

Abstract
Mx proteins belong to the dynamin superfamily of high molecular weight GTPases and interfere with multiplication of a wide variety of viruses. Earlier studies show that nuclear mouse Mx1 and human MxA designed to be localized in the nucleus inhibit the transcription step of the influenza virus genome. Here we set a transient influenza virus transcription system using luciferase as a reporter gene and cells expressing the three RNA polymerase subunits, PB1, PB2 and PA, and NP. We used this reporter assay system and nuclear-localized MxA proteins to get clues for elucidating the anti-influenza virus activity of MxA. Nuclear-localized VP16-MxA and MxA-TAg NLS strongly interfered with the influenza virus transcription. Over-expression of PB2 led to a slight resumption of the transcription inhibition by nuclear MxA, whereas over-expression of PB1 and PA did not affect the MxA activity. Of interest is that the inhibitory activity of the nuclear MxA was markedly neutralized by over-expression of NP. An NP devoid of its C-terminal region, but containing the N-terminal RNA binding domain, also neutralized the VP16-MxA activity in a dose-dependent manner, whereas an NP lacking the N-terminal region did not affect the VP16-MxA activity. Further, not only VP16-MxA but also the wild-type MxA was found to interact with NP in influenza virus-infected cells. This indicates that the nuclear MxA suppresses the influenza virus transcription by interacting with not only PB2 but also NP.
INTRODUCTION
Mx proteins are interferon-inducible and have antiviral activity against a wide variety of viruses (1–7). Studies on Mx proteins started with identification of murine Mx1 in a mouse strain, A2G, resistant to influenza virus infection (8). Mx1 accumulates in the nucleus and inhibits the influenza virus transcription (9), thereby inhibiting influenza virus multiplication. Thereafter, a number of Mx proteins were identified in higher eukaryotes including fish, birds and mammals as a homolog of murine Mx1 (10–16).
MxA, a human homolog of Mx1, accumulates in the cytoplasm and interferes with multiplication of orthomyxoviruses (2,17,18), paramyxoviruses (19), bunyaviruses (3) and togavirus (4). When cells expressing MxA are infected with influenza virus, mRNA synthesis by primary transcription with virion-associated RNA polymerases is undergone at the same level as in MxA-negative cells (17). In contrast, viral protein synthesis and genome amplification are strongly inhibited (17). However, the molecular function and exact target(s) of MxA in influenza virus-infected cells remain unknown. In Thogoto virus-infected cells, MxA interacts with viral nucleocapsids and inhibits transport of viral nucleocapsids into the nucleus (20,21). Recently it has been revealed that MxA inhibits multiplication of hepatitis B virus, a DNA virus (6). In HBV-infected cells, export of viral mRNA from the nucleus is blocked by MxA. These two cases suggest that MxA may interfere with translocation of viral components between the nucleus and the cytoplasm.
To approach the molecular mechanism of the anti-influenza virus activity of MxA, two lines of evidence by earlier studies could be useful clues: MxA designed to be present in the nucleus inhibits the transcription of the influenza virus genome as Mx1 (22), and the anti-influenza virus activity of Mx1 is suppressed by over-expression of PB2 (23,24). In order to get more clues for the anti-influenza virus action of MxA and to look for a candidate(s) for a viral target molecule(s) of MxA, we tried to design experiments based on the above two facts. We set a reporter assay system, which utilized plasmids used for the DNA transfection-mediated virus-like particle generation system (25). We constructed a host cell RNA polymerase I (Pol I)-based plasmid, from which an engineered influenza virus genome containing a reporter gene of negative sense is generated. When the plasmid is introduced into cells expressing the viral RNA polymerase and NP, the engineered viral RNA generated by Pol I is transcribed. With this system, we found that nuclear MxA interferes with expression of the reporter gene. This inhibition was neutralized by over-expression of PB2 but not by that of PB1 or PA. Interestingly, over-expression of NP led to a significant suppression of the inhibitory activity of nuclear MxA. Immunoprecipitation assays revealed that nuclear MxA and NP form a complex under certain conditions. These results suggest that nuclear MxA proteins interfere with the viral transcription directly or indirectly through NP. Further, we found that NP is also immunoprecipitated with the wild-type MxA when lysates prepared from influenza virus-infected cells were used. A possible function of MxA in the context of its interaction with NP is also discussed.
MATERIALS AND METHODS
Construction of plasmid vectors
A plasmid vector, from which an artificial influenza virus genome containing luciferase gene of negative polarity as a reporter is synthesized in cells by the mouse DNA-dependent Pol I, was constructed as follows. The mouse Pol I promoter region was amplified by PCR with primers, 5′-TAA TACGACTCACTATA-3′ and 5′-GTCGGTACCTATCTCCAGGTCCA-3′ using pMrBKSP11 (a gift from Dr K. Yamamoto) (26) as a template. PCR products were phosphorylated with T4 polynucleotide kinase (TOYOBO) and digested with KpnI (TaKaRa). This fragment was cloned into pHH21 containing the promoter region of human ribosomal RNA gene (a gift from Dr Y. Kawaoka) (25,27), which had been digested with BssHII followed by treatment with Klenow fragment (TaKaRa) and subsequent digestion with KpnI. Thus, the treatment described above removed the human promoter in pHH21 and then replaced it with the mouse promoter. The resultant plasmid was designated pHMP1. Next, a fragment containing luciferase gene sandwiched by 5′- and 3′-terminal sequences of influenza A virus (WSN/33) segment 8 encoding non-structural protein (NS) was amplified by PCR with primers, 5′-GTAGTAGAAACAAGGGTGTTTTTTACTCGAGATCTTACAATTTGGACTTTCCGCCCTT-3′ and 5′-GATCCGTCTCCGGGAGCAAAAGCAG GGTGACAAAGACATAATGCATATGGAAGACGCCAAAAACATAAAGAAAGG-3′ using pGV-B (TOYO Ink) as a template. pHMP1 was digested with KpnI, blunted with T4 DNA polymerase (TOYOBO) and de-phosphorylated with calf intestine alkaline phosphatase (TaKaRa). The amplified PCR product was phosphorylated with T4 polynucleotide kinase and cloned into pHMP1 treated as above, resulting in construction of pHMP1-pre-vNS-Luc. Then, pHMP1-pre-vNS-Luc was digested with BsmB1 and re-ligated to produce pHMP1-vNS-Luc, in which the luciferase gene of reverse orientation sandwiched with 23 and 26 nucleotide-long 5′- and 3′-terminal sequences of the influenza virus segment 8 is placed under the control of mouse Pol I promoter (Fig. 1A).
Figure 1.

Transient transfection assays. (A) A reporter assay system for the influenza virus transcription based on an engineered influenza virus genome. For details, see the text. (B) Localization of MxA proteins. Swiss3T3 cells were grown on a glass coverslip in a 24-well plate and transfected with 0.5 µg of pVP16-MxA or pHMG-TMxA as described in Materials and Methods. At 48 h post-transfection, cells were fixed with 3% paraformaldehyde solution, permeabilized with 0.1% NP-40, washed twice with PBS, and then treated with 1% skim milk. The cells were incubated with mouse anti-MxA (KM1132) at 1:500 dilutions in 1% skim milk for 60 min and washed twice with 0.1% NP-40 and once with PBS. Cells were then stained with FITC-conjugated goat anti-mouse IgG at 1:500 dilutions in 1% skim milk. The coverslip was washed with 0.1% NP-40 and mounted in 0.1% p-phenylendiamine and 80% glycerol. MxA in cells expressing VP16-MxA (left) or MxA-TAg NLS (right) was visualized under a fluorescence microscope.
