

Abstract
Wadi Al-Jufair, a tributary of Wadi Nisah, is one of the important wadis of Najd region (Saudi Arabia) sheltering a rich diversity of higher plants. The study area is extended into approximately 15 km2 encompassing the commonest geomorphological features encountered in desert wadis. The wadi supports several rare plants, including Maerua crassifolia Forssk., a regionally endangered tree, and Acacia oerfota (Forssk.) Schweinf., a rare shrub with restricted distribution. The present study aims to analyze the vegetation of wadi Al-Jufair and propose its designation as an important plant reserve. The vegetation type is fundamentally of chamaephytic nature with some phanerophytes, and distinguished into associations where the dominant perennial species give the permanent character of plant cover in each habitat. Four vegetation groups were identified with the application of TWINSPAN, DCA and CCA programs and named after the characteristic species as follows: Lycium shawii; A. oerfota; Acacia raddiana–Rhazya stricta and Artemisia monosperma. These plant associations demonstrate significant variation in soil texture, moisture, organic matter, pH, EC, and minerals of Wadi Al-Jufair.
Keywords: Plant diversity, Hyperarid wadi, Invasive species, Najd, Plant community, Saudi Arabia, Vegetation, Wadi Al-Jufair
1. Introduction
The wadi ecosystems in Najd Region are among the main plant diversity centers of central Saudi Arabia where biological research has been considerable but in-depth analysis on its floristic components are insubstantial (Vesey-Fitzgerald, 1957; Mandaville, 1990; Chaudhary, 1999). Moreover, the importance of wadi ecosystems for socioeconomic development is becoming increasingly recognized due to its ecological significance, physiographic variation and environmental gradients. The central region of Saudi Arabia that belongs to Saharo-Arabian phytogeographical zone, consists of small mountains, hillocks, plateaus, desert plains depressions “Raudhas” and wadis (Vesey-Fitzgerald, 1957; Shaltout and Mady, 1996; Sharaf El-Din et al., 1998; Alfarhan, 2001; Shaltout et al., 2010).
Several studies were conducted in the past to explore the plants in deserts (De Marco and Dinelli, 1974; Migahid, 1978), which, over the years, helped in strengthening the foundation of the desert studies in Saudi Arabia. Chaudhary (1983a), Mandaville (1986) and Al-Hemaid (1996) have demonstrated the vegetation of the sand dunes of Nafud, Dahna and Rub al-Khali, while Al-Turki (1997), El-Sheikh and Yousef (1981) and Schulz and Whitney (1986) have studied the vegetation and floras of the sabkhas, hillocks and other prominent mountains of the Najd region, such as Tuwaiq, Aja and Salma. Considerable efforts have also been made toward the elucidation of vegetation–environmental relationships in the ecosystems “raudhas” or depressions (Shaltout and Mady, 1996; Sharaf El-Din et al., 1998; Alfarhan, 2001) and plant communities of wadis, such as Wadi Al-Ammaria (Al-Yemeni, 2001) and Wadi Hanifa (Taia and El-Ghanem, 2001; EL Ghenem, 2006).
Wild plants in the Najd region are very much associated with drought and aridity. Out of 2243 species of higher plants from the entire country (Chaudhary, 1983a,b, 1999, 2000, 2001; Collenette, 1999), approximately 600 species are reported from the Central Region (Chaudhary, 1999, 2000, 2001; Zoghet and Alsheikh, 1999). Among these, many are either xerophytes or morphologically and physiologically adapted to cope with the harsh environment prevailing in the central region. Wild plants in the central region, though mostly annuals, have their own characteristics in combating desertification, improving the local climate, conserving soils, fixing sand dunes, preventing erosion and flooding (Zoghet and Alsheikh, 1999; Yousef and El-Sheikh, 1981a and Yousef and El-Sheikh, 1981b). Vegetation of wadis in general is not constant. It varies from year to year, depending upon the moisture (Siddiqui and Al-Harbi, 1995). Al-Farraj et al. (1997) conducted vegetation studies in some “Raudhas” to verify the abundance, frequency and density of each species, while Al-Yemeni and Al-Farraj (1995) reported the characteristics of seed bank and their relationship to the desert vegetation, a fundamental part of understanding the ecological and physiological process of several plants.
Generally, the soil of the central region can be considered as “Aridisols”, with no accumulation of clays or organic matter (Al-Nafie, 2004). Whereas the deposits in wadi basin are deep and fine-textured with a firm, flat vegetation-covered mud surface, supported by occasional rocky, sandy or phytogenic mounds (Batanouny, 1987). Since wadi Al-Jufair is vegetationally and floristically one of the richest wadis of the central region, the main objective of the present study is to analyze the vegetation of Wadi Al-Jufair and assess the role of edaphic factors and human impact that influences the vegetation and thereby proposes its designation as an important plant diversity center.
2. Study area
Wadi Al-Jufair (23°50′N, 46°14′E and 24°06′N, 46°19′E), a tributary of Wadi Nisah, is located about 120 km southwest of Riyadh City in Central Saudi Arabia (Fig. 1a). The study area extends 20 km2 and protects by Tuwayq mountain range which has an altitude ranging from 600 to 1000 m a.s.l. Wadi Nisah and Tuwayq mountains are the main features of the central physiographic province of a large central plateau, “Najd”, which is divided into “Higher Najd” in the western and “Lower Najd” in the eastern sides. It is characterized by the dissection of its landscape into an extensive system of large wadis which flow eastward, from the higher mountains in the west to the plains of Najd, responding to the general slope of the land. These wadis are not continuous, and at times covered and buried by the sand dunes of Dahna desert. Seasonal springs originate in these wadis during rainy days and often create shallow pools along the banks and depressions. Remnants of dense vegetation can be found along the stretch, reflecting a wetter climate of the past (Chapman, 1978; Al-Nafie, 2008). The dominant topography of the central province is made up of a nearly parallel sequence of several prominent crescent-shaped north–south escarpments, “Tuwayq Escarpment”, which is mainly of hard marine limestone capped with upper Jurassic limestone, extending for about 1200 km, with elevations up to 850 m a.s.l. and 240 m above the nearby plains (Al-Nafie, 2008).
Figure 1a.
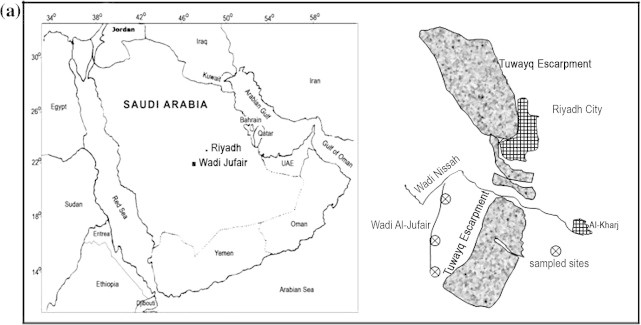
Location map of the study area and diagram of Wadi Al-Jufair.
