

Abstract
Man made ecosystems of dry lands are key habitats due to their ecological characteristics to survey biodiversity. This study investigated bird diversity in three oases of the Northern Algerian Sahara (i.e., Biskra in 2006, Oued Souf in 2008 and Ouargla in 2009), by using the spot-mapping method. Bird density “D” (pairs/10 ha), species richness “S” (number species), diversity (Shannon index) “H′” (in bits), and evenness “E” varied from one oasis to another (Biskra: D = 98.5, S = 47, H′ = 4.49, E = 0.81; Oued Souf: D = 96, S = 33, H′ = 3.9, E = 0.77; Ouargla: D = 91.5, S = 44, H′ = 4.39, E = 0.80). Differences in bird diversity between the monitored palm groves are due to the ecological characteristics of each environment. Documented literature outlined close taxonomic similarities between bird assemblages of the study area with many Northern Saharan oases. The Hybrid Sparrow Passer domesticus x Passer. hispaniolensis and some Columbidae species including Columba livia, Streptopelia turtur, Streptopelia senegalensis, and Streptopelia decaocto were the abundant species throughout surveyed oases in which they represented more than half (55.6%) of the sum of species densities (D = 286 pairs/10 ha). These synthropic species have known a huge expansion of their distribution range throughout Algerian oases. The correspondence analysis allowed the aggregation of both families and species into oasis they belong to. One-way ANOVA was tested to analyse variations of both family and species densities between studied oases. The ANOVA revealed there was no significant variation either in family densities (p = 0.937) or in bird densities (p = 0.622) between the surveyed oases because of the small size of bird populations.
Keywords: Oasis, Bird census, Biodiversity, Drylands, Biskra, Oued Souf, Ouargla, Algerian Sahara Desert
1. Introduction
Biodiversity has captured the world’s attention since its conservation maintains several ecosystem services and impacts on human wellbeing (MEA, 2005; Harrington et al., 2010). Preserving biological diversity in arid lands, which have severe ecological characteristics; infers deliberate human action to conserve the diversity of species, populations and ecosystems (Darkoh, 2003). Achieving this goal and implementing this intention means adopting the best possible strategies, which must be based on reliable scientific knowledge (Child, 2003).
Dry lands have been considered as areas of limited interest for biodiversity. However, this perception has changed since researches have now revealed that arid ecosystems are (i) the home to unique species with special characteristics, (ii) the origin of many important crops, and (iii) a significant resource of genes (Shachak et al., 2005). Moreover, the Sahara Desert is the largest and harshest desert worldwide that is characterized by severe edaphic and climatic conditions against the survival of living beings. However, this ecosystem has a very peculiar flora and fauna, which live harmoniously and efficiently by using the limited-resources at their disposal (WWF-World Wildlife Fund, McGinley, 2007).
Man made ecosystems are of high importance in reception and refuge of diverse life forms (Céréghino et al., 2008), including birds, particularly in habitats of hot and arid regions (Roshier et al., 2001; Mwaura, 2010; Chenchouni, 2012). Furthermore, date palm oases are key agrosystems due to their mesic and ecological characteristics for monitoring and assessing faunal diversity in hot-arid lands (Sellami, 2008). It should be noted that oases are considered a type of inland wetlands according to the Ramsar Convention (Ramsar, 2007).
The early data on birds of Saharan oases were published by (Heim de Balsac and Mayaud, 1926; 1962; Etchecopar and Hüe, 1964, 1964; Dupuy, 1966). Updated and synthesised data for Algerian avifana were presented by some workers (Ledant et al., 1981; Isenmann and Moali, 2000; Samraoui and Samraoui, 2008). However, the detailed monitoring, of one to three years, at local scale has been carried out by Boukhemza (1990) and Chenchouni (2012). Similar studies on birds at the oases of southern Tunisia have been undertaken by Selmi (2000). So, the current researches on wildlife animals, including avifauna of Northern Sahara, remain insufficient and often limited to qualitative-fragmentary and/or occasional observations. Therefore, this approach comes as a large-scale regional synthesis on bird assemblage diversity. This study also provides an updated assessment of bird diversity in the oases of Oued Souf, Ouargla and Biskra (Algeria) located at the northern Sahara Desert.