A plasmid encoding the wild-type MxA protein was constructed by subcloning of the full-length MxA fragment into a eukaryotic expression vector, pCAGGS (28). The MxA fragment prepared by digestion of pET3a-MxA (a gift from Dr Staeheli) with NdeI (BioLabs) and BamHI (TOYOBO) was once cloned into NdeI- and BamHI-digested pET14b (Novagen). The MxA fragment derived from pET14b-MxA by digestion with XbaI (TOYOBO) and EcoRV (TOYOBO) and blunted with Klenow fragment. Resultant DNA fragment was cloned into pCAGGS digested by XhoI (TOYOBO) and blunted with Klenow fragment. To construct a plasmid for expression of MxA fused to the trans-activation domain of herpes simplex virus VP16, pVP16 (Clontech), a plasmid for mammalian two-hybrid system was used. A fragment containing MxA gene was prepared from pET14b-MxA by digestion with NdeI followed by treatment with Klenow fragment and digestion with BamHI. The purified MxA fragment was cloned into pVP16 that had been digested with EcoRI (TOYOBO) and blunted with Klenow fragment followed by BamHI digestion. For expression of MxA tagged with the nuclear localization signal (NLS) sequence of SV40 T antigen, pHMG-TMxA was used (a gift from Dr Staeheli).
In order to construct pCHA-Mx1 encoding HA-tagged mouse Mx1, Mx1 cDNA was cloned into pCHA (29). The Mx1 cDNA portion was amplified by using KOD-plus (TOYOBO) with a previously constructed plasmid containing Mx1 cDNA (30) as a template and specific primers, 5′-TAG GCTAGCATGGATTCTGTGAATAATCTGTGC-3′ and 5′-TGAGCTAGCTTAATCGGAGAATTTGGCAAGCTT-3′. The PCR-amplified Mx1 fragment was digested with NheI (TOYOBO) and cloned into pCHA digested with NheI. pCHA-Mx1ΔC plasmid expressing Mx1 lacking its C-terminal (between amino acid positions 563 and 631) was spontaneously generated during the construction of pCHA-Mx1.
Plasmids encoding NP (pCAGGS-NP) and three subunits of the influenza virus RNA polymerase, PB1, PB2 and PA (pcDNA-PB1, pcDNA-PB2 and pcDNA-PA) were gifts from Dr Y. Kawaoka (25). In order to construct pCAGGS-NP-Myc for expression of Myc epitope-tagged NP, DNA fragments were amplified by PCR with primers, 5′-CCGAATTCCATGGCGTCTCAAGGCACCAAA-3′ and 5′-CGCGCCATGGCATTATCGTATTCCTCTGCA-3′ and a plasmid containing the NP gene as a template. The amplified fragment was digested with NcoI and ligated into NcoI-digested pBluescript II vector (Stratagene) containing a Myc tag sequence. Subsequently, a DNA fragment containing Myc-tagged NP sequence was excised from the pBluescript II and inserted into pCAGGS that had been digested with EcoRI and blunted with Klenow fragment. pSEAP2-control (Clontech) containing a secreted alkaline phosphatase (SEAP) gene under the control of the SV40 early promoter was used as a control for normalization of transfection. The nucleotide sequence of each plasmid was confirmed by DNA sequencing.
Cells and transfection
Swiss mouse 3T3 cell lines, Swiss3T3-Neo (Swiss3T3) and Swiss3T3-MxA cells (31), were kindly provided by Dr Haller. Either mouse Swiss3T3 cells or Clone 76 cells [a gift from Dr Nakata (28)] derived from mouse C127 cells were used as host cells for transfection experiments. Clone 76 cells contain PB2, PB1, PA and NP genes under the control of MMTV-LTR. The cells were maintained at 37°C in Dulbecco’s modified minimal essential medium containing 10% fetal calf serum. For transfection experiments, cells were grown in 24-well plates (up to 1 × 104 cells/well) or φ6 cm dishes (up to 5 × 104 cells/dish). DNA was transfected into cells using TransIT-LT1 transfection reagent (Mirus) according to the manufacturer’s instructions. The expression of influenza viral RNA polymerase subunits and NP in Clone 76 cells was induced by the addition of dexamethazone at the final concentration of 1 µM at 20 h post-transfection. In the case of Swiss3T3 cells, plasmids encoding three RNA polymerase subunits and NP were co-transfected with the reporter plasmid. At 48 h after either addition of dexamethazone or transfection, cells were harvested and their lysates were subjected to the luciferase assay or immunoprecipitation analyses.
Luciferase assay
The luciferase activity was determined using commercially available reagents (Promega) according to the manufacturer’s protocol. The relative luminescence intensity was measured with a luminometer for 20 s. The SEAP activity was determined according to the manufacturer’s instructions (TOYOBO). The luciferase activity was normalized as that relative to the SEAP activity/the amount of protein.
Preparation of antibodies, immunoprecipitation and western blot analysis
Detailed methods for preparation of mouse monoclonal anti-MxA antibodies will be described elsewhere. Briefly, recombinant MxA protein encoded on pET3a-MxA was expressed in Escherichia coli. MxA recovered in an insoluble fraction was purified by extensive washes with 6 M urea. The final insoluble fraction was dissolved in an SDS–PAGE sample buffer and subjected to SDS–PAGE. A protein band corresponding to MxA was excized, and MxA was eluted from the gel piece. MxA thus purified was used for immunization of mice (Balb/c). Preparation and selection of monoclonal cell lines were carried out essentially as described (29). Four monoclonal cell lines (KM1124, IgG1; KM1126, IgG1; KM1132, IgG2a and KM1135, IgG1) producing antibodies against MxA were established. It is reported that the antibody is useful in the measurement of MxA in virus-infected patients (32).
Swiss3T3-MxA stably expressing MxA or Swiss3T3 cells transfected with plasmid vectors of interest were infected with influenza A/PR/8/34 at m.o.i. of 5–10. After 8 h infection, the cells were harvested and lysed in buffer A containing 50 mM Tris–HCl (pH 8.0), 150 mM NaCl, 1 mM EDTA and 0.1% NP-40. The cell lysates were cleared by centrifugation at 4°C for 5 min at 10 000 g. After adding 1 µl of monoclonal mouse anti-MxA (a mixture of KM1132 and KM1126) or mouse anti-Myc (MBL), the lysates (200 µl) were incubated on ice for 2 h. Antibody–antigen complexes were mixed with protein A Sepharose beads (Pharmacia) and rotated at 4°C for a further 2 h. Protein A Sepharose beads were recovered by centrifugation and washed three times with buffer A. Beads were then suspended in an SDS-sample buffer and proteins were separated by electrophoresis through an 8% polyacrylamide gel in the presence of 0.1% SDS. Proteins were transferred to a polyvinylidene difluoride (PVDF) membrane. The membrane was exposed first to a specific primary antibody (monoclonal mouse anti-MxA, monoclonal mouse anti-Myc or anti-NP polyclonal rabbit antisera) and then to a horseradish peroxidase-conjugated second antibody against species-specific immunogloblin. Proteins were visualized with ECL detection kit (Amersham Life Science).