As the study area is located in the middle of three separate habitats (sand dunes, rocky hills and depression), the vegetation of Wadi Al-Jufair is influenced by the topography and soil type of these habitats. Wadi Al-Jufair is somewhat an enclosed habitat protected by Tuwayq Mountains and dissected into an extensive system of several small and big wadis. Despite harsh environment, Wadi Al-Jufair ecosystem is diverse in habitats and accordingly the vegetation is different from one habitat to another. During winter, the vegetation of the main wadi and its tributaries is represented by different plant communities, each comprised of trees, shrubs, sub-shrubs and seasonal vegetation represented by mesophytic herbs and grasses.
The study area is physiographically distinguished into plateau, slope and wadi bed. The major feature of the plateau is the flat rocky surface; its vegetation is restricted to notches and shallow drainage runnels where variable amounts of soil accumulate. The upper positions of slopes are steep and completely devoid of soil cover; and, therefore, support typical cliff vegetation. The middle slopes are less steep and covered by a shallow soil mixed with fragments of rocks of different sizes and support vegetation dominated by shrubby species of chasmophytic nature and grasses. The lower parts of slopes are gentle, where deep soil is accumulated by the run-off water and support dense vegetation. The fine soil has little chance to settle down due to the high velocity of flash floods during the rainy season. However, soil often filled up between large boulders where it supports the establishment of sparse vegetation. The wadi basin, which is opening into the main Wadi Nisah in the north, is characterized by sand dunes.
Meteorological data of the central region are characterized by mean air temperature ranging from 10 °C in January to 42 °C in July and an annual rainfall from 12 to 36 mm (Fig 2). The mean relative humidity ranges between 10% and 50%, while the mean evaporation value is 10.35 mm/day (Al-Nafie, 2008).
Figure 2.
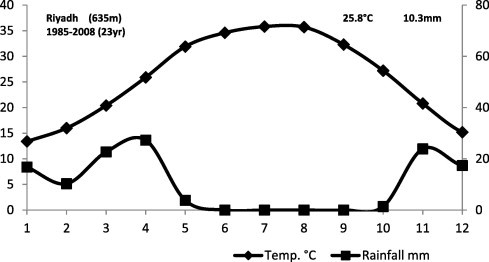
Climate diagram of Riyadh region.
3. Material and methods
3.1. Sampled stands
A total of 22 sites were selected. The stands were distributed along transect on the wadi that covered various landforms (6 on the wadi plateau, 6 on the wadi slope and 10 on the wadi bed and delta). The sampling process was carried out during spring season when most species were expected to be growing (Figs. 1a and b). The vegetation parameters included listing of all species, life forms and chorotypes. Species nomenclature followed Chaudhary (1999, 2000, 2001) and Collenette (1999). Plant cover was estimated by using the line intercept method (Canfield, 1941).
Figure 1b.
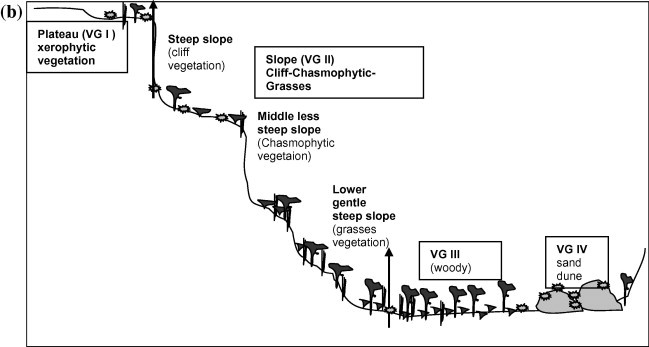
Schematic representation of the spatial hierarchal organization of the vegetation along the profile transect at Wadi Al-jufair. The vegetation groups are named as follows: VG I. Lycium shawii; VG II. Acacia oerfota; VG III. Acacia raddiana–Rhazya stricta and VG IV Artemisia monosperma.
3.2. Soil analysis
Three soil samples, down to 50 cm depth, were collected from each stand and mixed as one composite sample for each site. Soil texture was determined by hydrometer method (Allen et al., 1974). Total organic matter was determined based on loss-on-ignition at 450 °C. Soil water extract was prepared (1:5), by dissolving 100 g air dried soil in 500 ml distilled water for estimation of pH and electrical conductivity (EC) as mS cm−1. Soil nutrient elements (Ca, K, Na, Mg, Fe, N and P) were determined using spectrophotometer (model ICP MSEOS 6000 Series). All procedures are outlined by Allen et al. (1974).
3.3. Data analysis
The cover estimates of 77 plant species recorded in 22 stands were subjected to multivariate analysis; using TWINSPAN, DCA and CCA (Hill, 1979a,b; Ter Braak and Smilauer, 2002). Species richness (α-diversity) of the vegetation cluster was calculated as the average number of species per stand. Shannon–Wiener index for the relative evenness, and Simpson index for the relative concentration of dominance were calculated for each stand on the basis of the relative cover pi of the ith species (Pielou, 1975; Magurran, 1988). Relationships between the ordination axes on one hand, and community and soil variables on the other hand were tested using Pearson’s simple linear correlation coefficient (r). The variation in the species diversity, stand traits and soil variables in relation to plant community were assessed using one way analysis of variance (SAS, 1989–1996).
4. Results
4.1. Floristic diversity
One hundred and fifty-seven species, belonging to 133 genera in 40 families were recorded from various stands and adjoining areas, of which the most represented families are Poaceae and Asteraceae. Therophytes constituted 81 species (51%) of the total species) followed by Chamaephytes of 33 species (21%) and perennial herbs 31 species (18%) (Fig. 3a). Regarding the chorotype (Fig. 3b), the Saharo-Arabian region (auct, Zohary, 1973) has the highest share of species (41%), followed by the bi-regional and pleuri-regional elements that belong to the Sahelien-Somali Masai (10%), Saharo-Arabian-Somali Masai (8%), Saharo-Arabian-Irano-Turanian (8%), and Mediterranean-Irano-Turanian (8%).
Figure 3.
