

Abstract
A phase II study of NK cell therapy in treatment of patients with recurrent breast cancer has recently been reported. However, because of the complexities of tumor microenvironments, effective therapeutic effects have not been achieved in NK cell therapy. Radioiodine (I-131) therapy inhibits cancer growth by inducing the apoptosis and necrosis of cancer cells. Furthermore, it can modify cancer cell phenotypes and enhance the effect of immunotherapy against cancer cells. The present study showed that I-131 therapy can modulate microenvironment of breast cancer and improve the therapeutic effect by enhancing NK cell cytotoxicity to the tumor cells. The susceptibility of breast cancer cells to NK cell was increased by precedent I-131 treatment in vitro. Tumor burden in mice treated with I-131 plus NK cell was significantly lower than that in mice treated with NK cell or I-131 alone. The up-regulation of Fas, DR5 and MIC A/B on irradiated tumor cells could be the explanation for the enhancement of NK cell cytotoxicity to tumor cells. It can be applied to breast cancer patients with iodine avid metastatic lesions that are non-responsive to conventional treatments.
Introduction
Breast cancer is the most common cancer and the second most common cause of cancer-related death in women, with more than one million cases and nearly 600,000 deaths occurring annually worldwide [1]. Breast cancer is characterized by a distinct pattern of metastasis involving regional lymph nodes, bone, lung, and liver, and the distant metastasis is closely associated with poor prognosis [2]. Once breast cancer has metastasized, it is usually not cured by current therapies, including high dose chemotherapy, likely due to subpopulations of slow-dividing chemoresistant cells present in metastatic cells [3]. In addition, the triple negative breast cancers lack a therapeutic target and have a poor prognosis [4]. Therefore, establishment of new therapeutic strategies is crucial to improving the prognosis of advanced breast cancer.
Tumor-specific immunotherapy offers considerable potential in management of patients with breast cancer; one of the effective immunotherapies is use of natural killer (NK) cells [5]. NK cells, a subset of lymphocytes capable of mediating cytotoxicity against tumor cells and virally infected cells, constitute a key component of the innate immune system [6]. NK cells have been shown to play an important role in controlling the growth of various tumor cell lines in mice [6], [7]. A phase II study of NK cell therapy in treatment of patients with recurrent breast cancer has recently been reported [8]. However, because of the complexities of tumor microenvironments, effective therapeutic effects have not been achieved in NK cell therapy [9], [10].
Radioiodine (I-131) therapy inhibits cancer growth through induction of apoptosis and necrosis of cancer cells [11]. In addition, it was shown that I-131 therapy can modify cancer cell phenotypes and enhance the effect of immunotherapy against cancer cells [12]–[14]. In particular, irradiated cancer cells show up-regulated levels of Fas and tumor necrosis factor–related apoptosis inducing ligand (TRAIL) receptor [15]–[18]. In breast cancer, I-131 therapy can be used, as the majority of breast cancer (70–80%) expresses human sodium/iodide symporter (hNIS), which is a specialized active iodide transporter [11], [18], [19]. hNIS expression and I-131 uptake by breast cancer cells has been suggested to provide supportive evidence for use of I-131 as an additional modality for treatment of breast cancer [20]. Therefore, it is expected that pretreatment with I-131 will result in modification of cancer cell phenotypes and enhance the susceptibility of breast cancer cells to NK cell cytotoxicity.
In this study, we attempted to determine whether Fas and TRAIL receptors of breast cancer cells are up-regulated by I-131 therapy and whether I-131 therapy can enhance NK cell cytotoxicity in vitro and in vivo.
Materials and Methods
Cell Lines
The NK92-MI human NK cell line (NK92-MI) was obtained from the American Type Culture Collection (Rockville, MD, USA). NK92-MI cells were incubated in alpha modification of Eagle’s minimum essential medium (α-MEM; Invitrogen, Carlsbad, CA, USA) supplemented with 2 mM L-glutamine, 0.2 mM inositol, 0.02 mM folic acid, 0.01 mM 2-mercaptoethanol, 10% fetal bovine serum (FBS; Gemini Bio-Products, West Sacramento, CA, USA), and 10% horse serum (Sigma-Aldrich Corporation, St Louis, MO, USA). Human breast cancer cell line MDA-MB 231 (MDA-231) was purchased from the American Type Culture Collection. The MDA-MB 231 cell line coexpressing hNIS and enhanced firefly luciferase (effluc) genes (MDA-231/NF) was kindly provided by J.K Chung (Seoul National University, Seoul, Republic of Korea). MDA-231/NF cells were maintained in Roswell Park Memorial Institute medium (RPMI-1640; Sigma-Aldrich Corporation) supplemented with 10% FBS and 100 U/mL penicillin/streptomycin.
