

Abstract
Background
Previous studies have demonstrated that patients with treatment-resistant depression (TRD) and treatment-sensitive depression (TSD) differed at neural level. However, it remains unclear if these two subtypes of depression differ in the interhemispheric coordination. This study was undertaken for two purposes: (1) to explore the differences in interhemispheric coordination between these two subtypes by using the voxel-mirrored homotopic connectivity (VMHC) method; and (2) to determine if the difference of interhemispheric coordination can be used as a biomarker(s) to differentiate TRD from both TSD and healthy subjects (HS).
Methods
Twenty-three patients with TRD, 22 with TSD, and 19 HS participated in the study. Data of these participants were analyzed with the VMHC and seed-based functional connectivity (FC) approaches.
Results
Compared to the TSD group, the TRD group showed significantly lower VMHC values in the calcarine cortex, fusiform gyrus, hippocampus, superior temporal gyrus, middle cingulum, and precentral gyrus. Lower VMHC values were also observed in the TRD group in the calcarine cortex relative to the HS group. However, the TSD group had no significant change in VMHC value in any brain region compared to the HS group. Receiver operating characteristic curves (ROC) analysis revealed that the VMHC values in the calcarine cortex had discriminatory function distinguishing patients with TRD from patients with TSD as well as those participants in the HS group.
Conclusions
Lower VMHC values of patients with TRD relative to those with TSD and those in the HS group in the calcarine cortex appeared to be a unique feature for patients with TRD and it may be used as an imaging biomarker to separate patients with TRD from those with TSD or HS.
Introduction
Despite the rapid progress that has been made in the development of antidepressants over the years, approximately one-third of depressed patients still fail to respond to standard antidepressant treatments [1]. Patients who do not respond to a series of standard treatments are classified as having treatment-resistant depression (TRD), while those who respond to standard treatment are classified as having treatment-sensitive depression (TSD) [2]. Treating TRD remains a common therapeutic challenge for psychiatrists [3], [4]. To overcome this challenge is to develop more effective treatment for major depressive disorder (MDD), especially TRD. This effort requires better understanding of the pathogenesis of MDD [5].
As a part of the efforts to understand the pathogenesis of MDD, the structural and functional changes associated with MDD have been explored with imaging technologies. A bulk of positron emission tomography (PET) and functional magnetic resonance imaging (fMRI) studies have accumulated enough evidence to suggest that MDD is related to widespread local changes in many brain areas within the cortico-limbic circuit (See L et al. [6] for a review). In addition, electroencephalography (EEG) studies consistently reveal frontal alpha asymmetry in patients with MDD [7] which predicted treatment response in depression [8]. Several studies have shown that positive emotions are related to greater relative activity in left frontal area; whereas negative emotions are related to higher relative activity in right frontal area [9], [10]. Therefore, frontal hemispheric difference in EEG activity may be a characteristic presentation of patients with depression.
Moreover, MDD has been increasingly conceptualized as a disorder of brain disconnectivity. Abnormality of the limbic-cortical networks is speculated to play a key role in the pathogenesis of MDD. This speculation is supported by the findings of functional connectivity (FC) studies. For example, several studies revealed that patients with MDD had alterations of orbitofrontal cortex-precuneus coupling [11] and pregenual anterior cingulate cortex (ACC)-dorsomedial thalamus coupling [12]. Other studies have shown bilateral alterations in the prefrontal regions as well as the temporal limbic regions [13]. However, there has never been a study directly exploring TRD-related alterations in functional interhemispheric coordination between the cerebral hemispheres.
Functional homotopy, the synchrony in patterns of spontaneous activity between homotopic regions in each brain hemisphere, is one of the most salient aspects of the brain’s intrinsic functional architecture [14]. The degree of correlated activity between homotopic interhemispheric counterparts may represent the importance of interhemispheric communication to integrate brain function for underlying coherent cognition and behavior [15]. As mentioned above, the vast majority of task-based and resting-state fMRI (R-fMRI) studies in MDD suggest that the affected brain regions are bihemispheric [16]. Previously, by using a 3T Siemens version scanner, we found that homotopic resting-state FC was disrupted in first-episode, drug-naive MDD [17]. Based on these findings, homotopic resting-state FC studies may offer an opportunity to investigate regional function alterations to determine whether the change in functional homotopy can be used to distinguish TRD from both TSD and HS or distinguish TSD from HS.
Patients with TRD and TSD respond to antidepressants differently, suggesting that the two subtypes of depression differ at the neural level. In a seed-based FC study, TRD was related to disrupted FC mainly in thalamo-cortical circuits, while non-treatment refractory depression was associated with more distributed decreased FC in the limbic-striatal-pallidal-thalamic circuits [18]. Recently, using the coherence-based regional homogeneity (Cohe-ReHo) method, we detected differences between the two subtypes in regional alterations in the cerebellum [19]. These data further support that the two subtypes of MDD differ at the neural level. However, it remains equivocal if the two subtypes of depression differ in interhemispheric coordination.
To determine if there is any difference in interhemispheric coordination between patients with two types of depression as well as between depressed patients and healthy subjects, we analyzed an existing dataset [19] with voxel-mirrored homotopic connectivity (VMHC) method. The VMHC is a novel approach which directly compares the interhemispheric resting-state FC [20], and has been used in studying different psychiatric conditions [15], [21]. The VMHC quantifies the resting-state FC between each voxel in one hemisphere and its mirrored voxel in the opposite hemisphere. Based on the facts that: (1) patients with TRD or TSD respond to antidepressants differently; and (2) both groups of patients had disruption in different brain circuits [18], we hypothesized that the two patient groups would exhibit reduced VMHC in different brain regions. Given that frontal dysfunction is associated with MDD [11], [22]–[24], we expected that the frontal lobe would be particularly affected. We also hypothesized that the detected changes in VMHC could be utilized as biomarkers to separate patients with TRD from those with TSD and from healthy subjects (HS).
Methods
Subjects
Twenty-four patients with TRD, 31 with first-episode, treatment-naive MDD were recruited from the Mental Health Institute of the Second Xiangya Hospital, Key Laboratory of Psychiatry and Mental Health of Hunan Province, Central South University, Hunan, China. The subjects in the present study were from one of our previous studies [19]. Among the 27 female patients, 13 (6 patients with TRD and 7 patients with treatment-naive MDD) were maternal, but none had postpartum depression.