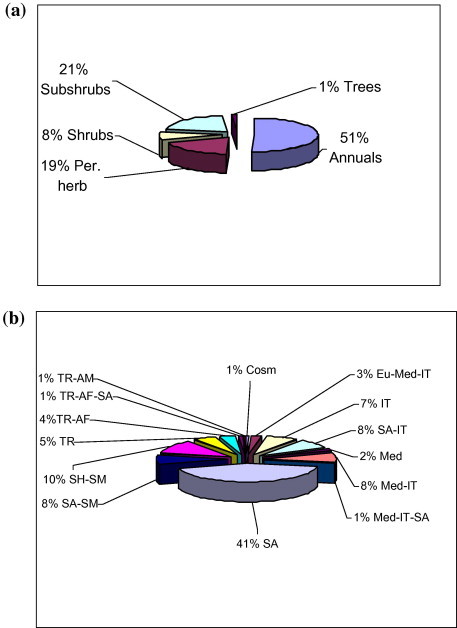
Life form (a) and Chorotype (b) relative spectra of the recorded species in Wadi Al-Jufair. (IT = Irano-Turanian; SA-IT = Saharo-Arabian-Irano-Turanian; SH-SM = Sahelian-Somali-Masai; TR = Tropical; SA = Saharo-Arabian; SA-SM = Saharo-Arabian-Somali-Masai; TR AF = Tropical African; TR AM = Torpical American; Med-IT = Mediterranean-Irano-Turanian; EU-Med-IT = Euro-Siberian-Mediterranean-Irano-Turanian; Med = Mediterranean; Med-IT-SA = Mediterranean-Irano-Turanian-Saharo-Arabian; Cosm = Cosmopolitan; TR AF-SA = Tropical Africa-Saharo-Arabian.
4.2. Multivariate analysis
TWINSPAN dendrogram divided the data set (22 stands × 72 species) into eight vegetation subgroups at level 5 and four vegetation groups (i.e. plant communities) at level 2. These four major plant communities were characterized and named after the dominant and subdominant species as follows: (I) Lycium shawii; (II) Acacia oerfota; (III) Acacia raddiana–Rhazya stricta and (IV) Artemisia monosperma. The application of DCA and CCA confirmed the separation between these communities and indicated relationships between environmental gradients and topographic aspects of wadi Al-Jufair (Table 1, Fig. 4a and b).
Table 1.
Synoptic table of species composition of the four vegetation groups (I–VI) identified after the application of TWINSPAN to the vegetation data of the 22 stands of Wadi Al-Jufair region. The cover levels are coded as follows: 1, ⩽10%; 2, 10–20%; 3, 20–30; 4, 30–40%; 5, ⩾40%. The vegetation groups are named as follows: I. Lycium shawii; II. Acacia oerfota; III. Acacia raddiana–Rhazya stricta and IV Artemisia monosperma. The life forms are: Th = Therophyte; Ch = Chamaephyte; Ph = Phanerophyte; He = Hemicryptophyte; Cr = Cryptophyte. The chorotypes are: IT = Irano-Turanian; SA-IT = Saharo-Arabian-Irano-Turanian; SH-SM = Sahelian-Somali-Masai; TR = Tropical; SA = Saharo-Arabian; SA-SM = Saharo-Arabian-Somali-Masai; TR AF = Tropical African; TR AM = Torpical American; Med-IT = Mediterranean-Irano-Turanian; EU-Med-IT = Euro-Siberian-Mediterranean-Irano-Turanian; Med = Mediterranean; Med-IT-SA = Mediterranean-Irano-Turanian-Saharo-Arabian; Cosm = Cosmopolitan; TR AF-SA = Tropical Africa-Saharo-Arabian.
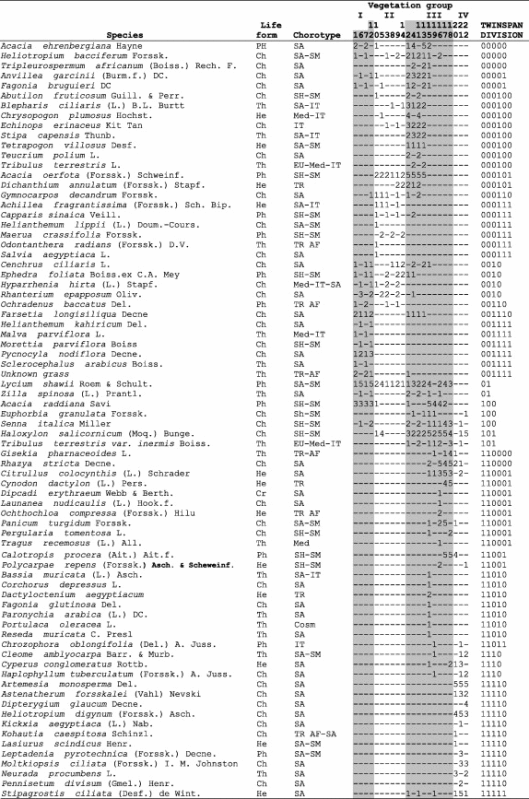 |
Figure 4.
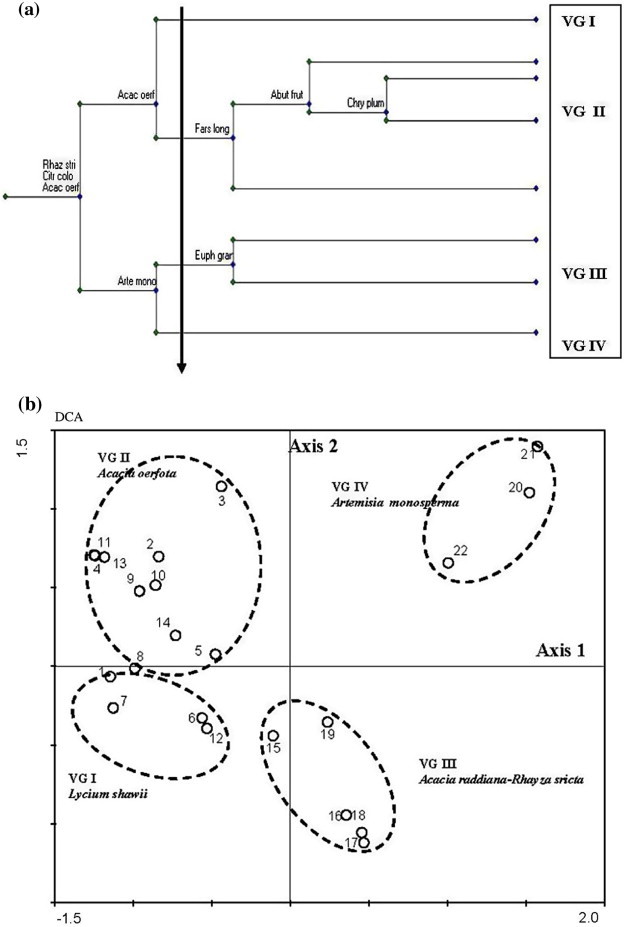
Relationship between the four plant communities after the application of TWINSPAN (a) and DCA (b).
