

Abstract
Estrogens exert a variety of effects in both reproductive and non-reproductive tissues. With the discovery of ERα splice variants, prior assumptions concerning tissue-specific estrogen signaling need to be re-evaluated. Accordingly, we sought to determine the expression of the classical estrogen receptors and ERα splice variants across reproductive and non-reproductive tissues of male and female mice. Western blotting revealed that the full-length ERα66 was mainly present in female reproductive tissues but was also found in non-reproductive tissues at lower levels. ERα46 was most highly expressed in the heart of both sexes. ERα36 was highly expressed in the kidneys and liver of female mice but not in the kidneys of males. ERβ was most abundant in non-reproductive tissues and in the ovaries. Because the kidney has been reported to be the most estrogenic non-reproductive organ, we sought to elucidate ER renal expression and localization. Immunofluorescence studies revealed ERα66 in the vasculature and the glomerulus. It was also found in the brush border of the proximal tubule and in the cortical collecting duct of female mice. ERα36 was evident in mesangial cells and tubular epithelial cells of both sexes, as well as podocytes of females but not males. ERβ was found primarily in the podocytes in female mice but was also present in the mesangial cells in both sexes. Within the renal cortex, ERα46 and ERα36 were mainly located in the membrane fraction although they were also present in the cytosolic fraction. Given the variability of expression patterns demonstrated herein, identification of the specific estrogen receptors expressed in a tissue is necessary for interpreting estrogenic effects. As this study revealed expression of the ERα splice variants at multiple sites within the kidney, further studies are warranted in order to elucidate the contribution of these receptors to renal estrogen responsiveness.
Introduction
Although estrogen was discovered in the 1920s [1], [2] and the hormone was proposed to act via a receptor, the human estrogen receptor (ERα66) was not cloned until 1986 [3]. Subsequent advances in the understanding of estrogen signaling came with the development of the estrogen receptor knockout mouse [4], the identification and cloning of a second estrogen receptor, ERβ [5], [6], and development of its knockout mouse [7]. Through these studies, the existence of multiple receptors for estrogen signaling with unique downstream events became evident. Novel roles for estrogens have been demonstrated in non-reproductive tissues, such as the skeletal [8] and cardiovascular systems [9]–[12], the brain [13], [14], and adipose tissue [15], [16], as well as their importance in male fertility [17]–[20]. Additionally, estrogen has been shown to work through both genomic pathways (transcriptional regulation through estrogen response elements) and non-genomic pathways (activation of cell-signaling cascades). Studies of a mouse model with ERα66 expressed only in the membrane [21] revealed the importance of genomic ERα66 in fertility.
In the past decade, it has become evident that there are at least two physiologically relevant splice variants of ERα66. The first, named ERα46 reflecting its molecular weight, was identified by Flouriot et al. [22] in the human breast cancer cell line, MCF7. The second, ERα36, was identified in 2005 [23]. Both ERα46 and ERα36 are truncated in the amino-terminus (173 amino acids) and lack the first transcriptional activation domain (AF1). ERα46 is identical to ERα66 in the remaining protein sequence. ERα36 lacks the second transcriptional activation domain (AF2) and has a unique C-terminus. The ERα36 protein is identical to the full-length receptor in its DNA binding domain and part of its dimerization and ligand binding domains. Because the ligand binding domain differs from that of ERα66, ERα36 may be capable of interacting with a wider array of estrogens than ERα66. Both ERα46 and ERα36 can form homodimers or they can heterodimerize with ERα66. ERα46 has a 2-fold higher binding affinity to the estrogen response element than ERα66 [24]. Post-translational modification by palmitoylation of ERα46 and myristoylation of ERα36 may target them to the membrane [23]. (For structure of the splice variants, see [25]).
These ERα splice variants have been studied primarily in human cancer cell lines. Their patterns of expression in mammalian organs and their contribution to estrogen signaling in the normal state are not known. It is evident that interactions among these receptors modulate estrogen signaling in the cell. An understanding of the tissue-specific expression of the various estrogen receptors is necessary for identifying the individual functional roles that each may play under physiologic conditions. As the kidney has been described as the most estrogen-responsive organ after gonadal tissue [26], it seems likely that ERα splice variants might contribute to the overall estrogen signaling in the kidney. Therefore, we hypothesized that the ERα splice variants and ERβ may be expressed differently in each organ in the body, and that estrogen receptors would be differentially localized to specific cell types within the kidney.
Methods
Ethics Statement
All animal procedures were approved and conducted in accordance with the guidelines of the Institutional Animal Care and Use Committee of the University of Nebraska Medical Center under protocols 10-081-12-FC and 10-082-10-FC.
Animal Procedures
Mice were housed five per cage and in a 12-hr light/dark cycle environment. Mice had ad libitum access to water and standard mouse chow. Male and female mice were sacrificed under intraperitoneal ketamine (100 mg/kg)/xylazine (10 mg/kg) anesthesia at 6–12 wks of age and organs were harvested for western blotting and immunofluorescence. In the female mice, kidneys, heart, liver, uterus, mammary and ovaries were collected. In the males, the kidneys, heart, liver and testes were harvested.