RESULTS
Construction of a reporter assay system for influenza virus transcription using an artificial influenza virus genome
Transient transfection assay systems using reporter genes such as chloramphenicol acetyltransferase (33) and luciferase (34) genes have been contributing to studies on fundamental and regulatory mechanisms of DNA transcription. Assay systems similar to those were developed and used for analysis of the mechanism of RNA synthesis of the negative-sense RNA genome (35). Recently, Neumann et al. established a plasmid DNA transfection-mediated influenza virus-like particle (VLP) generation system (25). In the system, influenza virus genome RNAs of exact size and orientation are synthesized by the cellular DNA-dependent RNA Pol I from plasmid DNAs encoding viral cDNAs cloned between the Pol I-promoter and terminator. Viral proteins are supplied from viral protein expression vectors cloned under the control of the cellular DNA-dependent RNA polymerase II promoter. The minimal requirement for production of infectious recombinant influenza viruses is a set of Pol I plasmids corresponding to eight viral RNA segments and four plasmids encoding the viral RNA polymerase subunits, PB2, PB1, and PA, and NP (25), suggesting that these four proteins are enough and sufficient for primary transcription of the viral genome within cells. Based on these observations, we have set a transient transfection system using a viral genome, in which the coding region for a viral gene is replaced with a reporter gene, luciferase, while cis-acting regulatory regions (36) remain intact (Fig. 1A, also see Materials and Methods). Pol I produces a negative-sense luciferase RNA sandwiched with viral terminal sequences. Luciferase mRNA is synthesized by transcription of the negative-sense RNA with the viral RNA polymerase and NP. The synthesized mRNA is subjected to translation. This system should be useful to study the mechanism of transcription of the influenza viral genome. Here we have examined the effect of nuclear MxA proteins on this system.
The effect of nuclear MxA on the transient transcription system
First, we tested whether the reporter plasmid, pHMP1-vNS-Luc (Fig. 1A), is active in mouse Clone 76 cells. Clone 76 cells contain PB2, PB1, PA and NP genes under the control of MMTV-LTR, so that these genes are expressed by the addition of dexamethazone (37). pHMP1-vNS-Luc contains the luciferase gene of reverse orientation sandwiched with 5′- and 3′-terminal sequences of influenza virus (A/WSN/33) segment 8, and vNS-luc RNA is synthesized under the control of the mouse Pol I promoter (Fig. 1A). As expected, the luciferase activity was detected in dexamethazone-treated Clone 76 cells transfected with pHMP1-vNS-Luc (data not shown).
We then tried to use the system in order to confirm that nuclear MxA proteins inhibit the viral transcription and to get a cue for the function of MxA. To this end, two plasmids encoding either MxA tagged with the NLS sequence of SV40 T antigen (MxA-TAg NLS) (22) or MxA fused to the transactivation domain of HSV VP16 (VP16-MxA) were constructed. Both MxA-TAg NLS and VP16-MxA derived from these plasmids were strictly localized in the nucleus (Fig. 1B). When clone 76 cells were co-transfected with pHMP1-vNS-Luc and plasmids expressing nuclear MxA and treated with dexamethazone for expression of the viral RNA polymerase and NP at 20 h post-transfection, the reporter luciferase activity was markedly reduced (Fig. 2A). Parental vector plasmids, one of which contains the VP16 portion, did not have any effect (data not shown). These results confirm that nuclear MxA inhibits viral transcription.
Figure 2.

The effect of Mx proteins on expression of the reporter gene. (A) The effect of nuclear MxA proteins. Clone 76 cells were grown to 50–60% confluence in 24-well plates and transfected with pHMP1-vNS-Luc (0.2 µg), pSEAP2-control (0.1 µg) and either parental vector, pVP16-MxA or pHMG-TMxA (0.3 µg) encoding nuclear MxA. The expression of influenza virus RNA polymerase subunits and NP was induced by treatment of cells with dexamethazone at 1 µM at the final concentration. At 48 h post-transfection, cells were harvested and assayed for luciferase activity. The expression level of SEAP was determined as described in Materials and Methods and used to standardize the efficiency of transfection. The luciferase activity was normalized as that relative to the SEAP activity/the amount of protein. (B) The effect of wild-type Mx1 and C-terminal deletion Mx1 mutant (Mx1ΔC) in Swiss3T3 cells. The cells were transfected with pHMP1-vNS-Luc (0.2 µg), pSEAP2-control (0.1 µg), plasmids encoding three RNA polymerase subunits and NP (0.1 µg of each plasmid), parental vector (0.3 µg), and either pCHA-Mx1 or pCHA-Mx1ΔC (0.01–0.3 µg each). The luciferase activity was determined as described above and in Materials and Methods. Error bars represent standard deviation (n = 3).
Next, we modified the system so that the viral RNA polymerase and NP are transiently supplied by transfection of plasmids encoding these components. Swiss3T3 cells (a kind gift from Dr Haller) were used in place of Clone 76 cells, because Swiss3T3 cells have been used for detailed analyses of MxA function. This assay would be meritorious, because the ratio of these proteins can be manipulated (see below). We could detect the luciferase expression in Swiss3T3 cells co-transfected with pHMP1-vNS-Luc and plasmids encoding the viral RNA polymerase and NP. Luciferase expression was totally dependent on the RNA polymerase and NP (data not shown). In agreement with earlier studies (9,17), luciferase expression was inhibited with a wild-type nuclear Mx1 in a dose-dependent manner but not with a mutant Mx1 (Mx1ΔC) devoid of its C-terminal region corresponding to the amino acid positions between 563 and 631 (Fig. 2B). These results indicate that the transient transfection assay system can be used for further analyses. Next, we tested the effect of wild-type and nuclear MxA proteins on luciferase expression in this system (Fig. 3). Nuclear MxA proteins, VP16-MxA and MxA-TAg NLS (Fig. 1B), inhibited the luciferase expression in Swiss3T3 cells (Fig. 3A) as effectively as in Clone 76 cells (Fig. 2A). The inhibition was dependent on increasing amounts of nuclear VP16-MxA (Fig. 3B). In contrast, the cytoplasmic wild-type MxA was less effective in inhibition of luciferase expression (Fig. 3B). C-terminal and internal deletion VP16-MxA mutants gave a limited level of inhibition, suggesting that these regions are important for the inhibition activity of MxA (Fig. 3C).
Figure 3.


The effect of transiently expressed nuclear MxA proteins. (A) The effect of nuclear MxA proteins in Swiss3T3 cells. The cells were transfected with pHMP1-vNS-Luc (0.2 µg), pSEAP2-control (0.1 µg), plasmids encoding three RNA polymerase subunits and NP (0.05 µg of each plasmid) and either parental vector, pVP16-MxA or pHMG-TMxA (0.3 µg each). Luciferase activity was determined and shown as described in Materials and Methods. Error bars represent standard deviation (n = 3). (B) Dose-dependent effect of wild-type MxA and VP16-MxA. The same procedure for (A) was carried out in the presence of increasing amounts of wild-type MxA or VP16-MxA. Error bars represent standard deviation (n = 3). (C) The effect of C-terminal (MxAΔC) and internal (MxAΔM) deletion VP16-MxA mutant proteins on reporter gene expression. The same procedure for (A) was carried out with mutant MxA proteins. To generate a VP16-MxA deletion mutant (MxAΔC) for expression of MxA lacking its C-terminal region (362–662), we amplified a fragment by PCR with specific primers, 5′-GGCATCCATATGGTTGTTTCCGAAGTGGACATCGCA-3′ and 5′-CGCGGATCCTTAACCATACTTTTGTAGCTCCTCTGT-3′ and pVP16-MxA as template. To generate a VP16-MxA deletion mutant (MxAΔM) lacking its internal region (362–573), a fragment was amplified by PCR with primers, 5′-GGCATCCATATGGTTGTTTCCGAAGTGGACATC GCA-3′ and 5′-CGCGGATCCTTAACCGGGGAACTGGGCAAGCCGGCG-3′ and pCHA-MxAΔM plasmid as a template. pCHA-MxAΔM was derived from previously constructed plasmid, pCHA-MxA by removal of an internal part of MxA by digestion with SalI (TOYOBO) and NcoI (TOYOBO) restriction enzymes. The main part of the plasmid was blunted with Klenow fragment and self-ligated. MxA fragments thus prepared were digested with NdeI, blunted with Klenow fragment and then digested with BamHI. These fragments were cloned into pVP16 plasmid digested with EcoRI followed by Klenow treatment and subsequent digestion with BamHI. Error bars represent standard deviation (n = 3).