CCA ordination was used to verify the correlation analysis between the dominant environmental factors and CCA axes (Fig. 5a and b and Table 2). Correlation analysis indicated that the separation of the species along the first axis is strongly affected positively by EC, sand, nitrogen content and species dominance (r = 0.310–0.851) and negatively by organic matter, clay, silt, Ca, Fe, K, Mg and P contents (r = −0.337 to −0.896). On the other hand, species richness (0.369) and clay content (0.310) are correlated positively with the second axis and negatively with pH, Fe and plant cover (r = −0.306 to −0.554). Therefore, the L. shawii (VG I) that occupies the wadi plateau and A. oerfota (VG II) that occupies the wadi slope were separated on the left hand of axis 1 from the A. raddiana–R. stricta (VG III) and A. monosperma (VG IV) which inhabited the wadi bed and on the right hand of axis 2 (Fig. 5a).
Figure 5.
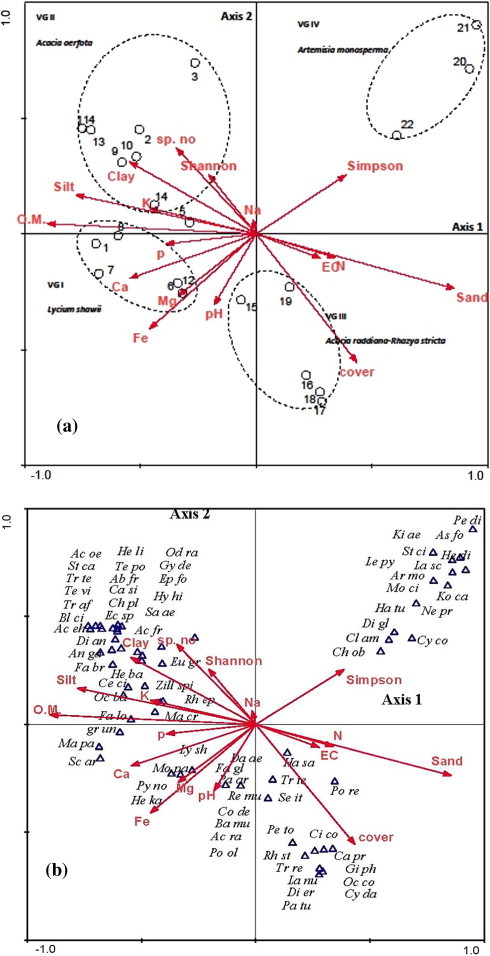
CCA biplot with environmental variables (arrows), the stands (a) and the abundant species represented by the first 4 letters of genus and species name (b). (For complete names of species, see Table 1).
Table 2.
Inter-set correlations of environmental variables with CCA axes. Significant values in bold.
| N | Name | AXIS-1 | AXIS-2 |
|---|---|---|---|
| 1 | pH | −0.18 | −0.31⁎ |
| 2 | EC (mS/cm) | 0.31⁎ | −0.11 |
| Bulk soil (%) | |||
| 3 | Organic matter | −0.90∗∗∗ | 0.04 |
| 4 | Sand | 0.85∗∗∗ | −0.24∗ |
| 5 | Clay | −0.054∗ | 0.31∗ |
| 6 | Silt | −0.78∗∗∗ | 0.17 |
| Minerals (ppm) | |||
| 7 | Ca | −0.54∗∗ | −0.19 |
| 8 | Fe | −0.46∗∗ | −0.41∗∗ |
| 9 | K | −0.50∗∗ | 0.11 |
| 10 | Mg | −0.34∗ | −0.27∗ |
| 11 | Na | −0.02 | 0.06 |
| 12 | P | −0.39∗∗ | −0.04 |
| 13 | N | 0.39∗ | −0.12 |
| Diversity indices | |||
| 14 | Species richness spp. stand−1 | −0.34∗ | 0.37∗ |
| 15 | Species cover m–100 m−1 | 0.43∗∗ | −0.56∗∗∗ |
| 16 | Conc. of dominance (C) | 0.39∗∗ | 0.25∗ |
| 17 | Relative evenness (Ĥ) | −0.21 | 0.26∗ |
p ⩽ 0.05.
p ⩽ 0.01.
p ⩽ 0.001.
Xerophytes occupied the fissures of flat stony habitat of L. shawii community (e.g. Bassia muricata, Corchorus depressus, Dactyloctenium aegyptium, Fagonia glutinosa, Helianthemum kahiricum, Paronychia arabica, Portulaca oleracea, Pycnocycla nodiflora, Reseda muricata and Sclerocephalus arabicus) on the lower negative part of axis 1 are correlated with P, Ca, Mg, pH and Fe content. The cliff, chasmophytic and grasses species of the A. oerfota community (e.g. Acacia ehrenbergiana, Anvillea garcinii, Capparis sinaica, Dichanthium annulatum, Gymnocarpos decandrum, Hyparrhenia hirta, Lasiurus scindicus, Ochradenus baccatus, Rhanterium epapposum, Stipa capensis, Tripleurospermum auriculatum and Zilla spinosa) on the upper negative part of axis 1 are correlated with organic matter, silt, clay, K, species number and relative evenness. On the other hand, the grasses, shrubs and sub-shrubs inhabit the wadi bed of the A. raddiana–R. stricta community, including some of the invasive species (e.g. Calotropis procera). Other species in this community (e.g. Citrullus colocynthis, Cynodon dactylon, Halocnemum strobilaceum, Ochthochloa compressa, Panicum turgidum, Pergularia tomentosa, Polycarpaea repens and Senna italica) on the lower positive part of the axis 1 are correlated with EC, nitrogen, sand and species cover. The psammophytic community inhabits the sand dunes of A. monosperma (e.g. Cyperus conglomeratus, Haloxylon salicornicum, Haplophyllum tuberculatum, Lasiurus scindicus, Leptadenia pyrotechnica, Moltkiopsis ciliata, Neurada procumbens, and Pennisetum divisum) on the upper positive part of axis 1 are correlated with species concentration of dominance. These combinations are typical of grass communities inhabiting the wadi bed and sand dunes (Fig. 5b).