In order to be eligible for this present study, all patients had to meet the following inclusion criteria: (1) In a current major depressive episode diagnosed with a Structured Clinical Interview for DSM-IV (SCID) [25]; (2) Within 18–50 years of age; (3) Right-handed Han Chinese; and (4) HRSD score of ≥18.
All patients with TRD were in a treatment-resistant state and had taken at least two classes of antidepressants before participating in the study. For patients with TRD there was a one week drug-free period prior to the MRI scan. This one week wash-out period was selected based on the fact that the half-life of most antidepressants (except for fluoxetine) is from 12- to 24-hours. Treatment resistance was defined as non-responsiveness to at least two adequate treatments with adequate dosage and duration (at least 6 weeks for each trial), compliance, and use of different types of antidepressants [26]. Non-responsiveness was defined as a reduction of <50% in the 17-item Hamilton Rating Scale for Depression (HRSD) [27] after the treatment with a minimum dose of 150 mg/day of imipramine equivalents for 6 weeks [28]. Detailed treatments and other clinical characters of patients with TRD are exhibited in Table S1 and S2.
The treatment-naive patients were directed to take an antidepressant at a minimum dose of 150 mg/day of imipramine equivalents for 6 weeks after the MRI scan. The antidepressant included one of the three typical classes: tricyclic antidepressants (TCAs), selective serotonin reuptake inhibitors (SSRIs), and serotonin-norepinephrine reuptake inhibitors (SNRIs). Treatment response was defined as more than a 50% reduction in the HRSD scores after treatment [26]. Only patients showing response after 6-week treatment were enrolled in the study as patients with TSD.
Twenty HS were recruited from the community. They were matched with the patient groups in age, gender, and education level. Patients or HS were excluded if they had any of the following: (1) Any history of neurological diseases, other medical illnesses, or other psychiatric disorders such as schizophrenia, bipolar disorders, or substance-induced mood disorder; (2) Any current Axis I comorbidities such as anxiety disorders, alcohol or drug dependence; (3) Any severe Axis II personality disorders or mental retardation; and (4) Any contraindication for MRI.
The study was approved by the Ethics Committee of the Second Xiangya Hospital. All participants were given information about the procedures and signed an informed consent document.
Image Acquisition
Scanning took place on a 1.5T GE scanner (General Electric, Fairfield, Connecticut, USA) equipped with high-speed gradients at the recruitment day. Participants were instructed to lie motionless, close their eyes and remain awake. The following parameters were used to obtain functional images: repetition time/echo time (TR/TE) = 2000/40 ms, 20 slices, 64×64 matrix, 90° flip angle, 24cm FOV, 5 mm slice thickness, 1 mm gap, and 180 volumes (6 min).
Data Preprocessing
Resting-state data were preprocessed in Matlab using the statistical parametric mapping software package (SPM8, http://www.fil.ion.ucl.ac.uk/spm). The first 10 volumes were discarded to ensure a steady state condition. Following steps including slice timing, head motion correction, and spatial normalization (voxel size: 3×3×3 mm3) were performed using Data Processing Assistant for Resting-State fMRI (DPARSF) [29]. The processed images were smoothed with an isotropic Gaussian kernel (full-width at half-maximum: 8 mm). Linearly trend removing and band-pass (0.01–0.08 Hz) filtering were conducted using REST [30]. Several sources of spurious covariates and their temporal derivatives were then removed using linear regression. These covariates included six head motion parameters obtained by rigid body correction, the signal from a ventricular region of interest (ROI), and the signal from a region centered in the white matter [31].
Interhemispheric Correlation
The VMHC was processed with software REST. Briefly, Pearson correlations (Fisher z-transformed) between a given voxel and a mirrored voxel in the opposite hemisphere were computed to generate VMHC maps. The details of VMHC obtainment were described in a previous study [20].
Voxel-based comparisons of the entire VMHC maps with ANOVA were conducted in REST with a significance threshold of p<0.005 for multiple comparisons using Gaussian Random Field (GRF) theory (min z>2.807, cluster significance: p<0.005, corrected). Post-hoc t-tests, restricted to the voxels showing significant differences by ANOVA analyses, were performed to identify differences between each pair of groups. The resulting statistical map was set at p<0.005 (GRF corrected). Since small amount of head motion can influence resting-state FC results [32], we computed framewise displacement (FD) for our data, which provided the temporal derivative of the motion parameters. These FD parameters were applied as covariates at the group level comparisons.
Brain regions exhibiting significant differences between the TRD and TSD groups and between the TRD and HS groups were designed as masks. Mean VMHC values were extracted within the masks for further receiver operating characteristic curves (ROC) analysis.
To examine the association of VMHC and clinical variables such as HRSD scores and illness duration, voxel-based correlations were implemented to the VMHC values of each depressed patient and clinical variables for each depressed group separately and together (p<0.005, GRF corrected).
Seed-based FC
Seed-based FC was performed using a temporal correlation approach [33]. Brain regions showing significant differences in VMHC maps between the TRD and TSD groups and between the TRD and HS groups were defined as seed ROIs. Correlation analyses were conducted between the seeds and the remaining voxels. The resulting r values were transformed to z values to improve the Gaussianity of their distribution. For each group and each seed, individual z maps were compared with a random effect one-sample t test to detect voxels showing significant correlations with the seeds (p<0.005, GRF correction). Afterwards, voxel-based comparisons of the entire brain z maps with ANOVA were conducted in REST by using an explicit mask from the union set of the one-sample t-test results (p<0.005, GRF correction). Then post-hoc t-tests were performed to identify differences between each pair of groups (p<0.005, GRF correction). The FD of each subject was used as a covariate at the group level analyses.
Results
Subjects
Data from four subjects (1 TRD, 2 TSD, and 1 HS) were excluded for further analyses due to excessive head motion. Data from 7 treatment-naive patients were discarded due to treatment non-response. Finally, the participants included 23 patients with TRD, 22 patients with TSD and 19 HS. Demographics and clinical characteristics of these participants are presented in Table 1. The three groups did not differ significantly in age, gender, education level, and the FD parameter. There was no significant difference in HRSD total scores and the scores of HRSD subscales between the patient groups. However, the TRD group had longer overall illness duration as well as current illness duration than those of the TSD group.