Western Blot (WB)
Tissue was snap-frozen in liquid nitrogen and stored at –80°C until homogenization and lysis in radioimmunoprecipitation assay (RIPA) buffer with Halt™ Protease and Phosphatase Inhibitor Cocktail (Thermo Fisher Scientific, Waltham, MA). Protein concentration was determined using the Bradford assay (Coomassie plus Protein, Thermo Fisher Scientific) with absorbance read at 595 nm on a uQuant microplate reader (Bio-Tek Instruments, Winooski, VT). After protein was boiled in 6X sample buffer (Boston Bioproducts, Ashland, MA), 33 µg were loaded on a 10% Tris-HCl gel (Bio-Rad, Hercules, CA). The gel was transferred to a PVDF membrane (Bio-Rad) and blocked in ODYSSEY blocking buffer (LI-COR Biosciences, Lincoln, NE) for 1 hr at room temperature. Then, membranes were incubated with primary antibodies in blocking buffer overnight at 4°C. The following morning, the membranes were washed in PBS with Tween 20, and then incubated in secondary antibodies for 1 hr at room temperature. After additional washes, membranes were visualized with the ODYSSEY® Infrared Imaging System (LI-COR Biosciences).
Immunofluorescence (IF)
Kidneys were rinsed in saline, decapsulated and bisected longitudinally. They were then submerged in a Tissue-Tek® Cryomold tissue cassette filled with O.C.T.™ Compound (Sakura Finetek, Torrance, CA), frozen in liquid nitrogen and stored at –80°C. Sections (6-µm) were cut on a Leica CM3050S Cryostat (Leica Microsystems, Buffalo Grove, IL), transferred onto ProbeOn Plus® slides (Thermo Fisher) and fixed in 4% paraformaldehyde at 4°C. The sections were blocked for 1 hr in a buffer containing 10% donkey serum with BSA and Triton X-100. Then, the sections were incubated in primary antibodies in blocking buffer overnight at 4°C. The following day, after washing in PBS, the sections were incubated with fluorescent secondary antibodies in the dark at room temperature. Slides were mounted in ProLong® Gold Antifade Reagent with DAPI (Molecular Probes|Life Technologies, Grand Island, NY), coverslipped and visualized on a confocal microscope (Leica TCS SP5, Leica Microsystems) at 630× magnification.
Antibodies
Rabbit anti-ERα36 (1∶150 IF females, 1∶50 IF males) was a kind gift of Zhao Yi Wang. Mouse anti-α smooth muscle actin (A5228; 1∶200 IF) was purchased from Sigma Aldrich (St. Louis, MO). Rabbit anti-ERα (H184, SC-7207; 1∶50 IF), rabbit anti-ERα (MC-20, SC-542; 1∶1000 WB), rabbit anti-ERβ (H-150, SC-8974; 1∶500 WB, 1∶50 IF), goat anti-synaptopodin (N-14, SC-21536; 1∶50 IF), goat anti-aquaporin 2 (C-17, SC-9882; 1∶50 IF) and goat anti-PECAM-1 (M-20, SC-1506; 1∶50 IF) antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA). The following antibodies labeled with Alexa Fluor® (AF) dyes were purchased from Invitrogen?Life Technologies: AF-488 donkey anti-rabbit IgG (A21206; 1∶200), AF-594 donkey anti-goat IgG (A11058; 1∶200), AF-594 donkey anti-mouse IgG (A21203; 1∶200) and AF-680 goat anti-mouse IgG (A21058; 1∶10,000 WB). Goat anti-rabbit IgG conjugated to IRDye® 800CW (611-131-122; 1∶10,000 WB) was from Rockland Immunochemicals (Gilbertsville, PA).
Separation of Membrane and Cytosolic Fractions
The kidney was dissected into cortex and medulla. The cortex was then separated into membrane and cytosolic fractions using the Mem-Per kit (Pierce, Rockford, IL) according to the manufacturer’s protocol. Briefly renal cortices were homogenized in TBS, centrifuged, re-suspended in provided reagents, centrifuged again and then the membrane and cytosolic fractions were separated. The fractions were dialyzed overnight prior to proceeding with a standard Western blotting protocol.
Statistics
Differences among groups were computed using two-way analysis of variance followed by Holm-Sidak method for pairwise multiple comparison analysis (Sigma Plot 11.0; Systat Software, San Jose, CA). One-way analysis of variance was performed within sex to account for unpaired reproductive tissue, followed by Tukey test. Probability values less than 0.05 were considered to be significant. Values are reported as means ± standard error.
Results
ERα66 in Male and Female Mice
Western blotting revealed that ERα66 protein level was more that 500-fold higher in the uterus and ovaries than in the kidney and other tissues; hence, the results are plotted in Figure 1 on a logarithmic scale. In the female mice, ERα66 expression was evident (in decreasing rank order) in the mammary gland, liver, heart, lung and kidney. In male mice, ERα66 protein level was highest in the liver, testis and kidney; however, the prominent band for testis on the Western blot ran at a slightly smaller molecular weight than evident in the uterus or ovaries. ERα66 protein expression was extremely low in the heart and lung of male mice.
Figure 1. ERα66 protein level in various organs of mice.
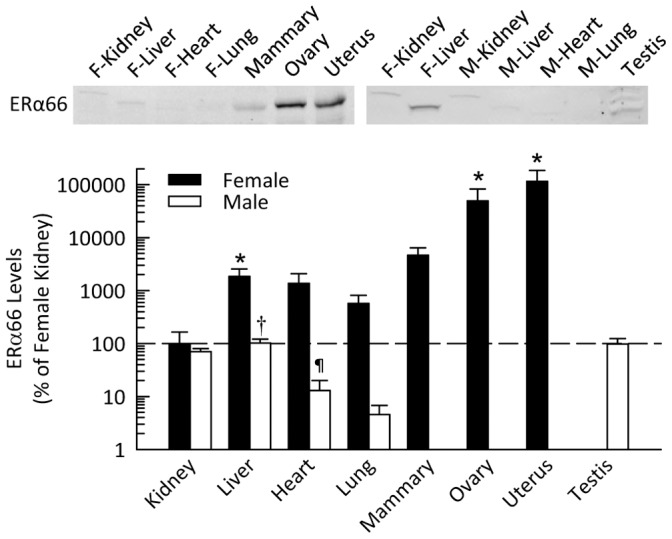
Shown are representative Western blots and quantification for expression in non-reproductive and reproductive organs harvested from female and male mice. Data are shown as percent of ERα66 in female kidney, plotted on a logarithmic scale. *P<0.05 vs. female kidney, † P<0.05 vs. female liver, ¶ P<0.05 vs. female heart (n = 4–5 per group).