The species richness was positively correlated with organic matter (0.405) and clay (0.673), and negatively with total cover, EC and sand (−0.272, −0.322 and −0.389, respectively) (Table 3). The species cover was positively correlated with pH (0.352) and sand (0.592), and negatively with organic matter, clay, silt and K contents (−0.454, −0.422, −0.526 and −0.410, respectively). The species concentration of dominance was positively correlated with Mg (0.394) and Na (0.548), and negatively with the species relative evenness (−0.522) and clay (−0.323). The species evenness was positively correlated with pH, EC and clay (0.468, 0.311 and 0.447, respectively) and negatively with Mg, Na and N (−0.559, −0.465 and −0.355, respectively).
Table 3.
Correlation between species diversity and edaphic variables. Significant values in bold.
| Variable | Species richness | Species cover m–100 m−1 | Conc. of dominance (C) | Relative evenness (Ĥ) |
|---|---|---|---|---|
| Diversity indices | ||||
| Species richness spp/stand | 1.00 | |||
| Species. cover m/100 m | −0.27⁎ | 1.00 | ||
| Conc. of dominance (C) | −0.02 | −0.02 | 1.00 | |
| Relative evenness (Ĥ) | 0.40⁎⁎ | −0.03 | −0.52⁎⁎ | 1.00 |
| Soil | ||||
| pH | 0.15 | 0.35⁎ | −0.27 | 0.47⁎⁎ |
| EC (mS/cm) | −0.32⁎ | −0.01 | −0.14 | 0.31⁎ |
| Bulk soil (%) | ||||
| Org. matter | 0.41⁎⁎ | −0.45⁎⁎ | −0.22 | 0.16 |
| Sand | −0.39⁎⁎ | 0.59⁎⁎⁎ | 0.04 | 0.04 |
| Clay | 0.67⁎⁎⁎ | −0.42⁎⁎ | −0.32⁎ | 0.45⁎⁎ |
| Silt | 0.23 | −0.52⁎⁎ | 0.05 | −0.18 |
| Mineral soil (ppm) | ||||
| Ca | 0.08 | 0.14 | −0.07 | 0.13 |
| Fe | 0.28⁎ | 0.07 | 0.08 | −0.13 |
| K | 0.10 | −0.41⁎⁎ | −0.09 | 0.13 |
| Mg | −0.20 | −0.25 | 0.39⁎⁎ | −0.56⁎⁎⁎ |
| Na | 0.18 | −0.30⁎ | 0.55⁎⁎ | −0.47⁎⁎ |
| P | 0.15 | −0.22 | 0.29⁎ | −0.27 |
| N | −0.10 | 0.07 | 0.17 | −0.35⁎ |
p ⩽ 0.05.
p ⩽ 0.01.
p ⩽ 0.001.
4.3. Plant community–soil relationship
The A. raddiana–R. stricta community (VG III) demonstrated the highest levels of species richness (14.4) and species cover (110.1 m–00 m−1). The A. oerfota (VG II) attained the highest relative evenness (0.92) and the lowest species cover (43.72 m–100 m−1) and dominance (0.13). A. monosperma (VG IV) showed the highest concentration of species dominance (0.48), while L. shawii (VG I) attained the lowest levels of species richness (12.75) and relative species evenness (0.69). The habitats of L. shawii community (VG I) are characterized by the highest contents of O.M., silt, Ca, Fe, Mg, P and the lowest of pH and sand. On the other hand, the A. oerfota (VG II) characterized the sites of the highest pH, EC, clay and K. A. monosperma community (VG IV) occupied sites with the highest values of sand and the lowest of the most soil variables (Table 4).
Table 4.
The mean ± standard deviation of soil variable and diversity indices. Significant values in bold.
| Variable | VGI | VGII | VGIII | VGIV | Total mean | F-value |
|---|---|---|---|---|---|---|
| Diversity indices | ||||||
| Species richness | 12.75 ± 9.8 | 13.70 ± 44.8 | 14.40 ± 43.2 | 13.66 ± 21.1 | 13.68 | 0.07 |
| Species cover | 54.27 ± 76.5 | 43.72 ± 81.3 | 110.1 ± 58.2 | 97.70 ± 6.0 | 68.08 | 3.29⁎ |
| Conc. of dominance (C) | 0.35 ± 78.6 | 0.13 ± 65.1 | 0.25 ± 30.9 | 0.48 ± 64.2 | 0.24 | 4.15⁎ |
| Relative evenness (Ĥ) | 0.69 ± 36. | 0.92 ± 17.9 | 0.75 ± 12.7 | 0.76 ± 20.9 | 0.82 | 2.24 |
| Soil | ||||||
| pH | 7.9 ± 0.0 | 8.12 ± 1.8 | 8.06 ± 5.3 | 7.93 ± 6.9 | 8.04 | 0.70 |
| EC (mS/cm) | 0.44 ± 65.0 | 0.52 ± 66.6 | 0.34 ± 65.0 | 0.43 ± 62.7 | 0.46 | 0.39 |
| Bulk soil (%) | ||||||
| O.M. | 1.66 ± 16.6 | 1.57 ± 23.9 | 0.76 ± 58.6 | 0.32 ± 53.7 | 1.23 | 13.9⁎⁎⁎ |
| Sand | 60.3 ± 1.64 | 65.2 ± 7.7 | 83.00 ± 11.8 | 86.69 ± 1.2 | 71.30 | 21.7⁎⁎⁎ |
| Clay | 12.16 ± 18.9 | 13.66 ± 45.8 | 10.56 ± 25.5 | 9.49 ± 6.0 | 12.11 | 0.85 |
| Silt | 27.54 ± 4.9 | 21.12 ± 40.9 | 6.43 ± 3.3 | 3.81 ± 39.3 | 16.59 | 11.3⁎⁎⁎ |
| Mineral soil (ppm) | ||||||
| Ca | 57.93 ± 23.8 | 31.73 ± 51.6 | 20.87 ± 84.4 | 8.26 ± 13.9 | 30.83 | 6.62⁎⁎⁎ |
| Fe | 2.24 ± 1.4 | 1.40 ± 31.6 | 1.53 ± 31.1 | 1.05 ± ± 22.1 | 1.53 | 6.2⁎⁎⁎ |
| K | 0.36 ± 11.9 | 0.41 ± 1.2 | 0.14 ± 35.6 | 0.11 ± 15.7 | 0.30 | 0.93 |
| Mg | 1.46 ± 2.7 | 0.54 ± 56.4 | 0.45 ± 26.4 | 0.21 ± 90.8 | 0.64 | 21.5⁎⁎⁎ |
| Na | 0.01 ± 7.9 | 0.01 ± 23.2 | 0.009 ± 15.1 | 0.01 ± 34.3 | 0.0098 | 3.91⁎ |
| P | 3.52 ± 4.1 | 0.64 ± 16.5 | 0.51 ± 29.1 | 0.48 ± 36.9 | 1.11 | 16.36⁎⁎⁎ |
| N | 0.01 ± 7.0 | 0.01 ± 3.2 | 0.008 ± 15.1 | 0.01±34.3 | 0.008 | 3.91⁎ |
p ⩽ 0.05.
p ⩽ 0.001.
