

Abstract
There has been a significant body of literature on species flock definition but not so much about practical means to appraise them. We here apply the five criteria of Eastman and McCune for detecting species flocks in four taxonomic components of the benthic fauna of the Antarctic shelf: teleost fishes, crinoids (feather stars), echinoids (sea urchins) and crustacean arthropods. Practical limitations led us to prioritize the three historical criteria (endemicity, monophyly, species richness) over the two ecological ones (ecological diversity and habitat dominance). We propose a new protocol which includes an iterative fine-tuning of the monophyly and endemicity criteria in order to discover unsuspected flocks. As a result nine « full » species flocks (fulfilling the five criteria) are briefly described. Eight other flocks fit the three historical criteria but need to be further investigated from the ecological point of view (here called « core flocks »). The approach also shows that some candidate taxonomic components are no species flocks at all. The present study contradicts the paradigm that marine species flocks are rare. The hypothesis according to which the Antarctic shelf acts as a species flocks generator is supported, and the approach indicates paths for further ecological studies and may serve as a starting point to investigate the processes leading to flock-like patterning of biodiversity.
Introduction
Thanks to the International Polar Year (IPY) 2007–2009 and the Census of Antarctic Marine Life (CAML), the recent increase of oceanographic explorations across the Southern Ocean has confirmed the status of the Antarctic continental shelf as an area of rich marine biodiversity [1]–[4]. The richness of the benthic Antarctic fauna was known before [5]–[7] but considered as yet underexplored. Such a richness was observed for many benthic groups such as pycnogonids, ascidians and polychaetes [8], teleost fishes [9], echinoderms [10]–[11], crustaceans [7], poriferans and hydrozoans [12]. [8] reported that while some groups appear more diverse there than elsewhere in terms of species richness (some isopod lineages, pycnogonids, bryozoans, sponges, ascidians), others appear equally diverse (polychaetes, amphipods, echinoderms), and others less diverse (decapods, molluscs, teleosts). All these organisms are a boon for research concerning processes generating biodiversity because they have flourished within a relatively isolated area for long periods [13]–[16]. A substantial part of the marine Antarctic species richness might well be the result of species flocks, especially within the benthic fauna. Indeed, the Antarctic continental shelf has been described as a giant species flocks generator [17], as explained in more detail below. Species flocks are bursts of closely related endemic species which are ecologically diverse and numerous relatively to surrounding areas [18]. We can't observe those bursts directly, however, they are responsible for certain patterns in modern biodiversity. The first steps to study species flocks are therefore in the detection of these patterns, since knowledge of the present patterns of biodiversity is necessary prior to further exploration of the processes having generated it [16].
Eastman and McCune [17] noticed physical similarities in terms of isolation, depth and age, between the Antarctic shelf and ancient lakes where species flocks were found. Indeed currents, sub-zero temperatures and distance isolate the Antarctic shelf from all other shelves of the Southern Ocean. The Antarctic shelf is approximately 450 m deep in average [8], i.e. eight times deeper than the world shelves average, because of the weight of the ice sheet on the continent [19]. The age of the shelf geological isolation dates back at least 40 Ma, and the shelf has existed under polar conditions for 14–12 Myr [20]. Moreover, repeated advances and retreats of the ice sheet on the shelf probably caused benthic faunal extinctions [21] but also stimulated speciation events [22] through population fragmentation in isolated areas of the shelf or population displacement in refugia in sub-antarctic islands or in the deep sea [16], [23]. All these physical and historical factors are likely to have promoted species flocks at different times in a geographic area including the Southern Ocean (i.e. south of the Polar Front) and the sub-Antarctic islands. Therefore we can expect an important part of benthic biodiversity to exhibit the genetic, geographical and ecological patterns of species flocks at different taxonomic levels.
The aim of the present survey is to evaluate the hypothesis of a species flock origin among different groups of the Antarctic benthic fauna, using the criteria of Eastman and McCune [17] for species flocks detection. We mainly focused on the rich sampling of the French-Australian cruises CEAMARC (2007–2008) and the French cruises REVOLTA (2009–2012) in the Eastern Antarctic coastal waters to answer the following questions: how frequent are species flocks? How useful and suitable are the criteria used by Eastman and McCune to detect them? Consequently, is it necessary to revisite the original criteria to identify a flock? The project tries to approach these questions with four distantly related taxonomic groups of the benthic fauna: notothenioid teleost fishes, crinoids (feather stars), echinoids (sea urchins) and crustacean arthropods. Some other groups (e.g. molluscs or pycnogonids) will also be occasionally mentioned.
What is a Species Flock?
It is important to distinguish between the theoretical definition of a species flock and the criteria to identify or detect them using empirical data. As to theoretical definitions, Ribbink [18] defined a species flock as « an assemblage of a disproportionately high number, relative to surrounding areas, of closely related species which apparently evolved rapidly within a narrowly circumscribed area to which all the member species are endemic ». The definition is ambiguous because it mixes empirical criteria (like high number of species, endemicity) with conjectural processes that generate species flocks (« evolved rapidly »). The former are embedded into the practice of identification, the latter are not linked to direct practice but rather belong to a theoretical scheme. Ribbink [18] did not put emphasis on monophyly while it is essential for Greenwood [24]. Starting from both definitions of Ribbink [18] and Greenwood [24], Eastman and McCune [17] focused on criteria to detect species flocks and retained five criteria: monophyly, high species diversity (called « speciosity »), high level of endemism, morphological and ecological diversity, and habitat dominance (in terms of biomass). Some of these criteria imply to determine the historical and geographic pattern of biodiversity (monophyly, endemism, speciosity), whereas others rely on ecological studies (ecological diversity, habitat dominance). The first three criteria deal with space and time, the two others with present dynamic interactions. Each of these criteria has to be assessed for a set of species in comparison to its sister lineage of the surrounding areas, especially speciosity, endemism [25], and ecological diversification. In spite of potential difficulties in gathering sufficient reliable data for each of these criteria, it is nevertheless possible to recognize sets of species that clearly correspond to species flocks (for instance the Notothenioidei at the scale of the Southern Ocean, as convincingly proposed by Eastman and McCune [17], and sets of species that clearly do not correspond to flocks (liparid fishes of the Antarctic shelf, see below). The present study will detect such conspicuous cases, but will also evaluate more intermediate situations, potentially due to difficulties in applying the criteria.
Model Case
The standard case for the Southern Ocean has been described by Eastman and McCune [17]. Notothenioid fishes are monophyletic [26] at the scale of the Southern Ocean (including sub-Antarctic islands). Within this area they have a level of endemicity of 97%, which is exceptionally high for a marine group. The species diversity of notothenioids (134 species, according to Fishbase, [27] is very high with regard to other components of the ichthyofauna. Notothenioids represent at least 50% of it. The notothenioids are also morphologically and ecologically diversified [9], [28]. This group is benthic in origin, and secondarily diversified into niches in the water column, involving pelagic or partially pelagic zooplanktivory and piscivory [28]. The group contains benthic, epibenthic, cryopelagic and pelagic species, with morphological diversification associated to vertical motion in the water column and neutral buoyancy control rather than to diversification in trophic morphology [28]. Finally, a number of studies summarized by Eastman and McCune [17] have confirmed that notothenioids clearly dominate the fish biomass, of which they represent more than 90% [9]. The notothenioids are therefore described as a giant species flock at the scale of the whole Southern Ocean by Eastman and McCune [17], providing a model and a point of reference for our further comparisons. The question remains: can we recognize other benthic species flocks within the Southern Ocean? If some sets of species fail to meet all criteria, which are the failing criteria? How to apply the multiplicity of criteria by which we recognize a flock?
