) after four pairings. * p< 0.05 vs 0 mg/kg cocaine (control)
An analysis of the proportion of rats in each group that spent at least 324 s in the initially least-preferred chamber during the test session revealed a significant effect of four cocaine conditioning trials on the percentage of rats meeting the CPP criterion (Fig. 3B, left, Fisher’s exact test, p<0.001). A priori comparisons revealed that four pairings of each dose of cocaine (5, 10, or 20 mg/kg, p<0.01) with the environment resulted in the expression of a CPP (Fig. 3B, left). These data were separately examined using a traditional analysis that compared the group mean time spent in the initially least-preferred chamber during the test session. In this analysis, an ANOVA revealed a significant main effect of cocaine conditioning (Fig. 3B, right, F3,43=5.68, p<0.01) on the mean time spent in the initially least-preferred chamber during the test session. A priori comparisons revealed a significant effect of each cocaine dose tested (Fig. 3B, right, p<0.05). These results demonstrate a somewhat enhanced ability of the criterion analysis for detecting conditioning of a cocaine CPP in the single-trial procedure, but not a four-trial procedure. The 324 s criterion has also been verified on an independent dataset in which the control group consisted of 16 rats and the cocaine group consisted of 40 rats (dela Cruz and Cunningham, in preparation). The 324 s criterion classified 75% of animals conditioned with cocaine as expressing a CPP and 63% of animals in the control group as not expressing a CPP (dela Cruz and Cunningham, in preparation). Thus, we consider the criterion of 324 s to appropriately differentiate control from cocaine conditioned rats, consistent with the true positive and true negative probabilities of the original data set.
Application of cocaine CPP criterion
The ability of MK 212 treatment to alter the expression of a single-trial cocaine CPP (20 mg/kg) was assessed as an application of the 324 s criterion. Criterion analysis of the percentage of rats that met the 324 s criterion for the expression of cocaine CPP revealed a significant main effect of cocaine conditioning (Fig. 4A; Wald chi-square=10.43, p<0.001), and a significant interaction between conditioning and MK 212 treatment (Wald chi-square=9.80, p<0.01), in the absence of a main effect of MK 212 treatment (Wald chi-square=1.47, NS). An a priori comparison revealed a significant difference between the control and cocaine groups administered vehicle before the expression test (p<0.001). The cocaine group expressed a CPP, as assessed by the percentage of rats meeting or exceeding the 324 s criterion. Fewer subjects met or exceeded the criterion in the cocaine group treated with the lower dose of MK 212 (0.125 mg/kg) before the expression test (Fig. 4A, p=0.02), indicating a blockade of CPP expression by the MK 212 treatment in a distinct sub-group of the rats tested. For comparison, a two-way ANOVA comparing the mean time spent in the originally least-preferred chamber during the test session revealed a significant main effect of conditioning (Fig. 4B, F1,129=33.69, p<0.001). There was no significant main effect of MK 212 treatment (F2,129=1.43, NS), although a significant cocaine conditioning x MK 212 treatment interaction (F2,129=3.13, p<0.05) was observed. An a priori comparison conducted with linear contrasts revealed a significant difference between the control and cocaine groups administered vehicle before the expression test (p<0.01). The cocaine group expressed a CPP, as assessed by the average time spent during the test session in the initially least-preferred chamber. In contrast to the results obtained with the criterion analysis, no differences were observed between the cocaine-conditioned groups, indicating no effect of MK 212 treatment on the expression of single-trial cocaine CPP in this analysis. A composite analysis (Fig. 4C) that overlays the amount of time during the test session each rat spent in the initially least-preferred chamber with the group means for this value allows for direct comparison of the criterion and traditional analyses. This analysis highlights the differences between groups in the variability of time spent during the test session in the initially least-preferred chamber. For example, the values for individuals (open circles) in the cocaine group administered vehicle prior to the test session are tightly clustered around the average value (filled square). In comparison, the time spent during the test session in the initially least-preferred chamber by the individual subjects in the cocaine group treated with MK 212 (0.125 mg/kg) are widely spaced, demonstrating sub-groups within the population in the response to MK 212 treatment in rats conditioned with cocaine. The criterion method of analysis allows for quantification of two separate sub-groups.
Figure 4. MK 212 treatment blocks expression of a single trial cocaine (20 mg/kg) CPP in the criterion but not traditional analysis.
