

Abstract
Adoptive transfer of ex vivo expanded CD4+CD25+FOXP3+ regulatory T cells is a successful therapy for autoimmune diseases and transplant rejection in experimental models. In man, equivalent manipulations in bone marrow transplant recipients appear safe, but questions regarding the stability of the transferred regulatory T cells during inflammation remain unresolved. In this study, protocols for the expansion of clinically useful numbers of functionally suppressive and stable human regulatory T cells were investigated. Regulatory T cells were expanded in vitro with rapamycin and/or all-trans retinoic acid and then characterized under inflammatory conditions in vitro and in vivo in a humanized mouse model of graft-versus-host disease. Addition of rapamycin to regulatory T-cell cultures confirms the generation of high numbers of suppressive regulatory T cells. Their stability was demonstrated in vitro and substantiated in vivo. In contrast, all-trans retinoic acid treatment generates regulatory T cells that retain the capacity to secrete IL-17. However, combined use of rapamycin and all-trans retinoic acid abolishes IL-17 production and confers a specific chemokine receptor homing profile upon regulatory T cells. The use of purified regulatory T-cell subpopulations provided direct evidence that rapamycin can confer an early selective advantage to CD45RA+ regulatory T cells, while all-trans retinoic acid favors CD45RA− regulatory T-cell subset. Expansion of regulatory T cells using rapamycin and all-trans retinoic acid drug combinations provides a new and refined approach for large-scale generation of functionally potent and phenotypically stable human regulatory T cells, rendering them safe for clinical use in settings associated with inflammation.
Introduction
Naturally occurring, thymus derived, CD4+CD25hiFOXP3+ T regulatory cells (Tregs) play a critical role in shaping many types of immune responses. They maintain peripheral tolerance to self-antigens, thereby controlling autoimmune diseases and limiting immune responses to foreign antigens such as pathogens and transplanted organs. In this respect, defective numbers or functions of Tregs have been associated with the pathogenesis of autoimmune diseases.1 In addition to their physiological role in vivo, it has emerged from experimental models that adoptive transfer of Tregs can ameliorate autoimmune disease, graft-versus-host disease (GvHD) and also prevent solid organ transplant rejection.2 These findings suggest that clinical therapy with human Tregs for the treatment of autoimmune diseases or for the induction of transplantation tolerance represents a promising strategy. Indeed, clinical trials using Tregs are already underway to prevent or treat GvHD.3–5 The majority of clinical trials published to date have utilized fresh, immunomagnetic bead-isolated Tregs derived from either cord or adult blood since clinical grade cell sorting facilities are not widely available. Good Manufacturing Practice procedures for CD4+CD25+ T-cell enrichment yield Treg preparations with a purity that is lower than 70%.6 In addition, the requirement to expand Tregs in vitro to achieve the number required for a therapeutic benefit further compromise the purity of Tregs at the end of the culture. Therefore, one of the major goals in Treg therapy is to maintain the purity and suppressive ability of Treg preparation after in vitro expansion and the requirement to limit the potential of expansion of cells that have the potential to produce pro-inflammatory cytokines, such as IL-17, particularly when Tregs are exposed to an inflammatory environment in vivo. Although both these limitations may be in part due to the presence of ‘contaminating’ effector T cells within bead-separated Treg preparations, the capacity for conversion of human Tregs into IL-17-producing cells has been well demonstrated.7,8 The application of tolerance permissive drugs to enhance Treg expansion in vitro has been investigated in recent years. Rapamycin (RAPA), an mTOR kinase inhibitor, is an immunosuppressive drug that inhibits effector T-cell proliferation, migration and cytokine production,9 and can selectively promote expansion of suppressive human CD4+CD25hiFOXP3+ T cells isolated from healthy donors and patients with diabetes.10,11 Likewise, all-trans retinoic acid (ATRA), a vitamin A metabolite, in combination with TGF-β, promotes differentiation of naive human and murine T cells into Tregs12–14 and more recently has been shown to increase in vitro Treg function.15 Selective retinoic acid receptor alpha (RARα) gene deletion in animal models results in significant loss of FOXP3 expression in Tregs, suggesting that ATRA may act to stabilize FOXP3 expression.16 As such, ATRA could represent a potential tool in combination with RAPA for ex vivo expansion of highly suppressive Tregs.17 However, as ATRA has been shown to play an important role in T-helper cell fate decisions, by inducing both effector T-cell (Teff) activation and differentiation to Th1 and Th17,18 its use for Treg expansion warrants further investigation.
To help formulate an optimal protocol for the expansion of bead-separated Tregs for clinical use, and to gain more insight into the effects of RAPA and ATRA on Tregs, these drugs were tested for their adjunctive effect on the expansion of human CD4+CD25+ Tregs in vitro. Here we demonstrate that RAPA and ATRA are equally effective in supporting the expansion of large numbers of pure, highly suppressive Tregs. Furthermore, these treatments maintained or up-regulated distinct molecular markers on expanded Tregs, including chemokine receptors. More importantly, we demonstrated in vitro and for the first time in vivo in a pre-clinical model of XenoGvHD that while RAPA treatment inhibited IL-17 expression by Tregs, ATRA was permissive for both IL-17 and IFN-γ production.
In order to shed some light on the different effects of drug treatments on Treg plasticity, we studied the fate of the Treg subpopulations identified by Miyara et al.19 Three phenotypically and functionally separate populations based on the expression of FOXP3 and CD45RA, namely CD4+CD25+CD45RA+ (P1), CD4+CD25hiCD45RA− (P2) and CD4+CD25+CD45RA− (P3) were identified and studied.19 In characterizing these subpopulations, the authors showed that while P1 had a stable phenotype, P3 had the potential to release pro-inflammatory cytokines. Our results clearly and directly demonstrated that the two drugs studied act by selecting and maintaining distinct subsets of Tregs during the in vitro expansion.
Design and Methods
Cell isolation and separation
Peripheral blood mononuclear cells (PBMC) from healthy donors were obtained from anonymized human leukocyte cones supplied by the National Blood Transfusion Service (NHS Blood and Transplantation, Tooting, London, UK). Human studies were conducted in accordance with the Declaration of Helsinki and approved by the Institutional Review Board of Guy’s Hospital (reference 09/H0707/86). Informed consent was obtained from all healthy donors prior to enrollment into the study. PBMC were isolated by Lymphocyte (PAA, Austria) density gradient centrifugation. CD4+CD25+ T cells were purified by negative selection of CD4+ T cells followed by positive selection of CD25+ T cells using miniMACS CD4+CD25+ T Regulatory Cell Isolation Kit (Miltenyi-Biotec, UK). The purity of CD4+CD25+ T cells was between 90–98%. CD4+CD25+ T cells were expanded using different treatments while aliquots of CD4+CD25− T cells (effector T cells, Teff) were cryopreserved and used as autologous responder cells in suppression assays. To sort Treg subpopulations, CD4+ T cells were enriched by negative selection and cells were stained with anti-CD4, anti-CD25 and anti-CD45RA antibodies to sort them into CD4+CD25+CD45RA+, CD4+CD25+CD45RA− and CD4+CD25hiCD45RA− T cells on a 3-laser FACS-Aria high-speed cell sorter (BD Biosciences).