Table 1. Characteristics of patients with TRD or TSD and HS.
| Variables(Mean±SD) | TRD | TSD | HS | P value |
| N (M/F) | 23(11/12) | 22(12/10) | 19(10/9) | 0.898a |
| Age, years | 27.35±7.26 | 28.09±9.91 | 24.37±4.18 | 0.269b |
| Education, years | 13.13±3.44 | 12.23±2.62 | 13.11±2.47 | 0.510b |
| Mean displacement (mm) | 0.09±0.04 | 0.09±0.03 | 0.10±0.04 | 0.468b |
| Overall illness duration, months | 27.43±35.89 | 2.95±1.73 | 0.004c | |
| Current illness duration, months | 12.22±6.08 | 2.95±1.73 | <0.001c | |
| HRSD score | 24.52±4.17 | 25.89±6.26 | 0.407c | |
| Factors of HRSD | ||||
| 1 Anxiety | 6.96±1.89 | 7.78±2.16 | 0.202c | |
| 2 Weight loss | 0.57±0.90 | 0.94±0.94 | 0.195c | |
| 3 Cognitive disturbance | 5.39±1.70 | 6.28±1.93 | 0.127c | |
| 4 Retardation | 7.96±1.52 | 7.67±1.94 | 0.595c | |
| 5 Sleep disturbance | 3.65±2.01 | 3.17±1.47 | 0.395c |
The P value for gender distribution in the three groups was obtained by chi-square test.
The P values were obtained by one-way analysis of variance tests.
The P values were obtained by two sample t-test.
TRD = treatment-resistant depression.
TSD = treatment-sensitive depression.
HS = healthy subjects.
HRSD = Hamilton Rating Scale for Depression.
VMHC: Group Differences
As shown in Figure 1 and Table 2, significant differences between TRD and TSD in the VMHC values were observed throughout the cerebellum, the frontal, temporal, parietal, occipital lobes and subcortical or limbic regions by ANOVA. Compared to the TSD group, the TRD group showed significantly lower VMHC values in the calcarine cortex, fusiform gyrus, hippocampus, superior temporal gyrus, middle cingulum and precentral gyrus. Besides, lower VMHC values were observed in the TRD group in the calcarine cortex relative to the participants in the HS group. However, there was no significant difference in the VMHC values in any brain region between the TSD group and the HS group. In addition, the pooled patients (TRD and TSD) exhibited lower VMHC in the postcentral gyrus compared to the HS group (Table S3).
Figure 1. Statistical maps showing VMHC differences in different brain regions between studied groups.

Red denotes VMHC differences in the ANOVA analyses and the yellow/red color bar indicates the F value from the ANOVA analyses. Blue denotes lower VMHC and the green/blue color bars indicate the T value from post hoc analysis between compared groups. Of note, the two-sample t-test results within a mask showed significant group differences in the ANOVA analysis. VMHC = voxel-mirrored homotopic connectivity, TRD = treatment-resistant depression, TSD = treatment-sensitive depression, HS = healthy subjects.
Table 2. Significant differences in VMHC values between groups.
| Cluster location | Peak (MNI) | Cluster size | T value | ||
| x | y | z | |||
| TRD<TSD | |||||
| Calcarine Cortex | ±18 | –69 | 6 | 27 | −3.9644 |
| Fusiform Gyrus | ±33 | −75 | −15 | 81 | −4.8120 |
| Precentral Gyrus | ±57 | 0 | 33 | 31 | −4.4508 |
| Hippocampus | ±24 | −33 | −3 | 18 | −4.0928 |
| Superior Temporal Gyrus | ±63 | −45 | 12 | 31 | −3.8810 |
| Middle Cingulum | ±9 | −39 | 45 | 22 | −4.1867 |
| TRD<HS | |||||
| Calcarine Cortex | ±15 | −72 | 12 | 29 | −3.8629 |
| TSD<HS | |||||
| None | |||||
VMHC = voxel-mirrored homotopic connectivity.
TRD = treatment-resistant depression.
TSD = treatment-sensitive depression.
HS = healthy subjects.
Seed-based FC
As mentioned above, the calcarine cortex exhibited lower VMHC values in the TRD group compared to both the TSD group and the HS group. We examined whole-brain resting-state FC associated with two ROIs (one calcarine cortex per hemisphere). Most of seed ROIs showed altered FC within the brain regions of the posterior perceptual and motor areas (such as the middle temporal gyrus, the middle occipital gyrus, the precentral gyrus and the cerebellum) along with a few subcortical regions (such as the thalamus) (Figure 2, 3 and Table 3).
Figure 2. Statistical maps showing seed-based FC differences between subject groups (seed: the left calcarine cortex).

Red denotes FC differences in the ANOVA analyses and the color bar indicate the F value from the ANOVA analyses. Blue denotes lower FC and the color bars indicate the T value from post hoc analysis between each pair of groups. Of note, the two-sample t-test results within a mask showed significant group differences in the ANOVA analysis. FC = functional connectivity, TRD = treatment-resistant depression, TSD = treatment-sensitive depression, HS = healthy subjects.
Figure 3. Statistical maps showing seed-based FC differences between subject groups (seed: the right calcarine cortex).

Red denotes FC differences in the ANOVA analyses and the color bar indicate the F value from the ANOVA analyses. Red and blue denote higher and lower FC respectively and the color bars indicate the T value from post hoc analysis between each pair of groups. Of note, the two-sample t-test results within a mask showed significant group differences in the ANOVA analysis. FC = functional connectivity, TRD = treatment-resistant depression, TSD = treatment-sensitive depression, HS = healthy subjects.