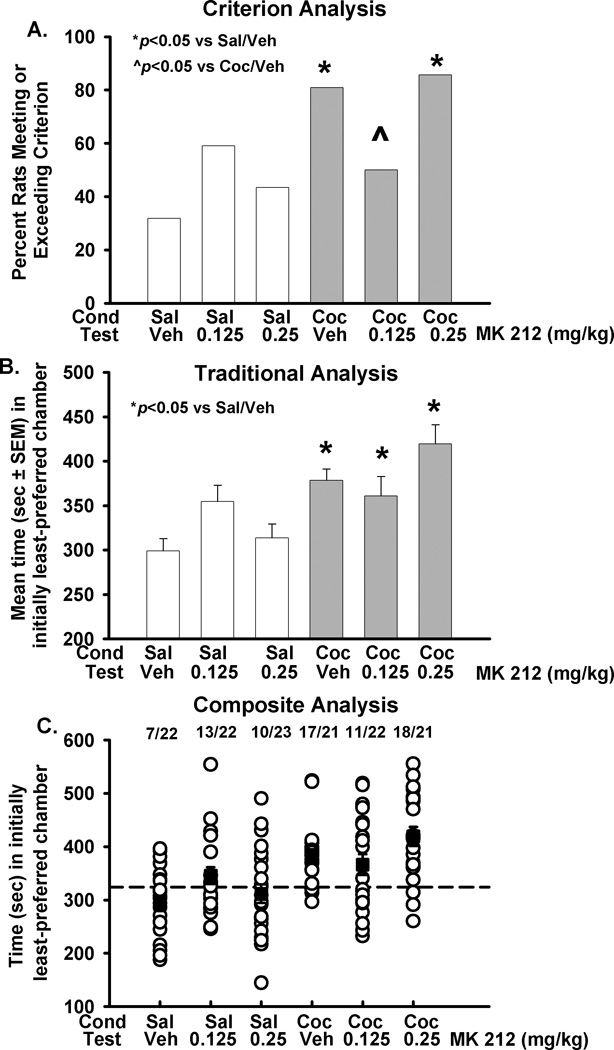
Following preconditioning, animals were conditioned with saline (open bars) or cocaine (20 mg/kg, grey bars) to the initially least-preferred side of the chamber. Ten minutes before the test session, animals were treated with MK 212 (0, 0.125, 0.25 mg/kg). Fig. 4A, Criterion analysis of the percentage of animals meeting or exceeding the 324 s criterion for CPP expression revealed a blockade of expression of a single trial cocaine CPP by MK 212 (0.125 mg/kg). Fig. 4B, Traditional analysis comparing the mean group time during the test session in the initially least-preferred chamber demonstrated that MK 212 treatment did not alter the expression of a single trial cocaine CPP. Fig. 4C, Composite analysis demonstrated overlap of criterion and traditional analyses; the amount of time spent during the test session in the initially least-preferred chamber by individual animals (open circles); proportion at top gives the number of animals meeting the criterion/number in treatment group. Dashed line (- - -) represents 324 s criterion. Black boxes represent group mean ± SEM. *p<0.05 vs vehicle-saline; ^ p<0.05 vs vehicle-cocaine; cond=conditioning drug, test=injection before test session.
Discussion
We established a criterion value (324 s) for the expression of a cocaine CPP, based on the time spent in the initially least-preferred chamber during the test session that effectively discriminated control from cocaine-conditioned rats. This criterion was reliable for two different groups of rats: those conditioned with either one pairing of 20 mg/kg of cocaine or those conditioned with four pairings of 10 mg/kg of cocaine with the CPP environment. The criterion method revealed an additional effective dose of cocaine in the single-trial procedure and a pharmacological blockade of CPP expression by MK 212 (0.125 mg/kg) in a sub-group of rats. The utility of the criterion method is the ability to identify and quantify the presence of two distinct sub-groups within a given conditioning group. In current methods for assessing CPP expression, the presence of a sub-group within a treatment group may be obscured by the need to compare averaged results to that of a control group (Adams et al. 2001; Daza-Losada et al. 2007). The proposed criterion method, by classifying the behavior of each subject as “CPP expressing” or “non-CPP expressing,” allows for the detection of such sub-groups. We propose that the criterion method is complementary to other methods of analysis of CPP data currently in use and may be a particularly useful adjunct in analyses of pharmacological manipulations in CPP paradigms as well as in analysis of molecular mechanisms that underlie CPP.