Rescue of nuclear MxA-mediated transcription inhibition by over-expression of influenza virus components involved in viral RNA synthesis
Since MxA, when present in the nucleus, has an inhibitory effect on the viral transcription, it is quite possible that a component(s) involved in viral RNA synthesis is a target of MxA. To test this, each one of the components was over-expressed and the inhibitory effect of VP16-MxA was analyzed. The amount of one of the expression plasmids for PB1, PB2, PA or NP was increased from 0.05 to 0.35 µg, the amount for the other plasmids being kept at 0.05 µg (Fig. 4A and B). When each plasmid was used at the amount of 0.05 µg, VP16-MxA inhibited expression of the luciferase. The MxA-mediated inhibition of the viral transcription was partially suppressed by over-expression of PB2, whereas over-expression of PB1 and PA had no remarkable effect (Fig. 4A). However, it is noted that over-expression of NP significantly rescued the luciferase expression (Fig. 4B). The effect of increasing amounts of NP on the reporter assay system was examined in the absence of MxA. It is shown that luciferase activity is not stimulated but rather slightly inhibited in Swiss3T3 cells transfected with additional amounts of the NP expressing plasmid. To further investigate the effect of NP over-expression, the plasmid expressing NP was increased from 0.1 to 0.5 µg in addition to the basal amount (0.15 µg), while the amount of plasmids expressing three RNA polymerase subunits was kept at 0.05 µg. The expression plasmids for NP and VP16-MxA were co-transfected at the various ratios into the Swiss3T3 cells (Fig. 4B). The MxA inhibitory activity was completely neutralized by over-expression of NP when cells were transfected with the low amount (0.1 µg) of pVP16-MxA. The luciferase expression remained inhibited at the high amount (0.5 µg) of pVP16-MxA even when increasing amounts of NP were supplied. Titration experiments of the plasmid encoding NP in the presence of 0.1 or 0.3 µg of pVP16-MxA were carried out (Fig. 4C). The amount of the plasmid for NP required for the rescue in the presence of 0.3 µg of pVP16-MxA was not exactly three times more but two to two-and-a-half times more than that in the presence of 0.1 µg of pVP16-MxA. Thus, more NP is needed for rescue from the inhibitory effect of more VP16-MxA. Next, we investigated the effect of C- and N-terminal deletion NP mutant proteins. The C-terminal deletion NP mutant that contains an RNA binding domain neutralized the MxA-mediated inhibition in cells transfected with 0.1 µg of pVP16-MxA although less effectively than the full-sized NP. In contrast, N-terminal deletion NP mutant did not show remarkable neutralizing activity (Fig. 4D). These results suggest that nuclear MxA proteins interfere with the viral transcription by directly or indirectly interacting with PB2 and/or NP. Further, the N-terminal domain of NP is involved in the inhibitory function of MxA.
Figure 4.


Rescue of the inhibitory activity of MxA proteins by over- expression of influenza RNA polymerase subunits and NP. (A) Effect of over-expression of viral RNA polymerase subunits. Swiss3T3 cells were transfected with pHMP1-vNS-Luc (0.2 µg), pSEAP (0.1 µg), pVP16-MxA (0 or 0.1 µg indicated – or +, respectively), and three RNA polymerase subunits and NP (0.05 µg each). The additional amount (0.3 µg) of a plasmid encoding either PB1, PB2 or PA was added as indicated. The luciferase activity was determined and shown as described in Materials and Methods. Error bars represent standard deviation (n = 3). (B) Effect of over-expression of NP. Swiss3T3 cells were transfected with pHMP1-vNS-Luc (0.2 µg), pSEAP (0.1 µg), pVP16-MxA (0, 0.1 or 0.5 µg), and three RNA polymerase subunits (0.05 µg each) and NP (0.15 µg). The additional amounts of the plasmid encoding NP were added as indicated. The luciferase activity was determined and shown as described in Materials and Methods. The result is shown as the average of two independent experiments. (C) Titration of plasmid encoding NP. Swiss3T3 cells were transfected with pHMP1-vNS-Luc (0.2 µg), pSEAP (0.1 µg), three RNA polymerase subunits (0.05 µg each) and NP (0.1, 0.3 or 0.5 µg) in the absence or presence of pVP16-MxA. The relative luciferase activity in the presence of MxA (0.1 µg, open circle; 0.3 µg, closed circle) was normalized with that in the absence of MxA, and plotted as a function of increasing amounts of NP. The result is shown as the average of two independent experiments. (D) Titration of VP16-MxA, and C-terminal (NPΔC) and N-terminal (NPΔN) deletion mutants of NP. The same procedure for (B) was carried out with mutant NPs. For expression of NP proteins lacking its C-terminal region (NPΔC) corresponding amino acid positions between 182 and 498 (where amino acid position 1 is set the first amino acid of the wild-type NP) and its N-terminal region (NPΔN) corresponding amino acid positions between 1 and 190, pCAGGS-NPΔC and pCAGGS-NPΔN plasmids were constructed as follows: portions of NP fragments were amplified by PCR with pCAGGS-NP as a template and a set of primers, 5′-TTGAATTCGCCACCATGGCGACCAAAGGCACC-3′ and 5′-TTGAATT CTTAAGCACCTGCGGCCCCAGACC-3′ for NPΔC and a set of primers, 5′-TTGAATTCGCCACCATGGAATTGATCAGAATG-3′ and 5′-GGGA ATTCTTAATTGTCGTACTCCTCTGCA-3′ for NPΔN. Synthesized NP fragments were digested with EcoRI and cloned into pCHA that had been digested with the same restriction enzyme. The additional amounts of the plasmid encoding deletion mutants of NP were transfected as indicated. The result is shown as the average of two independent experiments.
Physiological interaction of MxA with NP
To test whether MxA interacts with NP, immunoprecipitation assays were performed using lysates prepared from Swiss3T3 cells transfected with expression vectors for both VP16-MxA and NP tagged with the Myc epitope (NP-Myc). Proteins immunoprecipitated with either anti-MxA or anti-Myc antibody were separated by SDS–PAGE, transferred onto a PVDF filter, and visualized with a mixture of anti-MxA and anti-Myc antibodies. Figure 5A shows no significant interaction between MxA and NP in lysates prepared under the conditions described in Materials and Methods. Considering the possibility that viral RNA was needed for MxA to interact with NP, pHMP1-vNS-Luc was co-transfected. Co-expression of vNS-Luc RNA with NP and VP16-MxA did not affect the result. Further, we could not detect any interaction of VP16-MxA with NP in lysates prepared from cells transfected with VP16-MxA and super-infected with influenza virus A/PR/8/34 (data not shown).
Figure 5.

