

Abstract
Ephedra alata Decne is a perennial shrub and it is a very effective sand-binder. In Saudi Arabia, the species is associated with sand dunes formation, especially the mobile, non-saline and low moisture content ones. Its geographical distribution in Saudi Arabia includes the Northern, Eastern and Central regions. The aims of this study were to determine the effects of temperature, water potential and Sodium Chloride on germination of E. alata. Seeds were collected from King Khalid Centre of Wildlife Research and Development at Thumama (80 km north east of Riyadh), Saudi Arabia. Seeds were germinated at four alternating temperature regimes (8/22; 9/23; 13/27 and 18/35 °C). Seeds were also germinated under stress of aqueous Polyethylene Glycol (PEG) solutions mixed to create water potentials of 0; −0.3; −0.6; −1.2 and −1.5 MPa. Seed were also germinated in Sodium Chloride solutions of 0, 0.05, 0.1, 0.2 and 0.3 mol l−1. Optimum germination was attained at 13/28 °C that corresponds to temperatures prevailing during spring time. Seeds germinated in Polyethylene Glycol solutions exhibited significantly lower germination than control especially when water potential fell below −0.3 MPa. Germination was also negatively affected by 0.1 mol l−1 Sodium Chloride solution or above. Results indicated that the germination temperature responses of the nondormant seeds synchronize the event of germination with the season when environmental conditions are more favorable for subsequent growth and seedling establishment. Germination was also sensitive to both water potential and salinity.
Keywords: Ephedra alata, Salinity, Temperature regimes, Water potential, Germination
1. Introduction
Ephedra alata Decne is a gymnosperm (family Ephedraceae). Only two species, of Ephedra are represented in the native flora of Eastern Saudi Arabia, E. alata is a perennial shrub, stiff, yellow-green, densely branched, 40–100 cm tall and often wider than high. Twigs striate. Cones sessile and clustered in the axils (Mandaville, 1990; Migahid, 1996; Collenette, 1999). In Saudi Arabia, the species is associated with sand dunes formation, especially the mobile ones since it is a very effective sand-binder and resist the desertification (Abdallah and Chaieb, 2007). Its geographical distribution in Saudi Arabia includes the Northern, Eastern and Central regions. The Seeds were collected from King Khalid Centre of Wildlife Research and Development at Thumama (80 km north east of Riyadh), Saudi Arabia.
Ephedra have long been important as source of the alkaloid ephedrine, avaso-constricting drug formerly much used in the treatment of asthma and as general respiratory decongestant (Mandaville, 1990). The plant is exposed to extinction in Saudi Arabia due to excessive grazing (Guenther et al., 2005). The mechanisms which regulate germination are of great importance amongst the many processes, which constitute the adaptations of plants to their arid environment (Mahmoud et al., 1983). Two of the most important mechanisms responsible for coordination of germination with physical parameters of the environment, are the temperature response characteristics and the proportion of the seed with dormancy restriction on growth (Thomponson, 1973). The work described here shows the effects of temperature, water potential and Sodium Chloride solutions on germination of E. alata from Saudi Arabia. This study was undertaken to investigate the interactive effects of temperature with matric and osmotic potential on germination characteristics of E. alata. High molecular weight of Polyethylene Glycol (PEG) has been used with certain precautions, to simulate matric potential in germination studies (Kaufmann and Ross, 1970; McWilliam and Phillips, 1971; Sharma, 1973, 1976).
Temperature, soil water status and salinity are some of the most important environmental conditions that affect seed germination and hence seedling establishment and survival in arid areas. Sodium Chloride is conventionally used to study the effect of salinity (osmotic potential) on germination studies as it is easy to handle (Young et al., 1983). Also, Sodium Chloride is considered as one of the dominant salts in soils of Saudi Arabia (Bashour et al., 1983) as well as other arid rangelands. The effect of salt stress on desert seeds germination was reported by many authors (e.g. Demir et al., 2003; El-Keblawy and Al-Rawai, 2005; El-Khouly, 2006).
2. Material and methods
2.1. Experiment 1
The seeds of E. alata were collected from King Khalid Centre for Wildlife Research and Development at Thumama, 80 km northeast of Riyadh, Saudi Arabia. Seeds were germinated at four alternating temperature regimes (9/22, 10/24, 13/28 and 18/36 °C). These alternating temperature regimes were selected after consideration of metrological data at station within the habitat range of the species (Table 1). The seeds were germinated on filter papers placed in 10 cm diameter plastic dishes in dark incubators maintained at the appropriate temperature regimes. Four replicates (20 seeds each) were used. A seed was considered to have germinated when the radical emerged. Germinated seeds were checked daily and discarded immediately until no seed had germinated for seven successive days; counting involved removal of germinator lid and that allowed a change of air and briefly exposed the seeds to light.