Expansion of Treg lines
Healthy human Tregs were plated at 1×106/mL in X-Vivo 15 (Lonza, Switzerland) supplemented with 5% human AB serum (HS) (Biosera, UK), containing RAPA (LC-Laboratories, USA) and/or ATRA (Sigma-Aldrich, USA). RAPA (100nM) was used according to previous reports.10,20 Different doses of ATRA (0.002–2 μM) were investigated and 2 μM chosen for in vitro Treg expansion because of its strong biological effect on Treg phenotype. Cells were activated with anti-CD3 and anti-CD28 coated beads (Invitrogen, UK) at a bead:cell ratio of 1:1. IL-2 (1000 IU/mL, Proleukin-Novartis, UK) was added at Day 2 post-activation and replenished every two days. Beads were removed by magnetic adherence every seven days post-activation and fresh anti-CD3/CD28 beads (1:1 ratio), RAPA, ATRA and IL-2 (1000 IU/mL) were added. Expanded cells were used for further analysis 28 days post-activation.
Statistical analysis
Statistical analysis was carried out using GraphPad Prism software (GraphPad Software Inc., USA). Parametric and non-parametric data were calculated as the mean±s.d. and median (interquartile range, IQR), respectively. For comparison of parametric and non-parametric data, t-test, one- or two-way ANOVA with Bonferroni’s test comparison and Kruskal-Wallis test were used where appropriate.
Results
Highly suppressive Tregs are expanded in the presence of RAPA and/or ATRA
The clinical application of Tregs requires elevated numbers of highly suppressive and stable Tregs. Therefore, the effects and mechanisms of action of RAPA and ATRA on the expansion rate of human bead-enriched Tregs were further analyzed and compared. Immunomagnetic bead-separated Tregs, with a purity of more than 90% for FOXP3-positivity on isolation were cultured with anti-CD3/CD28 and IL-2, in the absence (untreated) or presence of RAPA (100nM), ATRA (2μM) or a combination of both drugs (RAPA+ATRA) (Figure 1A). Tregs were harvested weekly for up to 28 days and the co-expression of CD25 and FOXP3 molecules was evaluated (Figure 1A) together with the total cell number measured (Figure 1B and C). Analysis of the population doublings demonstrated that the presence of RAPA affected the expansion rate of Tregs during the first round of stimulation, as previously described by other authors6 (Figure 1B). However, the differences in the expansion rates between the Treg preparations decreased after the initial time of culture and similar Treg numbers were achieved at the second and third re-stimulation (Figure 1B). In addition, the same analysis demonstrated that the mean expansion after 28 days of culture for 10 different Treg lines was comparable between different culture conditions, although some variability was observed between Treg preparations derived from different donors (Figure 1C). Phenotypic analysis showed that no significant differences were detected in the percentage of Tregs expressing CD25 and FOXP3 molecules between the different cell lines at Day 28 (Figure 1A) or at any other time point analyzed (data not shown). However, similar to the findings of Golovina et al.,17 the mean fluorescent intensity (MFI) of CD25 expression was highest following exposure of Tregs to RAPA alone or in combination with ATRA. No significant differences were detected in FOXP3 MFI (Figure 1D).
Figure 1.

Expansion, phenotype and suppressive activity of untreated, RAPA-, ATRA- and RAPA+ATRA-treated human Tregs. (A) CD25 and FOXP3 expression of fresh cells and 28-day untreated or drug-treated Treg lines. Dot plots show a representative example from 7 independent experiments. (B) Population doublings during in vitro expansion from 10 different T-cell lines. Cell counts were performed at weekly intervals (R) throughout culture period. (C) Expansion rates of 10 different 28-day T-cell lines cultured in the presence of only IL-2 (1000 IU/mL) or IL-2 and RAPA (100nM), ATRA (2 μM) or RAPA+ATRA. Mid lines indicate median expansion. (D) Mean fluorescence intensity (MFI) of CD25 and FOXP3 expression in 28-day Treg lines. (E) Suppressive abilities at different ratios of Treg:Teff of 28-day Treg lines. Data, mean ± s.e.m. of 9 independent experiments, are expressed as percentage of inhibition of Teff proliferation. Statistical analysis shown only on significantly different data. *P<0.05, **P<0.01, ***P<0.001.
To compare the regulatory function of each Treg preparation, expanded Treg lines (after 4 restimulations, Day 28) were co-cultured with CFSE-labeled effector T cells (Teff) at different Teff:Treg ratios and examined for their ability to suppress a polyclonal stimulus (Figure 1E). RAPA- and RAPA+ATRA-treated Tregs had the greatest suppressive activity on Teff proliferation, followed by ATRA-conditioned Tregs, while untreated Treg cultures had a significantly reduced suppressive ability in comparison (Figure 1E). On adoptive transfer of ex vivo expanded Tregs, it is possible that Tregs may revert to an ‘untreated’ Treg phenotype in the prolonged absence of the conditioning drug used for expansion. Therefore, the suppressive ability of Tregs was re-evaluated after two weeks of drug withdrawal from cell cultures. Treg suppressive functions were maintained by each Treg line tested (Online Supplementary Figure S1).
RAPA and ATRA treatments confer distinct phenotypic and homing signatures on Tregs
Tregs have been shown to be heterogeneous in the expression of surface molecules. Some of these molecules have been shown to influence their function and migratory ability.21 To analyze phenotypic signatures conferred by RAPA and ATRA on Tregs, the surface expression of several molecules by Tregs was evaluated after 28 days of culture (Online Supplementary Figure S2A). All Treg preparations maintained a high percentage of cells expressing CTLA-4, GITR, ICOS and CD39 molecules, with low expression of CD127 (Online Supplementary Figure S2A). In addition, high percentages of Tregs co-expressing CD62L and CD27 molecules was observed in RAPA- and RAPA+ATRA-treated Treg preparations, while the same markers were expressed in a minority of untreated or ATRA-treated Tregs (Online Supplementary Figure S2B). It is worthy of note that the co-expression of these two molecules by Tregs has been previously associated with high suppressive ability both in vitro and in vivo.2,8,22,23 Finally, to further investigate the effects of different drug treatments on the phenotype of Tregs, we evaluated the expression of HLA-DR (Online Supplementary Figure S2C), previously shown to characterize a mature, functionally distinct sub-population of Tregs with the highest expression of FOXP324 (Online Supplementary Figure S2C). A significant difference in the percentage of HLA-DR+ Tregs was observed between untreated and RAPA-treated cells compared to ATRA-treated Tregs (Online Supplementary Figure S2C). This evidence suggests that ATRA treatment favors the expansion of a subset of Tregs HLA-DR.