Methods
We propose to rank the criteria in order to discover species flocks at various geographic scales within a given taxonomic group. Because evolutionary processes leading to flocks are not known or not obvious at the first glance, putative flocks are primarily recognized through the study of patterns of biodiversity: speciosity is obtained through taxonomy, endemicity from geographical distribution and monophyly from character distribution (i.e. from phylogenies, either molecular or morphological). Then, from a heuristic point of view, a flock can be primarily seen as a set of patterns (taxonomic, geographical, phylogenetic) associated with ecological diversity. The two ecological criteria are also important, but require a larger number of studies to be properly evaluated: when these criteria do not appear to be fulfilled this can be an artifact, i.e. a lack of precise knowledge hampering their assessment. The priority given here to historical criteria is therefore more practical than based on theoretical precedence. Geographic and taxonomic range seem to be the easiest criteria to assess. The question is therefore the following: at which geographic-taxonomic scales will all criteria be satisfied in their entirety? Starting from a given group in a given area as a candidate to be a species flock, we may reduce the taxonomic range, or expand the geographic one to reach monophyly, high endemism and speciosity within realistic limits, for example, those of the Southern Ocean. This corresponds to the modulation loop in Fig. 1, leading to the identification (or rejection) of a flock. A similar approach may be applied to uncover other larger or smaller flocks related to the firstly identified one. This approach should facilitate the discovery of nested species flocks. For example, the fact that notothenioids are described as a giant species flock would not prevent discovery of smaller, more recent, subflocks within the group. The two other criteria, which are more difficult to assess, can then be evaluated for the group. We provisionnally propose to call « core flock » a potential species flock for which the first three geographical and historical criteria are fulfilled, and « full flock » a potential species flock for which all the five criteria are fulfilled. Some flocks will probably remain as « core flocks » either because at least one ecological criteria failed or because there is not enough available data to assess one of these two criteria. The first three criteria are the ones that can be evaluated the most consistently. The approach allows finding where (and if) they are reached, and in which set of species. This method of assessment is not an unselective means of finding flocks everywhere. Instead, as shown below, some components of the benthic fauna of the Antarctic shelf are definitively not species flocks.
Figure 1. Protocol to use the five criteria of Eastman and McCune [17] to detect and evaluate species flocks.
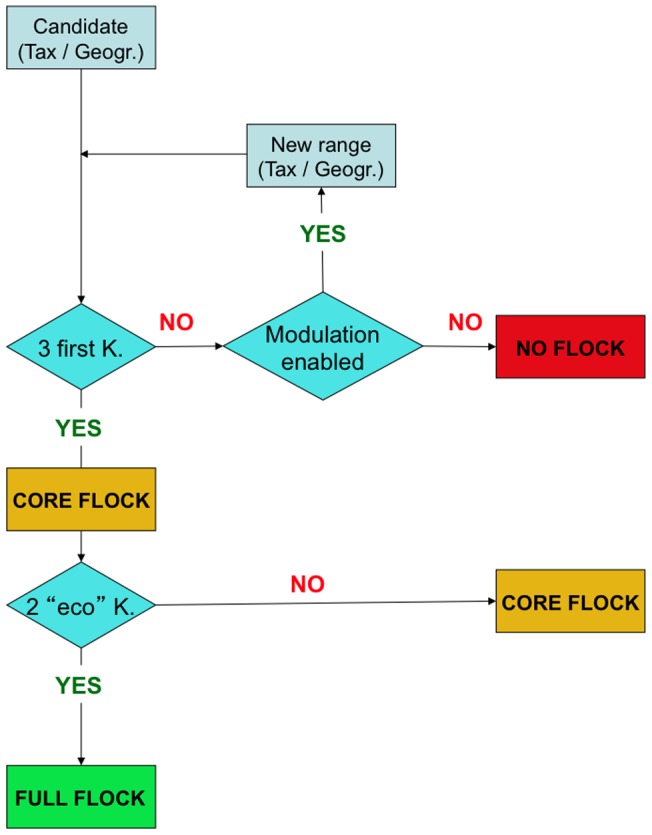
The first three criteria (3 K) are the species diversity (« speciosity ») of the taxonomic component, its level of endemicity, and its monophyly. The two other criteria (2 K) are habitat dominance (in biomass) and ecological diversity. The modulation loop means that the geographical range and the taxonomic rank may have to be redefined in order to discover unsuspected flocks.
Fig. 1 depicts a flow chart showing the two-step protocol used to detect species flocks. The three « historical-geographical » criteria (monophyly, endemism, speciosity) are the first ones to be assessed. It means that a failure to fulfill one of those three eliminates the taxon as a potential flock. A loop modulates the geographical range and the taxonomic rank to discover new flocks at not yet investigated levels. The two « ecological » criteria (morphological-ecological diversity, domination of habitat in terms of biomass) are considered afterwards. Results are summarized in Fig. 2.
Figure 2. Results on species flocks estimation concerning four taxonomic components: teleosts, echinoids, crinoids, crustaceans.

Amphi: amphipods, SO: Southern Ocean, AS: Antarctic Shelf, ©: newly discovered cryptic species are to be added.
Results
True Species Flocks (or « full » flocks)
The ecological and morphological diversification of the Nototheniidae, the most speciose notothenioid family (49 species) is very high and well documented [28], and the habitat domination at the scale of the Southern Ocean is not in doubt [9]. Endemicity is above 97%. However, the monophyly of the family, repeatedly considered as uncertain [29]–[32], has recently been seriously challenged [33]: four other families are embedded within it (Harpagiferidae, Artedidraconidae, Bathydraconidae and Channichthyidae). As a result, the « nototheniid flock » could actually be the flock of notothenioids with anti-freeze glycoproteins (AFGPs), i.e. notothenioids less the 13 notothenioid species of the subantarctic basal families Bovichtidae (11 species), Pseudaphritidae (1 species) and Eleginopsidae (1 species) [9], [32]. These proteins are sometimes seen as the key-innovation responsible for the sudden burst of diversification [22], [26], [34], but see [35]. In summary, there is evidence for a species flock here, but this flock should no longer be termed the « nototheniid flock », but rather the flock of the monophyletic group of notothenioids with the antifreeze glycoproteins (the « AFGP bearing notothenioids », Fig. 2, line 2). This species flock contains at least three nested flocks: the Trematominae, the Channichthyidae and the Artedidraconidae, which are detailed below.
With 13 species all endemic to the continental shelf and to some peri-insular plateaus of the Southern Ocean, the subfamily Trematominae (Teleostei, Notothenioidei, Nototheniidae) contains 10% of the notothenioids. However it is possibly underestimated because several species exhibit intra-specific chromosomal variability according to the geographic sector and sometimes within the same sector [36]–[37]. Their chromosome numbers and structures exhibit the highest diversity among the notothenioid clades. Chromosome diploid numbers and formulae differ from one species/population to another, except for Trematomus loennbergii which shows a highly polymorphic, unstable karyotype. They range from 2n = 24 (T. eulepidotus) to 2n = 58 (T. nicolai) and change according to Robertsonian fusion or fission events (reviewed in [37]). The group is monophyletic [30]–[31], [35], [38]. There is a noticeable degree of ecological diversity that does not reflect the phylogeny, i.e. niche changes do not appear to come from common ancestry but rather occur several times independently [31], [39]–[41]. The Trematoninae represent an important part of the biomass of coastal ichthyofauna [42]–[44]. Moreover they correspond to a sudden burst of diversification (to the exclusion of Trematomus scotti which is the sister-group of the rest of the subfamily) that occurred some 10 Ma [41], [45], though this feature is not among the criteria of Eastman and McCune [17]. Therefore the Trematominae can be considered as a smaller and more recent flock restricted to the Antarctic shelf and a few peri-insular shelves (e.g. South Georgia and South Sandwich islands [46].
The icefishes (family Channichthyidae, Notothenioidei) are endemic, monophyletic [32], [47] and display some degree of speciosity (16 species [48]) with regard to related families. They represent a very important part (more than 25%) of the Antarctic fish biomass [9]. They exhibit a noticeable ecological diversity [28], [49] linked to the ability to feed in the water column. Beyond these fulfilled criteria, it is also interesting to notice that their phyletic diversification seems to have occurred rapidly [35].
Artedidraconidae (plunderfishes) are speciose (30 species), monophyletic [29]–[30], [47] and endemic to the Southern Ocean. They show some degree of ecological diversification [50], though they need to be studied further, and possess only modest morphological diversity [9]. They are provisionally considered as a species flock [51].
The peracarid Crustacea is the most speciose animal group of the Southern Ocean, with more than 1000 strictly Antarctic species. Among them, amphipods are the most diverse, comprising 919 species in the Southern Ocean, and 547 species from the Antarctic region only (south of the Antarctic Polar Front), of which 417 species, or about 70%, are endemic [52]–[54]. In this environment, they have colonised a wide variety of ecological niches and achieved a successful eco-ethological diversification [55]. The genera Eusirus and Epimeria, though present elsewhere, each have components in the Southern Ocean that are monophyletic, given the present state of knowledge, and meet the five criteria for a species flock. A phylogenetic analysis of Antarctic Epimeria species confirmed a monophyletic assemblage of 26 Antarctic species and placed two potentially closely related non-Antarctic species (from New Zealand) as closest relatives [56]. Epimeria species are large, heavy, highly calcified and almost entirely benthic animals, with low mobility. They display a high diversity in morphology and trophic types. It is possible that the biogeographic range of Epimeria reflects its tolerance to a limited range of temperature, or cold stenothermy, which would provide an important clue as to its potential isolation in Antarctica from other parts of the world. To date, Epimeria provides the best example of a species flock within Antarctic amphipods. Eusirus spp. are medium- to large-sized predatory amphipods [57] with good swimming capacities [58], found between 0 and >7000 m [59] with benthic, pelagic or sympagic life styles [60]–[61]. Barnard and Karaman [59] listed 22 nominal species, of which seven have been recorded south of the Antarctic convergence. However, ongoing research indicates the existence of at least 23 described and undescribed species in the Southern Ocean [62]–[63]. The species of the group perdentatus which are particularly large Eusirus (60–100 mm), are found only south of the convergence; they include three named species and at least three multiple cryptic/pseudocryptic species [62]–[63]. The absence of dispersal across the Polar Front of the group perdentatus could be explained by their gigantism. It has been demonstrated that giant amphipods are restricted to waters with a maximum oxygen concentration, i.e. in truly icy waters [64]. Furthermore, all these species form a clade, which is geologically young (4–14 Ma) [63]. Ecological differentiation is present in this group, with most species benthic or benthopelagic, except for E. propeperdentatus which is entirely pelagic. Even though this group is only moderately diverse, it satisfies the criteria of a flock.