2. Materials and methods
2.1. Study area
Birds’ fauna was surveyed in several oases-agrosystems of three regions in the Algerian Northern Sahara i.e., Biskra, Oued Souf and Ouargla. All these oases, either modern (well maintained) or traditional (not maintained), were mainly composed of date palm grove plantations (Phoenix dactylifera), which are cultivated on sandy-saline soils. Climate is typically hyper-arid where the drought spreads throughout the year (Chenchouni, 2012). The coldest month is January in both Biskra and Oued Souf, and December in Ouargla with an average temperature of 10.2 °C, 11.6 °C and 11.4 °C, respectively. The hottest month is August in Biskra (37.0 °C) and July in Oued Souf and Ouargla with average temperatures of 35.7 °C and 38.2 °C, respectively. Precipitation occurs in the winter that extends over almost two months per year. Rainfall amount is characterized by a large interannual irregularity.
-
-
The oasis of Biskra is bordered at the north by the foothills of the southern Saharan Atlas Mountains (Aures Mountains). Birds were sampled from a modern and well-maintained palm grove plantation in the Filiach region (34°50′N, 5°45′E), located 5 km southeast of Biskra city and at an elevation of 87 m above the sea level (Fig. 1). The sampled palm grove included 24 ha of palm trees interspersed with fruit trees including olive, apricot and lemon trees and cereals. There are also green-windbreaks represented by Casuarina torulosa, Eucalyptus sp. and Tamarix gallica (Souttou et al., 2004).
-
-
Oued Souf Region (33°12′N to 33°35′N and 6°50′E to 6°51′E) is located between the southeast limit of Saharan Atlas Mountains and the northern borders of Oriental Erg (Fig. 1). This large sandy region is surrounded by large Saharan wetlands on the three sides; by large salt Lakes “Chotts” of Oued-Righ region in the west, by Chotts Merouane and Melrhir from the north, and by the large Tunisian Chott of El-Jerid from the east. Birds were surveyed in the traditional date palm cultivation system “Ghout” that is well known in Oued Souf, wherein palm trees are grown in large excavation-plots (1–12 ha) that enable palms to directly reach near aquifers without any further irrigation (Remini and Hallouche, 2008).
-
-
Oases of Ouargla (31°58′N, 5°20′E) are located in the southeast of Algeria at 800 km from Algiers (Fig.1), at a mean elevation of 134 m above the sea level. This area is located at the bottom of Oued Mya Valley and is bordered on the north by Bour El-Haicha. Birds were sampled from traditional (not maintained) oasis of 100 ha of area.
Figure 1.
Location of surveyed oases of Northern Algerian Sahara. The study area is indicated by the dashed square.
2.2. Bird sampling and data analysis
Census of bird species in each oasis was carried out from February through July, including the breeding season (mid February–May). The spot-mapping method, also known as territory-mapping (Bibby et al., 1992), was used. Birds were sampled in Biskra in the year 2006, in El Oued in 2008, and in Ouargla in the year 2009, where a set of twelve counts were carried out in each oasis (about two counts per mouth). Sampling duration was 120 min for each count. Within each palm-grove, a plot of 10 ha of area was chosen to record all individual birds seen or heard on a gridded map. The territory-mapping method is considered the most accurate method for measuring species densities (di) and then abundance of avian communities (D), particularly in open habitats; provided it is applied under favourable weather conditions (Bibby et al., 1992).
2.3. Data Analysis
Relative abundance (RA) was calculated for families by the number of species included in that family/the total number of species recorded in a given area. Species richness (S) was estimated by the total number of species or sub-species identified in a given oasis. Shannon index (H′ = -∑pi × log2pi) and evenness (E = H′/log2S) were applied for the measurement of bird diversity (α-diversity) in each oasis on the basis of the relative density (number of individual) pi of the ith species (Magurran, 2004).
First, a descriptive statistical analysis, using correspondence analysis (CA), was performed to examine segregation degree of families and species (considering their densities data) according to the surveyed oasis. In a second step, one-way ANOVA test was carried out to investigate the significance of bird density variation within the studied oases.