Interestingly, the treatments with the two drugs induced expression of very different homing receptors, and this evidence has important relevance for the use of these two drugs in the treatment of diseases with very well defined target organs. After 28 days of culture, RAPA-treatment induced co-expression of skin homing receptor CCR4 and CLA, whereas ATRA-treatment resulted in high percentages of cells co-expressing liver and gut homing receptors CCR9 and α4β7 integrin (Online Supplementary Figure S2D). Finally, Tregs treated with both drugs showed the combined homing characteristic features of Tregs stimulated by each drug (Online Supplementary Figure S2D).
While RAPA treatment inhibits, ATRA maintains inflammatory cytokine production by Tregs
One of the major concerns in Treg therapy is the plasticity of Tregs, as several studies have recently suggested that Tregs can convert to cells producing inflammatory cytokines.25 To evaluate the stability of bead-purified Tregs expanded in the presence of drugs, IL-17 and IFN-γ production was measured by ELISA following RAPA- and/or ATRA-treatment. Analysis of supernatants from 28-day expanded Treg lines (Figure 2A) demonstrated that there was no significant difference in IL-17 and IFN-γ production between untreated and ATRA-treated cells. In contrast, the presence of RAPA strongly reduced IL-17 and IFN-γ production (Figure 2A). To better characterize the nature of IL-17-producing Tregs, the expression of CD161 was analyzed. This has previously been shown to correlate with the ability of T cells to produce IL-17, as well as being described as a marker for precursors of IL-17-producing T cells.26,27 The results obtained demonstrated that while the addition of ATRA to Treg cultures favored expansion of FOXP3+CD161+ Treg, RAPA significantly suppressed proliferation of this subset (Figure 2B). Surprisingly, when both drugs were used in combination, the percentage of CD161+ Tregs was similar to cultures treated with ATRA alone, despite the reduction in IL-17 production, demonstrating that IL-17 release and CD161 expression may not be strictly correlated.
Figure 2.
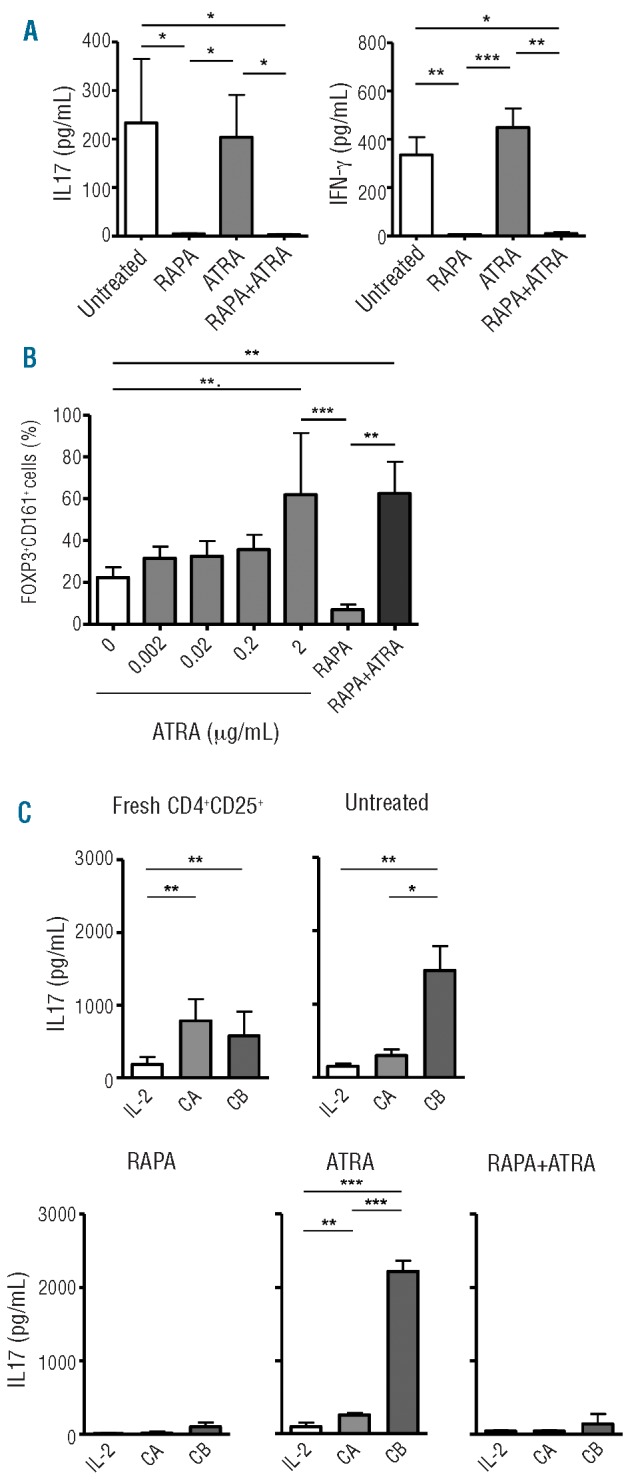
Analysis of Treg plasticity. (A) IL-17 and IFN-γ concentrations in 7-day culture supernatant from 28-day Treg cultures; cumulative mean ± s.d. from 5 experiments. (B) Percentage of FOXP3+CD161+ cells in the presence of RAPA (100 nM), RAPA+ATRA (100 nM and 2 μM, respectively) or different concentrations of ATRA; cumulative mean ± s.d. from 3 independent experiments. (C) Treg stability in the presence of pro-inflammatory milieu. IL-17 concentration in fresh CD4+CD25+ T-cell and Treg culture supernatants. IL-17 concentration was measured after 1-week culture in the absence of drugs and in the presence of only IL-2 (10 IU/mL) or 2 different cytokine cocktails. Cocktail A (CA): IL-2, IL-1β, IL-6, TGF-β. Cocktail B (CB): IL-2, IL-21, IL-23, TGF-β. (See text for details). Graphs show pooled mean ± s.d. from 3 independent experiments. Statistical analysis shown only on significantly different data. *P<0.05, **P<0.01, ***P<0.001.