The criterion analysis creates a binary, “all-or-none” classification for each subject, in that each rat in the study is classified as either “expressing” or “not expressing” a CPP. This method of analysis thus transforms continuous time recordings of seconds spent in a chamber into a discrete, all-or-none classification. Several groups have suggested that the dose-effect function for expression of cocaine CPP is not graded in that different doses of cocaine tend to produce the same level of CPP expression (Bardo & Bevins 2000). For example, in the present research (Fig. 3B, right), the time spent in the initially least-preferred chamber during the test session did not differ between rats conditioned with 5, 10, or 20 mg/kg of cocaine in the four-trial procedure. Similar results have been observed in other analyses of the dose-effect function for cocaine CPP in rats (Nomikos & Spyraki 1988; Durazzo et al. 1994; O'Dell et al. 1996) as well as in mice conditioned with cocaine (Brabant et al. 2005) or amphetamine (Mead et al. 1999), although exceptions to the all-or-none dose-effect function have been observed (Allan et al. 2001). Thus, the criterion method of data analysis, which classifies individual subjects as “CPP expressing” or “non-CPP expressing” parallels previous analyses that suggest a CPP can be observed following conditioning only with certain doses of cocaine (CPP is expressed) and not with other doses (CPP is not expressed). The criterion method may serve as an adjunct to traditional methods of analysis to help detect dose-effect functions for cocaine CPP, as the criterion detected an additional effective dose of cocaine (Fig. 3A, left). By converting the continuous variable of time into the discrete, “CPP expressing” vs “non-CPP expressing” classification, the ability to correlate time with another variable (i.e., locomotor activity) is lost. Thus, the criterion method of analysis is more valuable in some applications (presence of discrete groups) than others.
Our approach to establishing a criterion was to analyze a large body of CPP data collected in our laboratory using a three-chamber apparatus. Based on these data, we have established a criterion of 324 s spent in the initially least-preferred chamber during the test session. While the present data suggest that the 324 s criterion is appropriate for data collected in the commercially available three-chamber apparatus used in our laboratory, the applicability of this criterion will need further study due to the variability in the methods and equipment used in CPP studies and the inherent variability in behavioral procedures across laboratories (Crabbe et al. 1999). We anticipate that different labs may need to utilize different numerical values for the criterion. However, even if the exact criterion value cannot be applied in other labs, our method of compiling a large body of data from control and drug conditioned groups and subjecting the data to CART analysis to identify a criterion is widely applicable, and opens the possibility for distinguishing “CPP expressing” from “non-CPP expressing” subjects within a treatment group. Because this method can be so easily applied by other laboratories, a strength of this technique may prove to be the wide flexibility and adaptability of this type of analysis.
Our paradigm employed an apparatus in which subjects showed no initial preference for a conditioning chamber (unbiased apparatus) coupled with a biased design in which cocaine administration was paired with the chamber that was determined to be least-preferred for each subject. Control animals developed no natural bias with repeated exposure to the chamber during conditioning, as evidenced by equal time spent in the conditioning chambers during the test session. Although the use of a biased design has been criticized as difficult to interpret, the literature suggests that there is no difference in the magnitude of the CPP (assessed by three different dependent variables comparing control and ethanol-conditioned mice) when either a biased or unbiased design (chamber paired with conditioning drug is chosen randomly) is employed in an unbiased apparatus (Cunningham et al. 2003). Similarly, the magnitude of the morphine CPP established in a biased versus unbiased design did not differ (Blander et al. 1984), and preliminary data collected in our laboratory also found no difference between a biased and an unbiased design on the expression of cocaine CPP (Herin and Cunningham, unpublished observation). Thus, the biased design appears to be a valid method for the conditioning of a CPP.
The possibility of separating a cocaine conditioned group into “CPP expressing” and “non-CPP expressing” subjects may be particularly useful for investigators interested in the molecular mechanisms that drive expression of CPP. The “non-CPP expressing” animals serve as an ideal control, in that they have undergone the identical drug exposure and conditioning paradigm but failed to demonstrate CPP expression. Thus, changes in molecular targets observed in the “CPP expressing,” but not in the “non-CPP expressing,” rats can be considered as attributable to successful CPP conditioning.