Although endemicity and speciosity of several Antarctic isopod lineages is quite high, few of these meet all five criteria of species flocks. One of them is the Serolidae whose Antarctic component is monophyletic and comprises about half of the known serolid species [65]. Serolid isopods exhibit a wide variety of habitat use and lifestyles. They are known from the shallow waters to the deep sea, have semi-sessile to highly mobile rafting species [66], and inhabit soft-sediment and rocky bottoms. Antarctic serolids thus also fulfill the two ecological criteria making them full species flocks whereas many subordinate clades nested within the isopods remain core flocks due to many, often pseudo-cryptic species, which are only weakly differentiated ecologically. As the rate of discovery of new species through the use of molecular methods increases, it is expected that in the future more flocks at lower taxonomic levels will be described which currently violate the speciosity requirement only due to a lack of knowledge.
Core Flocks, but not « Full » Flocks
Some Antarctic isopod taxa appear to have radiated on the Antarctic shelf, and molecular studies have proven useful in demonstrating the presence of many morphologically similar, but genetically highly differentiated species [67]. The species inside these species complexes are monophyletic and can be locally very abundant but are generally only poorly differentiated in ecological terms, making them core but not full flocks (Fig. 2, e.g. Ceratoserolis trilobitoides, C. meridionalis and Glyptonotus antarcticus species complexes). Similarly species complexes are found using molecular tools in pycnogonids (Colossendeis megalonyx sensu lato [68]), which are not ecologically diverse. In other cases a lack of knowledge with regard to ecological diversity (like in the radiation of the gastropod Doris kerguelenensis, [69]) leads to the provisional conclusion of a “core flock” but not a “full flock”.
Echinoid taxonomic components fail to meet the five criteria, most of them lacking either ecological diversity or domination of habitat (Fig. 2). Two clades of echinoids fall within this category of « core flocks ». The Antarctic schizasterids consist of 27 morphologically recognized species of sea urchins. Over all a total of about 80 echinoid species have been recorded in the Southern Ocean so far, and they represent one third of the Antarctic echinoid speciosity. Molecular analyses based on several molecular markers confirm that the brooding species within this group of irregular sea urchins form a monophyletic group restricted to the Southern Ocean but not to the Antarctic shelf, some species being found in the Kerguelen Islands, Heard Island, and the southernmost part of South America. All species of the monophyletic brooding group are Subantarctic or Antarctic. Some species can locally dominate the echinoid fauna in term of abundance [70], but they display a limited ecological diversity (all are infaunal or partly infaunal deposit feeders) and consequently are not considered as a true flock. A recent population genetic study [71] confirms that the brooding schizasterids display highly reduced dispersal rates, a feature which may favour high speciation rates [72], independently of any key innovation.
In the regular echinoid family Cidaridae, the Ctenocidarinae form a monophyletic group of 21 morphologically recognized species that are restricted to the Southern Ocean [10], [73]. The genus Austrocidaris is found elsewhere in the Southern Ocean, but except for this genus, the Ctenocidarinae (without Austrocidaris) form a monophyletic group restricted to the Antarctic shelf. They are ecologically and morphologically well diversified. Some species are brooders while others are not, and their morphologically diverse primary spines are covered with numerous and various specific symbionts [74]–[75]. At both scales (Southern Ocean and Antarctic shelf), however, these two cidarid embedded groups of sea-urchins fail to be significant in terms of biomass, but this remains to be evaluated with more precision. Therefore, it is prudent to consider them as « core flocks », and not « full » species flocks.
We record as a « core flock » the Pogonophryne artedidraconid fish subgroup of 22 species because it is a remarkable example of a phyletic radiation without ecological or morphological diversification [9]. The morphology is so characteristic and, with the exception of the barbel, constant that the taxonomy of the group is the most difficult of all notothenioids. Their chromosome numbers and formulae are stable [76]–[78], whilst in other artedidraconids, they show some important structural interspecific changes. There has been sufficient collecting to indicate that their population densities and biomass are low: they do not exhibit habitat domination. However Pogonophryne are about 2-fold more diverse than any other Antarctic notothenioid genus, all having circum-Antarctic distributions and recently discovered species are coming from upper slope waters 1000–2000 m deep. So Pogonophryne is a large, recent, strictly benthic, non-adaptive (little morphological and ecological diversity) radiation, at a depth not colonized by most other clades of notothenioids. This is an interesting phenomenon contrasting, for example, with the adaptive radiation of trematomines on the shelf.
Criteria for a flock not met
For crinoids, the criterion of species diversity fails and ecological diversity is often poorly documented. The two widespread and most abundant species Promachocrinus kerguelensis and Florometra mawsoni together form a monophyletic group (case 1 in Table 1, Fig. 3) distinct from other Antarctic Heliometrinae [79]–[80]. These species are not restricted to the continental shelf but are found throughout the Southern Ocean [80]–[82]. Both of them show a large morphological variability [79], [81] and live in a great diversity of habitats [79]. Promachocrinus kerguelensis is composed of at least seven genetic circum-Antarctic lineages [83]–[84]. However these lineages represent two seperate species at best [84]. The species diversity criterion for a flock is therefore not met. Species from the genus Notocrinus are dominant among brooding crinoid species [80]. This genus is monophyletic and endemic to the Southern Ocean, from the continental shelf to Burdwood Bank [80]. Molecular results indicate that the genus Notocrinus is composed of more than the two morphologically known species: two cryptic species within N. mortenseni and four cryptic species within N. virilis [80] are suggested. Species diversity in Notocrinus still needs to be precisely assessed and the present lack of knowledge prevents the assignment of this group to a species flock.
Table 1. Three situations for the taxon of reference (columns) are to be considered: it is monophyletic, paraphyletic or polyphyletic.
| Members of the taxon present in the considered area: | Taxon: | ||
| Monophyletic | Paraphyletic | Polyphyletic | |
| Monophyletic | Flock | Case 1 | No |
| Paraphyletic | Case 2 | Case 3 | No |
| Polyphyletic | No | No | No |
There are also three situations to consider for the components of this taxon in the given area of reference (lines): monophyletic, paraphyletic or polyphyletic. To explain the table, we consider again the example of the non-notothenioid fish family Liparidae. It is monophyletic as a family (first column), however its components of the Antarctic shelf are polyphyletic because they are each related to Arctic liparids (bottom line). So the Antarctic liparid situation is the bottom left cell. Case 2 (when a taxon originating in the area of reference secondarily “exports” a part of its descent outside this area, Fig. 4) is discussed in the text.
Figure 3. Case 1 of Table 1.
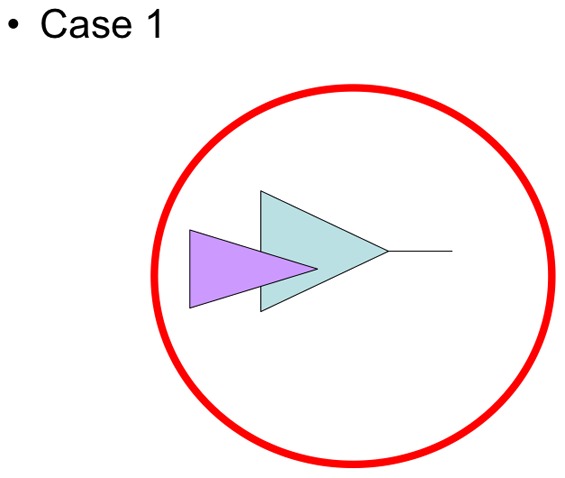
A set of species is paraphyletic (blue) with a taxonomic entity embedded within (purple) it that is restricted to the area of reference (red circle): A simple taxonomic decision could fulfill the two criteria of monophyly and endemicity. Indeed the taxonomic decision would render the whole set of species monophyletic (purple becomes blue).