Treatment of Tregs with RAPA inhibits their conversion to IL-17-producing cells in the presence of pro-inflammatory cytokines both in vitro and in vivo
One of the major risks for Treg therapy is that Tregs may acquire effector functions and lose their suppressive ability during inflammatory responses in vivo. To address this concern, Treg lines were cultured in the presence of pro-inflammatory cytokines previously reported to favor Th17 conversion.8,28 Two cytokine cocktails were selected: cocktail A (IL-1β, IL-2, IL-6 and TGF-β) and cocktail B (IL-2, IL-21, IL-23 and TGF-β). Analysis of supernatants from freshly isolated or 28-day untreated Tregs revealed that IL-17 production was increased by the addition of each cytokine cocktail. In contrast, the presence of RAPA in culture completely inhibited IL-17 production. ATRA-treatment in combination with pro-inflammatory cytokines and, in particular, of cocktail B significantly increased the production of IL-17 by Tregs (Figure 2C).
To extend these in vitro observations to a more relevant in vivo analysis, the plasticity of drug-conditioned Tregs was tested for the first time in a humanized mouse model during Xeno-GvHD29 using NSG mice reconstituted with human HLA-A2- PBMC. HLA-A2+ 28-day Tregs (untreated, RAPA-, ATRA- and RAPA+ATRA-treated) were adoptively transferred into animals with active Xeno-GvHD disease. After 48 h, splenocytes were recovered and HLA-A2+ Tregs analyzed (Figure 3A and Online Supplementary Figure S3). Intracellular staining was performed to evaluate the expression of human IL-17 and IFN-γ cytokines. As shown in Figure 3B, in vitro pre-treatment with RAPA or RAPA+ATRA completely prevented IL-17 and IFN-γ production in vivo while untreated and ATRA-treated Tregs produced both cytokines. The same Treg lines used for the injections were tested in vitro by intracellular staining (Figure 3D), with parallel findings (Figure 3C and E, respectively). Altogether these results demonstrated that the conditioning of Tregs with RAPA leads to the inhibition of IL-17 and IFN-γ production by Tregs both in vitro and in vivo in the presence of intense inflammatory conditions.
Figure 3.

Interleukin 17 and IFN-γ production from in vitro expanded and in vivo infused Tregs. (A) Representative dot plots showing the gating strategy to analyze in vivo infused Tregs. (B and C) IL-17 and IFN-γ production from Treg infused in a humanized mouse model of xeno-GvHD. Cumulative plots of percentages of IL-17+ or IFN-γ+ Tregs from different number of mice (n ≥ 3). Data are representative of 3 independent experiments. (B) In vitro Treg production of IL-17 and IFN-γ. Cumulative plots of percentages of IL-17+ or IFN-γ+ T cells from 6 and 5 Treg lines, respectively. Mid lines indicate mean expression. Statistical analysis shown only on significantly different data. *P<0.05.
RAPA favors naïve Treg subset expansion explaining its capacity to prevent IL-17 conversion
All the data presented so far have been obtained with ‘whole’ Treg preparations. As described before, human Tregs can be divided into at least three subpopulations,19 namely CD4+CD25+CD45RA+ (P1, naïve and stable), CD4+CD25hiCD45RA− (P2, effector) and CD4+CD25+CD45RA− (P3, memory and plastic). To understand whether the inhibitory effect of RAPA on the production of pro-inflammatory cytokines was due to a selective effect of the drug on the most stable of the subpopulations (P1), cell-sorted Treg subsets were cultured with RAPA and/or ATRA (Figure 4A). P2 did not expand at all within any of the in vitro culture conditions (data not shown), while P1 and P3 proliferated in the presence of the different drug treatments (Figure 4B and C). Treg cultures derived from the two subpopulations, obtained after cell sorting, showed equivalent expansion rates to the ‘whole’ Tregs described earlier. The treatments with RAPA or ATRA affected the initial expansion of the two subpopulations. P1 Tregs expanded in the presence of RAPA, while this drug did not affect P3 expansion (Figure 4D). In contrast, ATRA favored the expansion of P3 (Figure 4D).
Figure 4.
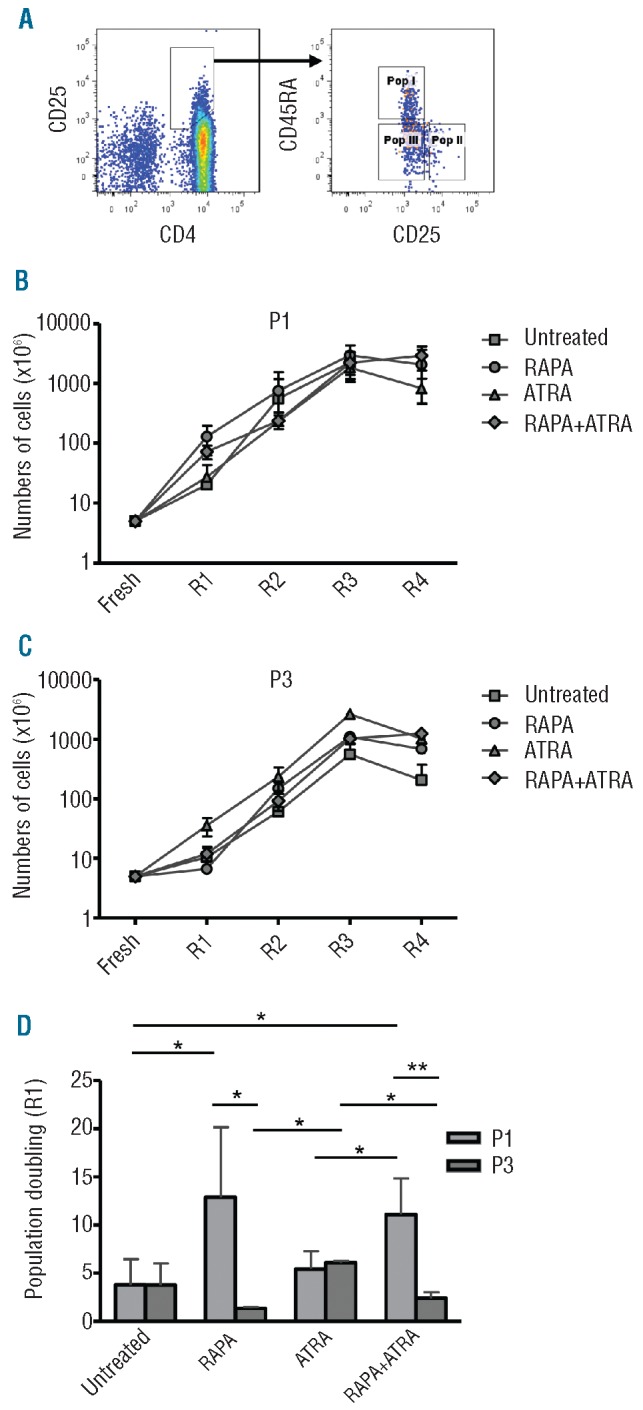
Treg subsets expansion in the presence of different treatment. (A) Gating strategy for Treg subset cell sorting. (B and C) Cumulative data from 5 different P1 (B) and P3 (C) Treg lines after 4 rounds of stimulation. (D) Population doublings after the first round of stimulation (R1) in differently treated P1 and P3 Treg lines. Data from 5 different Treg lines. Graphs show pooled mean ± s.d. Statistical analysis shown only on significantly different data. *P<0.05, **P<0.01.