An inhibitory role for the 5-HT2CR over the cellular and behavioral effects of cocaine has been well documented (Callahan & Cunningham 1995; Fletcher et al. 2002; Filip & Cunningham 2003; Filip et al. 2004), including for hyperactivity conditioned to cocaine (Liu & Cunningham 2006). Like CPP, the conditioned hyperactivity paradigm assesses the acquisition and expression of behavior that has become associated with cocaine exposure. In this assay, drug-free animals show hyperactivity in the test environment previously paired with cocaine. We previously demonstrated that treatment with the preferential 5-HT2CR agonist MK 212 significantly decreased, and the 5-HT2CR antagonist SB 242084 increased, cocaine-conditioned hyperactivity (Liu & Cunningham 2006), suggesting an inhibitory role for the 5-HT2CR over expression of a cocaine-evoked conditioned association. In the present study, MK 212, at a dose that did not alter basal motility (Filip et al. 2004) nor support acquisition of a CPP or conditioned place aversion (dela Cruz and Cunningham, unpublished observation), significantly suppressed expression of a single-trial cocaine CPP as assessed by the criterion analysis. Treatment with MK 212 also suppressed cue-induced reinstatement of cocaine self-administration (Neisewander & Acosta 2007), further evidence for an inhibitory role for the 5-HT2CR over the behavioral response to cocaine-associated cues. The U-shaped dose response relationship for MK 212 is in keeping with the effects of MK 212 on cocaine-induced locomotor activity (Filip et al. 2004). The shape of the dose response curve may be related to the partial selectively of MK 212 for the 5-HT2CR relative to other 5-HT2R (Kennett 1993), especially given that the 5-HT2AR and 5-HT2CR have been shown to have opposing effects on cocaine-regulated behaviors (Bubar & Cunningham 2008). This effect may also be related to the heterogeneity among populations of 5-HT2CR (for review see Bubar & Cunningham 2008). Thus, the value of employing both traditional and criterion methods of analysis is demonstrated by our ability to reveal the suppressive effect of MK 212 on a subset of CPP expression using the criterion analysis.
In conclusion, we utilized classification and regression tree analysis to identify a criterion for the expression of cocaine CPP that is reliable across multiple conditioning paradigms used in our laboratory. Using this criterion to determine the percentage of subjects within a treatment group that express a CPP, we demonstrate an enhanced sensitivity of this method to identify pharmacological blockade of CPP expression in a distinct sub-group.
Acknowledgements
This work was supported by NIDA DA 020314 (AMD), DA 015272 (DVH), DA 07287 (KAC), DA 06511 (KAC), and DA 16905 (KAC). The authors thank Dr. Marcy Bubar Jordan for her helpful comments during the writing of this manuscript and Bob Fox for expert technical assistance. Portions of this work were presented at the 2007 annual meeting of the College on Problems of Drug Dependence, Quebec City, Quebec.
References
- 1.Adams JU, Careri JM, Efferen TR, Rotrosen J. Differential effects of dopamine antagonists on locomotor activity, conditioned activity and conditioned place preference induced by cocaine in rats. Behav Pharmacol. 2001;12:603–611. doi: 10.1097/00008877-200112000-00004. [DOI] [PubMed] [Google Scholar]
- 2.Allan AM, Galindo R, Chynoweth J, Engel SR, Savage DD. Conditioned place preference for cocaine is attenuated in mice over-expressing the 5-HT(3) receptor. Psychopharmacology (Berl) 2001;158:18–27. doi: 10.1007/s002130100833. [DOI] [PubMed] [Google Scholar]
- 3.Bardo MT, Bevins RA. Conditioned place preference: what does it add to our preclinical understanding of drug reward? Psychopharmacology (Berl) 2000;153:31–43. doi: 10.1007/s002130000569. [DOI] [PubMed] [Google Scholar]