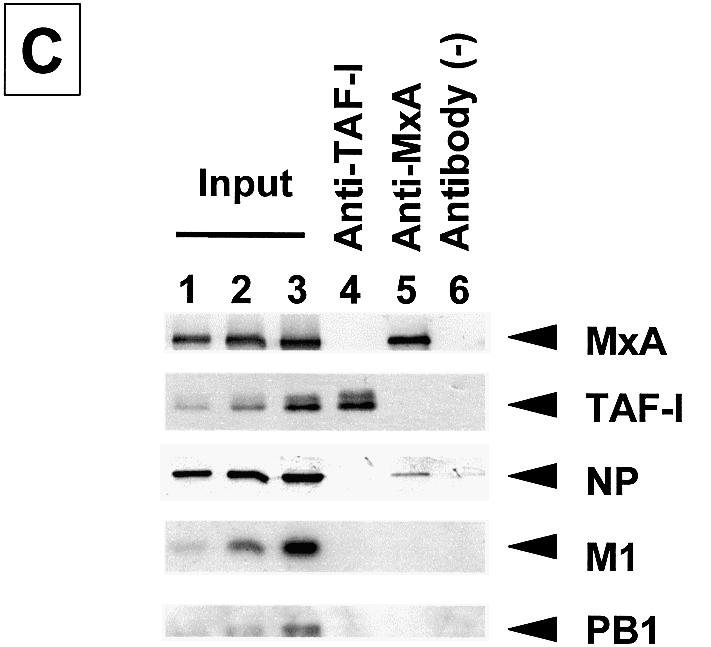
Immunoprecipitation analyses for interaction between MxA and NP. (A) Immunoprecipitation without cross-linking. Swiss3T3 cells were grown in φ6 cm dishes and transfected with plasmids encoding VP16-MxA (2.5 µg) and NP-Myc (2.5 µg) in the presence or absence of pHMP1-vNS-Luc (1 µg). At 48 h post-transfection, cells were lysed in buffer A. One microliter of 1 mg/ml monoclonal mouse anti-MxA (a mixture of KM1132 and KM1126), or anti-Myc was added to 200 µl of lysates and antibody–antigen complexes were precipitated as described in Materials and Methods. Immunoprecipitated proteins were denatured in 30 µl of SDS sample buffer containing 0.58 M β-mercaptethanol and separated through an 8% polyacrylamide gel. Proteins were transferred to a PVDF membrane, probed with mouse anti-MxA and anti-Myc antibodies mixture, and then visualized with ECL detection kit (Amersham Life Science). Lane 1 contains non-transfected Swiss3T3 cell lysates and lanes 2 and 3 contain 10% of the input of transfected Swiss3T3 lysates used for experiments in lanes 4–7. (B) Immunoprecipitation with cross-linking. Swiss3T3 cells transfected with pVP16-MxA and infected with influenza A/PR/8/34 was washed twice with PBS. The cells (2 × 106 equivalent) were collected into a 1.5 ml tube and resuspended in 495 µl of PBS. Proteins were cross-linked by addition of 0.1 M dithiobis(succinimidyl propionate) at a final concentration of 1 mM followed by incubation at 4°C for 60 min. The cross-linking reaction was stopped by addition of 25 µl of 1 M glycine. Cells were collected by centrifugation and lysed in 300 µl of buffer A. The cell lysates were centrifuged at 4°C for 5 min at 10 000 g and cleared lysates were transferred into a new tube. MxA and NP proteins in lysates were immunoprecipitated by using either mouse anti-MxA (a mixture of KM1132 and KM1126 mixture, lane 3) or anti-TAF-I (lane 2) (29), the antibody for nuclear TAF-I distinct from MxA as mentioned above. After being dissolved in an SDS sample buffer, proteins were separated through an 8% polyacrylamide gel and detected by western blotting using specific antibodies for MxA or NP. Lane 1 contains 5% of the input of lysates used for experiments in lanes 2 and 3 (C) Immunoprecipitation using lysates prepared from Swiss3T3-MxA cells infected with influenza A/PR/8/34 under the cross-linking condition. The same procedure for (B) was carried out for immunoprecipitation, except that influenza virus-infected Swiss3T3-MxA cells were used instead of VP16-MxA-transfected and influenza virus-infected Swiss3T3 cells. Proteins were visualized by western blotting with antibodies specific for MxA, TAF-I, NP, M1 and PB1. The antibody against PB1 was a gift from Dr Toyoda. Details for preparation of rabbit polyclonal anti-M1 and anti-NP antibodies with purified M1 (61) and NP (62) will be described elsewhere.
Considering the possibility that the interaction between NP and MxA, if any, is weak or/and transient, we carried out immunoprecipitation assays with lysates that had been treated with reversible cross-linking reagent dithiobis(succinimidyl propionate) prior to immunoprecipitation. Lysates were prepared from Swiss3T3 cells transfected with VP16-MxA and super-infected with influenza viruses (Fig. 5B). Immunoprecipitation assays (Fig. 5B) with cross-linked lysates revealed that VP16-MxA is associated with NP expressed in infected cells. In order to analyze the interaction between NP and the wild-type MxA, lysates were prepared from Swiss3T3-MxA cells infected with influenza viruses. After cross-linking, lysates were subjected to immunoprecipitation assay. Figure 5C showed that the wild-type MxA is associated with NP. Since we could not rule out the indirect interaction between MxA and NP, it is possible that MxA and NP are included in a putative complex through a molecule(s) bridging the two proteins. Viral M1 and PB1 proteins are not detected in the immunoprecipitates under the conditions employed here.
DISCUSSION
Human MxA is a cytoplasmic Mx and a structural and functional homolog of nuclear mouse Mx1. Mx1 is found to inhibit influenza virus transcription (9,17). It is suggested that PB2, a subunit of the influenza viral RNA polymerase, is a putative target of Mx1 (23,24), although there is no evidence showing a direct interaction between MxA and the viral RNA polymerase. Since a MxA mutant localized in the nucleus apparently functions as Mx1 (22), we tried to get a cue to know the function and a target(s) of MxA using nuclear type MxA and a system that allows us to monitor influenza virus transcription based on expression of a reporter gene. Systems for reverse-genetics of influenza virus have been developed (35). Transient transfection assay systems using a reporter gene have been developed and used for functional analyses on cis- and trans-acting factors involved in viral RNA synthesis of the influenza virus genome (for example 38). Our system described in this study is exactly the same as one that Fodor et al. used for analysis of PA subunit, except that our Pol I system is for mouse cells (39).
Several kinds of reporter assay system have been used for analyses of the MxA function. The multiplication of SFV, a positive-strand RNA virus, was inhibited by MxA at early steps in its replication cycle (4). The anti-SFV mechanism by MxA was examined using a transfected replicon system. Landis et al. established an SFV-based replicon system, in which a recombinant SFV genome containing SFV replicase and a reporter gene in place of viral structural protein genes is produced with in vitro transcription and transfected into cells (4). Using this system, viral structural proteins of SFV are found not to be required for the inhibition of SFV replication by MxA. Vaccinia virus-based transient expression systems were developed and used to study the mechanism of transcription and replication of orthomyxoviruses such as Thogoto virus (40) and influenza virus (41). In these systems, an artificial virus genome containing a reporter gene is reconstituted in vivo with the RNA polymerase and NP supplied by virus infection or T7 RNA polymerase provided by a recombinant vaccinia virus. Mx1 suppresses expression of a reporter gene from an influenza virus-like RNA in the vaccinia virus-based system (23). It was shown by using a reporter mini-replicon derived from a recombinant vaccinia virus that both wild-type cytoplasmic and nuclear forms of MxA significantly suppress Thogoto virus transcription (42). In this system, however, the effect of vaccinia virus infection could not be completely ruled out.
Using our transient transfection assay system, we found that nuclear MxA strongly interferes with influenza virus transcription (Figures 2 and 3). It has been reported that nuclear mouse Mx1 and human MxA-TAg NLS inhibit primary transcription in infected cells (9,17,22). Nuclear localization of Mx1 is necessary for inhibition of influenza virus transcription (43). The C-terminal deleted Mx1, which is localized in the cytoplasm (43 and data not shown), did not show an inhibitory effect on our reporter assay system (Fig. 2B). Thus, our results obtained by the transient transfection assay for influenza virus transcription are in good agreement with previous reports. Mx1 accumulates in the nucleus and inhibits influenza virus multiplication. Further, over-expression of PB2 neutralizes the anti-influenza virus activity of mouse Mx1 protein, but that of PB1, PA or NP does not (23,24). In our system, we examined the effect of over-expression of PB1, PB2, PA or NP on the transcription inhibitory activity of nuclear MxA. Over-expression of PB2 and NP neutralized the VP16-MxA activity at different levels, while PB1 and PA did not affect the MxA activity. The suppression of the VP16-MxA inhibitory activity by over-expression of PB2 may be due to the putative mechanism similar to that for Mx1. Since it is described that direct interaction of Mx1 with PB2 is not detected (23,24), the PB2 over-expression-mediated rescue is possibly interpreted as PB2 and Mx1 competing with each other for interaction with a common target molecule(s). However, the suppression level by PB2 against MxA in our system is quite low, so that the mode of interaction between MxA and its putative target and/or a target itself may be different from those of Mx1.