To further characterize the influence of the drugs on the two Treg subpopulations, the function and stability of each Treg subset after drug-conditioning were investigated. As previously shown for freshly isolated or in vitro expanded Treg subsets,19,30 P1 consistently demonstrated a tendency to have superior suppressive abilities independent of the culture treatment, compared to P3, although statistically significant differences were only observed upon Treg treatment with ATRA (Online Supplementary Figure S4). Cytokine analysis revealed that P1 did not produce IL-17 in any of the culture conditions (Figure 5A), while the same Treg subset showed some IFN-γ production particularly following treatment with ATRA (Figure 5B). The same cytokine analysis showed that untreated and ATRA Tregs derived from P3 produced large amounts of IL-17 that was inhibited only by the addition of RAPA (Figure 5A). No significant differences among the other culture conditions of P3 were observed for IFN-γ production (Figure 5B). The cytokine profile of the two Treg subsets cultured in the different conditions was confirmed by intracellular staining (Figure 5C). The cytokine profile of both subsets correlated very well with the expression of CD161 (Figure 5D). Tregs from P3 contained the highest percentage of cells expressing CD161 molecule, which was decreased when P3 was cultured with RAPA (Figure 5D). In contrast, P1 showed a lower number of cells expressing CD161, and again RAPA was able to further decrease it.
Figure 5.
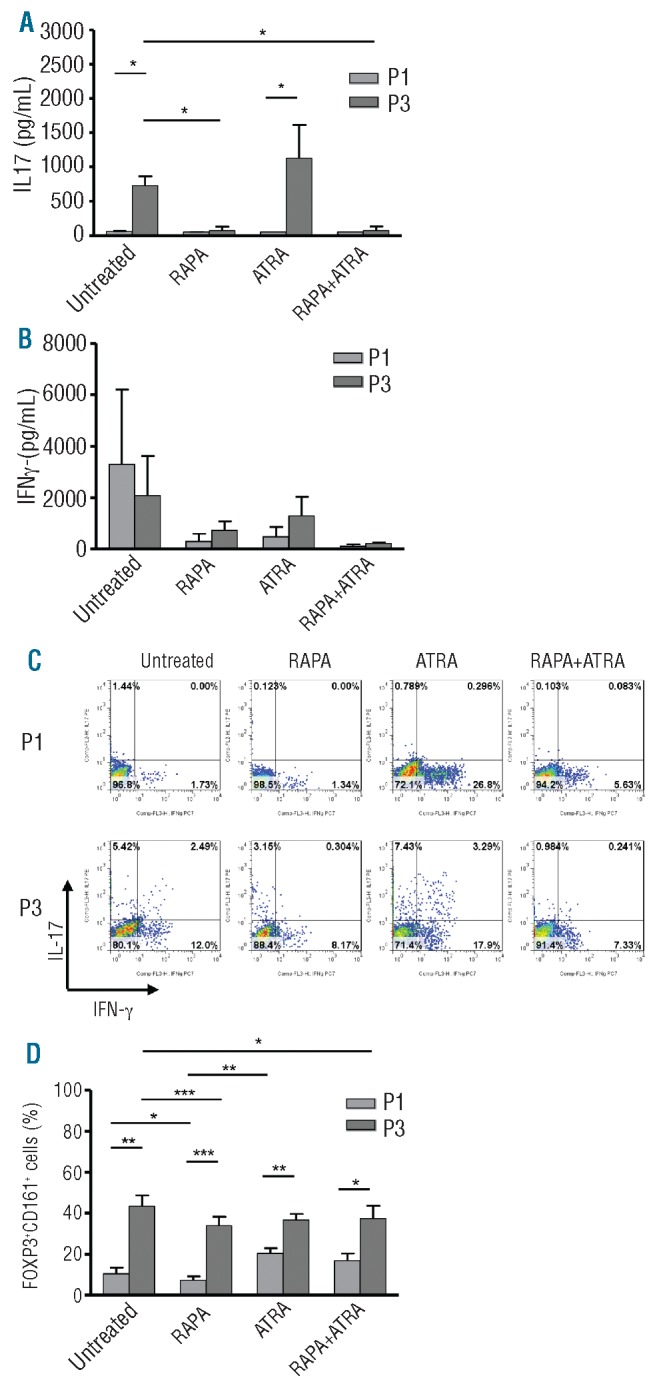
Treg subset stability and CD161 expression in the presence of different treatment. (A and B) Comparison of IL-17 (A) and IFN-γ (B) production by ELISA in 7-day supernatants of P1 and P3 Treg lines. (C) Representative dot plots showing by intracellular staining the percentage of IL-17+ and IFN-γ+ cells in CD25+FOXP3+ P1 and P3 cultured with different treatments. (D) Comparison of CD161 expression in P1 and P3 Treg lines. All data are from Treg lines after 4 rounds of stimulation. Plots are representative of 5 independent experiments. Statistical analysis shown only on significantly different data. *P<0.05, **P<0.01, ***P<0.001.
Discussion
In recent years, there has been an increasing interest in ex vivo expanded human CD4+CD25+FOXP3+ Tregs as a cell therapy for the induction of tolerance.31 The challenge in translating the therapeutic potential of Tregs is to expand clinically sufficient numbers of Tregs in vitro, which are highly pure, suppressive, and stable, with well defined Treg homing characteristics for in vivo Treg targeting to specific sites in order to optimize therapeutic benefit. In this study, we examined the use of RAPA and/or ATRA as suitable reagents to facilitate the expansion of natural Tregs for immunotherapeutic use from bead-separated populations. Results indicate how these two drugs affect the phenotype, suppressive and expansion properties of Tregs. Our study has focused in particular on Treg plasticity and attempted to identify ex vivo expansion protocols with the capacity to control it. We conclude that RAPA or the combined use of RAPA and ATRA provide an advantage for the expansion of a potent suppressive Treg subset displaying a broad range of homing receptors which, most importantly, sustains functional stability both in vitro and in vivo.