ERα46 in Male and Female Mice
ERα46 protein level was most abundant in the heart and uterus in female mice (Figure 2), and in the heart and testes of male mice. As was the case for ERα66, the prominent band for ERα46 in the testis ran at a smaller molecular weight than in other tissues. ERα46 was present at lower levels in the remaining tissues investigated, although it was barely detectable in the lungs of both sexes.
Figure 2. ERα46 protein level in various organs of mice.

Shown are representative Western blots and quantification for expression in non-reproductive and reproductive organs harvested from female and male mice. Data are shown as percent of ERα46 in female kidney. *P<0.05 vs. female kidney, † P<0.05 vs. female liver, ¶ P<0.05 vs. heart (of same sex), £ P<0.05 vs. uterus, ‡ P<0.05 vs. testis (n = 4–5 per group).
ERα36 in Male and Female Mice
As illustrated in Figure 3, ERα36 was most highly expressed in the kidney of female mice, but was almost non-detectable in the kidney of male mice (P<0.001 vs. females). In contrast, there was no sexual dimorphism with regard to ERα36 levels in other organs. This splice variant was expressed in all the tissues investigated but at significantly lower levels than in the female kidney, although the data for the ovary did not achieve statistical significance.
Figure 3. ERα36 protein level in various organs of mice.
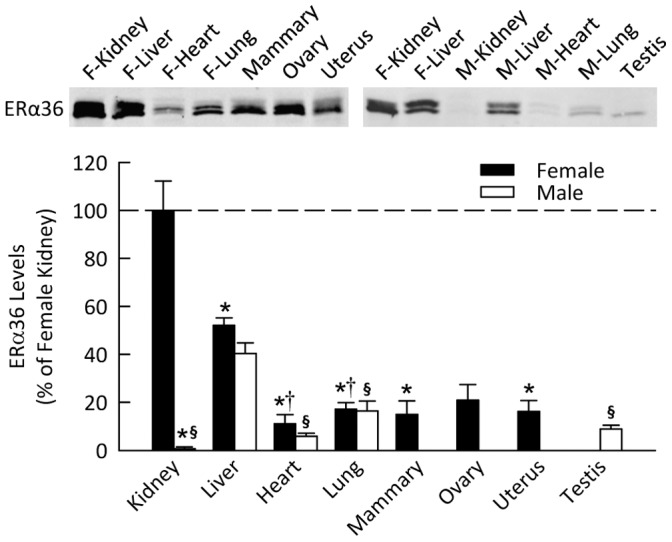
Shown are representative Western blots and quantification for expression in non-reproductive and reproductive organs harvested from female and male mice. Data are shown as percent of ERα36 in female kidney. *P<0.05 vs. female kidney, § P<0.05 vs. male liver, † P<0.05 vs. female liver (n = 4–5 per group).
ERβ in Male and Female Mice
In the female mice, ERβ protein level was highest in the liver, kidney, heart and ovary (Figure 4), with lower levels evident in mammary gland, uterus and lung. The male mice had similar expression of ERβ to that of females in all tissues except for the kidney, where levels in males were significantly lower than females. In the male mice, the liver and the heart had the most abundant level of ERβ.
Figure 4. ERβ protein level in various organs of mice.
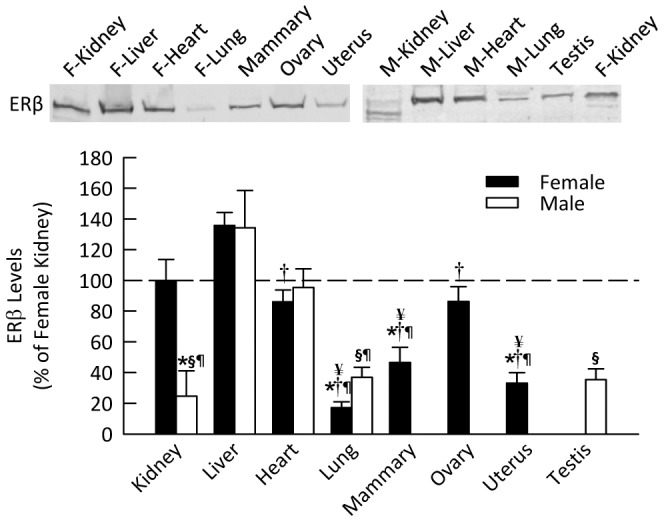
Shown are representative Western blots and quantification for expression in non-reproductive and reproductive organs harvested from female and male mice. Data are shown as percent of ERβ in female kidney. *P<0.05 vs. female kidney, § P<0.05 vs. male liver, † P<0.05 vs. female liver, ¶ P<0.05 vs. heart (of same sex), ¥ P<0.05 vs. ovary (n = 4 per group).
ER Localization in the Kidney
Because the kidney has been described as the most estrogenic non-reproductive organ, intrarenal ERα66, ERα36 and ERβ localization was investigated in frozen sections. ERα46 localization could not be investigated because of the lack of a specific antibody for this splice variant. ERα66 detected using an antibody against the N-terminus was found mainly in the vasculature and in the glomerulus, although not in podocytes (Figure 5). In females, it was also detectable in the brush border of the proximal tubules and in the apical aspect of cortical collecting duct principal cells where it co-localized with aquaporin-2 (AQP2).