5. Discussion and conclusion
The life form distribution of plants growing in arid regions is closely related with topography and landform (Kassas and Girgis, 1964; Zohary, 1973; Orshan, 1986; Shaltout et al., 2010). The Najd plateau in the rain-shadow of the Tuwayq Escarpment in the central Saudi Arabia has different wadis (Mandaville, 1986). Al-Jufair is a mature wadi characterized by its wide, deep valley-fill deposits, and well defined channels cutting older rocky limestone formations. The wadi ecosystem is divided into a number of habitats discernible on the ground of the soil thickness and plant cover. The vegetation is featured into associations where the dominant perennial species give the permanent character of plant cover in each habitat. This may be attributed to the rather scanty rainfall which is not adequate for the appearance of many annuals. On the other hand, the rainy season provides better chance for the appearance of a considerable number of annuals, which give a characteristic physiognomy to their vegetation (Shaltout and Mady, 1996; Schulz and Whitney, 1986; Hosni and Hegazy, 1996; Shaltout et al., 2010).
Floristic analysis indicated that the synanthropic species (e.g. Bassia eriophora, C. dactylon, Prosopis juliflora, Salsola imbricata and Tamarix nilotica) are rare or completely absent in Wadi Al-Jufair, indicating low human impact. However, these species have been recorded as common in a few other wadis and depressions (e.g. Raudhas) in the Najd region (Shaltout and Mady, 1996; Al-Farraj et al., 1997; Alfarhan, 2001; Taia and El-Ghanem, 2001; EL Ghenem, 2006; El-Ghanem et al., 2010). The presence of A. oerfota, a rare shrub with restricted distribution and Maerua crassifolia, an endangered tree (Mandaville, 1986) in the study area, can be considered as a positive sign that Wadi Al-Jufair is one of the few areas in the Najd region with less human impact.
The life form spectrum reflects a typical desert flora, the majority of species being therophytes and Chamaephytes (about 72%). These results agree with the spectra of vegetation in desert habitats in other parts of Saudi Arabia (e.g. El-Demerdash et al., 1995; Collenette, 1999; Chaudhary, 1999, 2000, 2001; Al-Turki and Al-Qlayan, 2003; Fahmy and Hassan, 2005; El-Ghanem et al., 2010). Life forms of desert plants are also closely related with topography (Kassas and Girgis, 1964; Zohary, 1973; Migahid, 1978; Orshan, 1986; Hosni and Hegazy, 1996; Hegazy et al., 1998; Shaltout et al., 2010). It may also be stated that the Saharo Arabian species which are restricted in their distribution to the central strip of Saudi Arabia are more abundant in habitats of more favorable micro-environmental conditions and those providing better protection (Zohary, 1973; Ghazanfar and Fisher, 1998; Hegazy et al., 1998; El-Ghanem et al., 2010). Besides high percentage of Saharo Arabian species in the study area, there are several other chorotypes attaining considerable values. This is due to the fact that the central region contains most of the rocky habitat types of the Peninsula and covers a wide range of bioclimatic zones. The central region falls within the transition zone from the Somalia-Masai regional center of endemism at low and middle altitudes to the Afromontane archipelago-like center of endemism at high altitudes, and above the tree line even taxa of the Mediterranean region and Irano-Turanian phytochorion are present (Zohary, 1973; Mandaville, 1990; White and Leonard, 1991; Hegazy et al., 1998; Ghazanfar and Fisher, 1998; Alfarhan, 1999).
Among the four vegetation groups in Wadi Al-Jufair ecosystem, vegetation group III, characterized by A. raddiana–R. stricta has clear separation with group IV dominated by A. monosperma. On the other hand, the groups I and II are less separated because they are characterized by mixed communities of shrubs, chasmophytes and grasses. In Saudi Arabia, Shaltout and Mady (1996), Al-Yemeni and Zayed (1999), Al-Yemeni (2001), Al-Wadie (2002) and EL Ghenem (2006) recognized several plant associations, some of which are comparable to those of the present study (e.g. A. raddiana–R. stricta which is comparable to that identified in neighboring countries: Batanouny, 1987; El-Bana and Al-Mathnani, 2009; Shaltout et al., 2010). The sand dune group A. monosperma has analogous with association recognized by Al-Yemeni and Zayed (1999) at Al-Thumamah sandy habitats. Communities in stony plateau and rocky outcrop slope in Wadi Talha at Asir, as recognized by Al-Wadie (2002), are, however, less comparable in Wadi Al-Jufair, which may be due to the variations in climate and topography.
L. shawii community inhabits the flat stony wadi plateau which consists of notches and shallow drainage runnels. A. oerfota community of cliff, chasmophytic shrubby and grassy species inhabits the outcrops of rocky slopes. Then comes the community of A. raddiana–R. stricta composed of dense woody and sparse short lived perennial species inhabits the main wide channel of the wadi bed. On the other hand, the wadi delta, the sand dunes inhabited by adapted psammophytic A. monosperma community. The most diverse groups II and III, inhabiting the habitats such as wadi slope outcrop, bed, etc. and characterized by A. oerfota and A. raddiana–R. stricta could have been related to high soil pH, high clay content in its soil and/or heterogeneity of substrate in a shallow soil mixed with fragments and a deep valley-fill soil deposits (Fig. 1b). The soil, accumulated by run-off water, supported dense woody and grasses vegetation with high cover during rainy season. Similar conclusions were made by Chaudhary (1983a, b), Siddiqui and Al-Harbi (1995), El-Demerdash et al. (1995), Shaltout and Mady (1996), Al-Yemeni (2001), Al-Wadie (2002), Abbadi and El-Sheikh (2002) and Springuel et al. (2006). A. monosperma group (IV), inhabiting the sand dunes at wadi delta, had low species diversity. This could be due to the increase in aridity at its loose and unstable soil surface, high salinity due to excessive evaporation at its surface and high content of sand (mostly red sand derived from bed rock) with poor fertility. This adverse habitat supported a few adapted psammophytic phytogenic populations of sand binding plants with highest concentration of their dominance and plays a major role in decreasing species diversity of central Saudi Arabia as a whole (Chaudhary, 1983a; Al-Hemaid, 1996; Shaltout and Mady, 1996; Al-Yemeni, 2000; Al-Wadie, 2002; Shaltout et al., 2010).