Table 3. Significant differences in seed-based resting-state FC between groups.
| Seeds | Connectedregions | Peak (MNI) | Clustersize | T value | ||
| x | y | z | ||||
| TRD>TSD | ||||||
| R Calcarine | Vermis 6 | 0 | −63 | −9 | 22 | 4.5880 |
| TRD<TSD | ||||||
| L Calcarine | R Inferior TemporalGyrus | 45 | −66 | −12 | 29 | −3.7247 |
| R Calcarine | 21 | −66 | 6 | 56 | −4.2185 | |
| L Middle OccipitalGyrus | −45 | −81 | 12 | 28 | −4.1227 | |
| L Cuneus | −18 | −78 | 15 | 33 | −3.5581 | |
| R Insula | 42 | −24 | 24 | 27 | −3.4162 | |
| L Middle OccipitalGyrus | −27 | −87 | 24 | 45 | −4.2684 | |
| R Calcarine | L Lingual Gyrus | −18 | −75 | −12 | 51 | −3.7115 |
| R Lingual Gyrus | 18 | −84 | −12 | 20 | −3.9199 | |
| L/R Calcarine | 0 | −87 | 12 | 213 | −4.1546 | |
| L Middle OccipitalGyrus | −27 | −84 | 24 | 41 | −3.6235 | |
| TRD<HS | ||||||
| L Calcarine | L Inferior TemporalGyrus | −45 | −51 | −12 | 409 | −5.5896 |
| R Lingual Gyrus | 15 | −39 | −3 | 38 | −3.4187 | |
| R Postcentral Gyrus | 60 | −21 | 18 | 846 | −5.5417 | |
| L Superior TemporalGyrus | −45 | −30 | 3 | 112 | −4.3217 | |
| L Precuneus | −24 | −45 | 0 | 66 | −3.9812 | |
| R Calcarine | 21 | −66 | 3 | 29 | −3.6190 | |
| R Middle TemporalGyrus | 45 | −63 | 9 | 152 | −4.4457 | |
| L Superior Temporal Gyrus | −60 | −12 | 3 | 39 | −3.3182 | |
| L Precentral Gyrus | −48 | −6 | 24 | 157 | −4.2126 | |
| R Postcentral Gyrus | 51 | −24 | 42 | 20 | −4.4052 | |
| R Calcarine | L Middle TemporalGyrus | −45 | −54 | −3 | 398 | −4.9090 |
| R Middle TemporalGyrus | 51 | −27 | −6 | 519 | −6.4587 | |
| L Middle TemporalGyrus | −54 | −12 | −12 | 59 | −4.1010 | |
| L Lingual Gyrus | −24 | −45 | 0 | 92 | −5.1995 | |
| R Middle OccipitalGyrus | 48 | −75 | 6 | 165 | −4.0451 | |
| R Thalamus | 15 | −27 | −3 | 19 | −3.7144 | |
| L Precentral Gyrus | −48 | −6 | 24 | 20 | −3.6062 | |
| R Postcentral Gyrus | 51 | −21 | 42 | 16 | −4.0321 | |
| TSD<HS | ||||||
| R Calcarine | R Middle TemporalGyrus | 48 | −24 | −9 | 54 | −4.4069 |
| L Middle TemporalGyrus | −48 | −54 | −3 | 15 | −3.4295 | |
FC = functional connectivity; MNI = Montreal Neurological Institute; TRD = treatment-resistant depression; TSD = treatment-sensitive depression; HS = healthy subjects; R = right; L = left.
ROC Analysis between the Patient Groups
Since the calcarine cortex exhibited significant VMHC differences between the patient groups and between patients with TRD and the HS group, it might be used as a biomarker to separate the patients with TRD from those with TSD and from HS. To test this possibility, mean VMHC values were extracted from the calcarine cortex (Figure 4) and ROC analysis was conducted. The areas under the curves of the calcarine cortex were relatively high. Further diagnostic analysis revealed that the sensitivity and specificity of separating TRD from TSD and from HS were relatively high (Figure 5 and Table 4).
Figure 4. Bar plots representing the mean (and standard error) VMHC value of the calcarine cortex.

Significant differences found between the TRD group and the TSD group or the HS group (TRD vs TSD: p<0.001; TRD vs HS: p<0.01). VMHC = voxel-mirrored homotopic connectivity, TRD = treatment-resistant depression, TSD = treatment-sensitive depression, HS = healthy subjects.
Figure 5. Receiver operating characteristic (ROC) curves by using the mean VMHC value in the calcarine cortex.
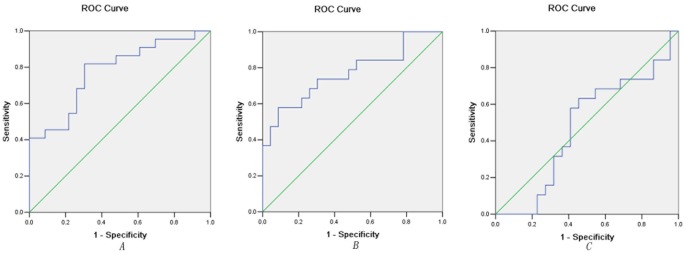
A. Separating TRD from TSD; B. Separating TRD from HS; C. Separating TSD from HS. VMHC = voxel-mirrored homotopic connectivity, TRD = treatment-resistant depression, TSD = treatment-sensitive depression, HS = healthy subjects.
Table 4. ROC analysis for differentiating different studied groups.
| Brain regions | Area Under the Curve | Cut-off point | Sensitivity | Specificity |
| Separating TRD from TSD | ||||
| Calcarine Cortex | 0.777 | 0.5497a | 73.91%(17/23) | 81.82%(18/22) |
| Separating TRD from HS | ||||
| Calcarine Cortex | 0.769 | 0.7823 | 91.30%(21/23) | 68.42%(13/19) |
| Separating TSD from HS | ||||
| Calcarine Cortex | 0.476 | 0.7498 | 54.55%(12/22) | 63.16%(12/19) |
By this cut-off point, the VMHC value of calcarine could correctly classify 17 of 23 patients with TRD and 18 of 22 patients with TSD, resulted in a sensitivity of 73.91% and a specificity of 81.82%. The means of other cut-off points were similar. VMHC = voxel-mirrored homotopic connectivity, TRD = treatment-resistant depression, TSD = treatment-sensitive depression, HS = healthy subjects.
Voxel-based Correlations between VMHC and Clinical Variables
No correlation was found between VMHC in any brain region and HRSD scores in each patient group and the combined patient group by voxel-based correlation analyses. There was also no correlation between VMHC and age, education level, the overall illness duration, or the current illness duration.
Discussion
To our knowledge, this is the first study of using the VMHC to explore interhemispheric resting-state FC in patients with TRD or TSD and healthy controls. We found that patients with TRD had lower VMHC values in the calcarine cortex compared to both TSD and HS groups. However, there was no significant difference between TSD and HS in VMHC values in the calcarine cortex.