Figure 5. Representative immunofluorescence images localizing ERα66 in mouse kidney.

Green fluorescence represents ERα66 in all images. Red fluorescence represents cell specific markers for co-localization (collecting duct marker, aquaporin 2; vascular smooth muscle & mesangial cell marker, α-smooth muscle actin; podocyte cell marker, synaptopodin). Nuclei are stained blue with DAPI. Scale bar = 20 µm.
As shown in Figure 6, ERα36 co-localized with the glomerular mesangial cell marker α-smooth muscle actin (αSMA) and was widely distributed in tubular epithelial cells, including the proximal tubule in both males and females. In females, ERα36 also co-localized with the podocyte-specific marker synaptopodin and the collecting duct principal cell marker AQP2; however, the data were inconclusive concerning its expression in vascular smooth muscle (not shown). ERα36 localization in males was similar to that evident in females, except for its failure to co-localize with synaptopodin in the glomerulus or α-SMA in vascular smooth muscle.
Figure 6. Representative immunofluorescence images localizing ERα36 in mouse kidney.

Green fluorescence represents ERα36 in all images. Red fluorescence represents cell specific markers for co-localization (collecting duct marker, aquaporin 2; vascular smooth muscle & mesangial cell marker, α-smooth muscle actin; podocyte cell marker, synaptopodin). Nuclei are stained blue with DAPI. Scale bar = 20 µm.
ERβ co-localized primarily with the podocyte-specific marker synaptopodin in females but not in males (Figure 7). ERβ was also present in the mesangial cells as evidenced by co-localization with αSMA, although the mesangial localization appeared to be more nuclear in the male kidneys. ERβ immunostaining was more prominent in proximal tubular cells than in distal nephron segments including the cortical collecting duct, and was present in vascular smooth muscle in both males and females (data not shown).
Figure 7. Representative immunofluorescence images localizing ERβ in mouse kidney.

Green fluorescence represents ERβ in all images. Red fluorescence represents cell specific markers for co-localization (collecting duct marker, aquaporin 2; vascular smooth muscle & mesangial cell marker, α-smooth muscle actin; podocyte cell marker, synaptopodin). Nuclei are stained blue with DAPI. Scale bar = 20 µm.
At the subcellular level, most of the renal ERα36 protein partitioned in the membrane fraction, with less than 10% in the cytoplasmic fraction (Figure 8). ERα46 was present in both the membrane and cytoplasmic fractions.
Figure 8. ERα46 and ERα36 protein level in membrane and cytosolic fractions from the renal cortices of female mice.
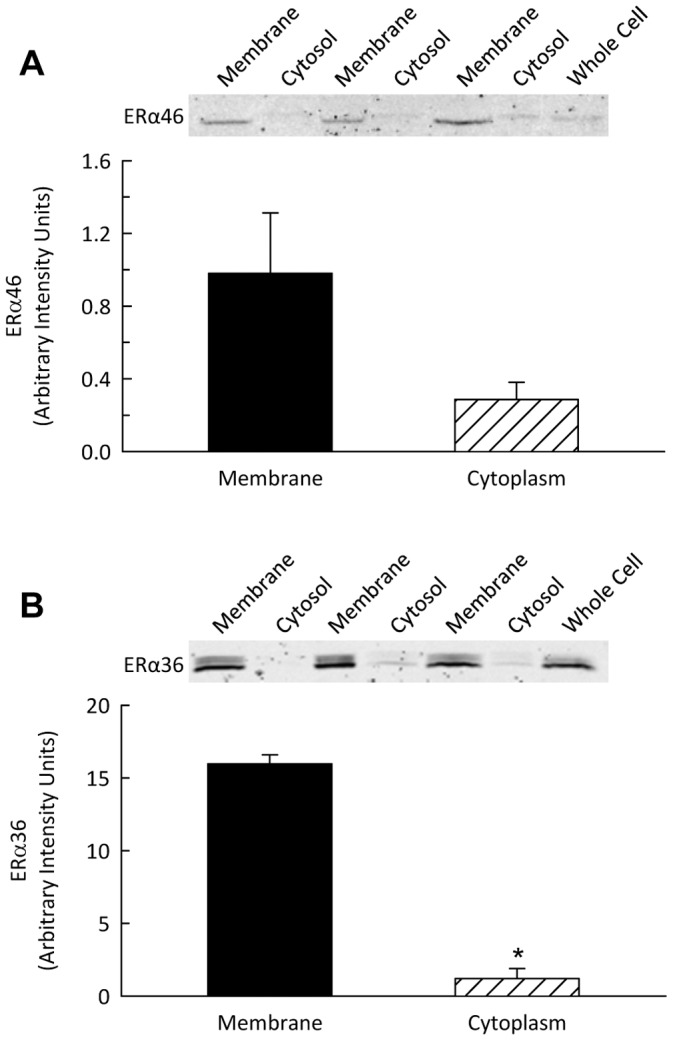
Shown are representative Western blots and quantification for ERα splice variant expression in membrane and cytosolic fractions from the renal cortices from female mice. *P<0.05 vs. membrane fraction.
Discussion
Expression of the various estrogen receptors differed widely among the organs of mice and also between males and females. The protein levels of ERα66 were low in all non-reproductive tissues. The highest expression was in the uterus and the ovaries. ERα66 expression was lower in males than in females in all the organs investigated, with levels in males averaging 70% of female levels in kidney, 6% of female values in liver, and 1% of female values in heart and lung. The bands on the Western blot were at a slightly higher molecular weight in the male and female kidney than the bands in other organs. The prominent band in the testis ran at a slightly lower molecular weight than that in the uterus and ovaries. This phenomenon has been reported previously by Baines [27], who suggested that this lower band was a degradation product. It is also possible that this represents a different isoform or that a post-translational modification present in the other tissues does not occur in the testis.