Harpagiferids (Teleostei, Notothenioidei) are a good example of a monophyletic family not corresponding to a flock: they consist of ten species of ecologically similar fishes [9]. The non-notothenioid family Liparidae (Teleostei, Cottoidei) also does not satisfy the criteria, because of the absence of monophyly of its components inhabiting the Antarctic shelf (bottom left in Table 1). Indeed, some species of the Antarctic shelf are more related to Arctic liparids than to other liparids of the Antarctic shelf [85]–[86] (but see page 116 in [87] who considers some subgroups of Antarctic liparids as probably forming flocks).
None of the decapod or euphausiid crustaceans of the Southern Ocean studied during this survey (Notocangron, Euphausia respectively) are species flocks: the monophyletic components are not speciose and do not include cryptic species, though they are very important in terms of biomass (for instance the krill species Euphausia superba and E. crystallorophias). This also applies to other eucarid taxa studied to date (Nematocarcinus and Chorismus [88]).
The peracarid taxa Liljeborgia and Orchomene sensu lato are also not species flocks. Liljeborgia spp. are medium-sized benthic amphipod crustaceans able to swim short distances [89] and are opportunistic feeders [90], recorded between 0 and 6000 m [59]. Of the 67 nominal species of Liljeborgia [89], [91], 24 are known from the cold parts of the Southern Hemisphere. Of these 24, 12 are only known south of the Antarctic Convergence and three are found on both sides of the convergence [92]–[93]. A high diversity of a genus in a region can be considered as an indication of the possible existence of a local flock. However, both the examination of morphological characters and molecular data indicate that Antarctic species do not form a clade ([92], bottom left in Table 1), and suggests that they have relatives in distant seas like the Norwegian Sea [89], [93]. In other words, it is an example of a diverse Antarctic taxon, which does not fulfill the monophyly criterion. Another example of a diverse Antarctic amphipod taxon, which does not comply with the monophyly criterion, is provided by the Antarctic species of the lysianassoid genus complex Orchomene sensu lato. The taxon initially represented a possible candidate for a species flock in the Southern Ocean, due to its relative species diversity and high degree of endemism. This genus complex harbours at least 28 endemic, valid species in the Southern Ocean, belonging to 5 different genera. Havermans et al. [94] identified three species new to science and four species complexes each consisting of at least two cryptic species. Furthermore, d'Udekem and Havermans [95] included one more undescribed species, so that 36 potential endemic species are present in the Southern Ocean sensu lato, with a rate of 39% of endemicity in a total of 93 species. The Antarctic species of Orchomene sensu lato also exemplify habitat dominance (shelf and abyssal depths) and a significant diversity in trophic adaptations, being opportunistic or exclusive scavengers, with corresponding modifications in their mouthpart morphology [60], [96]. Furthermore, there are indications of a recent and rapid diversification of the Antarctic component of Orchomene sensu lato ([94]). This group of Antarctic species was initially thought to be monophyletic [96] but further studies from the same team, including more non-Antarctic species, show that this monophyletic orchomenid clade also comprised strictly Atlantic and Magellan species. However, among a majority of species restricted to the Southern Ocean, the Atlantic and Magellan taxa have a derived, more apical position in the phylogeny, which might suggest their origin within the Southern Ocean (case 2 in Table 1, Fig. 4).
Figure 4. Case 2 of Table 1.
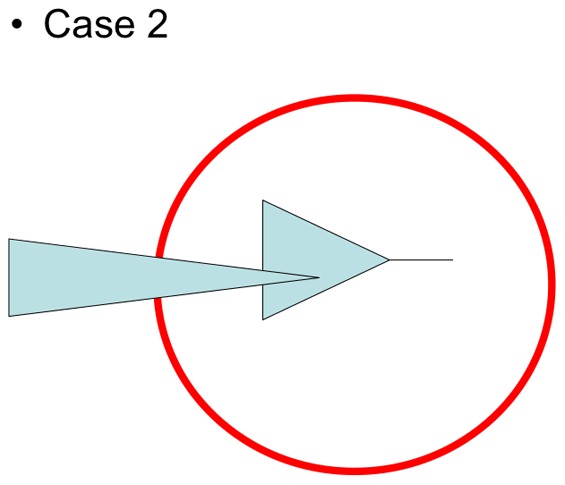
The set of species under focus is monophyletic but contains an internal subpart that is secondarily « exported » outside the area of reference (red circle). See text for discussion.
In the genus Sterechinus, which is a regular sea urchin, a monophyletic group is obtained when the genus Dermechinus is included into the clade. This group is mainly restricted to the Southern Ocean, but some representatives are present both in the Antarctic shelf and in South America as well as in the Kerguelen and other subantarctic regions [97], likely due to dispersal via planktotrophic larvae. The number of species in the clade “Sterechinus + Dermechinus” (case 1 in Table 1, Fig. 3) is rather limited (weak speciosity). There are half a dozen morphological species, but molecular data (mostly mitochondrial, complemented by recent nuclear data) suggest only three to four genetic species [98], and reveal a lack of congruence between genetics and geography; monophyletic groups of species (or even of haplotypes within species) do not correspond to geographical regions.
Discussion
Complex cases of « Nested Flocks » and « Exportations »
In some cases, when phylogenies are available, we detect an acceleration of cladogeneses for more internal subsets of the flock. These subsets may be regarded as nested flocks. For instance, within notothenioids, AFGP-bearing species constitute a nested flock probably allowed by the key-innovation of antifreeze glycoproteins available in the blood [22], [26], [35]. In molecular phylogenies this acceleration of cladogeneses corresponds to a weak resolution (or no resolution at all) in the branching patterns among the main nototheniid lineages with regard to the crown group [31]–[32], [35], [38]. Another case is illustrated by the subfamily Trematominae that contains a diversified set of 13 species of coastal fishes, which exhibit a later acceleration of cladogeneses in the sister-group of Trematomus scotti [41], [45]. However, despite of the fact that most of the flocks described exhibit a period of rapid cladogeneses (soft polytomy, [99]) or a simultaneous diversification (hard polytomy, [99]), the rate of diversification is not included among the criteria of Eastman and McCune [17], while it is in Ribbink [18]. Here nested flocks show situations where there is an acceleration (when studied): non-Austrocidaris ctenocidarins within the Ctenocidarinae, Trematominae within the notothenioids [35], [41], possibly Artedidraconidae and Channichthyidae within notothenioids [35].
When using the criteria in order to detect species flocks, a diversity of situations appear which are liste in Table 1. When a set of species is monophyletic within the area of reference to which it is endemic, two criteria of species flocks are fulfilled (top left in Table 1). For instance, this is the case for the Antarctic component of the amphipod genus Epimeria. When a set of species is monophyletic but its members in the area of reference are polyphyletic, i.e. not closely related to each other, the monophyly criterion fails and there is no species flock (bottom left in Table 1). For instance, this is the case for liparid fishes of the Antarctic shelf: liparids are monophyletic as a family but its Antarctic members are polyphyletic because each of their diverse components is directly related to Arctic liparids. Let's consider the case 1 of Table 1 (top middle cell): when a set of species is paraphyletic (Fig. 3, blue) with a taxonomic entity embedded within (Fig. 3, purple) it that is restricted to the area of reference (Fig. 3, red circle), a simple taxonomic decision could fulfill the two criteria of monophyly and endemicity. Indeed the taxonomic decision would render the whole set of species monophyletic (purple becomes blue). This is the case for the crinoids Promachocrinus kerguelensis, into which the crinoids of the genus Florometra are embedded. Renaming Florometra as Promachocrinus would render that genus monophyletic.
These situations are clearcut (italics in Table 1). Other situations are more complex. In the case 2 of Table 1 (middle left cell), the set of species under focus is monophyletic but contains an internal subpart that is secondarily « exported » outside the area of reference (Fig. 4). That pattern corresponds to the amphipod genus Orchomene sensu lato which Antarctic component contains an Atlantic subpart and the criterion of endemicity fails. However, viewed through time, the diversification could result from a two phases historical process: an initial burst of species and ecological diversity in a restricted area (i.e. the actual species flock), followed by an expansion of derived members of the flock outside this area. In such a case the species flock status could be maintained only when the rapid diversification is documented (for instance as in [41]). However, in the precise case of the Antarctic Orchomene sensu lato, the tempo of diversification remains to be investigated. Case 3 (middle cell of Table 1, Fig. 5) is a mix of case 1 and case 2. A taxonomic decision would simply lead to case 2, where supplementary data about the tempo of diversification would then be required.