Previous studies have identified RAPA as an ideal treatment to preferentially expand natural Tregs. RAPA has been shown to give a proliferative advantage to Tregs by affecting basic signal pathways such as Akt/mTor, which is not essential for Tregs.9 Other studies have shown that ATRA is also a suitable treatment for the induction of adaptive Tregs.14 Indeed, recently ATRA has been described as a treatment to expand human nTregs.17 The molecular pathway by which ATRA favors the expansion of Tregs is not completely clear, but it is thought to induce chromatin decondensation recruiting histone acetyltransferases and transcription machinery to the FOXP3 promoter.32,33 However, due to its multi-faceted effects on gene transcription, ATRA has also been shown to affect T-cell fate by contributing to Th1/Th17 differentiation.18 We have also identified significant differences between untreated, RAPA- and ATRA-treated Tregs in terms of homing receptor expression. Although others have previously reported that Tregs express multiple homing receptors34,35 such as CD62L,30,36,37 CCR4,34,37,38 CLA34,37,39 and CCR7,37,40 this is the first study in which the expression of homing receptors was evaluated on human Tregs during ex vivo expansion with drug conditioning. The migration of these Treg preparations in vivo is now under investigation.
One of the major concerns in Treg therapy is to prevent the induction and expansion of IL-17-producing cells which could arise from Teffs contaminating the Treg preparations,41 and/or FOXP3+ Tregs converting to IL-17-producing cells.8 Our data clearly show that RAPA has an inhibitory effect on the development of IL-17-producing cells, both in vitro and in vivo. This is consistent with previous descriptions in the literature of the inhibitory effect of RAPA on Th17 cells.29 In contrast, we found that ATRA maintained the expansion of IL-17 and IFN-γ producing FOXP3+ Tregs. The effect of ATRA on Th17 cells has been controversial. Some in vitro studies have shown that retinoic acid induces pro-inflammatory T-cell responses while others demonstrated that ATRA strongly inhibits Th17 polarization in vitro.12,13,42 Furthermore, Hall and coworkers have demonstrated in vivo that in mice fed with a vitamin A deficient diet, robust Th17 and Th1 responses are generated.18 The general conclusion is that the effect of retinoic acid is very much dependent upon its concentration and the inflammatory environment in which the immune response takes place.15,18,42 In this study, we demonstrate that ATRA expanded Tregs contained a small percentage of cells that lack the expression of HLA-DR and that these cells may be involved in the IL-17 production observed in this cell population.
To further interpret the results obtained with drug conditioning of ‘whole’ Tregs, Treg subsets19 were cell sorted and cultured with RAPA and/or ATRA. Our results demonstrate that the presence of RAPA provided an initial proliferative advantage to P1, while the same drug had an inhibitory effect on the P3 subset. In contrast, ATRA conditioning favored expansion of cells in P3. This leads to the hypothesis that when the two drugs were used with ‘whole’ Tregs, RAPA expands P1 to the detriment of P3, while ATRA allows the expansion of cells within the P3 that retain IL-17 secretion capacity. It is interesting to note that analysis of CD161 in general confirms this hypothesis by showing a higher percentage of Tregs expressing this marker in P3. However, when P3 is cultured with the combination of RAPA and ATRA, the same percentage of Tregs expressed CD161 while IL-17 production was reduced, suggesting that these two parameters do not necessarily correlate. In addition, it is also not clear why the percentage of Tregs expressing CD161 molecule increased when ‘whole’ Tregs are treated with ATRA but not when the single subpopulations are treated with the same drug. This result may be explained by the influence of the two subpopulations on each other when ‘whole’ Tregs are treated with ATRA; this needs further investigation.
Previous reports have clearly demonstrated the ability of human FOXP3+ Tregs, either freshly isolated or in vitro expanded in the presence43 or absence44,45 of rapamycin to ameliorate GvHD. In this manuscript, we have further extended this analysis by showing that ex vivo expanded Tregs conditioned with RAPA alone or in combination with ATRA remain functionally stable in the presence of a severe systemic inflammatory response, neither converting to IL-17 producing cells in vivo nor causing deterioration in clinical disease severity. These results are particularly relevant to the use of Tregs as an adoptive cell therapy. Very recently, two other groups have investigated the stability of Tregs in vivo using different models and protocols.6,28 Hippen and co-workers have shown that although human Tregs produced IL-17 in vitro, once injected in vivo together with peripheral blood mononuclear cells (PBMC), they remained functional and inhibited xenoGvHD.6 These results are very similar to our findings in vitro in which ATRA-treated Tregs, although they produce IL-17, are highly suppressive. These results provide some evidence that the observed level of IL-17-production by Tregs does not influence their capacity to suppress. Tresoldi et al. showed that RAPA-expanded Tregs did not produce IL-17 after injection of Tregs into NSG mice.28 However the transfer of Tregs was performed in the absence of PBMC injection, relying on the irradiation of the animals to produce in vivo inflammation that we predict would have been mostly composed of murine cytokines.28 The data presented in our study provide the first direct evidence that RAPA is a key drug in conferring stability on expanded Tregs even when injected in vivo during an inflammatory immune response.
In conclusion, we have identified a clinically applicable protocol for the expansion of human immunomagnetic bead-separated CD4+CD25+ Tregs. We present here additional data performed in vitro, and in particular in vivo, that describe how these two drugs affect Treg phenotype, function and plasticity during in vitro cell expansion. We demonstrate directly for the first time that one of the ways that RAPA stabilizes Tregs is by selecting and expanding the subpopulation of functional, not plastic naïve Tregs.19,30 In contrast, the effect that we observed with ATRA can be explained by the fact that this drug favors the expansion of P3 that contains cells that retain the capacity to produce IL-17. Although our findings further emphasize that Treg therapy with bead-separated Tregs requires RAPA for the expansion of functional and stable Tregs, the selection of an additional drug, such as ATRA, can be based on the type of organ or tissue targeted. We suggest that the combination of ATRA and RAPA may be an ideal culture condition to expand Tregs for their use in the induction of tolerance in inflammatory bowel disease (IBD).
Footnotes
The online version of this article has a Supplementary Appendix.