These data differ somewhat from previously reported results from Kuiper et al. [28], who detected high levels of ERα66 mRNA in the rat testis, male kidney and male adrenal gland, as well as the ovary and uterus. One explanation for the discrepancy is that their probes targeted the ERα DNA binding domain and, thus, would detect not only ERα66 but also ERα46 and ERα36. Additionally, the high levels of mRNA may not be translated into protein or may be rapidly degraded after translation and therefore not detectable by Western blot. It should also be noted that lower protein levels of ERα66 in the kidney do not necessarily equate with lack of activity. Indeed, despite its low relatively levels of expression in kidney, we and others have shown that loss of ERα66 results in physiologic changes in the kidney of female rodents [29]. This phenomenon likely reflects the ability of post-translational modification of proteins to alter function. In addition, post-translational modifications may interfere with antibody interaction at the epitope, thereby potentially impacting western blotting and immunofluorescence data.
In the present study using an antibody with an epitope in the N-terminus of the ERα66 protein, ERα66 was localized mainly to the vasculature and to the glomerulus. ERα66 was also found in the brush border of the proximal tubule and the apical aspect of collecting duct cells in female kidneys. The latter observations suggest a potential for ERα66 to modulate apical transport mechanisms in multiple segments of the nephron.
ERβ was present in all the tissues investigated. Males expressed ERβ levels similar to females in all organs except the kidney (25% of female value). These data again differ from the findings of Kuiper et al. [28], who did not detect ERβ mRNA in the male liver, kidney or heart. Similarly, Couse et al. [30] reported that ERβ mRNA was undetectable in the kidney of both male and female mice that were older than those used in the current study. Consistent with our observations, Catanuto et al. [31] have reported ERβ expression in the kidney. In a recent study in female rats, ERβ was detected in both the cortex and medulla [32]. The present studies revealed that ERβ was similarly expressed in the mesangial cells and vasculature of males and females. In contrast, prominent expression of ERβ was apparent in the podocytes of females but not males. Podocyte damage and apoptosis can underlie nephropathy and this sex difference in the expression of ERβ may contribute to increased severity of renal disease in males.
In both males and females, ERα46 protein level in the heart was approximately twice that found in the kidney. ERα46 was also highly expressed in the uterus and testes, but low in the mammary glands and ovaries. Recent studies have implicated ERα46 in both genomic and non-genomic estrogen signaling. ERα46 can inhibit transcription through the AF1 transactivation domain of ERα66, which may allow a proliferative phenotype [22]. Murphy et al. [33] demonstrated higher protein levels of ERα46 in differentiated macrophages than monocytes. Additionally, ERα46 in the membrane of endothelial cells can activate NOS3 [34]. Collectively these findings suggest that ERα46 can modulate the estrogenic effects of ERα66 in tissues that express both receptors, and that this occurs via both genomic and non-genomic mechanisms. Our observations suggest that this could occur in the heart and the uterus, which express both receptors; however, the extent of modulation might vary considerably due to the differences in the quantity of each receptor present in the tissue. This may be particularly true in the males that have low levels of ERα66 in the heart and testes. Esqueda et al. [32] reported a 46 kDa estrogen receptor in the cortex and medulla of female Dahl salt-sensitive and salt-resistant rats. Study of ERα46 is particularly difficult due to its identical protein sequence with ERα66. New methodologies need to be developed in order to differentiate the functional impact of ERα46 from that of ERα66.
ERα36 expression was low in all reproductive tissues investigated (mammary glands, ovaries, uterus and testes). ERα66 suppresses expression of ERα36 independent of estrogen, possibly through binding to the estrogen response element half site on the ERα36 promoter [35]. Therefore, it is not surprising that ERα36 expression is lowest in tissues known be predominantly regulated through ERα66. Even with low expression, however, ERα36 has been shown to be present in the plasma membrane of ovarian follicles and potentially influences non-genomic signaling [36].
The female kidney had the highest expression of ERα36, with the male kidney having the lowest expression of all organs studied (<1% of female kidney). ERα36 can exert a dominant negative effect by inhibiting ERα66 and ERβ transactivation through the AF1 and AF2 domains in the nucleus, but can also signal from the membrane [37]. This dominant negative role may be important in suppressing proliferative signaling through ERα66 in non-reproductive tissue. However, estrogen and estrogen receptor antagonists, such as tamoxifen and fulvestrant, can activate extracellular signal-regulated kinase (ERK1/2) signaling through ERα36 at the membrane, leading to proliferation [37]. This may suggest that genomic, rather than membrane-initiated, proliferative signaling is more prevalent in males.
In this study, the kidney was examined in greatest detail. Our findings suggest that the estrogen responsiveness of the female kidney may be primarily due to ligand binding to ERα36. In agreement with prior studies in cancer cells [23], ERα36 was distributed primarily in the membrane fraction of the renal cortex suggesting involvement in non-genomic signaling. Within the glomerulus, ERα36 co-localized with mesangial cells in both sexes and with podocytes in females only. We also found ERα36 present in the epithelial cells of the tubule. Thus, estrogen actions on a variety of renal cell types may involve non-genomic signaling via ERα36. The results of the present study also demonstrate sexual dimorphism of ER levels in the kidney, most strikingly with regard to the ERα36 splice variant but also for ERα46. Future studies are needed to elucidate the potential contribution of these receptors to sex differences in kidney function.