Figure 5. Case 3 of Table 1.
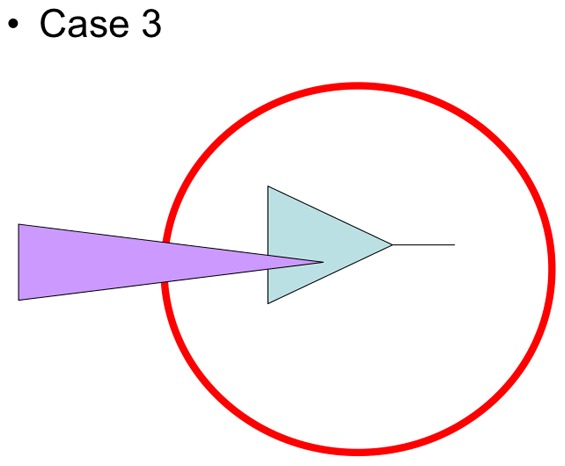
This situation is a mix of case 1 and case 2. A taxonomic decision would simply lead to case 2, where supplementary data about the tempo of diversification would then be required (see text).
Facing the case 2 of Table 1 (Fig. 4), two situations can be distinguished depending on the degree of the exportation. First, the status of species flock can be maintained by expanding the area of reference, all other criteria being satisfied, as long as the expanded new area remains within certain limits, consistent with the geographic criterion. This is an easy way, but the appraisal of a realistic expansion is a matter of empirism. In the present survey, we defined the Southern Ocean as the maximal area. For example, the icefishes (Channichthyidae) fit the definition of a species flock on the Antarctic continental shelf. Biogeography mapped onto their phylogeny (that is rather well resolved [47]–[49]) clearly shows that this benthic family originates on the shelf. However, some species are found outside the shelf (e.g. Channichthys rhinoceratus and Champsocephalus gunnari in Kerguelen Islands, Chaenocephalus aceratus in Bouvet Island, Champsocephalus esox in Magellan Strait). Consequently, the Channichthyidae is considered a species flock of the Southern Ocean.
The second way to answer is to rely on the history of the events. This is the case for deep-sea serolids of the Southern Hemisphere. Some of them occur outside the Southern Ocean (e.g. Acutiserolis spp. in [65]), but the molecular phylogeny places all of them firmly inside the radiation that took place in the shallow waters close to the Antarctic continent. This suggests that, subsequent to Antarctic speciations, the involved species had expanded their distribution area northward, rather than having arisen outside the Southern Ocean. This is also the case of the lysianassoid genus complex Orchomene sensu lato which clearly originates within the Southern Ocean, but has Atlantic and Magellan components well embedded within the Southern Ocean clade. These examples emphasize the importance of (1) reliable large-scale samplings and (2) historical information derived from phylogenetic trees. If the decision had rested exclusively on a geographic description of where the species occurred today, the monophly criterion would have been mistakenly violated even though the major evolutionary event (a major radiation occurring inside a narrowly circumscribed area) would have been true. Because a species flock is also defined by Ribbink [18] as a historical evolutionary event characterized by rapid species and ecological diversifications, the possibility for secondary exportation of some components outside the area from which it originates should be left open. This requires an early rapid diversification of the flock within the original area. Indeed the « flock » categorization within the initial geographic realm would be maintained only if we have precise information about the tempo of diversification: the « exported » subpart must originate crownward in the phylogeny of the flock, and phylogenies must exhibit early acceleration of cladogeneses at the base of the flock within the area of reference (soft polytomies being possibly due to unappropriate genetic markers or unsufficient data, the check for hard polytomies can be done using several markers separately). For example, the Antarctic species of Orchomene sensu lato are thought to have undergone a rapid and recent diversification [94] and the AFGP-bearing notothenioids exhibit a rapid tempo of diversification at the base of their tree [32]–[33], [35]. Among them, the genus Patagonotothen comprises 15 species branched crownwards, of which 14 have secondarily colonized northwards to the southern coasts of South America and the Falkland Islands [9]. Therefore, by recognizing notothenioids as a giant species flock, Eastman and McCune [17] already implicitly tolerated those secondary exportations. It is obvious that taking into account this phylogenetic-historical supplementary criterion, it may possibly contradict the initial criterion of endemicity. This discrepency is due to the fact that, even if a species flock is defined as a phyletic radiation with ecological diversification, the five criteria of Eastman and McCune [17] do not incorporate the radiation, i.e. do not estimate the tempo of phyletic diversification. This is mostly due to the fact that the species flock concept gets more complex when passing from a typological application to its historical interpretation. It is basically not a typological concept, but a historical one. The present work could lead to the recognition of a sixth criterion, fully recognizing the species flock concept as a historical one in the core of its application, the high rate of phyletic diversification, to be modulated with the criterion of endemicity, which consists in tolerating «secondary exportations » outside the area of reference.
An amended detection of flocks
Eastman and McCune [17] noticed that the identification of species flocks in the marine realm had received less attention than in freshwater lakes or in islands. The present study fills the gap. Our practice of Antarctic and subantarctic species flocks detection in various taxonomic groups is based on multidisciplinary practical experience (joining taxonomic expertise including morphology and anatomy, field work, molecular phylogenetics and cytogenetics, field ecology) that leads us to propose an improvement of species flock recognition, aiming to make it more operational and instrumental. First, it appears necessary to focus on the three robust, easier to determine criteria: monophyly, endemism, and speciosity. We recommend ranking the ecological criteria as secondary, hence suggesting the distinction between “core” flocks and “full” flocks. No criterion is actually easy to determine: species diversity is a continuous parameter that must be compared to species diversity of the sister-group and surrounding areas and assessment of endemism and monophyly heavily depend on reliable field samplings [25]. Species flocks may be underestimated because of a lack of detection of cryptic species, but they can be overestimated through overestimation of endemism due to uncomplete field sampling. Ecological criteria are the most difficult to document, because they require even more data to be robustly appraised, and because these data are in themselves complex to obtain in the field. Last but not least, there is a component of arbitrariness in the spatial and taxonomic delineation in the estimation of « habitat dominance » or the « ecological diversity ». For instance, notothenioids represent 90% of the fish biomass of the Antarctic shelf: however it is negligible with regard to the whole eukayote biomass. « Habitat dominance » depends on the arbitrary taxonomic realm of « fishes », a non-monophyletic group. Finally, in order to have more flexibility and to assess the robustness of flocks, we suggest introducing the possibility to tune up the spatial or the taxonomic ranges in order to meet the endemism/monophyly criteria respectively. The difficulty is to maintain the adjustments within realistic limits. Those limits can be set by logical comparisons with surrounding areas as originally suggested by Ribbink [18], but so far there is no more than a rule of thumb. The confrontation of Eastman and McCune's criteria with real organisms encountered in the field compelled us to assign priorities. The flow chart of Fig. 1 should be regarded as a guide in the process leading to flock identification. It operates as a prioritized protocol, as well as a synthesis, of our practical approach to flocks. However this flow chart does not yet encompass an aspect of Ribbink's definition of a species flock: the fact that the initial phyletic diversification occurred « rapidly ». Despite not being a criterion retained by Eastman and McCune, it could be an important, albeit difficult to obtain, complementary information.
Acknowledgments
We thank the crew and participants of the cruises involved in the capture of the samples, and especially the chief scientists of the CAML-CEAMARC cruises: G. Hosie, T. Ishimaru, M. Riddle, M. Stoddart. The CAML-CEAMARC cruises of R/V “Aurora Australis” and R/V “Umitaka maru” (IPY project n°53) were supported by the Australian Antarctic Division, the Japanese Science Foundation, the French polar institute “Institut Paul-Emile Victor” (and the IPEV programs 281 “ICOTA” co-directed by Catherine Ozouf-Costaz and Philippe Koubbi and 1124 “REVOLTA” directed by Guillaume Lecointre). The present work was also partly based on samples collected during the ICEFISH 2004 cruise (supported by National Science Foundation grant OPP 01–32032 to H. William Detrich (Northestern University) and the EPOS cruise (1989) for which we are grateful to the Alfred Wegener Institute and the European Science Foundation.