3. Results
3.1. Bird taxonomic diversity
A total of 59 bird species classified into seven orders, 23 families and 38 genera were recorded in the oases of North-eastern Algerian Sahara (Table 1). Of these, 47 species belonging to six orders, 21 families and 34 genera were observed in the Biskra oases (Fig. 2), of which family Turdidae was represented by the highest number of species i.e., nine (RA = 19.1% of the total inventory), followed by Columbidae and Sylviidae (five species each, RA = 10.6%). Moreover, 44 species (29 genera, 17 families and five orders) were censused in Ouargla. Turdidae was the most dominant family with eight species (RA = 18.6%), followed by Sylviidae with seven species (RA = 16.3%). In 2008, a set of 33 birds was inventoried in Oued Souf; these species belonged to 24 genera, 17 families and five orders (Fig. 2). Sylviidae held six species (RA = 18.8%) followed by the Columbidae and Turdidae with four species each (RA = 12.5%). Values of diversity indices (species richness, Shannon Index and evenness) recorded in the oasis of Biskra were high (S = 47, H′ = 4.49 bits, E = 0.81) compared to the oasis of Oued Souf (S = 33, H′ = 3.9 bits, E = 0.77) which had the lowest values (Table 1).
Table 1.
Species density (expressed in number of pairs/10 ha) of birds recorded during the breeding seasons in three oasis regions of northern Algerian Sahara with diversity indices (—: absent species).
| FAMILY Species | Specific density “di” (pair/10 ha) |
FAMILY Species | Specific density “di” (pair/10 ha) |
||||
|---|---|---|---|---|---|---|---|
| Biskra | Oued Souf | Ouargla | Biskra | Oued Souf | Ouargla | ||
| PHASIANIDAE | STURNIDAE | ||||||
| Alectoris barbara | 3.5 | — | — | Sturnus vulgaris | 1.5 | 2 | — |
| FALCONIDAE | SYLVIIDAE | ||||||
| Falco tinnunculus | 1 | — | — | Hippolais pallida | 0.5 | 1 | 0.5 |
| Falco biarmicus | — | — | 2 | Sylvia nana | — | — | 0.5 |
| Falco peregrinus | — | 0.5 | 1 | Sylvia deserticola | 1 | 1 | 1.5 |
| TYTONIDAE | Sylvia melanocephala | — | 0.5 | — | |||
| Tyto alba | 1 | 1 | 1.5 | Sylvia atricapilla | — | — | 0.5 |
| STRIGIDAE | TURDIDAE | ||||||
| Bubo ascalaphus | 0.5 | — | 0.5 | Turdus merula | 2 | 1.5 | — |
| Athene noctua saharae | 0.5 | 1 | 2 | Cercotrichas galactotes | 1.5 | — | 1 |
| COLUMBIDAE | MUSCICAPIDAE | ||||||
| Columba livia | 4.5 | 4 | 4.5 | Erithacus rubecula | 0.5 | — | 0.5 |
| Columba palumbus | 0.5 | — | — | Luscinia megarhynchos | 1 | — | — |
| Streptopelia turtur | 5.5 | 7 | 8.5 | Phoenicurus phoenicurus | 1 | 0.5 | 1 |
| Streptopelia decaocto | 2.5 | 6 | 6.5 | Saxicota caprata | — | — | 0.5 |
| Streptopelia senegalensis | 6.5 | 15.5 | 12 | Saxicota torquata | — | — | 1 |
| MEROPIDAE | Oenanthe oenanthe | 1 | 1 | 1 | |||
| Merops persicus | 0.5 | — | — | Oenanthe deserti | 0.5 | 0.5 | 1 |
| Merops apiaster | 2.5 | 1.5 | 1.5 | Oenanthe leucopyga | 1 | — | 1 |
| UPUPIDAE | Oenanthe leucura | 1 | 1.5 | — | |||
| Upupa epops | 1 | 0.5 | 1 | Muscicapa striata | 1.5 | 1 | 1 |
| LANIIDAE | Ficedula hypoleuca | 1 | 1.5 | 1 | |||
| Lanius meridionalis elegans | 1 | 1.5 | 1 | TIMALIIDAE | |||
| Lanius senator | 0.5 | 0.5 | 1 | Turdoides fulvus | 2.5 | 4.5 | 3 |
| PARIDAE | MOTACILLIDAE | ||||||
| Parus caeruleus | 0.5 | — | — | Motacilla alba | 1 | 2 | 1 |
| ALAUDIDAE | Motacilla flava | 0.5 | 1 | 1.5 | |||
| Ammomanes cincturus | 1.5 | — | 1 | PASSERIDAE | |||
| Passer domesticus × | |||||||
| Ammomanes deserti | 1 | 1.5 | 1 | P. hispaniolensis | 26.5 | 28 | 21 |