Pre-menopausal women have lower blood pressure than men, and the kidney is largely responsible for chronic regulation of blood pressure [38], [39]; however, the mechanism underlying this sexual dimorphism in blood pressure control is not completely understood. One obvious possibility is involvement of the vasodilator effect of estrogen, which arises through differential signaling mechanisms in various vascular beds of female rats [40]. Moreover, oxidative stress is more pronounced in healthy young men than in healthy, premenopausal women [41] and increases in women after menopause [42]. Estrogen receptor subtypes have been studied in the reduction of cardiovascular oxidative stress [12] and estradiol has been shown to attenuate superoxide production in an experimental model of hypertension [43]. In addition, male sex is a risk factor for nephropathy [44]. Estrogen has been shown to regulate matrix metalloproteinases which prevent collagen deposition in the glomerulus [45]–[47] and may contribute to the decreased risk in females. Fewer estrogen receptors in the male mouse kidney, particularly with regard to ERα66 and ERβ in podocytes, may render males more susceptible to apoptosis and glomerular injury. Finally, it is widely appreciated that female kidneys are smaller than male kidneys in the normal state, and evidence suggests that ERα66 is involved in compensatory renal growth after uninephrectomy in female mice [48]. Consistent with this observation, prominent proximal tubular brush border localization of this receptor could contribute to this phenomenon.
Viewed en masse, the results of the present study establish that estrogen receptor expression varies widely in different tissues. This new information expanding our understanding of which cells express each receptor can aid our interpretation of overall estrogen responsiveness in multiple tissues. In particular, the finding that the female kidney has the highest level of expression of ERα36 among the organs studied, in concert with an apparent sexual dimorphism in its intrarenal localization, should fuel future investigation to define the roles of this ERα splice variant in normal renal physiology and pathophysiology.
Acknowledgments
We would like to thank Dr. Zhao-Yi Wang (Creighton University Medical Center, Omaha, NE) for the generous gift of the ERα36 antibody. William J. Langer provided expert technical assistance.
Additionally, we thank Dr. Lie Gao for the use of the confocal microscope and cryostat.
Funding Statement
This work was funded by the University of Nebraska Medical Center Departments of Pediatrics and of Cellular and Integrative Physiology. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Allen E, Doisy EA (1983) Landmark article Sept 8, 1923. An ovarian hormone. Preliminary report on its localization, extraction and partial purification, and action in test animals. By Edgar Allen and Edward A. Doisy. JAMA 250: 2681–2683. [DOI] [PubMed] [Google Scholar]
- 2. Marrian GF, Butenandt A (1931) Oestrus-producing hormones. Science 74: 547 74/1926/547 [pii];10.1126/science.74.1926.547 [doi]. [DOI] [PubMed] [Google Scholar]
- 3. Green S, Walter P, Kumar V, Krust A, Bornert JM, et al. (1986) Human oestrogen receptor cDNA: sequence, expression and homology to v-erb-A. Nature 320: 134–139 10.1038/320134a0 [doi]. [DOI] [PubMed] [Google Scholar]
- 4. Lubahn DB, Moyer JS, Golding TS, Couse JF, Korach KS, et al. (1993) Alteration of reproductive function but not prenatal sexual development after insertional disruption of the mouse estrogen receptor gene. Proc Natl Acad Sci U S A 90: 11162–11166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Kuiper GG, Enmark E, Pelto-Huikko M, Nilsson S, Gustafsson JA (1996) Cloning of a novel receptor expressed in rat prostate and ovary. Proc Natl Acad Sci U S A 93: 5925–5930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Mosselman S, Polman J, Dijkema R (1996) ERβ: identification and characterization of a novel human estrogen receptor. FEBS Lett 392: 49–53. 0014-5793(96)00782-X [pii]. [DOI] [PubMed]
- 7. Krege JH, Hodgin JB, Couse JF, Enmark E, Warner M, et al. (1998) Generation and reproductive phenotypes of mice lacking estrogen receptor β. Proc Natl Acad Sci U S A 95: 15677–15682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Lindberg MK, Moverare S, Skrtic S, Gao H, Dahlman-Wright K, et al. (2003) Estrogen receptor (ER)-β reduces ERα-regulated gene transcription, supporting a “ying yang” relationship between ERα and ERβ in mice. Mol Endocrinol 17: 203–208. [DOI] [PubMed] [Google Scholar]
- 9.Wittnich C, Tan L, Wallen J, Belanger M (2013) Sex differences in myocardial metabolism and cardiac function: an emerging concept. Pflugers Arch. 10.1007/s00424-013-1232-1 [doi]. [DOI] [PubMed]
- 10. Sullivan JC, Pardieck JL, Brinson K, Kang KT (2009) Effects of estradiol on renal cyclic guanosine monophosphate and oxidative stress in spontaneously hypertensive rats. Gend Med 6: 498–510 S1550-8579(09)00095-3 [pii];10.1016/j.genm.2009.09.003 [doi]. [DOI] [PubMed] [Google Scholar]