Funding Statement
This work was supported by the Centre national pour la Recherche scientifique (CNRS), the Muséum national d'Histoire naturelle (MNHN) and the Agence nationale pour la Recherche (ANR, grant number “ANTFLOCKS”, USAR 07-BLAN-0213-01 to G.L.), the “Service de Systématique Moléculaire” of the Muséum national d'Histoire naturelle (UMS 2700 CNRS), the “Consortium national de Recherche en Génomique”: it is part of the agreement number 2005/67 between the Genoscope and the Muséum national d'Histoire naturelle on the project ‘Macrophylogeny of life’ directed by G.L. This work was further supported by DFG project SCHU-1460-8 to C.S. and C.He., the Belgian Science Policy (“Action I” grant number MO/36/022 to C.U.A., “Action II” grant number WI/36/H04 to C.Ha., “PADDII” grant number SD/BA/02B to C.D.R.). This project is a contribution to the EBA-SCAR program. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Griffiths HJ (2010) Antarctic Marine Biodiversity – What Do We Know About the Distribution of Life in the Southern Ocean? PLoS One 5. doi:10.1371/journal.pone.0011683. [DOI] [PMC free article] [PubMed]
- 2. Grant RA, Griffiths HJ, Steinke D, Wadley V, Linse K (2011) Antarctic DNA barcoding; a drop in the ocean? Polar Biol 34: 775–780. [Google Scholar]
- 3. De Broyer C, Danis B (2011) How many species in the Southern Ocean? Towards a dynamic inventory of the Antarctic marine species. Deep Sea Res Part 2 Top Stud Oceanogr 58(1–2): 5–17 doi:dx.doi.org/10.1016/j.dsr2.2010.10.007 [Google Scholar]
- 4.Brandt A, De Broyer C, Ebbe B, Ellingsen KE, Gooday AJ, et al.. (2012) Southern Ocean Deep Benthic Biodiversity. Antarctic Ecosystems: An Extreme Environment in a Changing World (ed. by A. D. Rogers, N. M. Johnson, E. J. Murphy, and A. Clark), 291–334. Wiley-Blackwell, Oxford.
- 5. Starmans A, Gutt J (2002) Mega-epibenthic diversity: a polar comparison. Mar Ecol Prog Ser 225: 45–52. [Google Scholar]
- 6. Clarke A, Johnston NM (2003) Antarctic marine benthic diversity. Oceanogr Mar Biol Ann Rev 41: 47–114. [Google Scholar]
- 7. Brandt A, Gooday AJ, Brandão SN, Brix S, Brökeland W, et al. (2007) First insights into the biodiversity and biogeography of the Southern Ocean deep sea. Nature 447: 307–311. [DOI] [PubMed] [Google Scholar]
- 8. Aronson RB, Thatje S, Clarke A, Peck LS, Blake DB, et al. (2007) Climate Change and Invasibility of the Antarctic Benthos. Annu Rev Ecol Evol Syst 38: 129–154. [Google Scholar]
- 9. Eastman JT (2005) The nature of the diversity of Antarctic fishes. Polar Biol 28: 93–107. [Google Scholar]
- 10.David B, Choné T, Mooi R, De Ridder C (2005a) Antarctic Echinoidea. Synopses of the Antarctic benthos vol 10 (ed. by J. W. Wägele, and J. Sieg). Koeltz Scientific books Publishing, Königstein.
- 11. David B, Choné T, Festeau A, Mooi R, De Ridder C (2005b) Biodiversity of Antarctic echinoids: a comprehensive and interactive database. Sci Mar 69: 201–203. [Google Scholar]
- 12. Ragua-Gil JM, Gutt J, Clarke A, Arntz WE (2004) Antarctic shallow-water mega-epibenthos: shaped by circumpolar dispersion or local conditions? Mar Biol 144: 29–40. [Google Scholar]
- 13. Clarke A, Barnes DKA, Hodgson DA (2005) How isolated is Antarctica? Trends Ecol Evol 20(1): 1–3. [DOI] [PubMed] [Google Scholar]
- 14.Clarke A, Crame JA (1989) The origin of the Southern Ocean marine fauna. Origins and Evolution of the Antarctic Biota (ed. by J. A. Crame), 253–268. Geol Soc Spec Pub 47, London.
- 15.Clarke A, Crame JA (1997) Diversity, latitude and time: patterns in the shallow sea. In: R.G.F Ormond, J.D Gage, and M.V Angel, editors. Marine Biodiversity: causes and consequences. Cambridge: Cambridge Univ. Press. p. 122–147.
- 16. Clarke A, Crame JA (2010) Evolutionary dynamics at high latitudes: speciation and extinction in polar marine faunas. Philos Trans R Soc Lond B Biol Sci 365(1558): 3655–3666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Eastman JT, McCune AR (2000) Fishes on the Antarctic continental shelf: evolution of a marine species flock? J Fish Biol 57: 84–102. [Google Scholar]
- 18.Ribbink AJ (1984) Is the species flock concept tenable? Evolution of Fish Species Flocks (ed. by A. A. Echelle, and I. Kornfield), 21–25.University of Maine at Orono Press, Orono.
- 19.Anderson JB (1999) Antarctic Marine Geology. Cambridge: Cambridge Univ Press. 289 p.
- 20.Kennett JP (1982) Marine Geology. Prentice-Hall, Englewood Cliffs.
- 21.Krug AZ, Jablonski D, Roy K, Beu AG (2010) Differential Extinction and the Contrasting Structure of Polar Marine Faunas. PLoS One 5. [DOI] [PMC free article] [PubMed]
- 22. Patarnello T, Verde C, Prisco G di, Bargelloni L, Zane L (2011) How will fish that evolved at constant sub-zero temperatures cope with global warming? Notothenioids as a case study. Bioessays 33: 260–268. [DOI] [PubMed] [Google Scholar]
- 23. Thatje S, Hillenbrand CD, Larter R (2005) On the origin of Antarctic marine benthic community structure. Trends Ecol Evol 20: 534–540. [DOI] [PubMed] [Google Scholar]
- 24.Greenwood PH (1984) What is a species flock? Evolution of Fish Species Flocks (ed. by A. A. Echelle, and I. Kornfield), 13–19. editors. Orono Press, University of Maine, Orono.
- 25. Castelin M, Puillandre N, Lozouet P, Sysoev A, Richer de Forges B, et al. (2011) Molluskan species richness and endemism on New Caledonian seamounts: Are they enhanced compared to adjacent slopes? Deep Sea Res Part 1 Oceanogr Res Pap 58: 637–646. [Google Scholar]
- 26.Matschiner M, Haner R, Salzburger W (2011) On the Origin and trigger of the Notothenioid Adaptive Radiation. PLoS One 6. doi:10.1371/journal.pone.0018911. [DOI] [PMC free article] [PubMed]
- 27.Froese R, Pauly D (2011) FishBase. World Wide Web electronic publication. Available: http://www.fishbase.org.
- 28.Eastman JT (1993) Antarctic Fish Biology. Academic Press, San Diego.
- 29. Bargelloni L, Marcato S, Zane L, Patarnello T (2000) Mitochondrial phylogeny of Notothenioids: a molecular approach to antarctic fish evolution and biogeography. Syst Biol 49(1): 114–129. [DOI] [PubMed] [Google Scholar]
- 30. Near TJ, Pesavento JJ, Cheng CC (2004) Phylogenetic investigations of Antarctic notothenioid fishes (Perciformes: Notothenioidei) using complete gene sequences of the mitochondrial encoded 16S rRNA. Mol Phylogenet Evol 32: 881–891. [DOI] [PubMed] [Google Scholar]
- 31. Sanchez S, Dettaï A, Bonillo C, Ozouf-Costaz C, Detrich WH, et al. (2007) Molecular and morphological Phylogenies of the Nototheniidae, with on taxonomic focus on the Trematominae. Polar Biol 30: 155–166. [Google Scholar]
- 32.Lecointre G (2012) Phylogeny and systematics of Antarctic teleosts: methodological and evolutionary issues. Adaptation and Evolution in Marine Environments – The Impacts of Global Change on Biodiversity (ed. by G. di Prisco, and C. Verde), 97–117. Springer-Verlag, Berlin, Heidelberg.
- 33. Dettaï A, Berkani M, Lautrédou AC, Li B, Couloux A, et al. (2012) Tracking the elusive monophyly of nototheniid fishes (Teleostei) with multiple mitochondrial and nuclear markers. Mar Genomics 8: 49–58 doi:10.1016/j.margen.2012.02.003 [DOI] [PubMed] [Google Scholar]
- 34. Logsdon JM, Doolittle FW (1997) Origin of antifreeze protein genes: A cool tale in molecular evolution. Proc Natl Acad Sci USA 94: 3485–3487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Near TJ, Dornburg A, Kuhn KL, Eastman JT, Pennington JN, et al. (2012) Ancient climate change, antifreeze, and the evolutionary diversification of Antarctic fishes. Proc Natl Acad Sci USA 109(9): 3434–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Ozouf-Costaz C, Pisano E, Thaeron C, Hureau JC (1999) Karyological survey of the Notothenioid fish occuring in Adelie Land (Antarctica). Proceedings of the Vth Indo-Pac Fish Conf, Noumea (ed. by B. Séret, and J. Y. Sire), 427–440. Soc fr Ichthyol, Paris.