| Galerida cristata | 3.5 | — | — | Passer hispaniolensis | 0.5 | — | — |
| PYCNONOTIDAE | Passer simplex | — | 1.5 | — | |||
| Pycnonotus barbatus | 1 | — | — | FRINGILLIDAE | |||
| HIRUNDINIDAE | Serinus serinus | 1.5 | 2 | — | |||
| Hirundo rustica | — | 1.5 | 2 | Chloris chloris | 0.5 | — | 1 |
| Delichon urbica | — | — | 1 | Carduelis cannabina | — | — | 0.5 |
| ORIOLIDAE | Carduelis carduelis | 0.5 | — | 0.5 | |||
| Oriolus oriolus | — | — | 0.5 | EMBERIZIDAE | |||
| CORVIDAE | Emberiza striolata | 7.5 | — | — | |||
| Corvus corax | 1 | 1.5 | 1 | ||||
| PHYLLOSCOPIDAE | Total densities “D” | 98.5 | 96 | 91.5 | |||
| Phylloscopus collybita | 1 | 1.5 | 1 | Average density (±SD) | 2.1 ± 4.0 | 3.0 ± 5.4 | 2.2 ± 3.7 |
| Phylloscopus fuscatus | 1 | 0.5 | 1 | Species richness “S” | 47 | 33 | 44 |
| ACROCEPHALIDAE | Shannon’s index (bits) | 4.49 | 3.90 | 4.39 | |||
| Acrocephalus schoenobaenus | 0.5 | — | 0.5 | Evenness | 0.81 | 0.77 | 0.80 |
Figure 2.
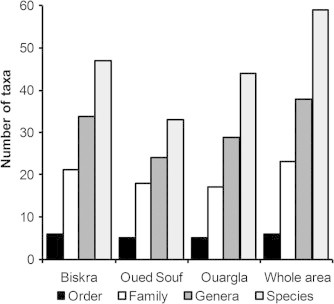
Number of bird taxa identified in the oases of Northern Algerian Sahara according to orders, families, genera and species.
3.2. Quantitative distribution of species through surveyed oases
Quantitative bird data generated from spot-mapping method revealed that the Hybrid sparrow Passer domesticus × Passer hispaniolensis has more than one third of the total number of individuals of all species inventoried in Biskra (di = 26.5 pairs/10 ha, D = 98.5 pairs/10 ha), in Oued Souf (di = 28 pairs/10 ha, D = 96 pairs/10 ha), and in the Ouargla region (di = 21 pairs/10 ha, D = 91.5 pairs/10 ha). The hybrid sparrow with Columbidae species including Columba livia, Streptopelia turtur, Streptopelia senegalensis, and Streptopelia decaocto (mean di is 4.3, 7.0, 11.3, and 5.0 pairs/10 ha, respectively) were the most abundant species throughout the surveyed oases of Northern Algerian Sahara in which they held more than half (55.6%) of sum species densities (D = 286 pairs/10 ha) (Table 1).
Correspondence analysis (CA), applied to family and species densities in relation with different sampled oases, was represented on the factorial symmetrical plot of two axes F1 × F2 with the maximum of inertia (100% hereafter). Both CA applied for densities of families (F1 = 81.54%, F2 = 18.46%) and species (F1 = 67.21%, F2 = 32.79%) showed that the x-axis separated the oasis of Ouargla and the oasis of Oued Souf. Y-axis distinguished the oasis of Biskra from the two previous oases (Fig. 3). The agglomeration of points representing the families/species on the factorial plan of the CA showed two cases: (i) observations aggregated at the centre of the plot (near the reference of two axes) are the families/species common between three oases that are dispersed over different quarters of the symmetric plot; and (ii) points located at the periphery of the factorial plot, away from its centre. Either located close to one oasis or at an intermediate position between two oases, these observations (families/species) characterize the nearest oasis (Fig. 3).
Figure 3.
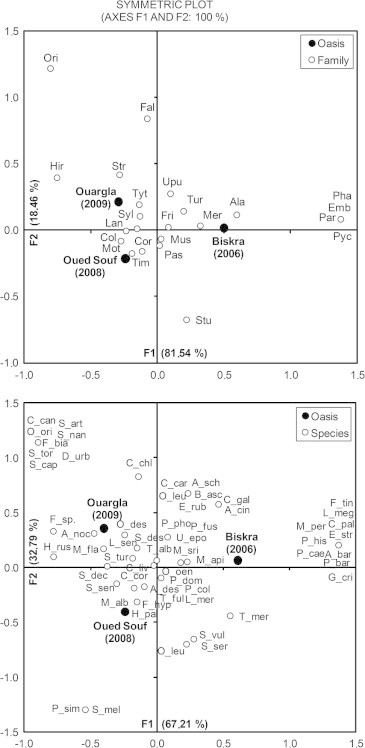
Factorial plot of the Classification Analysis (CA) applied to the distributions of family densities (a) and densities of species (b) on sampled oases. (Codification of families was reported as first three letters of each family name. Each species was coded by capitalized first letter of genus followed by first three letters of scientific name of the species).