Animals
Specific pathogen-free six-week-old female BALB/c nude mice (Hamamatsu, Shizuoka, Japan) were used in in vivo study. All animal experiment protocols were conducted in accordance with National Institutes of Health guidelines for the care and use of laboratory animals and approved by the Committee for the Handling and Use of Animals of Kyungpook National University.
RT-PCR Analysis for hNIS and Effluc Genes
MDA-231 and MDA-231/NF cells and homogenized human thyroid tissue were lysed using a Trizol solution (Invitrogen), and total RNA was extracted according to the manufacturer’s instructions. Reverse transcription was performed using a RevertAid First Strand cDNA Synthesis kit (Fermentas, Ontario, Canada). After denaturation of the samples for 1 min at 94°C, 30 cycles for 25s at 94°C, 30 s at 57°C, and 30 s at 72°C were followed with an additional 10 min at 72°C. Two units of Taq DNA polymerase (Takara, Shiga, Japan) using a GeneAmp PCR system (Bio-Rad, Hercules, CA, USA) and the following primers were used: hNIS gene, forward: 5′-CTACGAGTACCTGGAGATGC-3′, reverse: 5′-GTCGCAGTCAGTGTAGAACA-3′; GAPDH, forward: 5′-GCCAAAAGGGTCATCATCTC-3′, reverse: 5′-GATGAGGCAGGGATGATGTT-3′; effluc gene, forward: 5′-GCACAAGGCCATGAAGAGAT-3′, reverse: 5′-CTTCTTGCTCACGAACACCA-3′. Samples were separated by electrophoresis in an ethidium bromide-stained agarose gel.
I-125 Uptake Assay
The day before iodine uptake, 1×105, 2×105, 4×105, and 8×105 MDA-231 and MDA-231/NF cells were plated in 24-well plates. The level of I-125 uptake was determined by incubation of the cells with 500 µl of Hank’s balanced salt solution (HBSS) containing 0.5% bovine serum albumin (bHBSS), 3.7 kBq carrier-free I-125, and 10 µmol/L sodium iodide (specific activity of 740 MBq/mmol) at 37°C for 30 min. The blocking control study for hNIS was performed in an identical manner, with the exception of the addition of 1mM KClO4 to the incubation buffer. After incubation, the cells were washed twice as quickly as possible with ice-cold bHBSS buffer and detached using 500 µL of trypsin. Radioactivity was measured using a gamma-counter (CobraII, Packard, Perkin Elmber, Waltham, MA, USA).
To evaluate the functional expression of the hNIS gene in vivo, MDA-231/NF cells (5×105) in phosphate buffered saline (PBS) were implanted subcutaneously into the right flank of three mice. Fourteen days after tumor implantation, Tc-99m pertechnetate SPECT/CT scan was performed using the Inveon small animal imaging system (Siemens Medical Solutions, Knoxville, TN, USA). The mice were placed in a spread-prone position at 40 min after injection of Tc-99m pertechnetate (7.4 MBq/0.2 mL of 0.9% NaCl) into the tail vein, and scanned for 20 min. A 20% window was centered at the 140 keV photopeak of Tc-99m. The 3-D ordered subset expectation–maximization (OSEM) algorithm was used in reconstruction. The voxel size of the image matrix was 0.5×0.5×0.5 mm. All reconstructed images were normalized using a correction matrix derived from a uniform cylindrical phantom image prior to reconstruction.
Luciferase Assay
In examination of the luciferase assay in vitro, 1×105, 2×105, 4×105, and 8×105 MDA-231 and MDA-231/NF cells were plated in 96-well white plates and cultured with Dulbecco’s modified eagle medium (DMEM; Sigma-Aldrich Corporation, St Louis, MO, USA) containing 10% FBS. After 24 hr incubation, 6 µL of D-luciferin (30 mg/mL) was added to each well, followed by measurement of bioluminescence using a microplate luminometer (Molecular Devices, Sunnyvale, CA, USA).