- 37.Pisano E., Ozouf-Costaz C (2002) Cytogenetics and evolution in extreme environment: the case of Antarctic Fishes. Fish adaptation. Val e G. Kapoor A, editor, 311–3338. Oxford IBH Publishing, New Delhi.
- 38. Near TJ, Cheng CC (2008) Phylogenetics of notothenioid fishes (Teleostei: Acanthomorpha): Inferences from mitochondrial and nuclear gene sequences. Mol Phylogenet Evol 47: 832–840. [DOI] [PubMed] [Google Scholar]
- 39. Klingenberg CP, Ekau W (1996) A combined morphometric and phylogenetic analysis of an ecomorphological trend: pelagization in Antarctic fishes (Perciformes: Nototheniidae). Biol J Linn Soc 59: 143–177. [Google Scholar]
- 40. Lautrédou AC, Bonillo C, Denys G, Cruaud C, Ozouf-Costaz C, et al. (2010) Molecular taxonomy and identification within the Antarctic genus Trematomus (Notothenioidei, Teleostei): How valuable is barcoding with COI? Polar Science 4(2): 333–352. [Google Scholar]
- 41. Lautrédou AC, Hinsinger D, Gallut C, Cheng CH, Berkani M, et al. (2012) Phylogenetic footprints of an Antarctic radiation: the Trematominae (Notothenioidei, Teleostei). Mol Phylogenet Evol 65(1): 87–101. [DOI] [PubMed] [Google Scholar]
- 42. Ekau W, Gutt J (1991) Notothenioid fishes from the Weddell Sea and their habitat, observed by underwater photography and television. Polar Biol 4: 36–49. [Google Scholar]
- 43. La Mesa M, Eastman JT, Vacchi M (2004) à The role of notothenioid fish in the food web of the Ross Sea shelf waters: a review. Polar Biol 27: 321–338. [Google Scholar]
- 44. Causse R, Ozouf-Costaz C, Koubbi P, Lamy D, Eléaume M, et al. (2011) Demersal ichthyofauna from the Dumont d'Urville Sea (East Antarctica) during the CEAMARC surveys in 2007–2008. Polar Science 5: 272–285. [Google Scholar]
- 45. Janko K, Marshall C, Musilová Z, Van Houdt J, Couloux C, et al. (2011) Multilocus analyses of an Antarctic fish species flock (Teleostei, Notothenioidei, Trematominae): Phylogenetic approach and test of the early-radiation event. Mol Phylogenet Evol 60: 305–316. [DOI] [PubMed] [Google Scholar]
- 46. Jones CD, Anderson ME, Balushkin AV, Duhamel G, Eakin RR, et al. (2008) Diversity, relative abundance, new locality records and population structure of Antarctic demersal fishes from the northern Scotia Arc islands and Bouvetoya. Polar Biol 31: 1481–1497. [Google Scholar]
- 47. Derome N, Chen WJ, Dettaï A, Bonillo C, Lecointre G (2002) Phylogeny of Antarctic dragonfishes (Bathydraconidae, Notothenioidei, Teleostei) and related families based on their anatomy and two mitochondrial genes. Mol Phylogenet Evol 24: 139–152. [DOI] [PubMed] [Google Scholar]
- 48. Near TJ, Pesavento JJ, Cheng CC (2003) Mitochondrial DNA, morphology and the phylogenetic relationships of Antarctic icefishes (Notothenioidei: Channichthyidae). Mol Phylogenet Evol 28: 87–98. [DOI] [PubMed] [Google Scholar]
- 49.Chen WJ, Bonillo C, Lecointre G (1998) Phylogeny of the Channichthyidae (Notothenioidei, Teleostei) Based on Two Mitochondrial Genes. Fishes of Antarctica. A Biological Overview (ed. by G. di Prisco, E. Pisano, and A. Clarke), 287–298. Springer-Verlag, Milano.
- 50. Lombarte A, Olaso I, Bozzano A (2003) Ecomorphological trends in the Artedidraconidae (Pisces: Perciformes: Notothenioidei) of the Weddell Sea. Antarct Sci 15: 211–218. [Google Scholar]
- 51. Lecointre G, Gallut C, Bonillo C, Couloux A, Ozouf-Costaz C, et al. (2011) The Antarctic Fish Genus Artedidraco is paraphyletic (Teleostei, Notothenioidei, Artedidraconidae). Polar Biol 34(8): 1135–1145. [Google Scholar]
- 52. De Broyer C, Guerra-Garcia JM, Takeuchi I, Robert H, Meerhaeghe A (2004) Biodiversity of the Southern Ocean: a catalogue of the Antarctic and sub-Antarctic Caprellidae and Cyamidae (Crustacea: Amphipoda) with distribution and ecological data. Bull Inst r Sci nat Belg 74: 61–99. [Google Scholar]
- 53. De Broyer C, Lowry JK, Jazdzewski K, Robert H (2007) Catalogue of the Gammaridean and Corophiidean Amphipoda (Crustacea) of the Southern Ocean, with distribution and ecological data. Bull Inst r Sci nat Belg 77: 1–325. [Google Scholar]
- 54. Zeidler W, De Broyer C (2009) Catalogue of the hyperiidean Amphipoda (Crustacea) of the Southern Ocean with distribution and ecological data. Bull Inst r Sci nat Belg 79: 1–100. [Google Scholar]
- 55. Dauby P, Scailteur Y, Chapelle G, De Broyer C (2001a) Potential impact of the main benthic amphipods on the eastern Weddell Sea shelf ecosystem (Antarctica). Polar Biol 24: 657–662. [Google Scholar]
- 56. Lörz AN, Maas EW, Linse K, Coleman CO (2009) Do circum-Antarctic species exist in peracarid Amphipoda? A case study in the genus Epimeria Costa, 1851. (Crustacea, Peracarida, Epimeriidae). Zookeys 18: 91–128. [Google Scholar]
- 57. Klages M, Gutt J (1990) Observations on the feeding behaviour of the Antarctic gammarid Eusirus perdentatus Chevreux, 1912 (Crustacea: Amphipoda) in aquaria. Polar Biol 10: 359–364. [Google Scholar]
- 58. Macnaughton MO, Thormar J, Berge J (2007) Sympagic amphipods in the Arctic pack ice: redescriptions of Eusirus holmii Hansen, 1887 and Pleusymtes karstensi (Barnard, 1959). Polar Biol 30: 1013–1025. [Google Scholar]
- 59.Barnard JL, Karaman GS (1991) The families and genera of Marine Gammaridean Amphipoda (Except Marine Gammaroidea). Rec Aust Mus: 41–412.
- 60. De Broyer C, Scailteur Y, Chapelle G, Rauschert M (2001) Diversity of epibenthic habitats of gammaridean amphipods in the eastern Weddell Sea. Polar Biol 24: 744–753. [Google Scholar]
- 61. Krapp RH, Berge J, Flores H, Gulliksen B, Werner I (2008) Sympagic occurence of Eusirid and Lysianassoid amphipods under Antarctic pack ice. Deep Sea Res Part 2 Top Stud Oceanogr 55 (8–9): 1015–1023. [Google Scholar]
- 62. Baird HP, Miller KJ, Stark JS (2011) Evidence of hidden biodiversity, ongoing speciation and diverse patterns of genetic structure in giant Antarctic amphipods. Mol Ecol 20: 3439–3454. [DOI] [PubMed] [Google Scholar]
- 63.Verheye M (2011) Systématique et diversité génétique des Eusirus de l'océan austral (Crustacea, Amphipoda, Eusiridae). Mémoire de l'Université catholique de Louvain, Louvain-la-Neuve. p. 127.