ANOVA analysis highlighted the variation of densities of bird families and species within the sampled oases. The test revealed no significance in density variations, either for families (F = 0.48, p = 0.622) or for species (F = 0.07, p = 0.937), between the three surveyed oases (Table 2).
Table 2.
One-way analysis of variance (ANOVA) test between densities of bird families and species within three the studied oases (Biskra, El Oued, Ouargla).
| Variable | Source of Variation | Df | SS | MS | F | P |
|---|---|---|---|---|---|---|
| Density of families | Between groups | 2 | 8.61 | 4.31 | 0.07 | 0.937 |
| Within groups | 53 | 3 491.75 | 65.88 | |||
| Total | 55 | 3 500.36 | 63.64 | |||
| Density of species | Between groups | 2 | 17.77 | 8.89 | 0.48 | 0.622 |
| Within groups | 118 | 2 196.23 | 18.61 | |||
| Total | 120 | 2 214.00 | 18.45 |
4. Discussion and conclusion
The present study presents the first comprehensive ornithological survey data on birds of Algerian oases at large regional scale since works of Heim de Balsac and Mayaud (1926, 1962). In this study, all the 59 species of birds were recorded from all the surveyed oases that represents 16.9% of total Algerian species (about 350 species according to Ledant et al., 1981) and 14.5% over 406 species of birds recorded by Isenmann and Moali (2000).
The obtained results are in good agreement with the following previous studies on several local Northern African oases:
-
-
Souttou et al. (2004), nearby Filiach region, have identified 26 bird species distributed among 20 genera, 16 families and six orders. On a larger scale in the Ziban region (including Filiach oasis), on a six-year survey, Farhi and Belhamra (2009) have inventoried 124 species (including waterbirds) distributed among 15 orders and 39 families, where Passeriformes represent 58 species and 12 families.
-
-
In south Tunisian oases, Selmi (2000) reported 86 species distributed among 13 families and four orders. The most abundant family was Sylviidae with three species, followed by Columbidae, Laniidae, Turdidae and Fringillidae (two species each).
-
-
In addition, this study’s species richness values, recorded at Ouargla oases, were higher than those recorded by (Hadjaidji-Benseghier, 2009) in the same area, who noted the presence of 36 species distributed among 28 genera, 19 families and 8 orders.
-
-
This study’s findings in Oued Souf oases were very comparable to those done in some palm groves by (Degachi, 1992), who recorded 40 bird species distributed among 31 genera, 18 families and seven orders.
-
-
Besides, a qualitative bird study in Timimoun (SW Algeria) described 100 species belonging to 59 genera, 28 families and 12 orders. This large number of species may be explained by the diversity of sampled habitats such as oases, salt lake, reeds, and suburban area (Boukhemza, 1990). This author reported 36 birds (18 families and 12 orders) living particularly in oases habitats. Thus, bird taxonomic diversity values obtained in this study were also in agreement with those obtained in palm groves by Boukhemza (1990).
Compared with previous works, this detailed assessment gives clear outlines on huge similarities that exist between bird assemblages living in Northern Saharan and Algerian oases. Conversely, the slight differences observed in some bird abundances within our investigated oases may be explained by ecological heterogeneities in the types of surveyed habitats, microclimate and vegetation structure.
The five abundant species of this survey (hybrid sparrow with four Columbidae species) are synthropic species, which are known to have a huge expansion of their distribution range over the Mediterranean basin (Benyacoub, 1998; Bergier et al., 1999; Eraud et al., 2007; Bendjoudi et al., 2012) and worldwide (Barnard and Thuiller, 2008; Bonter et al., 2010; Fujisaki et al., 2010). These latter references highlighted, that at regional or global scale, the ecological impacts and other mechanisms lead to invasion and/or rapid expansion of introduced or expanding bird species. In addition, even if spatiotemporal invasion patterns of bird species at the Sahara remain little known, it is expected that the installation of new man made habitats such as large cereal crops in the Algerian Sahara during recent decades has led to this rapid expansion and outbreak of sparrows and Columbidae species, regarding their main granivorous diet. Besides, these species are considered recently as the characteristic of abundant birds and the most common in all oases of the Tunisian Sahara (Selmi, 2000).