Table 1.
Duration of temperatures °C (maximum; minimum and mean) and distribution of rainfall (mm) of Thumama region, Riyadh, Saudi Arabia.
| Month | Temperature (°C) |
Rainfall (mm) | ||
|---|---|---|---|---|
| Maximum | Minimum | Mean | ||
| January | 20.0 | 6.9 | 13.4 | 16.8 |
| February | 23.0 | 9.0 | 16.0 | 10.3 |
| March | 27.6 | 13.1 | 20.4 | 22.7 |
| April | 33.3 | 18.2 | 25.9 | 27.3 |
| May | 39.4 | 23.3 | 31.9 | 3.8 |
| June | 42.5 | 25.0 | 34.6 | 0.0 |
| July | 43.7 | 26.3 | 35.8 | 0.0 |
| August | 43.7 | 26.2 | 35.7 | 0.0 |
| September | 40.5 | 22.7 | 32.3 | 0.0 |
| October | 35.4 | 18.1 | 27.2 | 1.4 |
| November | 28.2 | 13.4 | 20.8 | 13.9 |
| December | 22.1 | 8.5 | 15.2 | 17.4 |
2.2. Experiment 2
Different water potentials were obtained with different solutions of Polyethylene Glycol (PEG) 8000. PEG solutions were prepared according to Michel (1983) to give water potentials of 0; −0.3; −0.6; −0.9; −1.2 and −1.5 MPa. Seeds were kept in moist plastic dishes with diameter 9.0 cm and were incubated in dark incubators maintained at 13/28 °C. All seeds with at least 1 mm radical length were considered as germinated. This was preferred to a moist solid matrix as it allowed continues monitoring germination of seeds.
2.3. Experiment 3
This experiment was conducted in order to assess the effect of different concentrations of Sodium Chloride solution on germination of E. alata. Seeds were germinated on substrata moistened with 100 ml (0; −0.05; −0.1; −0.2 and −0.3 mol l−1) of Sodium Chloride. Germination was made in plastic dishes kept in dark incubators maintained at 13/28 °C. The germinated seeds were discarded immediately and seed counts were mad daily until no seeds germinated for 14 successive days.
3. Results
3.1. Experiment 1
The germination percentage of seeds at different alternating temperature is shown in Fig. 1. The germination percentages increased with rise of temperature attaining their maximum at 13/28 °C being lower at 10/24 °C and even lower at 9/22 °C. The germination temperature responses of seeds of E. alata indicate that the most favorable temperature regime is that of 13/28 °C.
Figure 1.
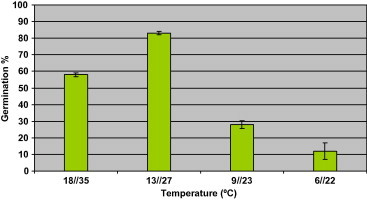
Percentage of seed germination of Ephedra alata in response to temperature.
3.2. Experiment 2
The percentages of seed germination at different water potential are shown in Fig. 2. Germination rate deceases concomitantly with decreased of water potential. A reduction of up to 36% was observed in percent of germination when seeds were exposed to −1.5 MPa compared to the control treatment (80%). A water potential lower than −0.6 MPa may be a critical to germination. Percent of germination was reduced to half when water potential fell from 0 to −1.5 MPa.
Figure 2.
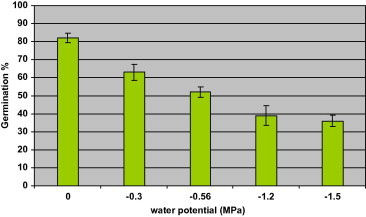
Percentage of seed germination of Ephedra alata in response to water potential.
3.3. Experiment 3
Data in Fig. 3 shows germination percentage at different concentrations of Sodium Chloride. Germination E. alata seeds were poor in response to osmotic potential. The reduction of germination percentage due to the highest NaCl concentration examined (0.1 mol l) was 96% compared to the control treatment. At 0.2 and 0.3 mol l−1 NaCl seed of E. alata failed completely to germinate.
Figure 3.