To evaluate the functional expression of the effluc gene in vivo, MDA-231/NF cells in PBS were implanted subcutaneously into the right hind-flank (1×105), left hind-flank (3×105), and right fore-flank (9×105) of three mice. After tumor implantation, bioluminescence imaging was performed using the IVIS lumina II imaging system (Caliper, Alameda, CA, USA), which included a highly sensitive CCD camera mounted on a light-tight specimen chamber.
Phenotype Marker Analysis
To determine the levels of Fas, TRAIL receptor2 (DR5), major histocompatibility complex (MHC) class I chain-related molecule A and B (MIC A/B), and human leukocyte antigen -A, B, and C (HLA-ABC) expression in cancer cells, MDA-231/NF cells were grown in 75 cm2 flasks and incubated for 7 hr at 37°C in HBSS only or in HBSS containing 14.8 MBq/5 mL of I-131. The reaction was terminated by removal of the medium containing the radioisotope and by washing cells twice with HBSS. Cells were then grown for three days. These cells were then stained with phycoerythrin (PE)-anti-Fas (BD Biosciences, San Jose, CA, USA), PE-anti-DR5 (eBioscience, San Diego, CA, USA), PE-anti-MIC A/B (BD Biosciences), and PE-anti-HLA-ABC (BD Biosciences). Flow cytometric analysis was performed using a Becton Dickinson FACScan unit using CELL Quest software (Becton Dickinson Immunocytometry Systems, San Jose, CA, USA).
Cytotoxicity Assay
Cytotoxic activity of NK92-MI cells was assessed using the Calcein-AM release test. Calcein-AM was purchased from Invitrogen as a 1 mg/mL solution in dimethyl sulfoxide (DMSO; Sigma, St. Louis, MO, USA). MDA-231 and MDA-231/NF cells were grown in 75 cm2 flasks and incubated for 7 hr at 37°C in HBSS only or in HBSS containing 14.8 MBq of I-131. After 7 hr, I-131 containing medium was removed and cells were washed twice with HBSS. Cells were grown for three days. Then, irradiated or non-irradiated MDA-231/NF cells were re-suspended in the complete medium at a final concentration of 106 cells/mL and were incubated with 5 µM Calcein-AM for 20 min at 37°C, allowing Calcein-AM to enter the target cells. The labeled target cells were then washed twice and dispensed at a concentration of 1×104 target cells/well in a round-bottom, 96-well plate (Nunc, Roskilde, Denmark). The effectors were distributed in triplicate at effector : target (E : T) cell ratios from 2.5 : 1, 5 : 1, and 10 : 1 with at least three replicate wells for spontaneous (only target cells in complete medium) and maximum release (only target cells in medium plus 1% NP-40 cell lysis buffer). After incubation at 37°C in 5% CO2 for 4 hr, each supernatant was harvested and transferred into new plates. Samples were measured using a multilabelcounter (VICTOR3, Perkin Elmer, San Diego, CA, USA) (excitation filter: 485±9 nm; band-pass filter: 530±9 nm). Data were expressed as arbitrary fluorescent units. NK cell cytotoxicity was calculated using the following equation:
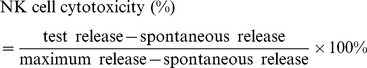 |
In vivo Animal Experiments
Twelve mice were divided into four groups for assessment of therapeutic effects (three mice per group); the experimental groups were referred to as the control, I-131, NK, and combined groups. In 12 mice, MDA-231/NF cells (5×105) were implanted subcutaneously into the right flank.
In the control group, intravenous injection of PBS was administered at 14 days post-challenge. In the I-131 group, intraperitoneal injection of 29.6 MBq of I-131 was administered at 14 days post-challenge. In the NK group, NK92-MI cells (5×106) were injected intravenously via tail vein at 17 and 18 days. A total of two doses were administered to each mouse with two days apart. The combined group received treatment with both I-131 at 14 days and NK92-MI cells at 17 and 18 days.