- 64. Chapelle G, Peck LS (1999) Polar gigantism dictated by oxygen availability. Nature 399: 114–115. [Google Scholar]
- 65. Held C (2000) Phylogeny and biogeography of serolid isopods (Crustacea, Isopoda, Serolidae) and the use of ribosomal expansion segments in molecular systematics. Mol Phylogenet Evol 15(2): 165–178. [DOI] [PubMed] [Google Scholar]
- 66. Leese F, Agrawal S, Held C (2010) Long-distance island hopping without dispersal stages: transportation across major zoogeographic barriers in a Southern Ocean isopod. Naturwissenschaften 97(6): 583–594. [DOI] [PubMed] [Google Scholar]
- 67. Leese F, Held C (2008) Identification and characterization of microsatellites from the Antarctic isopod Ceratoserolis trilobitoides: nuclear evidence for cryptic species. Conserv Genet 9: 1369–1372. [Google Scholar]
- 68. Krabbe K, Leese F, Mayer C, Tollrian R, Held C (2010) Cryptic mitochondrial lineages in the widespread pycnogonid Colossendeis megalonyx Hoek, 1881 from Antarctic and Subantarctic waters. Polar Biol 33: 281–292. [Google Scholar]
- 69. Wilson NG, Schrödl m, Halanych KM (2009) Ocean barriers and glaciation: evidence for explosive radiation of mitochondrial lineages in the Antarctic sea slug Doris kerguelenensis (Mollusca, Nudibranchia). Mol Ecol 18: 965–984. [DOI] [PubMed] [Google Scholar]
- 70. Poulin E, Féral J-P (1995) Pattern of spatial distribution of a brood-protecting schizasterid echinoid, Abatus cordatus, endemic to the Kerguelen Islands. Mar Ecol Prog Ser 118: 179–186. [Google Scholar]
- 71. Ledoux JB, Tarnowska K, Gérard K, Lhuillier E, Jacquemin B, et al. (2012) Fine-scale spatial genetic structure in the brooding sea urchin Abatus cordatus suggests vulnerability of the Southern Ocean marine invertebrates facing global change. Polar Biol 35: 611–623. [Google Scholar]
- 72. Poulin E, Féral J-P (1996) Why are there so many species of brooding Antarctic echinoids? Evolution 50(2): 820–830. [DOI] [PubMed] [Google Scholar]
- 73.Lockhart SJ (2006) Molecular evolution, phylogenetics and parasitism in Antarctic cidaroid echinoids. Unpublished PhD thesis, University Santa Cruz, California. 151 p.
- 74. Hétérier V, David B, De Ridder C, Rigaud T (2008) Ectosymbiosis is a critical factor in the local benthic biodiversity of the Antarctic deep sea. Mar Ecol Prog Ser 364: 67–76. [Google Scholar]
- 75. Hardy C, David B, Rigaud T, De Ridder C, Saucède T (2011) Ectosymbiosis associated with cidaroids (Echinodermata: Echinoidea) promotes benthic colonization of the seafloor in the Larsen Embayments, Western Antarctica. Deep Sea Res Part 2 Top Stud Oceanogr 58: 84–90. [Google Scholar]
- 76. Morescalchi A, Morescalchi MA, Odierna G, Stingo V, Capriglione T (1996) Karyotype and genome size of zoarcids and notothenioids (Teleostei, Percifirmes) from the Ross Sea: cytotaxonomic implications. Polar Biol 16: 559–564. [Google Scholar]
- 77. Ozouf-Costaz C, Hureau JC, Beaunier M (1991) Chromosome studies on fish of the suborder Notothenioidei collected in the Weddell Sea during EPOS 3 cruise. Cybium. 15(4): 275–290. [Google Scholar]
- 78. Ghigliotti L, Near TJ, Ferrando S, Vacchi M, Pisano E (2010) Cytogenetic diversity in the Antarctic plunderfishes(Notothenioidei: Artedidraconidae). Antarct Sci 22(6): 805–814. [Google Scholar]
- 79.Eléaume M (2006) Approche morphométrique de la variabilité phénotypique: conséquences systématiques et évolutives. Application aux crinoïdes actuels (Crinoidea: Echinodermata). Unpublished PhD dissertation, Muséum national d'Histoire naturelle, Paris. 402 p.
- 80.Hemery LG (2011) Diversité moléculaire, phylogéographie et phylogénie des Crinoïdes (Echinodermes) dans un environnement extrême: l'océan Austral. Unpublished PhD Dissertation, Muséum national d'Histoire naturelle, Paris. 398 p.
- 81. Clark AH, Clark Am (1967) A monograph of the existing crinoids, Vol. 1: the comatulids, part 5, Suborders Oligophreata (concluded) and Macrophreata. Bulletin of the United States National Museum 82: 1–795. [Google Scholar]
- 82. Speel JA, Dearborn JH (1983) Comatulid crinoids from the R/V Eltanin cruises in the Southern Ocean. Antarct Res Ser 38: 1–60. [Google Scholar]
- 83. Wilson NG, Hunter RL, Lockhart SJ, Halanych KM (2007) Multiple lineage and absence of panmixia in the “circumpolar” crinoid Promachocrinus kerguelensis from the Atlantic sector of Antarctica. Mar Biol 152: 895–904. [Google Scholar]
- 84. Hemery LG, Eléaume M, Roussel V, Améziane N, Gallut C, et al. (2012) Comprehensive sampling reveals circumpolarity and sympatry in seven mitochondrial lineages of the Southern Ocean crinoid species Promachocrinus kerguelensis (Echinodermata). Mol Ecol 21: 2502–2518. [DOI] [PubMed] [Google Scholar]
- 85.Lautrédou AC (2009) Relations de parenté et phylogéographie au sein des Liparidae (Gill, 1861). Unpublished MSc dissertation, Muséum national d'Histoire naturelle, Paris.
- 86. Duhamel G, Hautecoeur M, Dettaï A, Causse R, Pruvost P, et al. (2010) Liparids from the Eastern sector of Southern Ocean and first information from molecular studies. Cybium 34(4): 319–343. [Google Scholar]
- 87. Stein DL (2012) Snailfishes (Family Liparidae) of the Ross Sea, and closely adjacent waters. Zootaxa 3285: 1–120. [Google Scholar]
- 88. Raupach MJ, Thatje S, Dambach J, Rehm P, Misof B, Leese F (2010) Genetic homogeneity and circum-Antarctic distribution of two benthic shrimp species of the Southern Ocean, Chorismus antarcticus and Nematocarcinus lanceopes . Mar Biol 157: 1783–1797. [Google Scholar]
- 89. Udekem d'Acoz C d' (2010) Contribution to the knowledge of European Liljeborgiidae (Crustacea, Amphipoda), with considerations on the family and its affinities. Bull Inst r Sci nat Belg 80: 127–259. [Google Scholar]
- 90. Dauby P, Scailteur Y, De Broyer C (2001b) Trophic diversity within the eastern Weddell Sea amphipod community. Hydrobiologia 443: 69–86. [Google Scholar]
- 91. Udekem d'Acoz C d' (2009) New records of Liljeborgia from Antarctic and sub-Antarctic seas, with the description of two new species (Crustacea, Amphipoda, Liljeborgiidae). Bull Inst r Sci nat Belg 79: 243–304. [Google Scholar]
- 92. Udekem d'Acoz C d' (2008) Shelf and abyssal Liljeborgia Bate, 1861 of the Southern Ocean (Crustacea, Amphipoda, Liljeborgiidae). Bull Inst r Sci nat Belg 78: 45–286. [Google Scholar]
- 93. Udekem d'Acoz C d', Vader W (2009) On Liljeborgia fissicornis (M. Sars, 1858) and three related new species from Scandinavia, with a hypothesis on the origin of the group fissicornis . Journal of natural history 43: 2087–2139. [Google Scholar]
- 94. Havermans C, Nagy ZT, Sonet G, De Broyer C, Martin P (2011) DNA barcoding reveals new insights into the diversity of Antarctic species of Orchomene sensu lato (Crustacea: Amphipoda: Lysianassoidea). Deep Sea Res Part 2 Top Stud Oceanogr 58: 230–241. [Google Scholar]
- 95. Udekem d'Acoz C d', Havermans C (2012) Two new Pseudorchomene species from the Southern Ocean, with phylogenetic remarks on the genus and related species (Crustacea, Amphipoda, Lysianassoidea). Zootaxa 3310: 1–50. [Google Scholar]
- 96. Havermans C, Nagy ZT, Sonet G, De Broyer C, Martin P (2010) Incongruence between molecular phylogeny and morphological classification in amphipod crustaceans: A case study of Antarctic lysianassoids. Mol Phylogenet Evol 55: 202–209. [DOI] [PubMed] [Google Scholar]
- 97. Pierrat B, Saucède T, Laffont R, De Ridder C, Festeau A, et al. (2012) Large-scale distribution analysis of Antarctic echinoids using ecological niche modelling. Mar Ecol Prog Ser 463: 215–230 doi:10.3354/meps09842 [Google Scholar]
- 98. Díaz A, Féral J-P, David B, Saucède T, Poulin E (2011) Evolutionary pathways among shallow and deep-sea echinoids of the genus Sterechinus in the Southern Ocean. Deep Sea Res Part 2 Top Stud Oceanogr 58: 205–211. [Google Scholar]
- 99. Maddison WP (1989) Reconstructing character evolution on polytomous cladograms. Cladistics 5: 365–377. [DOI] [PubMed] [Google Scholar]