Correlation analysis in the present study indicates that the species diversity (richness and evenness) is positively correlated with increasing organic matter, clay pH and EC. These factors may reflect the degree of wadi bed maturation in the study area (Kassas and Imam, 1954). On the other hand, species diversity decreased with increasing Mg, Na, N, EC, sand and species cover (which correlates positively with pH and sand). In such cases, most of the total cover is accounted by one or two species (e.g. the community of A. monosperma) that can apparently make the best use of available resources as a result of their high competitive capacities under environmental stress. Similar correlations were reported by El-Demerdash et al. (1995), Abbadi and El-Sheikh (2002), El-Sheikh et al. (2006) and El-Sheikh et al. (2010).
Acknowledgements
This project was supported by King Abdulaziz City for Science and Technology, Riyadh, Saudi Arabia (Project grant number 10-ENV1295-02) through the National Plan for Sciences and Technology (NPST), King Saud University, Riyadh. We also extend our appreciation to Prof. A.K. Hegazy, Department of Botany & Microbiology, College of Science, King Saud University, for his valuable comments.
References
- Abbadi G.A., El-Sheikh M.A. Vegetation analysis of Failaka Island (Kuwait) Journal of Arid Environments. 2002;50:153–165. [Google Scholar]
- Alfarhan A. A floristic account on Raudhat Khuraim, Central Province Saudi Arabia. Saudi Journal of Biological Sciences. 2001;8(1):80–103. [Google Scholar]
- Alfarhan A.H. A phytogeographical analysis of the floristic elements in Saudi Arabia. Pakistan Journal of Biological Sciences. 1999;2(3):702–711. [Google Scholar]
- Al-Farraj M.M., Al-Farhan A., Al-Yemeni M. Ecological studies on rawdhat system in Saudi Arabia I. Rawdhat Khorim. Pakistan Journal of Botany. 1997;29(1):75–88. [Google Scholar]
- Al-Hemaid F.M. Vegetation and distribution of the sand seas in Saudi Arabia. Geobios. 1996;23:2–15. [Google Scholar]
- Allen S.E., Grimshaw H.M., Parkinson J.H., Quarmby C. Blackwell Scientific Publication; Oxford: 1974. Chemical Analysis of Ecological Materials. [Google Scholar]
- Al-Nafie A.H. King Fahd National Library; El-Riyadh, Saudi Arabia: 2004. Botanical Geography of Saudi Arabia. [Google Scholar]
- Al-Nafie A.H. Phytogeography of Saudi Arabia. Saudi Journal of Biological Sciences. 2008;15(1):159–176. [Google Scholar]
- Al-Turki T.A. A preliminary checklist of the flora of Qassim, Saudi Arabia. Feddes Repertorium. 1997;108(3–4):259–280. [Google Scholar]
- Al-Turki T.A., Al-Qlayan H.A. Contribution to the flora of Saudi Arabia: Hail region. Saudi Journal of Biological Sciences. 2003;10:190–222. [Google Scholar]
- Al-Wadie H. Floristic composition and vegetation of Wadi Talha, Aseer mountains, south west Saudi Arabia. Journal of Biological Sciences. 2002;2(5):285–288. [Google Scholar]
- Al-Yemeni M., Al-Farraj M.M. The seed bank of desert soil in Central Saudi Arabia. Pakistan Journal of Botany. 1995;27:309–319. [Google Scholar]
- Al-Yemeni M.N. Ecological studies on Sand dunes vegetation in Al-Kharj region, Saudi Arabia. Saudi Journal of Biological Sciences. 2000;7(1):64–87. [Google Scholar]
- Al-Yemeni M.N. Ecology of some plant communities in Wadi Al-Ammaria, Riyadh, Saudi Arabia. Saudi Journal of Biological Sciences. 2001;8(2):145–165. [Google Scholar]
- Al-Yemeni M.N., Zayed K.M. Ecology of some plant communities along Riyadh-Al-Thumamah Road, Saudi Arabia. Saudi Journal of Biological Sciences. 1999;6(1):9–26. [Google Scholar]
- Batanouny K.H. Current knowledge of plant ecology in the Arab Gulf countries. Catena. 1987;14:291–316. [Google Scholar]
- Canfield R. Application of the line intercept method in sampling range vegetation. Journal of Forestry. 1941;39:288–393. [Google Scholar]
- Chapman R.W. Geomorphology of Arabian Peninsula. In: Sayari A., Zotl J., editors. Quaternary Period in Saudi Arabia. Springer-Verlag; New York: 1978. pp. 19–29. [Google Scholar]
- Chaudhary S.A. Vegetation of Great Nafud. Journal of Saudi Arabian Natural History Society. 1983;2:32–33. [Google Scholar]
- Chaudhary S.A. Ministry of Agriculture and Water; Riyadh: 1983. Acacia and Other Genera of Mimosoideae in Saudi Arabia. [Google Scholar]
- Chaudhary S.A. vol. I. Ministry of Agriculture and Water; Riyadh: 1999. (Flora of the Kingdom of Saudi Arabia). [Google Scholar]
- Chaudhary, S.A., 2000. Flora of the Kingdom of Saudi Arabia, vol II, parts 1–3. Ministry of Agriculture and Water, Riyadh.