Based on the distribution of families and species within a factorial plot of the CA, the majority of them are common among the three surveyed oases, and thus can be considered as cosmopolitan, because of their central position within the factorial plot. Nevertheless, each oasis is actually bounded by its characteristic species, which are present only in that oasis or owned, therelarge densities compared to other oases. Thus, specific and distinct species at the Oued Souf oasis are Sylvia melanocephala and Passer simplex. Characteristic species of the Biskra oasis are numerous; they belong mainly to the families of Phasianidae, Pycnonotidae, Paridae and Emberizidae. Regarding abundant species, specific for the oasis of Ouargla, they are many and they mostly belong to Falconidae, Hirundinidae and Oriolidae. These precise distributions of species and their families on the factorial plot provide accurate information on ornithological particularities and the composition of each oasis. Indeed, arid ecosystems are known to shelter a variety of species that give them their originalities and particularities, (Roshier et al., 2001) wetland ecosystems located in drylands are more attractive for birds (Chenchouni, 2012) since every species is concentrated in the refuge or habitat that responds favourably to its ecological requirements (Barnard and Thuiller, 2008; Céréghino et al., 2008; Mwaura, 2010).
There was a trend for the variability in species densities (F = 0.48, p = 0.622) to be higher between sampled oases than the variability in family densities (F = 0.07, p = 0.937). The similarities in bird abundances through the Saharan oases may be explained by large continental-scale interactions of desert bird populations, which are the contrary to the water birds of wetlands in these environments (Roshier et al., 2001; Chenchouni, 2012). In addition, small sized populations (0.5–1.5 pairs/10 ha) initially recorded make it more difficult to detect a significant variability within habitats, even if bird taxonomical differences (Fig.2) clearly appeared between the monitored palm groves, which may be originally due to ecological peculiarities of each environment (Mwaura, 2010).
The presence of several bird species in the surveyed oases is closely linked to their phenology. Considering this, many non-breeder species as well as water birds were recorded in several oases. In accordance with Isenmann and Moali (2000), the following species are not breeding in the Algeria oases: Columba palumbus, Merops apiaster, Hirundo daurica, Delichon urbicum, Motacilla alba, Motacilla flava, Lanius senator, Phylloscopus collybita, and Phylloscopus fuscatus. These species may be qualified as accidental, however some of them breed effectively in the oases (Lanius senator, Hirundo daurica) (Hadjaidji-Benseghier, 2009) while others are trans-Saharan spring migrants like Motacilla flava (Wood, 1992). In fact, the actual status and distribution of birds in the study area is quite different than those known in the literature, mainly in Isenmann and Moali (2000). This may be due to changes that happen constantly in bird communities including those of the Sahara Desert (Gaskell, 2005). Moreover, the record of Acrocephalus schoenobaenus and Sylvia deserticola (differently from what the name leads to think) occurs especially in the winter period, because these species breed on Saharan Atlas Mountains while during winter, they move southward. This probably is the case for other species too (Sylvia melanocephala, S. atricapilla, Muscicapa striata, Ficedula hypoleuca, Erithacus rubecula, Luscinia megarhynchos, Phoenicurus phoenicurus, Turdus merula, Saxicola torquatus, Oenanthe oenanthe, Sturnus vulgaris, Carduelis chloris, C. Cannabina, and Oriolus oriolus). Many of the listed species are not linked to hot-hyperarid lands, but during trans-Sahara migration, they stop on oases to refuel energy (Bairlein, 1985; Biebach et al., 1986; Lavee et al., 1991). Much work has been carried out on the bird migration strategies over the Sahara (Bairlein, 1992; Biebach et al., 2000; Herremans, 1991). Some bird species were observed in the oases during their hunting activity, such as Falco biarmicus and Falco peregrines, which breed on cliffs.
Acknowledgements
We would like to thank with gratitude the assistance of Dr. Muhammad Khalid Mukhtar (University of Sargodha, Pakistan), Mr. Youcef Alioua (University of Ouargla, Algeria) and Dr. Ashraf A. Ramadan (Environment & Life Sciences Center, Kuwait Institute for Scientific Research), for their linguistic editing of an earlier version of the manuscript.