- 4.Belzung C, Barreau S. Differences in drug-induced place conditioning between BALB/c and C57Bl/6 mice. Pharmacol Biochem Behav. 2000;65:419–423. doi: 10.1016/s0091-3057(99)00212-9. [DOI] [PubMed] [Google Scholar]
- 5.Bevins RA, Cunningham CL. Place conditioning: a methodological analysis. In: Anderson MJ, editor. Tasks and Techniques: A Sampling of Methodologies for the Investigation of Animal Learning, Behavior, and Cognition. New York: Nova Science Publishers, Inc; 2006. pp. 99–110. [Google Scholar]
- 6.Blander A, Hunt T, Blair R, Amit Z. Conditioned place preference: an evaluation of morphine's positive reinforcing properties. Psychopharmacology (Berl) 1984;84:124–127. doi: 10.1007/BF00432040. [DOI] [PubMed] [Google Scholar]
- 7.Brabant C, Quertemont E, Tirelli E. Influence of the dose and the number of drug-context pairings on the magnitude and the long-lasting retention of cocaine-induced conditioned place preference in C57BL/6J mice. Psychopharmacology (Berl) 2005;180:33–40. doi: 10.1007/s00213-004-2138-6. [DOI] [PubMed] [Google Scholar]
- 8.Bubar MJ, Cunningham KA. Prospects for serotonin 5-HT2R pharmacotherapy in psychostimulant abuse. Prog Brain Res. 2008;172:319–346. doi: 10.1016/S0079-6123(08)00916-3. [DOI] [PubMed] [Google Scholar]
- 9.Callahan PM, Cunningham KA. Modulation of the discriminative stimulus properties of cocaine by 5-HT1B and 5-HT2C receptors. J Pharmacol Exp Ther. 1995;274:1414–1424. [PubMed] [Google Scholar]
- 10.Carr G, Fibiger HC, Phillips AG. Conditioned place preference as a measure of drug reward. In: Liebman JM, Cooper SJ, editors. The Neuropharmacological Basis of Reward. Oxford, U.K.: Clarendon; 1989. pp. 264–319. [Google Scholar]
- 11.Childress AR, Ehrman R, Rohsenow DJ, Robbins SJ, O'Brien CP. Classically conditioned factors in drug dependence. In: Lowinson JH, Ruiz P, Millman RB, Langrod JG, editors. Substance Abuse: A Comprehensive Textbook. Baltimore: Williams and Wilkins; 1992. pp. 56–69. [Google Scholar]
- 12.Childress AR, Mozley PD, McElgin W, Fitzgerald J, Reivich M, O'Brien CP. Limbic activation during cue-induced cocaine craving. Am J Psychiatry. 1999;156:11–18. doi: 10.1176/ajp.156.1.11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Crabbe JC, Wahlsten D, Dudek BC. Genetics of mouse behavior: interactions with laboratory environment. Science. 1999;284:1670–1672. doi: 10.1126/science.284.5420.1670. [DOI] [PubMed] [Google Scholar]
- 14.Cunningham CL, Ferree NK, Howard MA. Apparatus bias and place conditioning with ethanol in mice. Psychopharmacology (Berl) 2003;170:409–422. doi: 10.1007/s00213-003-1559-y. [DOI] [PubMed] [Google Scholar]
- 15.Daza-Losada M, Ribeiro Do CB, Manzanedo C, Aguilar MA, Rodriguez-Arias M, Minarro J. Rewarding Effects and Reinstatement of MDMA-Induced CPP in Adolescent Mice. Neuropsychopharmacology. 2007;32:1750–1759. doi: 10.1038/sj.npp.1301309. [DOI] [PubMed] [Google Scholar]
- 16.Durazzo TC, Gauvin DV, Goulden KL, Briscoe RJ, Holloway FA. Cocaine-induced conditioned place approach in rats: the role of dose and route of administration. Pharmacol Biochem Behav. 1994;49:1001–1005. doi: 10.1016/0091-3057(94)90255-0. [DOI] [PubMed] [Google Scholar]
- 17.Filip M, Cunningham KA. Hyperlocomotive and discriminative stimulus effects of cocaine are under the control of serotonin(2C) (5-HT(2C)) receptors in rat prefrontal cortex. J Pharmacol Exp Ther. 2003;306:734–743. doi: 10.1124/jpet.102.045716. [DOI] [PubMed] [Google Scholar]
- 18.Filip M, Bubar MJ, Cunningham KA. Contribution of serotonin (5-hydroxytryptamine; 5-HT) 5-HT2 receptor subtypes to the hyperlocomotor effects of cocaine: acute and chronic pharmacological analyses. J Pharmacol Exp Ther. 2004;310:1246–1254. doi: 10.1124/jpet.104.068841. [DOI] [PubMed] [Google Scholar]
- 19.Fletcher PJ, Grottick AJ, Higgins GA. Differential effects of the 5-HT(2A) receptor antagonist M100907 and the 5-HT(2C) receptor antagonist SB242084 on cocaine-induced locomotor activity, cocaine self-administration and cocaine-induced reinstatement of responding. Neuropsychopharmacology. 2002;27:576–586. doi: 10.1016/S0893-133X(02)00342-1. [DOI] [PubMed] [Google Scholar]