Of importance is that over-expression of NP is found to be much more effective than that of PB2 to rescue viral transcription activity from the VP16-MxA inhibitory activity (Fig. 4). The neutralization effect with NP on MxA was cancelled by increased amounts of MxA. These results suggest that MxA inhibits an NP and/or NP-related function(s) involved in influenza virus transcription. NP is thought to be an essential factor for elongation of RNA chain (44). Thus, nuclear MxA might inhibit the elongation step of viral RNA synthesis. Immunoprecipitation assays revealed a physiological interaction of MxA with NP. However, this interaction was found only under a cross-linking condition, suggesting that the interaction is weak and/or transient. Alternatively, the interaction may be bridged by one or more mediator molecules, so that a chain interaction among NP, MxA and putative mediators may be negatively cooperated, even if the affinity between each two molecules is reasonable. It is interesting to speculate that MxA titrates out not only a free form of NP but also a molecule that associates and allows NP to be active or to be placed appropriately in the transcription machinery, since several proteins that interact with NP are identified (45,46).
Studies using transgenic mice and transfected cells expressing MxA indicate that MxA has an antiviral effect against a broad range of RNA viruses (1,3–5,47). Different kinds of inhibitory mechanism are postulated for the anti-virus mechanism of MxA. In Thogoto virus-infected cells, MxA interacts with viral nucleocapsids and blocks nuclear import of the viral genome (20,21). MxA suppresses multiplication of HBV by inhibiting nuclear export of viral mRNA (6). These suggest two possible MxA functions: MxA may interact with viral RNA possibly associated with NP- or RNP-binding proteins and MxA may function in the mechanism of intracellular protein/RNA translocation. Indeed, MxA is a member of the dynamin superfamily of proteins, which are involved in intracellular vesicle/protein-sorting (48–51). In this respect, it is shown that a part of MxA is localized to the endoplasmic reticulum (ER) and tubulated lipid (52).
An important question is raised of whether native MxA also affect the NP and/or NP-related function during the course of influenza virus infection in the cytoplasm. To analyze the MxA function in inhibition of Thogoto virus multiplication, two kinds of DNA-based transcription/replication system were used (42). In the vaccinia virus-based system, MxA effectively inhibits the reporter gene expression. In contrast, no significant inhibitory effect of MxA is observed in the Pol I-based system. It is worthwhile to note that vRNP is formed in the cytoplasm in the vaccinia virus system, whereas vRNP formation in the Pol I-based system is in the nucleus. These suggest that the intracellular distribution of MxA and the intracellular compartment for vRNP formation are critical for the MxA function. We showed that the nuclear VP16-MxA is co-immunoprecipitaed with NP (Fig. 5B). Further, we examined interaction of cytoplasmic MxA and NP using lysates prepared from Swiss3T3-MxA infected with influenza virus. Swiss3T3 cells (a gift from Dr Haller) constitutively express MxA that confers anti-influenza virus activity (31). Under cross-linked conditions employed here, NP was shown to be immunoprecipitated with MxA when lysates prepared from infected cells were used (Fig. 5C). With this result, it would be worthwhile to extend discussion on the function of MxA, in particular, that is mediated through the interaction between MxA and NP in the cytoplasm. One of the cytoplasmic forms of NP must be associated to incoming (infecting) or progeny RNP complexes, so that MxA may interact with influenza virus RNP complexes as it does with viral RNP complexes in Thogoto virus-infected cells (20). However, this may not be the case. Pavlovic et al. concluded MxA does not block nuclear import of incoming influenza virus RNP complexes (17), since primary transcription is not inhibited in cells expressing MxA. Alternatively, nuclear export of newly formed vRNP may be blocked by MxA at the cytoplasmic face as MxA inhibits nuclear export of viral mRNAs in HBV-infected cells (6). Newly synthesized NP is the other cytoplasmic form of NP. Since a portion of NP is associated with importin-α (45), MxA may affect this interaction. It is also possible that MxA interacts with nascent NP or a molecule(s) associated with nascent NP on ER. On this line, we have shown that expression of MxA affects cellular functions including translation and cell death not only in influenza virus-infected cells but also in uninfected cells (53). Thus, it is presumed that MxA inhibits virus multiplication indirectly by affecting host cellular functions.
It appears that MxA gives its antiviral activity through a variety of virus multiplication steps and cellular pathways (1,3,4,6,17,19,20,22,54–56). The COP9 (constitutive photomorphogenesis 9) complex termed signalosome consists of multi-components. The components interact with a wide variety of proteins including transcription factors, translation factors, signal transducers, intracellular modifiers and proteosome subunits and regulate those functions as a sort of buffer (57). MxA exists in cells as a giant complex possibly with or without other molecules (58–60). It is assumed that a MxA complex as a multi-valent complex may interact with a variety of host and viral factors. Thus, systematic identification of factors that interact with MxA is an important issue.
Acknowledgments
ACKNOWLEDGEMENTS
We thank Drs O. Haller (University of Freiburg, Freiburg, Germany), P. Staeheli (University of Freiburg, Freiburg, Germany), Y. Kawaoka (Institute of Medical Science, University of Tokyo, Tokyo, Japan), T. Toyoda (Kurume University, Kurume, Japan) and K. Yamamoto (Nagasaki University, Nagasaki, Japan) for the generous gifts of Swiss3T3 and Swiss3T3-MxA cells (to O.H.), pET3a-MxA and pHMG-TMxA (to P.S.), plasmid vectors, pCAGGS-NP, pcDNA-PB1, pcDNA-PB2, pcDNA-PA and pHH21 (to Y.K.), anti-PB1 antibody (to T.T.), and pMrBKSP11 (to K.Y.). This work was supported in part by a grant-in-aid from the Ministry of Education, Culture, Sports, Science and Technology of Japan (to K.N.), a grant for the Bioarchitect Program from RIKEN (Institute of Physical and Chemical Research, Japan) (to K.N.), a grant from the Sumitomo Foundation (to K.N.), an Invitation Fellowship for Research in Japan from the Japan Society for the Promotion of Science for (to K.T.), and a Fellowship from the 34th UNESCO International Course for Advanced Research in Chemistry and Chemical Engineering (to K.T.).