- 11. Reckelhoff JF (2006) Cardiovascular disease, estrogen deficiency, and inflammatory cytokines. Hypertension 48: 372–373 01.HYP.0000235866.97871.9d [pii];10.1161/01.HYP.0000235866.97871.9d [doi]. [DOI] [PubMed] [Google Scholar]
- 12.Arias-Loza PA, Muehlfelder M, Pelzer T (2013) Estrogen and estrogen receptors in cardiovascular oxidative stress. Pflugers Arch. 10.1007/s00424-013-1247-7 [doi]. [DOI] [PubMed]
- 13.Li W, Zheng T, Altura BM, Altura BT (2001) Sex steroid hormones exert biphasic effects on cytosolic magnesium ions in cerebral vascular smooth muscle cells: possible relationships to migraine frequency in premenstrual syndromes and stroke incidence. Brain Res Bull 54: 83–89. S0361-9230(00)00428-7 [pii]. [DOI] [PubMed]
- 14.Schreihofer DA, Ma Y (2013) Estrogen receptors and ischemic neuroprotection: Who, what, where and when? Brain Res. S0006-8993(13)00330-2 [pii];10.1016/j.brainres.2013.02.051 [doi]. [DOI] [PubMed]
- 15. Gormsen LC, Host C, Hjerrild BE, Pedersen SB, Nielsen S, et al. (2012) Estradiol acutely inhibits whole body lipid oxidation and attenuates lipolysis in subcutaneous adipose tissue: a randomized, placebo-controlled study in postmenopausal women. Eur J Endocrinol 167: 543–551 EJE-12-0422 [pii];10.1530/EJE-12-0422 [doi]. [DOI] [PubMed] [Google Scholar]
- 16. Amengual-Cladera E, Llado I, Gianotti M, Proenza AM (2012) Sex differences in the effect of high-fat diet feeding on rat white adipose tissue mitochondrial function and insulin sensitivity. Metabolism 61: 1108–1117 S0026-0495(12)00009-1 [pii];10.1016/j.metabol.2011.12.016 [doi]. [DOI] [PubMed] [Google Scholar]
- 17. Fisher JS, Millar MR, Majdic G, Saunders PT, Fraser HM, et al. (1997) Immunolocalisation of oestrogen receptor-α within the testis and excurrent ducts of the rat and marmoset monkey from perinatal life to adulthood. J Endocrinol 153: 485–495. [DOI] [PubMed] [Google Scholar]
- 18. Saunders PT, Fisher JS, Sharpe RM, Millar MR (1998) Expression of oestrogen receptor beta (ERβ) occurs in multiple cell types, including some germ cells, in the rat testis. J Endocrinol 156: R13–R17. [DOI] [PubMed] [Google Scholar]
- 19. Joseph A, Hess RA, Schaeffer DJ, Ko C, Hudgin-Spivey S, et al. (2010) Absence of estrogen receptor alpha leads to physiological alterations in the mouse epididymis and consequent defects in sperm function. Biol Reprod 82: 948–957 biolreprod.109.079889 [pii];10.1095/biolreprod.109.079889 [doi]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Ruz R, Gregory M, Smith CE, Cyr DG, Lubahn DB, et al. (2006) Expression of aquaporins in the efferent ductules, sperm counts, and sperm motility in estrogen receptor-α deficient mice fed lab chow versus casein. Mol Reprod Dev 73: 226–237 10.1002/mrd.20390 [doi]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Pedram A, Razandi M, Kim JK, O’Mahony F, Lee EY, et al. (2009) Developmental phenotype of a membrane only estrogen receptor alpha (MOER) mouse. J Biol Chem 284: 3488–3495 M806249200 [pii];10.1074/jbc.M806249200 [doi]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Flouriot G, Brand H, Denger S, Metivier R, Kos M, et al. (2000) Identification of a new isoform of the human estrogen receptor-alpha (hER-α) that is encoded by distinct transcripts and that is able to repress hER-alpha activation function 1. EMBO J 19: 4688–4700 10.1093/emboj/19.17.4688 [doi]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Wang Z, Zhang X, Shen P, Loggie BW, Chang Y, et al. (2005) Identification, cloning, and expression of human estrogen receptor-α36, a novel variant of human estrogen receptor-α66. Biochem Biophys Res Commun 336: 1023–1027 S0006-291X(05)01951-0 [pii];10.1016/j.bbrc.2005.08.226 [doi]. [DOI] [PubMed] [Google Scholar]
- 24. Penot G, Le PC, Merot Y, Grimaud-Fanouillere E, Ferriere F, et al. (2005) The human estrogen receptor-α isoform hERα46 antagonizes the proliferative influence of hERα66 in MCF7 breast cancer cells. Endocrinology 146: 5474–5484 en.2005-0866 [pii];10.1210/en.2005-0866 [doi]. [DOI] [PubMed] [Google Scholar]
- 25. Taylor SE, Martin-Hirsch PL, Martin FL (2010) Oestrogen receptor splice variants in the pathogenesis of disease. Cancer Lett 288: 133–148 S0304-3835(09)00430-3 [pii];10.1016/j.canlet.2009.06.017 [doi]. [DOI] [PubMed] [Google Scholar]
- 26. Jelinsky SA, Harris HA, Brown EL, Flanagan K, Zhang X, et al. (2003) Global transcription profiling of estrogen activity: estrogen receptor α regulates gene expression in the kidney. Endocrinology 144: 701–710. [DOI] [PubMed] [Google Scholar]
- 27. Baines H, Nwagwu MO, Hastie GR, Wiles RA, Mayhew TM, et al. (2008) Effects of estradiol and FSH on maturation of the testis in the hypogonadal (hpg) mouse. Reprod Biol Endocrinol 6: 4 1477-7827-6-4 [pii];10.1186/1477-7827-6-4 [doi]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Kuiper GG, Carlsson B, Grandien K, Enmark E, Haggblad J, et al. (1997) Comparison of the ligand binding specificity and transcript tissue distribution of estrogen receptors α and β. Endocrinology 138: 863–870. [DOI] [PubMed] [Google Scholar]