Bioluminescence imaging was performed using the IVIS lumina II imaging system (Caliper). From 14, 24, and 34 days post-challenge, mice received intraperitoneal injection with 100 µL of D-luciferin (30 mg/mL). After 5 min, mice were placed individually in the specimen chamber and images were then acquired. Grayscale photographic images and bioluminescent color images were superimposed using LIVING IMAGE, version 2.12 (Caliper, Alameda, CA, USA), and IGOR image analysis FX software (WaveMetrics, Lake Oswego, OR, USA). Bioluminescent signals were expressed in units of photons per cm2 per second per steradian (P/cm2/sec/sr).
Statistical Analysis
All data are expressed as means ± SDs and are representative of at least two separate experiments. The unpaired Student’s t test and ANOVA analysis were used for determination of statistical significance. P values of <0.05 were considered statistically significant.
Results
Verification of MDA-231 Expressing hNIS and Effluc Genes
Expression of hNIS and effluc genes of MDA-231/NF cells was confirmed by RT-PCR analysis. Human thyroid tissue was used as positive control for hNIS expression in MDA-231/NF cells. RT-PCR revealed hNIS mRNA expression in MDA-231/NF and human thyroid tissue. RT-PCR fragments had lengths of 583 bp and 316 bp for hNIS and effluc in MDA-231/NF cells, however, these bands did not appear in MDA-231 cells (Figure 1).
Figure 1. RT-PCR analysis of human sodium/iodide symporter (hNIS) and enhanced firefly luciferase (effluc) gene expression in MDA-231, MDA-231/NF cells and human thyroid tissue.

RT-PCR revealed hNIS mRNA expression in MDA-231/NF cells and human thyroid tissue, and effluc mRNA expression in MDA-231/NF cells. RT-PCR fragments have lengths of 583 bp and 316 bp for hNIS and effluc in MDA-231/NF cells; however, these bands do not appear in MDA-231 cells.
In vitro I-125 uptake assay showed that I-125 uptake by MDA-231/NF cells increased according to cell number, whereas I-125 uptake by MDA-231 cells and MDA-231/NF cells blocked by KClO4 remained at the basal level (Figure 2A). I-125 uptake in MDA-231/NF cells was 17-fold higher than the uptake observed in MDA-231 cells. The presence of 1mM KClO4 inhibited I-125 uptake completely in MDA-231/NF cells. In vitro luciferase assay was performed for MDA-231/NF and MDA-231 cells. Bioluminescence signals of MDA-231/NF cells increased according to cell number, whereas bioluminescence signals of MDA-231 cells remained at background level (Figure 2B). The signal intensity was approximately 1,180-fold higher in MDA-231/NF cells than in MDA-231 cells.
Figure 2. In vitro I-125 uptake assay and luciferase assay in MDA-231 and MDA-231/NF cells.

(A) I-125 uptake by MDA-231/NF cells increased according to cell number. I-125 uptake by MDA-231 cells remained at the basal level. *** p<0.001 compared with MDA-231 and MDA-231/NF cells blocked by KClO4. (B) Bioluminescence signals of MDA-231/NF cells increased according to cell number. Bioluminescence signal of MDA-231 cells remained at the basal level. CPM: count per minute, RLU: Relative Light Units, *** p<0.001 compared with MDA-231 cells.
To evaluate the functional expression of the hNIS gene in a tumor xenograft, Tc-99m pertechnetate SPECT/CT scan was performed in a mouse animal model. Focal tracer uptake was observed in the right flank of the MDA-231/NF tumor xenograft (Figure 3). To evaluate the functional expression of the effluc gene in the tumor xenograft, bioluminescence imaging was performed in a mouse animal model. Bioluminescence signals from implanted MDA-231/NF cells were clearly visualized in the right fore-flank and bilateral hind-flanks. The signal intensity from tumor cells increased with an increasing number of implanted cells.
Figure 3. Characterization image of MDA-231/NF cells.