REFERENCES
- 1.Staeheli P. and Pavlovic,J. (1991) Inhibition of vesicular stomatitis virus mRNA synthesis by human MxA protein. J. Virol., 65, 4498–4501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Frese M., Kochs,G., Meier-Dieter,U., Siebler,J. and Haller,O. (1995) Human MxA protein inhibits tick-borne Thogoto virus but not Dhori virus. J. Virol., 69, 3904–3909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Frese M., Kochs,G., Feldmann,H., Hertkorn,C. and Haller,O. (1996) Inhibition of bunyaviruses, phleboviruses and hantaviruses by human MxA protein. J. Virol., 70, 915–923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Landis H., Simon-Jodicke,A., Kloti,A., Di Paolo,C., Schnorr,J.J., Schneider-Schaulies,S., Hefti,H.P. and Pavlovic,J. (1998) Human MxA protein confers resistance to Semliki Forest virus and inhibits the amplification of Semliki Forest virus-based replicon in the absence of viral structural proteins. J. Virol., 72, 1516–1522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Chieux V., Chehadeh,W., Harvey,J., Haller,O., Wattre,P. and Hober,D. (2001) Inhibition of coxsackievirus B4 replication in stably transfected cells expressing human MxA protein. Virology, 283, 84–92. [DOI] [PubMed] [Google Scholar]
- 6.Gordien E., Rosmorduc,O., Peltekian,C., Garreau,F., Brechot,C. and Kremsdorf,D. (2001) Inhibition of hepatitis B virus replication by the interferon-inducible MxA protein. J. Virol., 75, 2684–2691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Jin H.K., Yoshimatsu,K., Takada,A., Ogino,M., Asano,A., Arikawa,J. and Watanabe,T. (2001) Mouse Mx2 protein inhibits hantavirus but not influenza virus replication. Arch. Virol., 146, 41–49. [DOI] [PubMed] [Google Scholar]
- 8.Staeheli P., Colonno,R.J. and Cheng,Y.S. (1983) Different mRNAs induced by interferon in cells from inbred mouse strains A/J and A2G. J. Virol., 47, 563–567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Krug R.M., Shaw,M., Broni,B., Shapiro,G. and Haller,O. (1985) Inhibition of influenza viral mRNA synthesis in cells expressing the interferon-induced Mx gene product. J. Virol., 56, 201–206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Horisberger M.A., Wathelet,M., Szpirer,J., Szpirer,C., Islam,Q., Levan,G., Huez,G. and Content,J. (1988) cDNA cloning and assignment to chromosome 21 of IFI-78K gene, the human equivalent of murine Mx gene. Somat. Cell Mol. Genet., 14, 123–131. [DOI] [PubMed] [Google Scholar]
- 11.Meier E., Fah,J., Grob,M.S., End,R., Staeheli,P. and Haller,O. (1988) A family of interferon-induced Mx-related mRNAs encodes cytoplasmic and nuclear proteins in rat cells. J. Virol., 62, 2386–2393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Staeheli P., Yu,Y.X., Grob,R. and Haller,O. (1989) A double-stranded RNA-inducible fish gene homologous to the murine influenza virus resistance gene Mx. Mol. Cell Biol., 9, 3117–3121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Muller M., Winnacker,E.L. and Brem,G. (1992) Molecular cloning of porcine Mx cDNAs: new members of a family of interferon-inducible proteins with homology to GTP-binding proteins. J. Interferon Res., 12, 119–129. [DOI] [PubMed] [Google Scholar]
- 14.Bazzigher L., Schwarz,A. and Staeheli,P. (1993) No enhanced influenza virus resistance of murine and avian cells expressing cloned duck Mx protein. Virology, 195, 100–112. [DOI] [PubMed] [Google Scholar]
- 15.Ellinwood N.M., McCue,J.M., Gordy,P.W. and Bowen,R.A. (1998) Cloning and characterization of cDNAs for a bovine (Bos taurus) Mx protein. J. Interferon Cytokine Res., 18, 745–55. [DOI] [PubMed] [Google Scholar]
- 16.Leong J.C., Trobridge,G.D., Kim,C.H., Johnson,M. and Simon,B. (1998) Interferon-inducible Mx proteins in fish. Immunol. Rev., 166, 349–363. [DOI] [PubMed] [Google Scholar]
- 17.Pavlovic J., Haller,O. and Staeheli,P. (1992) Human and mouse Mx proteins inhibit different steps of the influenza virus multiplication cycle. J. Virol., 66, 2564–2569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Marschall M., Zach,A., Hechtfischer,A., Foerst,G., Meier-Ewert,H. and Haller,O. (2000) Inhibition of influenza C viruses by human MxA protein. Virus Res., 67, 179–188. [DOI] [PubMed] [Google Scholar]
- 19.Zhao H., De,B.P., Das,T. and Banerjee,A.K. (1996) Inhibition of human parainfluenza virus-3 replication by interferon and human MxA. Virology, 220, 330–338. [DOI] [PubMed] [Google Scholar]
- 20.Kochs G. and Haller,O. (1999) Interferon-induced human MxA GTPase blocks nuclear import of Thogoto virus nucleocapsids. Proc. Natl Acad. Sci. USA, 96, 2082–2086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kochs G. and Haller,O. (1999) GTP-bound human MxA protein interacts with the nucleocapsids of Thogoto virus (Orthomyxoviridae). J. Biol. Chem., 274, 4370–4376. [DOI] [PubMed] [Google Scholar]
- 22.Zurcher T., Pavlovic,J. and Staeheli,P. (1992) Mechanism of human MxA protein action: variants with changed antiviral properties. EMBO J., 11, 1657–1661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Huang T., Pavlovic,J., Staeheli,P. and Krystal,M. (1992) Overexpression of the influenza virus polymerase can titrate out inhibition by the murine Mx1 protein. J. Virol., 66, 4154–4160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Stranden A.M., Staeheli,P. and Pavlovic,J. (1993) Function of the mouse Mx1 protein is inhibited by overexpression of the PB2 protein of influenza virus. Virology, 197, 642–651. [DOI] [PubMed] [Google Scholar]
- 25.Neumann G., Watanabe,T., Ito,H., Watanabe,S., Goto,H., Gao,P., Hughes,M., Perez,D.R., Donis,R., Hoffmann,E., Hobom,G. and Kawaoka,Y. (1999) Generation of influenza A viruses entirely from cloned cDNAs. Proc. Natl Acad. Sci. USA, 96, 9345–9350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Yamamoto K., Koga,A., Yamamoto,M., Nishi,Y., Tamura,T., Nogi,Y. and Muramatsu,M. (2000) Identification of a novel 70 kDa protein that binds to the core promoter element and is essential for ribosomal DNA transcription. Nucleic Acids Res., 28, 1199–1205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Neumann G., Zobel,A. and Hobom,G. (1994) RNA polymerase I-mediated expression of influenza viral RNA molecules. Virology, 202, 477–479. [DOI] [PubMed] [Google Scholar]
- 28.Niwa H., Yamamura,K. and Miyazaki,J. (1991) Efficient selection for high-expression transfectants with a novel eukaryotic vector. Gene, 108, 193–199. [DOI] [PubMed] [Google Scholar]
- 29.Nagata K., Saito,S., Okuwaki,M., Kawase,H., Furuya,A., Kusano,A., Hanai,N., Okuda,A. and Kikuchi,A. (1998) Cellular localization and expression of template-activating factor I in different cell types. Exp. Cell Res., 240, 274–281. [DOI] [PubMed] [Google Scholar]
- 30.Nakayama M., Yazaki,K., Kusano,A., Nagata,K., Hanai,N. and Ishihama,A. (1993) Structure of mouse Mx1 protein. Molecular assembly and GTP-dependent conformational change. J. Biol. Chem., 268, 15033–15038. [PubMed] [Google Scholar]
- 31.Pavlovic J., Zurcher,T., Haller,O. and Staeheli,P. (1990) Resistance to influenza virus and vesicular stomatitis virus conferred by expression of human MxA protein. J. Virol., 64, 3370–3375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Yoshimasu T., Manabe,A., Ebihara,Y., Tanaka R., Ooi,J., Iseki,T., Shirafuji,N., Maekawa,T., Asano,S., Yoshikawa,N. and Tsuji,K. (2003) MxA expression in patients with viral infection after allogeneic stem cell transplantation. Bone Marrow Transplant, 32, 313–316. [DOI] [PubMed] [Google Scholar]