- Chaudhary S.A. vol. III. Ministry of Agriculture and Water; Riyadh: 2001. (Flora of the Kingdom of Saudi Arabia). [Google Scholar]
- Collenette I.S. National Commission for Wildlife Conservation and Development; Riyadh: 1999. Wildflowers of Saudi Arabia. [Google Scholar]
- De Marco G., Dinelli A. First contribution to the floristic knowledge of Saudi Arabia. Annali Di Botanica. 1974;36:209–236. [Google Scholar]
- EL Ghenem W.M. Ecological study at wadi Al-Ammaria in El-Riyadh city-Saudi Arabia. Bulletin of Pure and Applied Sciences, Section B. 2006;25(1):11–19. [Google Scholar]
- El-Bana M.I., Al-Mathnani A. Vegetation–soil relationships in the Wadi Al-Hayat area of the Libyan Sahara. Australian Journal of Basic and Applied Sciences. 2009;3:740–747. [Google Scholar]
- El-Demerdash M.A., Hegazy A.K., Zilay M.A. Vegetation–soil relationships in Tihamah coastal plains of Jazan region, Saudi Arabia. Journal of Arid Environments. 1995;30:161–174. [Google Scholar]
- El-Ghanem W.A., Hassan L.M., Galal T.M., Badr A. Floristic composition and vegetation analysis in Hail region north of central Saudi Arabia. Saudi Journal of Biological Sciences. 2010;17:119–128. doi: 10.1016/j.sjbs.2010.02.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- El-Sheikh A.M., Yousef M.M. Halophytic and xerophytic vegetation near Al-Kharj springs. Journal of College of Science, Riyadh University. 1981;12:5–21. [Google Scholar]
- El-Sheikh M.A., El-Ghareeb R.M., Testi A. Diversity of plant communities in coastal salt marshes habitat in Kuwait. Rendiconti Fisiche Accademia Lincei. 2006;17:311–331. [Google Scholar]
- El-Sheikh M.A., Abbadi G.A., Bianco P. Vegetation ecology of phytogenic hillocks (nabkhas) in coastal habitats of Jal Az-Zor National Park, Kuwait. Flora. 2010;205:832–840. [Google Scholar]
- Fahmy A.G., Hassan L.M. Plant diversity of wadi el Ghayl, Aseer Mountains, Saudi Arabia. Egyptian Journal of Desert Research. 2005;55:39–52. [Google Scholar]
- Ghazanfar S.A., Fisher M. Kluwer; London: 1998. Vegetation of the Arabian Peninsula. [Google Scholar]
- Hegazy A.K., El-Demerdash M.A., Hosni H.A. Vegetation, species diversity and floristic relations along an altitudinal gradient in south-west Saudi Arabia. Journal of Arid Environments. 1998;38:3–13. [Google Scholar]
- Hill M.O. Cornell University; Ithaca, NY: 1979. DECORANA – A FORTRAN Program from Detrended Correspondence Analysis and Reciprocal Averaging. [Google Scholar]
- Hill M.O. Cornell University; Ithaca, NY: 1979. TWINSPAN – A FORTRAN Program from Arranging Multivariate Data in an Order Two Way Table by Classification of the Individuals and Attributes. [Google Scholar]
- Hosni H.A., Hegazy A.K. Contribution to the flora of Asir, Saudi Arabia. Candollea. 1996;51:169–202. [Google Scholar]
- Kassas M., Girgis W.A. Habitat and plant communities in the Egyptain desert. V: The limestone plateau. Journal of Ecology. 1964;52:107–119. [Google Scholar]
- Kassas M., Imam M. Habitats and plant communities in the Egyptian desert. III. The wadi bed ecosystem. Journal of Ecology. 1954;42:424–441. [Google Scholar]
- Magurran A.E. Princeton University Press; Princeton, NJ: 1988. Ecological Diversity and its Management. [Google Scholar]
- Mandaville, J.P., 1986. Plant life in the Rubál-Khali (the Empty Quarter), south central Arabia. In: Hedge, I.C. (Ed.), Plant Life of South West Asia. Proceedings of the Royal Society Edinburgh, Section B, vol. 89, pp. 147–157.
- Mandaville J.P. Kegan Paul; London and NCWCD, Riyadh: 1990. Flora of Eastern Saudi Arabia. [Google Scholar]
- Migahid A.M. vols. 1 & 2. Riyadh University; Riyadh, Saudi Arabia: 1978. (Flora of Saudi Arabia). [Google Scholar]
- Orshan G. The desert of the Middle East. In: Evenari M., Noy-Meir I., Goodall D.W., editors. Ecosystems of the World, 12 B, Hot Deserts and Arid Shrublands. Elsevier; Amsterdam: 1986. pp. 1–28. [Google Scholar]
- Pielou E.C. John Wiley; New York: 1975. Ecological Diversity. [Google Scholar]
- SAS, 1989–1996. SAS/STAT User’s Guide. SAS Institute Inc., Cary, NC, USA.
- Schulz E., Whitney J.W. Vegetation in north-central Saudi Arabia. Journal of Arid Environment. 1986;10:175–186. [Google Scholar]
- Shaltout K.H., Mady M. Analysis of raudhas vegetation in Central Saudi Arabia. Biodiversity and Conservation. 1996;5:27–36. [Google Scholar]
- Shaltout K.H., Sheded M.G., Salem A.M. Vegetation spatial heterogeneity in a hyper arid biosphere reserve area in north Africa. Acta Botanica Croatea. 2010;69(1):31–46. [Google Scholar]
- Sharaf El-Din A., El-Kady H.F., Shaltout K.H., Mady M.I. Nutrative value of the Raudhas plants in Central Saudi Arabia. Arab Gulf Journal of Scientific Research. 1998;16(3):537–553. [Google Scholar]
- Siddiqui A.Q., Al-Harbi A.H. A preliminary study of the ecology of Wadi Hanifah stream with reference to animal communities. Arab Gulf Journal of Scientific Research. 1995;13(3):695–717. [Google Scholar]
- Springuel I., Sheded M., Darius F., Bornkamm R. Vegetation dynamics in an extreme desert wadi under the influence of episodic rainfall. Polish Botanical Studies. 2006;22:459–472. [Google Scholar]
- Taia W.K., El-Ghanem W.M. City vegetation analysis of three habitats at El-Riyadh. Bulletin of Pure and Applied Sciences. 2001;20B(1):53–65. [Google Scholar]
- Ter Braak, C.F.G., Smilauer, P., 2002. CANOCO Reference Manual and CanoDraw for Window’s User’s Guide: Software for Canonical Community Ordination (version 4.5). Microcomputer Power, Ithaca, NY.
- Vesey-Fitzgerald D.F. The vegetation of central and eastern Saudi Arabia. Journal of Ecology. 1957;45:779–798. [Google Scholar]
- White F., Leonard J. Phytogeographical links between Africa and southwest Asia. Flora et Vegetation Mundi. 1991;9:229–246. [Google Scholar]
- Yousef M.M., El-Sheikh A.M. The vegetation alongside a running water canal at Al-Kharj. Journal of College of Science. 1981;12(1):23–51. [Google Scholar]
- Yousef M.M., El-Sheikh A.M. Observations on the vegetation of gravel desert areas with special reference to succession in Central Saudi Arabia. Journal of College Science. 1981;12(2):331–351. [Google Scholar]
- Zoghet M.F., Alsheikh A.A. Academic Publishing & Press, King Saud University; Riyadh: 1999. Wild Plants in the Region of Riyadh. 234 p. [Google Scholar]
- Zohary M. Geobotanical Foundations of the Middle East Gus. Fischer Verlage; Stuttgart: 1973. [Google Scholar]