(A–C) Tc-99m pertechnetate SPECT/CT images show focal tracer uptake (arrow) in the right flank of the MDA-231/NF tumor xenograft. There is also noted tracer uptake at the stomach (arrow head). (D–F) In vivo bioluminescence imaging for a mouse with MDA-231/NF cell implantation. (D) MDA-231/NF cells were implanted subcutaneously into the right hind-flank (1×105; arrow head), left hind-flank (3×105; double arrow head), and right fore-flank (9×105; arrow) of the mouse. (E) Bioluminescence signals from implanted MDA-231/NF cells are clearly visualized in the right hind-flank (arrow head), left hind-flank (double arrow head), and right fore-flank (arrow). (F) The signal intensity from tumor cells increased with increasing cell number. * p<0.05 compared with 1×105 cells, *** p<0.001 compared with 1×105 and 3×105 cells.
Phenotypic Modulation of Cancer Cells by I-131 Therapy in vitro
To investigate the effect of I-131 therapy at surface levels of Fas, DR5, MIC A/B, and HLA-ABC of MDA-231/NF cells in vitro, flow cytometric analysis was performed in cells that received I-131 or PBS. The levels of Fas and DR5 were significantly higher in irradiated cells than in non-irradiated cells, respectively (15.1% vs. 6.0% and 44.0% vs. 20.3%; p = 0.027 and p<0.001). In addition, the level of MIC A/B was significantly higher in irradiated cells than in non-irradiated cells (13.1% vs. 3.9%; p = 0.004) (Figure 4). Although statistically not significant, the level of HLA-ABC was lower in irradiated cells than in non-irradiated cells (27.8% vs. 37.0%; p = 0.061).
Figure 4. Phenotype analysis in MDA-231/NF cells by flow cytometry.

(A, B and C) The levels of DR5, Fas, and MIC A/B expression in tumor cells that received I-131 were significantly higher than in tumor cells received. (D) I-131 therapy resulted in decreased expression of HLA-A,B,C, but without statistical significance. * p<0.05, ** p<0.005, *** p<0.001 compared with control.
Enhancement of NK Cell Cytotoxicity by I-131 Therapy in vitro
To determine whether I-131 therapy can enhance NK cell cytotoxicity to breast cancer cells, the cytotoxicity assay was performed in MDA-231/NF cells that received I-131 or PBS at E : T cell ratios of 2.5 : 1, 5 : 1, and 10 : 1. Cytotoxicity to the target cells was significantly increased by high E : T ratio in both non-irradiated and irradiated MDA-231/NF cells (p<0.001 and p = 0.001). NK cell cytotoxicities to non-irradiated cells were 2.6%, 6.0%, and 8.4% at an E : T ratio of 2.5 : 1, 5 : 1, and 10 : 1. NK cell cytotoxicities to the irradiated cells were 6.4%, 9.9%, and 17.5% at an E : T ratio of 2.5 : 1, 5 : 1, and 10 : 1 respectively (Figure 5). NK cell cytotoxicities were significantly higher in irradiated cells than in non-irradiated cells at all E : T ratios of 2.5 : 1, 5 : 1, and 10 : 1, respectively (p = 0.020, p = 0.001 and p = 0.003).
Figure 5. In vitro Cytotoxicity assay using Calcein-AM.

NK cell cytotoxocity is significantly increased according to effector : target cell (E : T) ratio in the control and I-131 groups. NK cell cytotoxicities were significantly higher in irradiated cells than in non-irradiated cells at all E : T ratios of 2.5 : 1, 5 : 1, and 10 : 1. * p<0.05, ** p<0.005 compared with control.
Enhancement of NK Cell Cytotoxicity by I-131 Therapy in vivo
The mice were divided into four groups (control, I-131, NK, and combined groups) and PBS, I-131, NK92-MI or I-131 and NK92-MI were applied to each group. Tumor burden was monitored by bioluminescence imaging 14, 24, and 34 days after tumor inoculation.
Mice in the control group who received PBS showed a continuous increase in their tumor signal. The I-131 and NK groups showed moderate increases in tumor signals and the tumor signals of the I-131 and NK groups were consistently lower than that of the control group and significantly lower than that of the control group at 34 days, respectively (p = 0.017 and p = 0.001). The combined group showed a stationary tumor signal over time, which was consistently lower than that of the I-131 and NK groups, and significantly lower than that of the I-131 and NK groups at 34 days, respectively (p = 0.030 and p = 0.038) (Figure 6). Tumor burdens measured by bioluminescence imaging did not differ significantly between the NK and I-131 groups (p = 0.155).