The calcarine cortex is where the primary visual cortex is located and plays an important role in “refocusing” of attention [34]. Lower VMHC in this region may disrupt the lower-level function of attentional control. Normally, the fronto-parietal cortex is the primary source of top-down attentional control signals [35]. The modulation of visual input with attentional engagement follows as a consequence of top-down control processes [36]. In addition to direct interhemispheric interaction, lower-level areas like the calcarine cortex may act by sending projections to the high-level areas, such as the prefrontal cortex (PFC). Thereafter, reduced VMHC in this area in patients with TRD will interfere with the function of high-level areas and lead to a top-down disconnection. This kind of top-down disconnection is supported at least by one fMRI study showing that attentional shifts generate activity not only within the fronto-parietal cortex but also within the calcarine cortex [34].
Reduced VMHC values in other brain regions in patients with TRD compared to patients with TSD suggest that the disconnection of other brain regions may play a role in the pathogenesis of TRD. The fusiform gyrus is involved in the perception of facial emotion. The impairment of the fusiform gyrus has been observed in MDD [37]. As a core region of the limbic system, the hippocampus is involved not only in learning and memory [38] but also in the regulation of motivation and emotion [39]. The superior temporal gyrus plays a distinct role in emotional processing and social cognition [40]. Moreover, hypoactivity of the middle cingulum and precentral gyrus was associated with executive dysfunction and psychomotor symptoms in MDD [41]. Reduced VMHC in these regions may be the neuronal basis of depressive symptoms of TRD. Since the reduced VMHC values in these regions were only found between TRD and TSD, but not between TRD and HS or between TSD and HS, they might be state-related changes in the TRD group.
The insignificant difference between TSD and HS groups in the VMHC values is somewhat unexpected. There are a few possibilities. First is the duration of illness. The TSD group included young adults with short overall/current illness duration (mean: 2.95 months), first-episode, and drug-naive MDD. The short duration of the illness may be responsible for the insignificant VMHC change in the TSD group. Second is the sensitivity of scanners. Using a 3T Siemens version scanner, we found that patients with first-episode and drug-naive MDD had reduced VMHC values in the medial prefrontal cortex (MPFC) and the posterior cingulate cortex/precuneus (PCC/PCu) [17]. The TSD patients in the current study were similar to those in the previous study [17]. Since the original data from these two studies were obtained from different scanners, we can not make a direct group comparison. However, the results of the current study suggest that the VMHC method used in the current study might not be sensitive enough to detect VMHC change in TSD patients. Third is the statistical power. When we loosed our statistical power to uncorrected voxel-wise threshold of p<0.005, postcentral gyrus also exhibited significantly decreased VMHC values (Table S4). These data suggest that disrupted functional homotopy in certain brain areas may also attribute to the pathogenesis of TSD.
There have been reports that the PFC acts in a key role with both cognition and emotion. Abnormal activity in the PFC has been observed in patients with MDD by using both R-fMRI [42]–[44] and task-based fMRI methods [45]. These studies suggest that PFC changes are a consistent feature of depressed patients. In light of these studies, the lack of the PFC changes in both the TRD and TSD groups in the present study is somewhat unexpected. In addition to factors such as depression severity, heterogeneity of subjects, and medication treatments, there are at least two other possibilities which might account for this inconsistency. First, the inconsistency could be due to different analytic methods used in different studies. The VMHC is designed to evaluate the interhemispheric resting-state FC. It is possible that the lower-level perceptual and motor areas send unilateral projections to the high-level areas (such as the PFC) in addition to projecting directly to both hemispheres in order to interhemispherically execute their influences [46]. Thus, reduced VMHC in the lower-level areas could contribute to the deficits in the high-level areas although impaired interhemispheric interaction can not be discovered in these areas. This assumption is partly supported by our previous Cohe-ReHo analysis, which showed that the patients in the current study exhibited lower Cohe-ReHo in the bilateral superior frontal gyrus [19]. The Cohe-ReHo method was developed to measure the local signal coherence or synchronization within a functional cluster (usually 27 voxels) [47]. Evidence from functional neuroimaging studies on humans suggests that top-down attentional control processes encompass the superior frontal, inferior parietal and superior temporal cortex [36]. Combined with the previous results, the present results support the idea that the lower-level areas (such as the calcarine cortex in the present study) send unilateral projections to the high-level areas (such as the PFC) in addition to direct interhemispherically mediated influences [46].
Another possibility is the relatively strict corrected p value adopted in the present study. When we loosed our statistical power to uncorrected voxel-wise threshold of p<0.005, some PFC areas also exhibited significantly decreases in VMHC values (Table S4). This phenomenon implies that disrupted functional homotopy in the PFC areas may also attribute to the pathogenesis of MDD to some extent.
The VMHC approach adopted here to identify brain regions with abnormal interhemispheric FC has proved to be informative. However, the mechanism behind these deficits in VMHC remains equivocal. There are several possibilities for the deficits in VMHC. First, white matter deficits in the corpus callosum could disrupt the synchrony between mirrored connected regions because neural signals are not transmitted with fidelity. However, our previous analyses using the tract-based spatial statistics (TBSS) method in the same group of patients with TRD or TSD did not show any white matter alteration between the mirrored regions with decreased VMHC [48], [49]. In addition, no direct relationship between VMHC and white matter diffusion was observed previously [15], [21]. These findings imply that structural and functional metrics estimate different aspects of interhemispheric connectivity.
Second, gray matter dysfunctions might account for the deficits in VMHC. In the intrinsic neural activity, the correlation of fMRI signals of gray matter is an indirect index of synchrony [50], [51]. Similar to interhemispheric coherence, the correlation of fMRI signals between homotopically connected brain regions has been employed to measure the strength of resting-state FC [21]. The findings indicate that normal interhemispheric FC may arise from more complex pathways, such as common subcortical drivers or complex network-level synchronization, neither of which needs direct structural connectivity between cortical counterparts. VMHC is one of the most salient aspects of the brain intrinsic functional architecture [14], [16], which possibly represents the importance of interhemispheric communication to integrate brain function underlying coherent cognition and behavior [15]. Clinically, the disruption in VMHC has been reported in schizophrenia [46], depression [17], autism [21] and cocaine addiction [15].