- 20.Isaac WL, Nonneman AJ, Neisewander J, Landers T, Bardo MT. Prefrontal cortex lesions differentially disrupt cocaine-reinforced conditioned place preference but not conditioned taste aversion. Behav Neurosci. 1989;103:345–355. doi: 10.1037//0735-7044.103.2.345. [DOI] [PubMed] [Google Scholar]
- 21.Kennett GA. 5-HT1C receptors and their therapeutic relevance. Current Opinion in Investigational Drugs. 1993;2:317–362. [Google Scholar]
- 22.Kupperman M. On Comparing Two Observed Frequency Counts. Applied Statistics. 1960;9:37–42. [Google Scholar]
- 23.Lemon SC, Roy J, Clark MA, Friedmann PD, Rakowski W. Classification and regression tree analysis in public health: methodological review and comparison with logistic regression. Ann Behav Med. 2003;26:172–181. doi: 10.1207/S15324796ABM2603_02. [DOI] [PubMed] [Google Scholar]
- 24.Lewis RJ. An introduction to classification and regression tree (CART) analysis. 2000 [Google Scholar]
- 25.Liu S, Cunningham KA. Serotonin(2C) receptors (5-HT(2C)R) control expression of cocaine-induced conditioned hyperactivity. Drug Alcohol Depend. 2006;81:275–282. doi: 10.1016/j.drugalcdep.2005.07.007. [DOI] [PubMed] [Google Scholar]
- 26.Martin-Iverson MT, Reimer AR. Classically conditioned motor effects do not occur with cocaine in an unbiased conditioned place preferences procedure. Behav Pharmacol. 1996;7:303–314. doi: 10.1097/00008877-199608000-00001. [DOI] [PubMed] [Google Scholar]
- 27.Mead AN, Vasilaki A, Spyraki C, Duka T, Stephens DN. AMPA-receptor involvement in c-fos expression in the medial prefrontal cortex and amygdala dissociates neural substrates of conditioned activity and conditioned reward. Eur J Neurosci. 1999;11:4089–4098. doi: 10.1046/j.1460-9568.1999.00828.x. [DOI] [PubMed] [Google Scholar]
- 28.Neisewander JL, Acosta JI. Stimulation of 5-HT2C receptors attenuates cue and cocaine-primed reinstatement of cocaine-seeking behavior in rats. Behav Pharmacol. 2007;18:791–800. doi: 10.1097/FBP.0b013e3282f1c94b. [DOI] [PubMed] [Google Scholar]
- 29.Nomikos GG, Spyraki C. Cocaine-induced place conditioning: importance of route of administration and other procedural variables. Psychopharmacology (Berl) 1988;94:119–125. doi: 10.1007/BF00735892. [DOI] [PubMed] [Google Scholar]
- 30.O'Brien MS, Anthony JC. Risk of becoming cocaine dependent: epidemiological estimates for the United States, 2000–2001. Neuropsychopharmacology. 2005;30:1006–1018. doi: 10.1038/sj.npp.1300681. [DOI] [PubMed] [Google Scholar]
- 31.O'Dell LE, Khroyan TV, Neisewander JL. Dose-dependent characterization of the rewarding and stimulant properties of cocaine following intraperitoneal and intravenous administration in rats. Psychopharmacology (Berl) 1996;123:144–153. doi: 10.1007/BF02246171. [DOI] [PubMed] [Google Scholar]
- 32.Roma PG, Riley AL. Apparatus bias and the use of light and texture in place conditioning. Pharmacol Biochem Behav. 2005;82:163–169. doi: 10.1016/j.pbb.2005.08.004. [DOI] [PubMed] [Google Scholar]
- 33.Sheskin DJ. Handbook of parametric and nonparametric statistical procedures. Boca Raton: Chapman & Hall/CRC; 2004. [Google Scholar]
- 34.Shippenberg TS, Heidbreder C. Sensitization to the conditioned rewarding effects of cocaine: pharmacological and temporal characteristics. J Pharmacol Exp Ther. 1995;273:808–815. [PubMed] [Google Scholar]
- 35.Spyraki C, Fibiger HC, Phillips AG. Cocaine-induced place preference conditioning: lack of effects of neuroleptics and 6-hydroxydopamine lesions. Brain Res. 1982;253:195–203. doi: 10.1016/0006-8993(82)90686-2. [DOI] [PubMed] [Google Scholar]
- 36.Zhang H, Holford T, Bracken MB. A tree-based method of analysis for prospective studies. Stat Med. 1996;15:37–49. doi: 10.1002/(SICI)1097-0258(19960115)15:1<37::AID-SIM144>3.0.CO;2-0. [DOI] [PubMed] [Google Scholar]