Figure 6. Tumor burdens monitored by bioluminescence imaging 14, 24, and 34 days after tumor inoculation.

(A and B) The tumor signals in the I-131 and NK groups were significantly lower than those in the control group at 34 days. The combined group showed the lowest stationary tumor signal over time, which was significantly lower than that in the I-131 and NK groups at 34 days. * p<0.05 compared with I-131 and NK groups,** p<0.005 compared with control group.
Discussion
Contemporary management of breast cancer with early detection, newer local control techniques, improved chemotherapy regimens, and targeted treatments has resulted in immense gains in survival in individuals with breast cancer [21]. Unfortunately, the triple negative breast cancers, a subset of breast cancers clinically defined by the absence of the estrogen receptor, progesterone receptor, and HER-2 over expression, have a higher propensity to metastasize to distant visceral organs, and have a worse outcome with a high rate of recurrence after adjuvant treatment [4]. Thus, the need for development of successful therapeutic options in an attempt to improve the outcome is urgent. An attractive approach to reducing the rate of recurrences in these individuals is use of immunotherapeutic strategies [6], [22]. However, because of the complexities of tumor microenvironments, effective therapeutic effects may not be feasibly achieved in immunotherapy. Several researchers have reported obstacles to successful immunotherapy, such as tumor-derived suppression cytokines, the absence of danger signals, loss of MHC class molecules, and low antigen levels [22], [23]. To overcome these impediments, modulation of the tumor microenvironment is essential. The current study showed that I-131 therapy can modulate microenvironment of breast cancer and improve the therapeutic effect through enhancement of NK cell cytotoxicity to tumor cells.
NK cells comprise 10–15% of all circulating lymphocytes and are also found in peripheral tissues, including the liver, peritoneal cavity, and placenta. Resting NK cells circulate in the blood; however, following activation by cytokines, they are capable of extravasation and infiltration into most tissues that contain pathogen infected or malignant cells [24]. NK92 is a human NK cell line first established in 1994 from a 50-year-old male patient with an aggressive NK cell lymphoma [25]. The NK92 cell line has been examined clinically as a treatment for advanced sarcoma and leukemia [25], [26]. The parental NK92 cell line is highly dependent on the cytokine IL-2 and therapies involving these cells in vivo require superphysiological amounts of IL-2. However, an IL-2 independent cell line, NK92-MI, which has been shown to be virtually identical to the parental cell line, may be a more appropriate choice for clinical therapies [26], [27]. Nagashima et al. [28] reported that NK92-MI sustained proliferation in the absence of exogenously supplied IL-2 and showed greater in vivo anti-tumor activity in mice. The current study also showed that NK92-MI have anti-tumor activity to a breast cancer cell line in vitro and in vivo without IL-2 supplementation.
The cytotoxicity of NK cells are carried out by two main mechanisms. The first mechanism is granule-dependent cytotoxicity, where upon triggering by activating receptors [6]. Upon recognition of the ligands on the surface of the target cell surface by activating NK cell receptors, various intracellular signaling pathways drive NK cells toward cytotoxic action, which results in cytolysis of target cells [29]. When NK cells are activated by MIC A/B, which are ligands for the activating receptor NKG2D on the tumor surface, perforin and granzyme B are released to the tumor cell, resulting in mediation of apoptosis [30]. However, these processes are tightly controlled by a group of inhibitory receptors. These receptors act as negative regulators of NK cytotoxicity and inhibit the action of NK cells against ‘self’ targets. A main group of this type of receptors is NK cell immunoglobulin-like receptors (KIRs), which are mainly specific for self MHC Class-I molecules. Members of the KIR family recognize HLA-A, B and C alleles [31]. The second mechanism is the triggering of apoptosis pathways in the target cell via stimulation of death receptors by TRAIL or Fas ligand expressed on the surface of NK cells as well as secretion of TNF-α. NK cells express Fas ligand and TRAIL, which are both members of the TNF family and have been shown to induce target cell apoptosis when they bind their receptors on target cells [32]. In the current study, the levels of surface expression of Fas, DR5, and MIC A/B showed an increase on cells treated with I-131 in in vitro study. Up-regulation of Fas, DR5, and MIC A/B on the surface of irradiated tumor cells could explain the enhancement of NK cell cytotoxicity to tumor cells in both in vitro and in vivo studies.