Footnotes
Peer review under responsibility of King Saud University.

References
- Bairlein F. Body weight and fat deposition of Palaearctic passerine migrants in the central Sahara. Oecologia. 1985;66:141–146. doi: 10.1007/BF00378566. [DOI] [PubMed] [Google Scholar]
- Bairlein F. Recent prospects on trans-Saharan migration of songbirds. Ibis. 1992;134:41–46. [Google Scholar]
- Barnard P., Thuiller W. Introduction. Global change and biodiversity: future challenges. Biology Letters. 2008;4:553–555. doi: 10.1098/rsbl.2008.0374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bendjoudi, D., Chenchouni H., Doumandji, S., Voisin, J.F. (in press). Bird diversity in Plain Mitidja (N. Algeria) with emphasis on dynamics of invasive and expanding species. Acrocephalus 33.
- Benyacoub S. La Tourterelle turque Streptopelia decaocto en Algérie. Alauda. 1998;66:251–253. [Google Scholar]
- Bergier P., Franchimont J., Thévenot M. Implantation et expansion géographique de deux espèces de Columbidés au Maroc: La Tourterelle turque Streptopelia decaocto et la. Tourterelle maillée Streptopelia senegalensis. Alauda. 1999;67:23–36. [Google Scholar]
- Bibby C.J., Burgess N.D., Hill D.A. Academic Press; London: 1992. Bird census techniques. [Google Scholar]
- Biebach H., Friedrich W., Heine G. Interaction of body-mass, fat, foraging and stopover period in trans-Sahara migrating passerine birds. Oecologia. 1986;69:370–379. doi: 10.1007/BF00377059. [DOI] [PubMed] [Google Scholar]
- Biebach H., Biebach I., Friedrich W., Heine G., Partecke J., Schmidl D. Strategies of passerine migration across the Mediterranean Sea and the Sahara Desert: a radar study. Ibis. 2000;142:623–634. [Google Scholar]
- Bonter D.N., Zuckerberg B., Dickinson J.L. Invasive birds in a novel landscape: habitat associations and effects on established species. Ecography. 2010;33:494–502. [Google Scholar]
- Boukhemza M. National Institute of Agronomy; El Harrach, Algeria: 1990. Contribution à l’étude de l’avifaune de la région de Timimoun (Gourara): inventaire et données bioécologiques. Magister dissertation. [Google Scholar]
- Céréghino R., Ruggiero A., Marty P., Angélibert S. Biodiversity and distribution patterns of freshwater invertebrates in farm ponds of a southwestern French agricultural landscape. Hydrobiologia. 2008;597:43–51. [Google Scholar]
- Chenchouni H. Diversity assessment of vertebrate fauna in a wetland of hot hyperarid lands. Arid Ecosystems. 2012;2:253–263. [Google Scholar]
- Child G. Setting and achieving objectives for conserving biological diversity in arid environments. Journal of Arid Environments. 2003;54:47–54. [Google Scholar]
- Darkoh M.B.K. Regional perspectives on agriculture and biodiversity in the drylands of Africa. Journal of Arid Environments. 2003;54:261–279. [Google Scholar]
- Degachi, A., 1992. Contribution à l’étude bioécologique des peuplements d’oiseaux dans les palmeraies d’El Oued. Engineer dissertation, National Institute of Agronomy, El Harrach, Algeria.
- Dupuy A. Catalogue ornithologique du Sahara Algérien. L’Oiseau et R.F.O. 1966;39:140–160. [Google Scholar]
- Eraud C., Boutin J.M., Roux D., Faivre B. Spatial dynamics of an invasive bird species assessed using robust design occupancy analysis: the case of the Eurasian collared dove (Streptopelia decaocto) in France. Journal of Biogeography. 2007;34:1077–1086. [Google Scholar]
- Etchecopar R., Hüe E. Ed. N Boubée et Cie; Paris, France: 1964. Les oiseaux du nord de l’Afrique. [Google Scholar]
- Farhi, Y., Belhamra, M., 2009. Typologie de l’avifaune des Ziban. Poster presentation at the Int. Conference SIBFA, 22–24 Nov. 2009, Univ. of Ouargla, Algeria.