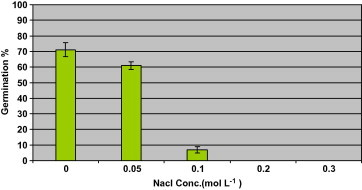
Germination percentage of Ephedra alata seeds in response to concentration of Sodium Chloride.
Statistical analysis: Mean values were taken from measurements of four replicates and Standard Error of the mean calculated. Analyses were done using the SPSS (version 15).
4. Discussion
In desert, where rainfall is unpredictable, it is important that seeds do not germinate unless the soil contains enough water to enable the resulting seedlings to complete their life cycles. It is therefore significant for germination to be controlled by a regulated response to the environment. One mechanism that regulates germination and contributes to the survival of desert plants is the presence, in their seed, of water-soluble, water-leachable germination inhibitors (Mahmoud, 1985). At the fluctuating temperature regimes the final germination percentages attained by the seed was high at the 13/28 °C regime, but it decreased at 9/22 °C (Fig. 1). Evidence from Meteorological data (Figs. 4 and 5) shows that summer was hot and dry (43 °C = mean maximum, 26 °C = mean minimum air temperature), but the winter begins with cool rainy season (22 °C = mean maximum, 9 °C = mean minimum air temperature) (Fig. 5). Germination may start at the beginning of the warm rainy season, these responses synchronize the event of germination with the season of optimal environment conditions for subsequent growth and establishment of seedlings. The optimum temperature response of E. alata seed germination is in harmony with the climatic conditions prevailing in its natural habitat in Riyadh. In other words, the germination temperature response of seeds of E. alata is at its best at 13/28 °C (temperature in spring time). Similarly, Al-Qarawi et al. (1996) found that Achillea fragrantissima germinated best during the period of spring.
Figure 4.
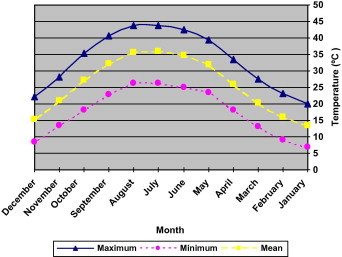
Meteorological data obtained at Riyadh where seed of Ephedra alata were collected. Mean monthly temperature, maximum and minimum. Data averages for the period 1978–2008.
Figure 5.
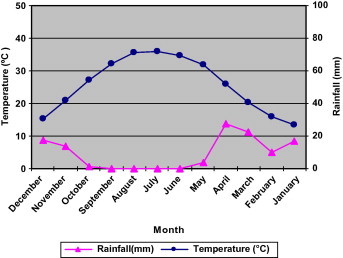
Meteorological data obtained at Riyadh where seed of Ephedra alata were collected. Data averages for the period 1978–2008.
Germination was sensitive to water potential (−0.6 to −1.5 MPa) since it lower germination at potential below −0.3 MPa. Similarly, Pangou et al. (2008) found that germination of Entandrophragma cylindricum was also very sensitive to water potential (0 to −0.75 MPa) with no germination occurring at potential below −0.25 MPa. The seed germinated better at low water potential in the vicinity of the optimum temperatures.
In general, it is agreed that salinity affects germination by creating a potential sufficiently low to inhibit water uptake and/or by providing conditions for the entry of ions that may be toxic to the embryo (Bewley and Black, 1982). In saline seedbeds, however, osmotic potential is not necessarily additive to soil matric potential effect (Roundy, 1987). Sharma (1973) said that NaCl may lowers the osmotic potential and thus facilitating hydration of germinated seeds. The osmotic potential of a 0.3 mol l−1 solution of NaCl is about −1.5 MPa. Comparing the effect of both types of water potential, the present results indicate that the germination of E. alata seeds is lower in NaCl solutions. Ions may enter the seeds affecting their viability.
In conclusion, the best conditions for the germination of the seeds of E. alata is in spring season. The seed of E. alata allows germination to take place only when ecological conditions would permit further establishment. Otherwise the survival of the species is at stake. When moisture and temperature are favorable, the seeds of E. alata can germinate in 0.05 mol l−1 NaCl. This gives an increased probability of propagates finding fresh habitats that is suitable.