Irradiation can alter immunogenecity of the tumor as well as the circumstances of the immunologic condition. Several groups have reported that radiation therapy concomitantly up-regulates the levels of Fas, DR5, and MIC A/B in several tumor cells. Ishikawa et al. [13] reported that external radiation therapy enhanced Fas and DR5 expression in glioma cell lines and cytotoxicity of NK cells was enhanced after radiation therapy. Zhou et al. [17] reported that DR5 expression was enhanced in melanoma cell lines by external radiation therapy and treatment with TRAIL resulted in significantly increased tumor cell apoptosis caused by radiation therapy. Xu et al. [33] also reported that radiation therapy up-regulated the level of MIC A/B and increased the sensitivity of NK cell killing in a pancreatic cancer cell line. However, external radiation therapy is limited in treatment of multiple metastatic lesions. Like external radiation therapy, I-131 therapy can also alter immunogenicity of tumor cells by providing radiation to the cells. In addition, I-131 therapy is applicable to treatment of multiple metastatic breast cancer, which can take up I-131 by hNIS expression [34]. Jeon et al. [15] reported that I-131 therapy can lead to up-regulated expression of Fas and enhance the killing activities of cytotoxic T cells in a colon cancer cell line. In accordance with previous studies, the present study demonstrated that I-131 therapy up-regulated the level of Fas, DR5 and MIC A/B expression in breast cancer cell line. Because the majority of breast cancers are known to express hNIS and take up iodide, it would be applicable in the clinical setting [19].
A limitation of the current study is the use of MDA-231/NF cells expressing hNIS, instead of parental MDA-231 cells, for I-131 therapy, and it is not straightforward in clinical practice. Expression of hNIS is known to be enhanced by certain agents such as a retinoic acid, and enhancement by hNIS inducible agents has been well investigated in breast cancer cells [35]–[37]. hNIS expression in breast cancer cells by the inducible agents (not by transduction of the hNIS gene) would be more appropriate in the clinical field. For clinical translation of this strategy, conduct of further studies on enhancement of NK cell cytotoxicity to parental breast cancer cells with hNIS inducible agents followed by I-131 therapy is needed.
Conclusion
I-131 therapy would lead to up-regulation of the level of death receptors in breast cancer cells and improve therapeutic efficiency of NK cell therapy through enhancement of the cytotoxic effect of NK cells to the cancer. Enhancement of NK cell cytotoxicity by I-131 pretreatment can be applied to breast cancer patients with iodine avid metastatic lesions that are non-responsive to conventional treatments.
Funding Statement
This research was supported by National Nuclear R&D Programs through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (No.2012M2A2A7014020, 2009-0078234) and by a grant of the Korea Health Technology R&D Project, Ministry of Health & Welfare, Republic of Korea (A111345, A091224). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. DeSantis C, Siegel R, Bandi P, Jemal A (2011) Breast cancer statistics, 2011. CA Cancer J Clin 61: 409–418. [DOI] [PubMed] [Google Scholar]
- 2. Ugnat AM, Xie L, Morriss J, Semenciw R, Mao Y (2004) Survival of women with breast cancer in Ottawa, Canada: variation with age, stage, histology, grade and treatment. Br J Cancer 90: 1138–1143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Nieto Y (2003) The verdict is not in yet. Analysis of the randomized trials of high-dose chemotherapy for breast cancer. Haematologica 88: 201–211. [PubMed] [Google Scholar]
- 4. Dent R, Trudeau M, Pritchard KI, Hanna WM, Kahn HK, et al. (2007) Triple-negative breast cancer: clinical features and patterns of recurrence. Clin Cancer Res 13: 4429–4434. [DOI] [PubMed] [Google Scholar]
- 5. Dewan MZ, Terunuma H, Takada M, Tanaka Y, Abe H, et al. (2007) Role of natural killer cells in hormone-independent rapid tumor formation and spontaneous metastasis of breast cancer cells in vivo. Breast Cancer Res Treat 104: 267–275. [DOI] [PubMed] [Google Scholar]