Since epidemiological studies have shown that women have a higher risk for developing MDD than men, gender difference in brain structure and function has attracted researchers’ attention. However, the gender difference in imaging data is far from settled [52]. Some researchers have shown that there was no gender difference in whole gray matter volume, and the activity of the frontal lobe and the temporal lobe when the brain volume was controlled [53]. However, smaller brain volume has been observed in females than in males. Females had a higher percentage of gray matter, but males had a higher percentage of white matter [54]. It would be useful to conduct an fMRI study between males and females with different stages of MDD. The sample sizes in the current study are not large enough to address gender specific questions. Larger sample studies comparing gender difference in brain functioning in patients at different stages of MDD are warranted.
Since clinical variables such as the duration and severity of a depressive episode have been reported as predictors for treatment non-response [55]–[58], the finding in the present study of no correlation between the decreased VMHC values in the calcarine cortex and these variables was somewhat unexpected. Although this finding could be confounded by the small sample size of our study, it is also possible that the alterations of VMHC in the calcarine cortex may be a trait marker for patients with TRD independent of the severity of symptoms and other related factors.
In addition to the relatively small sample size, the present study has several other limitations. First, patients with TRD were in the current treatment-resistant state and had tried at least two types of antidepressants before the recruitment. Previous studies have shown that antidepressants made brain activity of depressed patients more similar to that of healthy subjects [59], [60]. Hence, the current findings in the TRD group were more likely due to the nature of the illness rather than the effects of the drugs. Second, the effect of the overall/current illness duration on the disorder remains to be debated. The TRD group had longer overall illness duration as well as longer current illness duration than those of the TSD group. The two variables were not used as covariates in the group comparisons for the reason that illness chronicity is an inherent characteristic of the TRD group and can not be removed [61]. However, no correlation was found between the VMHC in any brain region and the overall illness duration or the current illness duration by voxel-based correlation analyses. Third, cortical structures are not completely symmetrical. Using a symmetrical standard template in the current study might have biased the final results. Finally, global signal regression (GSReg) can bias correlations and complicate the explanation of negative correlations. The GSReg can also introduce a regionally varying correlation bias that is dependent on the unknown true underlying correlation structure [62]. Therefore, the GSReg was not utilized in the present study. It is unknown whether the current findings have been biased by the global signal without GRReg since the GSReg has distorting effect on interregional correlations.
In conclusion, the decreased VMHC values in the calcarine cortex may represent an imaging biomarker for TRD and may be used to separate patients with TRD from those with TSD as well as from HS. The findings also suggest that VMHC may offer an important new avenue of exploring the homotopic FC in both TRD and TSD in order to better understand the nature of the deficits in patients with MDD.
Supporting Information
Current treatment details of patients with TRD.
(DOC)
The number of episodes of patients with TRD.
(DOC)
Significant VMHC differences between pooled patients and HS (p<0.005, GRF correction).
(DOC)
Significant VMHC differences between groups (uncorrected voxel-wise threshold of p<0.005).
(DOC)
Acknowledgments
The authors express their gratitude to Mr. Timothy Warneka, a research coordinator, at the Mood Disorders Program of the University Hospitals Case Medical Center for grammatical assistance.
Funding Statement
The study was supported by grants from National Natural Science Foundations of China (grant no. 30900483, no. 81260210, no. 61035006 and no. 30971053) and the “10th Five-year-plan” of National Key Technologies R&D Program of China (grant no. 2004DA7210A21), and the special funding by Ministry of Health of the peoples’ Republic of China (grant no. 201002003). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Petersen T, Gordon JA, Kant A, Fava M, Rosenbaum JF, et al. (2001) Treatment resistant depression and axis I co-morbidity. Psychol Med 31: 1223–1229. [DOI] [PubMed] [Google Scholar]
- 2. Stimpson N, Agrawal N, Lewis G (2002) Randomised controlled trials investigating pharmacological and psychological interventions for treatment-refractory depression. Systematic review. Br J Psychiatry 181: 284–294. [DOI] [PubMed] [Google Scholar]
- 3. Little A (2009) Treatment-resistant depression. Am Fam Physician 80: 167–172. [PubMed] [Google Scholar]
- 4. Bschor T (2010) Therapy-resistant depression. Expert Rev Neurother 10: 77–86. [DOI] [PubMed] [Google Scholar]
- 5. Fagiolini A, Kupfer DJ (2003) Is treatment-resistant depression a unique subtype of depression? Biol Psychiatry 53: 640–648. [DOI] [PubMed] [Google Scholar]
- 6. L W, D FH, I BH, J L (2012) A systematic review of resting-state functional-MRI studies in major depression. J Affect Disord 142: 6–12. [DOI] [PubMed] [Google Scholar]
- 7. Carvalho A, Moraes H, Silveira H, Ribeiro P, Piedade RA, et al. (2011) EEG frontal asymmetry in the depressed and remitted elderly: is it related to the trait or to the state of depression? J Affect Disord 129: 143–148. [DOI] [PubMed] [Google Scholar]
- 8. Baskaran A, Milev R, McIntyre RS (2012) The neurobiology of the EEG biomarker as a predictor of treatment response in depression. Neuropharmacology 63: 507–513. [DOI] [PubMed] [Google Scholar]