- 29. Lovegrove AS, Sun J, Gould KA, Lubahn DB, Korach KS, et al. (2004) Estrogen receptor α-mediated events promote sex-specific diabetic glomerular hypertrophy. Am J Physiol Renal Physiol 287: F586–F591 10.1152/ajprenal.00414.2003 [doi];00414.2003 [pii]. [DOI] [PubMed] [Google Scholar]
- 30. Couse JF, Lindzey J, Grandien K, Gustafsson JA, Korach KS (1997) Tissue distribution and quantitative analysis of estrogen receptor-alpha (ERα) and estrogen receptor-beta (ERβ) messenger ribonucleic acid in the wild-type and ERalpha-knockout mouse. Endocrinology 138: 4613–4621. [DOI] [PubMed] [Google Scholar]
- 31. Catanuto P, Doublier S, Lupia E, Fornoni A, Berho M, et al. (2009) 17β-estradiol and tamoxifen upregulate estrogen receptor β expression and control podocyte signaling pathways in a model of type 2 diabetes. Kidney Int 75: 1194–1201 ki200969 [pii];10.1038/ki.2009.69 [doi]. [DOI] [PubMed] [Google Scholar]
- 32. Esqueda ME, Craig T, Hinojosa-Laborde C (2007) Effect of ovariectomy on renal estrogen receptor-α and estrogen receptor-β in young salt-sensitive and -resistant rats. Hypertension 50: 768–772 HYPERTENSIONAHA.107.095265 [pii];10.1161/HYPERTENSIONAHA.107.095265 [doi]. [DOI] [PubMed] [Google Scholar]
- 33. Murphy AJ, Guyre PM, Wira CR, Pioli PA (2009) Estradiol regulates expression of estrogen receptor ERα46 in human macrophages. PLoS One 4: e5539 10.1371/journal.pone.0005539 [doi]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Li L, Haynes MP, Bender JR (2003) Plasma membrane localization and function of the estrogen receptor alpha variant (ER46) in human endothelial cells. Proc Natl Acad Sci U S A 100: 4807–4812 10.1073/pnas.0831079100 [doi];0831079100 [pii]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Zou Y, Ding L, Coleman M, Wang Z (2009) Estrogen receptor-alpha (ER-α) suppresses expression of its variant ER-α 36. FEBS Lett 583: 1368–1374 S0014-5793(09)00239-7 [pii];10.1016/j.febslet.2009.03.047 [doi]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Chakraborty P, Roy SK (2013) Expression of estrogen receptor α36 (ESR36) in the hamster ovary throughout the estrous cycle: effects of gonadotropins. PLoS One 8: e58291 10.1371/journal.pone.0058291 [doi];PONE-D-12-38435 [pii]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Wang Z, Zhang X, Shen P, Loggie BW, Chang Y, et al. (2006) A variant of estrogen receptor-α, hER-α36: transduction of estrogen- and antiestrogen-dependent membrane-initiated mitogenic signaling. Proc Natl Acad Sci U S A 103: 9063–9068 0603339103 [pii];10.1073/pnas.0603339103 [doi]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Dubey RK, Jackson EK (2001) Estrogen-induced cardiorenal protection: potential cellular, biochemical, and molecular mechanisms. Am J Physiol Renal Physiol 280: F365–F388. [DOI] [PubMed] [Google Scholar]
- 39. Reckelhoff JF (2005) Sex steroids, cardiovascular disease, and hypertension: unanswered questions and some speculations. Hypertension 45: 170–174. [DOI] [PubMed] [Google Scholar]
- 40.Reslan OM, Yin Z, do Nascimento GR, Khalil RA (2013) Subtype-specific estrogen receptor-mediated vasodilator activity in the cephalic, thoracic and abdominal vasculature of female rat. J Cardiovasc Pharmacol. 10.1097/FJC.0b013e31828bc88a [doi]. [DOI] [PMC free article] [PubMed]
- 41. Ide T, Tsutsui H, Ohashi N, Hayashidani S, Suematsu N, et al. (2002) Greater oxidative stress in healthy young men compared with premenopausal women. Arterioscler Thromb Vasc Biol 22: 438–442. [DOI] [PubMed] [Google Scholar]
- 42. Helmersson J, Mattsson P, Basu S (2002) Prostaglandin F2α metabolite and F2-isoprostane excretion rates in migraine. Clin Sci (Lond) 102: 39–43. [PubMed] [Google Scholar]
- 43.Ji H, Zheng W, Menini S, Pesce C, Kim J, et al.. (2007) Female protection in progressive renal disease is associated with estradiol attenuation of superoxide production. Gend Med 4: 56–71. S1550-8579(07)80009-X [pii]. [DOI] [PubMed]
- 44. Neugarten J, Acharya A, Silbiger SR (2000) Effect of gender on the progression of nondiabetic renal disease: a meta-analysis. J Am Soc Nephrol 11: 319–329. [DOI] [PubMed] [Google Scholar]
- 45. Potier M, Elliot SJ, Tack I, Lenz O, Striker GE, et al. (2001) Expression and regulation of estrogen receptors in mesangial cells: influence on matrix metalloproteinase-9. J Am Soc Nephrol 12: 241–251. [DOI] [PubMed] [Google Scholar]
- 46. Guccione M, Silbiger S, Lei J, Neugarten J (2002) Estradiol upregulates mesangial cell MMP-2 activity via the transcription factor AP-2. Am J Physiol Renal Physiol 282: F164–F169 10.1152/ajprenal.00318.2000 [doi]. [DOI] [PubMed] [Google Scholar]
- 47. Dixon A, Maric C (2007) 17β-Estradiol attenuates diabetic kidney disease by regulating extracellular matrix and transforming growth factor-β protein expression and signaling. Am J Physiol Renal Physiol 293: F1678–F1690 00079.2007 [pii];10.1152/ajprenal.00079.2007 [doi]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Sun J, Langer WJ, Devish K, Lane PH (2006) Compensatory kidney growth in estrogen receptor-α null mice. Am J Physiol Renal Physiol 290: F319–F323 00271.2005 [pii];10.1152/ajprenal.00271.2005 [doi]. [DOI] [PubMed] [Google Scholar]