- Fujisaki I., Pearlstine E.V., Mazzotti F.J. The rapid spread of invasive Eurasian Collared Doves Streptopelia decaocto in the continental USA follows human-altered habitats. Ibis. 2010;152:622–632. [Google Scholar]
- Gaskell J. Recent changes in the status and distribution of birds in Libya. Sandgrouse. 2005;27:126–138. [Google Scholar]
- Hadjaidji-Benseghier, F., 2009. Analyse bibliographique de l’avifaune nicheuse des palmeraies de la cuvette d’Ouargla. Oral presentation, Int. Conf. SIBFA, 22–24 Nov. 2009, Univ. of Ouargla, Algeria.
- Harrington R., Anton C., Dawson T.P., de Bello F., Feld C.K., Haslett J.R. Ecosystem services and biodiversity conservation: concepts and a glossary. Biodiversity and Conservation. 2010;19:2773–2790. [Google Scholar]
- Heim de Balsac H., Mayaud N., editors. Contribution à l’ornithologie du Sahara central et du Sud algérien. Le Typo-litho; Algiers: 1926. [Google Scholar]
- Heim de Balsac H., Mayaud N. Paul Lechevalier; Paris, France: 1962. Les oiseaux du Nord-Ouest de l’Afrique. [Google Scholar]
- Herremans M. Trans-Saharan migration strategies. Ringing & Migration. 1991;12:55. [Google Scholar]
- Isenmann P., Moali A., editors. Birds of Algeria. Societé d’études ornithologiques; Paris, France: 2000. [Google Scholar]
- Lavee D., Safriel U.N., Meilijson I. For how long do trans-Saharan migrants stop over at an oasis? Ornis Scandinavica. 1991;22:33–44. [Google Scholar]
- Ledant J.P., Jacobs J.P., Jacobs P., Malher F., Ochando B., Roché J. Mise à jour de l’avifaune algérienne. Gerfault. 1981;71:295–398. [Google Scholar]
- Magurran A.E., editor. Measuring biological diversity. Wiley-Blackwell; New York: 2004. p. 256. [Google Scholar]
- MEA . World Resources Institute; Washington, DC: 2005. Ecosystems and human wellbeing: wetlands and water synthesis. Millennium ecosystem assessment (MEA) [Google Scholar]
- Mwaura F. The influence of geographic and morphometric factors on the distribution of water bird species in small high altitude tropical man made reservoirs, Central Rift Valley, Kenya. African Journal of Ecology. 2010;48:676–690. [Google Scholar]
- Ramsar, 2007. Designating Ramsar sites: The Strategic Framework and guidelines for the future development of the List of Wetlands of International Importance. Ramsar handbooks for the wise use of wetlands, 3rd edition, vol. 14. Ramsar Convention Secretariat, Gland, Switzerland, 110p.
- Remini B., Hallouche W. Les Ghouts dans les oasis d’El Oued (Algérie) Revue des régions arides. 2008;21:909–914. [Google Scholar]
- Roshier D.A., Robertson A.I., Kingsford R.T., Green D.G. Continental-scale interactions with temporary resources may explain the paradox of large populations of desert waterbirds in Australia. Landscape Ecology. 2001;16:547–556. [Google Scholar]
- Samraoui B., Samraoui F. An ornithological survey of the wetlands of Algeria: Important Bird Areas, Ramsar sites and threatened species. Wildfowl. 2008;58:71–98. [Google Scholar]
- Sellami, M.H., 2008. A Scientific Guide for Agricultural Water Management and Biodiversity Conservation inside the Northern African Oasis, In: Veritas R.I. (Ed.), Biodiversity Research Developments: Nova Publishers, pp. 45–86.
- Selmi S. Données nouvelles sur les avifaunes des oasis du sud Tunisien. Alauda. 2000;68:201–212. [Google Scholar]
- Shachak M., Gosz J.R., Pickett S.T.A., Perevolotsky A. Oxford University Press; 2005. Biodiversity in Drylands: Toward a Unified Framework. [Google Scholar]
- Souttou K., Guezoul O., Baziz B., Doumandji S. Note sur les oiseaux des palmeraies et des alentours de Filiach (Biskra, Algérie) Ornithologia Algirica. 2004;4:5–10. [Google Scholar]
- Wood B. Yellow Wagtail Motacilla flava migration from West Africa to Europe: pointers towards a conservation strategy for migrants on passage. Ibis. 1992;134:66–76. [Google Scholar]
- WWF-World Wildlife Fund, McGinley, M., 2007. Sahara desert. In: Cleveland CJ (Ed.), Encyclopedia of Earth. Environmental Information Coalition, National Council for Science and the Environment. Washington DC.