- 9. Bruder GE, Fong R, Tenke CE, Leite P, Towey JP, et al. (1997) Regional brain asymmetries in major depression with or without an anxiety disorder: a quantitative electroencephalographic study. Biol Psychiatry 41: 939–948. [DOI] [PubMed] [Google Scholar]
- 10. Putnam KM, McSweeney LB (2008) Depressive symptoms and baseline prefrontal EEG alpha activity: a study utilizing Ecological Momentary Assessment. Biol Psychol 77: 237–240. [DOI] [PubMed] [Google Scholar]
- 11. Frodl T, Scheuerecker J, Albrecht J, Kleemann AM, Muller-Schunk S, et al. (2009) Neuronal correlates of emotional processing in patients with major depression. World J Biol Psychiatry 10: 202–208. [DOI] [PubMed] [Google Scholar]
- 12. Anand A, Li Y, Wang Y, Lowe MJ, Dzemidzic M (2009) Resting state corticolimbic connectivity abnormalities in unmedicated bipolar disorder and unipolar depression. Psychiatry Res 171: 189–198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. de Asis JM, Stern E, Alexopoulos GS, Pan H, Van Gorp W, et al. (2001) Hippocampal and anterior cingulate activation deficits in patients with geriatric depression. Am J Psychiatry 158: 1321–1323. [DOI] [PubMed] [Google Scholar]
- 14. Salvador R, Suckling J, Schwarzbauer C, Bullmore E (2005) Undirected graphs of frequency-dependent functional connectivity in whole brain networks. Philos Trans R Soc Lond B Biol Sci 360: 937–946. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Kelly C, Zuo XN, Gotimer K, Cox CL, Lynch L, et al. (2011) Reduced interhemispheric resting state functional connectivity in cocaine addiction. Biol Psychiatry 69: 684–692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Stark DE, Margulies DS, Shehzad ZE, Reiss P, Kelly AM, et al. (2008) Regional variation in interhemispheric coordination of intrinsic hemodynamic fluctuations. J Neurosci 28: 13754–13764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Guo W, Liu F, Dai Y, Jiang M, Zhang J, et al. (2013) Decreased interhemispheric resting-state functional connectivity in first-episode, drug-naive major depressive disorder. Prog Neuropsychopharmacol Biol Psychiatry 41: 24–29. [DOI] [PubMed] [Google Scholar]
- 18. Lui S, Wu Q, Qiu L, Yang X, Kuang W, et al. (2011) Resting-state functional connectivity in treatment-resistant depression. Am J Psychiatry 168: 642–648. [DOI] [PubMed] [Google Scholar]
- 19. Guo WB, Liu F, Chen JD, Gao K, Xue ZM, et al. (2012) Abnormal neural activity of brain regions in treatment-resistant and treatment-sensitive major depressive disorder: A resting-state fMRI study. J Psychiatr Res 46: 1366–1373. [DOI] [PubMed] [Google Scholar]
- 20. Zuo XN, Kelly C, Di Martino A, Mennes M, Margulies DS, et al. (2010) Growing together and growing apart: regional and sex differences in the lifespan developmental trajectories of functional homotopy. J Neurosci 30: 15034–15043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Anderson JS, Druzgal TJ, Froehlich A, DuBray MB, Lange N, et al. (2010) Decreased interhemispheric functional connectivity in autism. Cereb Cortex 21: 1134–1146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Gilbert AM, Prasad K, Goradia D, Nutche J, Keshavan M, et al. (2010) Grey matter volume reductions in the emotion network of patients with depression and coronary artery disease. Psychiatry Res 181: 9–14. [DOI] [PubMed] [Google Scholar]
- 23. Kennedy SH, Konarski JZ, Segal ZV, Lau MA, Bieling PJ, et al. (2007) Differences in brain glucose metabolism between responders to CBT and venlafaxine in a 16-week randomized controlled trial. Am J Psychiatry 164: 778–788. [DOI] [PubMed] [Google Scholar]
- 24. Liu F, Guo W, Liu L, Long Z, Ma C, et al. (2013) Abnormal amplitude low-frequency oscillations in medication-naive, first-episode patients with major depressive disorder: A resting-state fMRI study. J Affect Disord 146: 401–406. [DOI] [PubMed] [Google Scholar]
- 25.First MB, Spitzer RL, Gibbon M, Williams JBW (1997) Structured Clinical Interview for DSM-IV Axis I Disorders (SCID). Washington, DC, American Psychiatric Press.
- 26. Furtado CP, Maller JJ, Fitzgerald PB (2008) A magnetic resonance imaging study of the entorhinal cortex in treatment-resistant depression. Psychiatry Res 163: 133–142. [DOI] [PubMed] [Google Scholar]
- 27. Hamilton M (1967) Development of a rating scale for primary depressive illness. Br J Soc Clin Psychol 6: 278–296. [DOI] [PubMed] [Google Scholar]
- 28.Nierenberg AA, Amsterdam JD (1990) Treatment-resistant depression: definition and treatment approaches. J Clin Psychiatry 51 Suppl: 39–47; discussion 48–50. [PubMed]
- 29. Yan C, Zang Y (2010) DPARSF: A MATLAB toolbox for "pipeline" data analysis of resting-state fMRI. Front Syst Neurosci 4: 13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Song XW, Dong ZY, Long XY, Li SF, Zuo XN, et al. (2011) REST: a toolkit for resting-state functional magnetic resonance imaging data processing. PLoS One 6: e25031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Fox MD, Snyder AZ, Vincent JL, Corbetta M, Van Essen DC, et al. (2005) The human brain is intrinsically organized into dynamic, anticorrelated functional networks. Proceedings of the National Academy of Sciences of the United States of America 102: 9673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Power JD, Barnes KA, Snyder AZ, Schlaggar BL, Petersen SE (2012) Spurious but systematic correlations in functional connectivity MRI networks arise from subject motion. Neuroimage 59: 2142–2154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Fox MD, Snyder AZ, Vincent JL, Corbetta M, Van Essen DC, et al. (2005) The human brain is intrinsically organized into dynamic, anticorrelated functional networks. Proc Natl Acad Sci U S A 102: 9673–9678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Liu T, Slotnick SD, Serences JT, Yantis S (2003) Cortical mechanisms of feature-based attentional control. Cereb Cortex 13: 1334–1343. [DOI] [PubMed] [Google Scholar]
- 35. Corbetta M, Shulman GL (2002) Control of goal-directed and stimulus-driven attention in the brain. Nat Rev Neurosci 3: 201–215. [DOI] [PubMed] [Google Scholar]
- 36. Hopfinger JB, Buonocore MH, Mangun GR (2000) The neural mechanisms of top-down attentional control. Nat Neurosci 3: 284–291. [DOI] [PubMed] [Google Scholar]