References
- Abdallah L., Chaieb M. Water status and growth phonology of a Sahran shrub in north Africa. Afr. J. Ecol. 2007;1(45):80–85. [Google Scholar]
- Al-Qarawi A.A., Assaeed A.M., Al-Doss A.A. Effect of time of sowing on emergence and seedling growth of Achillea fragrantissima. Egypt. J. Appl. Sci. 1996;11:168–175. [Google Scholar]
- Bashour I.I., Al-Mashhady A.S., Prasad J.D., Miller T., Mazroa M. Morphology and composition of some soils under cultivation in Saudi Arabia. Geoderma. 1983;29:327–340. [Google Scholar]
- Bewley J.D., Black M. Physiology and Biochemistry of Seeds in Relation to Germination. vol. 2. Springer-Verlag; New York: 1982. (Viability Dormancy and Environmental Control). [Google Scholar]
- Collenette, S. 1999. Wildflowers of Saudi Arabia. National Commission for Wildlife Conservation and Development (NCWCD). Riyadh, Kingdom of Saudi Arabia, 799p.
- Demir I., Mavi K., Ozcoban M., Okcu G. Effect of salt stress on germination and seedling growth in serially harvested aubergine (Solanum melongena L.) seeds during development. Isr. J. Plant Sci. 2003;51:125–131. [Google Scholar]
- El-Keblawy A., Al-Rawai A. Effects of salinity, temperature and light on germination of invasive Prosopis juliflora (Sw.) DC. J. Arid Environ. 2005;61:555–565. [Google Scholar]
- El-Khouly A. Influence of water stress and salinity on germination and seedling growth of three medicinal desert plant species. Al-Azhar Bull. Sci. 2006;17(1):103–129. [Google Scholar]
- Guenther R., Gilbert F., Zalat S., Salem K.A. Vegetation and grazing in the St. Katherine protectorate, South Sinai, Egypt. Egypt. J. Biol. 2005;7:55–66. [Google Scholar]
- Kaufmann M.R., Ross K.J. Water potential, temperature and kinetin effects on seed germination in soil and solute systems. Am. J. Bot. 1970;57:413–419. [Google Scholar]
- Mahmoud A. Germination of Cassia italica from Saudi Arabia. Arab Gulf J. Sci. Res. 1985;3(2):437–447. [Google Scholar]
- Mahmoud A., El-Sheikh A.M., Abdulbaset S. Germination of Artemisia abyssinica sch. Bip. J. Coll. Sci. King Saud Univ. 1983;14:253–272. [Google Scholar]
- Mandaville, J.P., 1990. Flora of Eastern Saudi Arabia. Kegan Paul International London and New York Jointly with the National Commission for Wildlife Conservation and Development. Riyadh, Saudi Arabia, p. 482.
- McWilliam J.R., Phillips P.J. Effect of osmotic and matric potentials on the availability of water for seed germination. Aust. J. Biol. Sci. 1971;24:423–431. [Google Scholar]
- Michel B.E. Evaluation of the water potentials of polyethylene glycol 8000 both in the presence and absence of other solutes. Plant Physiol. 1983;72:66–70. doi: 10.1104/pp.72.1.66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Migahid, A.M., 1996. Flora of Saudi Arabia, fourth ed. vol. 1. King Saud Univ. Press, Riyadh, Saudi Arabia, 251p.
- Pangou S.V., Zoysa De N., Tsouboula M.L., Lechon G. Effect of temperature and water optional on germination of Entandrophragma cylindricum (Sprague) seed of different provenances. Seed Sci. Technol. 2008;36(1):462–466. [Google Scholar]
- Roundy, B.A., 1987. Seedbed salinity and the establishment of range plants. In: Frasier, G.W., Evans, R.A. (Eds.), Proceedings of Symposium-Seedbed Ecology of Rangeland Plants. USDA, Agr. Res. Serv., Nat. Tech. Inform. Serv., 5285 Port Royal Rd., Springfield, VA 22161, pp. 68–81.
- Sharma M.L. Simulation of drought and its effect on germination of five pasture species. Agron. J. 1973;65:423–431. [Google Scholar]
- Sharma M.L. Interaction of water potential and temperature effects on germination of three semi-arid plant species. Agron. J. 1976;68:390–394. [Google Scholar]
- Thomponson P.A. Seed germination in relation to ecological and geographical distribution. In: Heywood V.H., editor. Taxonomy and Ecology. Academic Press; London, New York: 1973. pp. 93–119. [Google Scholar]
- Young, J.A., Evans, R.A., Roundy, B.A., Cluff, G.J. 1983. Moisture stress and seed germination. USDA Agri. Res. Serv. ARM-W-36. Oakland, CA. 41p.