Funding
The research was funded by the National Institute for Health Research (NIHR) Biomedical Research Centre based at Guy’s and St Thomas’ NHS Foundation Trust and King’s College London, UK.. The views expressed are those of the authors and not necessarily those of the NHS, the NIHR or the Department of Health. The authors acknowledge financial support from the Medical Research Council (to BA and RIL), the British Heart Foundation, Guy’s and St Thomas’ Charity Trust (RIL and GL), the EU FP7 EU Framework 7 Research Projects NAIMIT (to MP), and ONE STUDY (to CS). The authors acknowledge the support of the MRC Centre for Transplantation at King’s College London, UK.
Authorship and Disclosures
Information on authorship, contributions, and financial & other disclosures was provided by the authors and is available with the online version of this article at www.haematologica.org.
References
- 1.Miyara M, Gorochov G, Ehrenstein M, Musset L, Sakaguchi S, Amoura Z. Human FoxP3(+) regulatory T cells in systemic autoimmune diseases. Autoimmun Rev. 2011;10(12):744–55 [DOI] [PubMed] [Google Scholar]
- 2.Issa F, Hester J, Goto R, Nadig SN, Goodacre TE, Wood K. Ex vivo-expanded human regulatory T cells prevent the rejection of skin allografts in a humanized mouse model. Transplantation. 2010;90(12):1321–7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Di Ianni M, Falzetti F, Carotti A, Terenzi A, Castellino F, Bonifacio E, et al. Tregs prevent GVHD and promote immune reconstitution in HLA-haploidentical transplantation. Blood. 2011;117(14):3921–8 [DOI] [PubMed] [Google Scholar]
- 4.Trzonkowski P, Bieniaszewska M, Juscinska J, Dobyszuk A, Krzystyniak A, Marek N, et al. First-in-man clinical results of the treatment of patients with graft versus host disease with human ex vivo expanded CD4+CD25+CD127− T regulatory cells. Clin Immunol. 2009;133(1):22–6 [DOI] [PubMed] [Google Scholar]
- 5.Brunstein CG, Miller JS, Cao Q, McKenna DH, Hippen KL, Curtsinger J, et al. Infusion of ex vivo expanded T regulatory cells in adults transplanted with umbilical cord blood: safety profile and detection kinetics. Blood. 2011;117(3):1061–70 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hippen KL, Merkel SC, Schirm DK, Sieben CM, Sumstad D, Kadidlo DM, et al. Massive ex Vivo Expansion of Human Natural Regulatory T Cells (Tregs) with Minimal Loss of in Vivo Functional Activity. Sci Transl Med. 2011;3(83):83ra41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Afzali B, Lombardi G, Lechler RI, Lord GM. The role of T helper 17 (Th17) and regulatory T cells (Treg) in human organ transplantation and autoimmune disease. Clin Exp Immunol. 2007;148(1):32–46 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Koenen HJ, Smeets RL, Vink PM, van Rijssen E, Boots AM, Joosten I. Human CD25highFoxp3pos regulatory T cells differentiate into IL-17-producing cells. Blood. 2008;112(6):2340–52 [DOI] [PubMed] [Google Scholar]
- 9.Peter C, Waldmann H, Cobbold SP. mTOR signalling and metabolic regulation of T cell differentiation. Curr Opin Immunol. 2010;22(5):655–61 [DOI] [PubMed] [Google Scholar]
- 10.Battaglia M, Stabilini A, Migliavacca B, Horejs-Hoeck J, Kaupper T, Roncarolo MG. Rapamycin promotes expansion of functional CD4+CD25+FOXP3+ regulatory T cells of both healthy subjects and type 1 diabetic patients. J Immunol. 2006;177(12): 8338–47 [DOI] [PubMed] [Google Scholar]
- 11.Putnam AL, Brusko TM, Lee MR, Liu W, Szot GL, Ghosh T, et al. Expansion of human regulatory T-cells from patients with type 1 diabetes. Diabetes. 2009;58(3):652–62 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Mucida D, Park Y, Kim G, Turovskaya O, Scott I, Kronenberg M, et al. Reciprocal TH17 and regulatory T cell differentiation mediated by retinoic acid. Science. 2007; 317(5835):256–60 [DOI] [PubMed] [Google Scholar]
- 13.Elias KM, Laurence A, Davidson TS, Stephens G, Kanno Y, Shevach EM, et al. Retinoic acid inhibits Th17 polarization and enhances FoxP3 expression through a Stat-3/Stat-5 independent signaling pathway. Blood. 2008;111(3):1013–20 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Mucida D, Pino-Lagos K, Kim G, Nowak E, Benson MJ, Kronenberg M, et al. Retinoic acid can directly promote TGF-beta-mediated Foxp3(+) Treg cell conversion of naive T cells. Immunity. 2009;30:(4)471–2; author reply 472–3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Wang J, Huizinga TW, Toes RE. De novo generation and enhanced suppression of human CD4+CD25+ regulatory T cells by retinoic acid. J Immunol. 2009;183(6):4119–26 [DOI] [PubMed] [Google Scholar]
- 16.Lu L, Ma J, Li Z, Lan Q, Chen M, Liu Y, et al. All-trans retinoic acid promotes TGF-beta-induced Tregs via histone modification but not DNA demethylation on Foxp3 gene locus. PLoS One. 2011;6(9):e24590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Golovina TN, Mikheeva T, Brusko TM, Blazar BR, Bluestone JA, Riley JL. Retinoic acid and rapamycin differentially affect and synergistically promote the ex vivo expansion of natural human T regulatory cells. PLoS One. 2011;6(1):e15868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Hall JA, Cannons JL, Grainger JR, Dos Santos LM, Hand TW, Naik S, et al. Essential role for retinoic acid in the promotion of CD4(+) T cell effector responses via retinoic acid receptor alpha. Immunity. 2011;34(3): 435–47 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Miyara M, Yoshioka Y, Kitoh A, Shima T, Wing K, Niwa A, et al. Functional delineation and differentiation dynamics of human CD4+ T cells expressing the FoxP3 transcription factor. Immunity. 2009;30(6): 899–911 [DOI] [PubMed] [Google Scholar]