- 37. Stuhrmann A, Suslow T, Dannlowski U (2011) Facial emotion processing in major depression: a systematic review of neuroimaging findings. Biol Mood Anxiety Disord 1: 10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Eichenbaum H, Yonelinas AP, Ranganath C (2007) The medial temporal lobe and recognition memory. Annu Rev Neurosci 30: 123–152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Seminowicz DA, Mayberg HS, McIntosh AR, Goldapple K, Kennedy S, et al. (2004) Limbic-frontal circuitry in major depression: a path modeling metanalysis. Neuroimage 22: 409–418. [DOI] [PubMed] [Google Scholar]
- 40. Gallagher HL, Frith CD (2003) Functional imaging of 'theory of mind'. Trends Cogn Sci 7: 77–83. [DOI] [PubMed] [Google Scholar]
- 41. Wang L, Li K, Zhang QE, Zeng YW, Jin Z, et al. (2013) Interhemispheric functional connectivity and its relationships with clinical characteristics in major depressive disorder: a resting state FMRI study. PLoS One 8: e60191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Yao Z, Wang L, Lu Q, Liu H, Teng G (2009) Regional homogeneity in depression and its relationship with separate depressive symptom clusters: a resting-state fMRI study. J Affect Disord 115: 430–438. [DOI] [PubMed] [Google Scholar]
- 43. Zhang J, Wang J, Wu Q, Kuang W, Huang X, et al. (2011) Disrupted brain connectivity networks in drug-naive, first-episode major depressive disorder. Biol Psychiatry 70: 334–342. [DOI] [PubMed] [Google Scholar]
- 44. Liu F, Guo W, Yu D, Gao Q, Gao K, et al. (2012) Classification of Different Therapeutic Responses of Major Depressive Disorder with Multivariate Pattern Analysis Method Based on Structural MR Scans. PLoS One 7: e40968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Chantiluke K, Halari R, Simic M, Pariante CM, Papadopoulos A, et al. (2012) Fronto-striato-cerebellar dysregulation in adolescents with depression during motivated attention. Biol Psychiatry 71: 59–67. [DOI] [PubMed] [Google Scholar]
- 46. Hoptman MJ, Zuo XN, D'Angelo D, Mauro CJ, Butler PD, et al. (2012) Decreased interhemispheric coordination in schizophrenia: a resting state fMRI study. Schizophr Res 141: 1–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Liu D, Yan C, Ren J, Yao L, Kiviniemi VJ, et al. (2010) Using coherence to measure regional homogeneity of resting-state FMRI signal. Front Syst Neurosci 4: 1–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Guo WB, Liu F, Chen JD, Xu XJ, Wu RR, et al. (2012) Altered white matter integrity of forebrain in treatment-resistant depression: A diffusion tensor imaging study with tract-based spatial statistics. Prog Neuropsychopharmacol Biol Psychiatry 38: 201–206. [DOI] [PubMed] [Google Scholar]
- 49. Guo WB, Liu F, Xue ZM, Gao K, Wu RR, et al. (2012) Altered white matter integrity in young adults with first-episode, treatment-naive, and treatment-responsive depression. Neuroscience Letters 522: 139–144. [DOI] [PubMed] [Google Scholar]
- 50. Fox MD, Raichle ME (2007) Spontaneous fluctuations in brain activity observed with functional magnetic resonance imaging. Nat Rev Neurosci 8: 700–711. [DOI] [PubMed] [Google Scholar]
- 51. Biswal BB, Mennes M, Zuo XN, Gohel S, Kelly C, et al. (2010) Toward discovery science of human brain function. Proc Natl Acad Sci U S A 107: 4734–4739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Scheibe S, Preuschhof C, Cristi C, Bagby RM (2003) Are there gender differences in major depression and its response to antidepressants? J Affect Disord 75: 223–235. [DOI] [PubMed] [Google Scholar]
- 53. Koch K, Pauly K, Kellermann T, Seiferth NY, Reske M, et al. (2007) Gender differences in the cognitive control of emotion: An fMRI study. Neuropsychologia 45: 2744–2754. [DOI] [PubMed] [Google Scholar]
- 54. Dai XJ, Gong HH, Wang YX, Zhou FQ, Min YJ, et al. (2012) Gender differences in brain regional homogeneity of healthy subjects after normal sleep and after sleep deprivation: a resting-state fMRI study. Sleep Med 13: 720–727. [DOI] [PubMed] [Google Scholar]
- 55. Katon W, Unutzer J, Russo J (2010) Major depression: the importance of clinical characteristics and treatment response to prognosis. Depress Anxiety 27: 19–26. [DOI] [PubMed] [Google Scholar]
- 56. Riedel M, Moller HJ, Obermeier M, Adli M, Bauer M, et al. (2011) Clinical predictors of response and remission in inpatients with depressive syndromes. J Affect Disord 133: 137–149. [DOI] [PubMed] [Google Scholar]
- 57. Holtzheimer PE, 3rd, Russo J, Claypoole KH, Roy-Byrne P, Avery DH (2004) Shorter duration of depressive episode may predict response to repetitive transcranial magnetic stimulation. Depress Anxiety 19: 24–30. [DOI] [PubMed] [Google Scholar]
- 58. Gildengers AG, Houck PR, Mulsant BH, Dew MA, Aizenstein HJ, et al. (2005) Trajectories of treatment response in late-life depression: psychosocial and clinical correlates. J Clin Psychopharmacol 25: S8–13. [DOI] [PubMed] [Google Scholar]
- 59. Anand A, Li Y, Wang Y, Wu J, Gao S, et al. (2005) Antidepressant effect on connectivity of the mood-regulating circuit: an FMRI study. Neuropsychopharmacology 30: 1334–1344. [DOI] [PubMed] [Google Scholar]
- 60. Fu CH, Williams SC, Brammer MJ, Suckling J, Kim J, et al. (2007) Neural responses to happy facial expressions in major depression following antidepressant treatment. Am J Psychiatry 164: 599–607. [DOI] [PubMed] [Google Scholar]
- 61. Miller GA, Chapman JP (2001) Misunderstanding analysis of covariance. J Abnorm Psychol 110: 40–48. [DOI] [PubMed] [Google Scholar]
- 62. Saad ZS, Gotts SJ, Murphy K, Chen G, Jo HJ, et al. (2012) Trouble at rest: how correlation patterns and group differences become distorted after global signal regression. Brain Connect 2: 25–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Current treatment details of patients with TRD.
(DOC)
The number of episodes of patients with TRD.
(DOC)
Significant VMHC differences between pooled patients and HS (p<0.005, GRF correction).
(DOC)
Significant VMHC differences between groups (uncorrected voxel-wise threshold of p<0.005).
(DOC)