- 33.Gorman C.M., Moffat,L.F. and Howard,B.H. (1982) Recombinant genomes which express chloramphenicol acetyltransferase in mammalian cells. Mol. Cell. Biol., 2, 1044–1051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.deWet J.R., Wood,K.V., DeLuca,M., Helinski,D.R. and Subramani,S. (1987) Firefly luciferase gene: structure and expression in mammalian cells. Mol. Cell Biol., 7, 725–737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Neumann G., Whitt,M.A. and Kawaoka,Y. (2002) A decade after the generation of a negative-sense RNA virus from cloned cDNA—what have we learned? J. Gen. Virol., 83, 2635–2662. [DOI] [PubMed] [Google Scholar]
- 36.Neumann G. and Kawaoka,Y. (1999) Genetic engineering of influenza and other negative-strand RNA viruses containing segmented genomes. Adv. Virus Res., 53, 265–300. [DOI] [PubMed] [Google Scholar]
- 37.Kimura N., Nishida,M., Nagata,K., Ishihama,A., Oda,K. and Nakada,S. (1992) Transcription of a recombinant influenza virus RNA in cells that can express the influenza virus RNA polymerase and nucleoprotein genes. J. Gen. Virol., 73, 1321–1328. [DOI] [PubMed] [Google Scholar]
- 38.Pleschka S., Jaskunas,R., Engelhardt,O.G., Zurcher,T., Palese,P. and Garcia-Sastre,A. (1996) A plasmid-based reverse genetics system for influenza A virus. J. Virol., 70, 4188–4192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Fodor E., Crow,M., Mingay,L.J., Deng,T., Sharps,J., Fechter,P. and Brownlee,G.G. (2002) A single amino acid mutation in the PA subunit of the influenza virus RNA polymerase inhibits endonucleolytic cleavage of capped RNAs. J. Virol., 76, 8989–9001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Weber F., Jambrina,E., Gonzalez,S., Dessens,J.T., Leahy,M., Kochs,G., Portela,A., Nuttall,P.A., Haller,O., Ortin,J. and Zurcher,T. (1998) In vivo reconstitution of active Thogoto virus polymerase: assays for the compatibility with other orthomyxovirus core proteins and template RNAs. Virus Res., 58, 13–20. [DOI] [PubMed] [Google Scholar]
- 41.Fuerst T.R., Niles,E.G., Studier,F.W. and Moss,B. (1986) Eukaryotic transient-expression system based on recombinant vaccinia virus that synthesizes bacteriophage T7 RNA polymerase. Proc. Natl Acad. Sci. USA, 83, 8122–8126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Weber F., Haller,O. and Kochs,G. (2000) MxA GTPase blocks reporter gene expression of reconstituted Thogoto virus ribonucleoprotein complexes. J. Virol., 74, 560–563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Garber E.A., Hreniuk,D.L., Scheidel,L.M. and van der Ploeg,L.H. (1993) Mutations in murine Mx1: effects on localization and antiviral activity. Virology, 194, 715–723. [DOI] [PubMed] [Google Scholar]
- 44.Honda A., Ueda,K., Nagata,K. and Ishihama,A. (1988) RNA polymerase of influenza virus: role of NP in RNA chain elongation. J. Biochem. (Tokyo), 104, 1021–1026. [DOI] [PubMed] [Google Scholar]
- 45.Wang P., Palese,P. and O’Neill,R.E. (1997) The NPI-1/NPI-3 (karyopherin alpha) binding site on the influenza a virus nucleoprotein NP is a nonconventional nuclear localization signal. J. Virol., 71, 1850–1856. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Momose F., Basler,C.F., O’Neill,R.E., Iwamatsu,A., Palese,P. and Nagata,K. (2001) Cellular splicing factor RAF-2p48/NPI-5/BAT1/UAP56 interacts with the influenza virus nucleoprotein and enhances viral RNA synthesis. J. Virol., 75, 1899–1908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Pavlovic J., Arzet,H.A., Hefti,H.P., Frese,M., Rost,D., Ernst,B., Kolb,E., Staeheli,P. and Haller,O. (1995) Enhanced virus resistance of transgenic mice expressing the human MxA protein. J. Virol., 69, 4506–4510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.van der Bliek A.M. and Meyerowitz,E.M. (1991) Dynamin-like protein encoded by the Drosophila shibire gene associated with vesicular traffic. Nature, 351, 411–414. [DOI] [PubMed] [Google Scholar]
- 49.Chen M.S., Obar,R.A., Schroeder,C.C., Austin,T.W., Poodry,C.A., Wadsworth,S.C. and Vallee,R.B. (1991) Multiple forms of dynamin are encoded by shibire, a Drosophila gene involved in endocytosis. Nature, 351, 583–586. [DOI] [PubMed] [Google Scholar]
- 50.Vater C.A., Raymond,C.K., Ekena,K., Howald-Stevenson,I. and Stevens,T.H. (1992) The VPS1 protein, a homolog of dynamin required for vacuolar protein sorting in Saccharomyces cerevisiae, is a GTPase with two functionally separable domains. J. Cell Biol., 119, 773–786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Sever S. (2002) Dynamin and endocytosis. Curr. Opin. Cell Biol., 14, 463–467. [DOI] [PubMed] [Google Scholar]
- 52.Accola M.A., Huang,B., Al Masri,A. and McNiven,M.A. (2002) The antiviral dynamin family member, MxA, tubulates lipids and localizes to the smooth endoplasmic reticulum. J. Biol. Chem., 277, 21829–21835. [DOI] [PubMed] [Google Scholar]
- 53.Mibayashi M., Nakad,K. and Nagata,K. (2002) Promoted cell death of cells expressing human MxA by influenza virus infection. Microbiol. Immunol., 46, 29–36. [DOI] [PubMed] [Google Scholar]
- 54.Schnorr J.J., Schneider-Schaulies,S., Simon-Jodicke,A., Pavlovic,J., Horisberger,M.A. and ter Meulen,V. (1993) MxA-dependent inhibition of measles virus glycoprotein synthesis in a stably transfected human monocytic cell line. J. Virol., 67, 4760–4768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Schneider-Schaulies S., Schneider-Schaulies,J., Schuster,A., Bayer,M., Pavlovic,J. and ter Meulen,V. (1994) Cell type-specific MxA-mediated inhibition of measles virus transcription in human brain cells. J. Virol., 68, 6910–6917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Schuster A., Johnston,I.C., Das,T., Banerjee,A.K., Pavlovic,J., ter Meulen,V. and Schneider-Schaulies,S. (1996) Expression of the human MxA protein is associated with hyperphosphorylation of VSV P protein in human neural cells. Virology, 220, 241–245. [DOI] [PubMed] [Google Scholar]
- 57.Karniol B. and Chamovitz,D.A. (2000) The COP9 signalosome: from light signaling to general developmental regulation and back. Curr. Opin. Plant Biol., 3, 387–393. [DOI] [PubMed] [Google Scholar]
- 58.Richter M.F., Schwemmle,M., Herrmann,C., Wittinghofer,A. and Staeheli,P. (1995) Interferon-induced MxA protein. GTP binding and GTP hydrolysis properties. J. Biol. Chem., 270, 13512–13517. [PubMed] [Google Scholar]
- 59.Schumacher B. and Staeheli,P. (1998) Domains mediating intramolecular folding and oligomerization of MxA GTPase. J. Biol. Chem., 273, 28365–28370. [DOI] [PubMed] [Google Scholar]
- 60.DiPaolo C., Hefti,H.P., Meli,M., Landis,H. and Pavlovic,J. (1999) Intramolecular backfolding of the carboxyl-terminal end of MxA protein is a prerequisite for its oligomerization. J. Biol. Chem., 274, 32071–32078. [DOI] [PubMed] [Google Scholar]
- 61.Watanabe K., Handa,H., Mizumoto,K. and Nagata,K. (1996) Mechanism for inhibition of influenza virus RNA polymerase activity by matrix protein. J. Virol., 70, 241–247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Yamanaka K., Ishihama,A. and Nagata,K. (1990) Reconstitution of influenza virus RNA-nucleoprotein complexes structurally resembling native viral ribonucleoprotein cores. J. Biol. Chem., 265, 11151–11155. [PubMed] [Google Scholar]