- 20.Strauss L, Czystowska M, Szajnik M, Mandapathil M, Whiteside TL. Differential responses of human regulatory T cells (Treg) and effector T cells to rapamycin. PLoS One. 2009;4(6):e5994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Shevach EM. Mechanisms of foxp3+ T regulatory cell-mediated suppression. Immunity. 2009;30(5):636–45 [DOI] [PubMed] [Google Scholar]
- 22.Koenen HJ, Fasse E, Joosten I. CD27/CFSE-based ex vivo selection of highly suppressive alloantigen-specific human regulatory T cells. J Immunol. 2005;174(12):7573–83 [DOI] [PubMed] [Google Scholar]
- 23.Nadig SN, Wieckiewicz J, Wu DC, Warnecke G, Zhang W, Luo S, et al. In vivo prevention of transplant arteriosclerosis by ex vivo-expanded human regulatory T cells. Nat Med. 2010;16(7):809–13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Baecher-Allan C, Wolf E, Hafler DA. MHC class II expression identifies functionally distinct human regulatory T cells. J Immunol. 2006;176(8):4622–31 [DOI] [PubMed] [Google Scholar]
- 25.Hori S. Developmental plasticity of Foxp3+ regulatory T cells. Curr Opin Immunol. 2010;22(5):575–82 [DOI] [PubMed] [Google Scholar]
- 26.Cosmi L, De Palma R, Santarlasci V, Maggi L, Capone M, Frosali F, et al. Human interleukin 17-producing cells originate from a CD161+CD4+ T cell precursor. J Exp Med. 2008;205(8):1903–16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Maggi L, Santarlasci V, Capone M, Peired A, Frosali F, Crome SQ, et al. CD161 is a marker of all human IL-17-producing T-cell subsets and is induced by RORC. Eur J Immunol. 2010;40(8):2174–81 [DOI] [PubMed] [Google Scholar]
- 28.Tresoldi E, Dell’albani I, Stabilini A, Jofra T, Valle A, Gagliani N, et al. Stability of human rapamycin-expanded CD4+CD25+ T regulatory cells. Haematologica. 2011;96(9): 1357–65 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.King MA, Covassin L, Brehm MA, Racki W, Pearson T, Leif J, et al. Human peripheral blood leucocyte non-obese diabetic-severe combined immunodeficiency interleukin-2 receptor gamma chain gene mouse model of xenogeneic graft-versus-host-like disease and the role of host major histocompatibility complex. Clin Exp Immunol. 2009; 157(1):104–18 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hoffmann P, Eder R, Boeld TJ, Doser K, Piseshka B, Andreesen R, et al. Only the CD45RA+ subpopulation of CD4+CD25high T cells gives rise to homogeneous regulatory T-cell lines upon in vitro expansion. Blood. 2006;108(13):4260–7 [DOI] [PubMed] [Google Scholar]
- 31.Lombardi G, Sagoo P, Scotta C, Fazekasova H, Smyth L, Tsang J, et al. Cell therapy to promote transplantation tolerance: a winning strategy? Immunotherapy. 2011;3(4 Suppl):28–31 [DOI] [PubMed] [Google Scholar]
- 32.Kang SG, Lim HW, Andrisani OM, Broxmeyer HE, Kim CH. Vitamin A metabolites induce gut-homing FoxP3+ regulatory T cells. J Immunol. 2007;179(6): 3724–33 [DOI] [PubMed] [Google Scholar]
- 33.McGrane MM. Vitamin A regulation of gene expression: molecular mechanism of a prototype gene. J Nutr Biochem. 2007;18(8): 497–508 [DOI] [PubMed] [Google Scholar]
- 34.Hirahara K, Liu L, Clark RA, Yamanaka K, Fuhlbrigge RC, Kupper TS. The majority of human peripheral blood CD4+CD25highFoxp3+ regulatory T cells bear functional skin-homing receptors. J Immunol. 2006;177(7):4488–94 [DOI] [PubMed] [Google Scholar]
- 35.Zhang N, Schroppel B, Lal G, Jakubzick C, Mao X, Chen D, et al. Regulatory T cells sequentially migrate from inflamed tissues to draining lymph nodes to suppress the alloimmune response. Immunity. 2009;30 (3):458–69 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Baecher-Allan C, Brown JA, Freeman GJ, Hafler DA. CD4+CD25high regulatory cells in human peripheral blood. J Immunol. 2001;167(3):1245–53 [DOI] [PubMed] [Google Scholar]
- 37.Iellem A, Colantonio L, D’Ambrosio D. Skin-versus gut-skewed homing receptor expression and intrinsic CCR4 expression on human peripheral blood CD4+CD25+ suppressor T cells. Eur J Immunol. 2003;33 (6):1488–96 [DOI] [PubMed] [Google Scholar]
- 38.Lee I, Wang L, Wells AD, Dorf ME, Ozkaynak E, Hancock WW. Recruitment of Foxp3+ T regulatory cells mediating allograft tolerance depends on the CCR4 chemokine receptor. J Exp Med. 2005;201 (7):1037–44 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Booth NJ, McQuaid AJ, Sobande T, Kissane S, Agius E, Jackson SE, et al. Different proliferative potential and migratory characteristics of human CD4+ regulatory T cells that express either CD45RA or CD45RO. J Immunol. 2010;184(8):4317–26 [DOI] [PubMed] [Google Scholar]
- 40.Hoffmann P, Eder R, Kunz-Schughart LA, Andreesen R, Edinger M. Large-scale in vitro expansion of polyclonal human CD4(+)CD25high regulatory T cells. Blood. 2004;104(3):895–903 [DOI] [PubMed] [Google Scholar]
- 41.Cosmi L, Maggi L, Santarlasci V, Capone M, Cardilicchia E, Frosali F, et al. Identification of a novel subset of human circulating memory CD4(+) T cells that produce both IL-17A and IL-4. J Allergy Clin Immunol. 2010;125:(1)222–30 e1–4 [DOI] [PubMed] [Google Scholar]
- 42.Uematsu S, Fujimoto K, Jang MH, Yang BG, Jung YJ, Nishiyama M, et al. Regulation of humoral and cellular gut immunity by lamina propria dendritic cells expressing Toll-like receptor 5. Nat Immunol. 2008;9(7):769–76 [DOI] [PubMed] [Google Scholar]
- 43.Golovina TN, Mikheeva T, Suhoski MM, Aqui NA, Tai VC, Shan X, et al. CD28 costimulation is essential for human T regulatory expansion and function. J Immunol. 2008;181(4):2855–68 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Cao T, Soto A, Zhou W, Wang W, Eck S, Walker M, et al. Ex vivo expanded human CD4+CD25+Foxp3+ regulatory T cells prevent lethal xenogenic graft versus host disease (GVHD). Cell Immunol. 2009;258 (1):65–71 [DOI] [PubMed] [Google Scholar]
- 45.Kleinewietfeld M, Starke M, Di Mitri D, Borsellino G, Battistini L, Rotzschke O, et al. CD49d provides access to “untouched” human Foxp3+ Treg free of contaminating effector cells. Blood. 2009;113(4):827–36 [DOI] [PubMed] [Google Scholar]