- 6. Sutlu T, Alici E (2009) Natural killer cell-based immunotherapy in cancer: current insights and future prospects. J Intern Med 266: 154–181. [DOI] [PubMed] [Google Scholar]
- 7. Castriconi R, Dondero A, Cilli M, Ognio E, Pezzolo A, et al. (2007) Human NK cell infusions prolong survival of metastatic human neuroblastoma-bearing NOD/scid mice. Cancer Immunol Immunother 56: 1733–1742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Geller MA, Cooley S, Judson PL, Ghebre R, Carson LF, et al. (2011) A phase II study of allogeneic natural killer cell therapy to treat patients with recurrent ovarian and breast cancer. Cytotherapy 13: 98–107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Kruschinski A, Moosmann A, Poschke I, Norell H, Chmielewski M, et al. (2008) Engineering antigen-specific primary human NK cells against HER-2 positive carcinomas. Proc Natl Acad Sci U S A 105: 17481–17486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Ames E, Hallett WH, Murphy WJ (2009) Sensitization of human breast cancer cells to natural killer cell-mediated cytotoxicity by proteasome inhibition. Clin Exp Immunol 155: 504–513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Ahn BC (2012) Sodium iodide symporter for nuclear molecular imaging and gene therapy: from bedside to bench and back. Theranostics 2: 392–402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Nowosielska EM, Cheda A, Wrembel-Wargocka J, Janiak MK (2011) Anti-neoplastic and immunostimulatory effects of low-dose X-ray fractions in mice. Int J Radiat Biol 87: 202–212. [DOI] [PubMed] [Google Scholar]
- 13. Ishikawa E, Tsuboi K, Saijo K, Takano S, Ohno T (2004) X-irradiation to human malignant glioma cells enhances the cytotoxicity of autologous killer lymphocytes under specific conditions. Int J Radiat Oncol Biol Phys 59: 1505–1512. [DOI] [PubMed] [Google Scholar]
- 14. Schmidt J, Patrut EM, Ma J, Jager D, Knaebel HP, et al. (2006) Immunomodulatory impact of interferon-alpha in combination with chemoradiation of pancreatic adenocarcinoma (CapRI). Cancer Immunol Immunother 55: 1396–1405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Jeon YH, Choi Y, Kim CW, Kim YH, Youn H, et al. (2010) Human sodium/iodide symporter-mediated radioiodine gene therapy enhances the killing activities of CTLs in a mouse tumor model. Mol Cancer Ther 9: 126–133. [DOI] [PubMed] [Google Scholar]
- 16. Hori T, Kondo T, Kanamori M, Tabuchi Y, Ogawa R, et al. (2010) Ionizing radiation enhances tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-induced apoptosis through up-regulations of death receptor 4 (DR4) and death receptor 5 (DR5) in human osteosarcoma cells. J Orthop Res 28: 739–745. [DOI] [PubMed] [Google Scholar]
- 17. Zhou Y, Song X, Jia R, Wang H, Dai L, et al. (2010) Radiation-inducible human tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) gene therapy: a novel treatment for radioresistant uveal melanoma. Pigment Cell Melanoma Res 23: 661–674. [DOI] [PubMed] [Google Scholar]
- 18. Chatterjee S, Malhotra R, Varghese F, Bukhari AB, Patil A, et al. (2013) Quantitative immunohistochemical analysis reveals association between sodium iodide symporter and estrogen receptor expression in breast cancer. PLoS One 8: e54055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Ryan J, Curran CE, Hennessy E, Newell J, Morris JC, et al. (2011) The sodium iodide symporter (NIS) and potential regulators in normal, benign and malignant human breast tissue. PLoS One 6: e16023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Upadhyay G, Singh R, Agarwal G, Mishra SK, Pal L, et al. (2003) Functional expression of sodium iodide symporter (NIS) in human breast cancer tissue. Breast Cancer Res Treat 77: 157–165. [DOI] [PubMed] [Google Scholar]
- 21. G EBCTC (2005) Effects of chemotherapy and hormonal therapy for early breast cancer on recurrence and 15-year survival: an overview of the randomised trials. Lancet 365: 1687–1717. [DOI] [PubMed] [Google Scholar]
- 22. Smyth MJ, Godfrey DI, Trapani JA (2001) A fresh look at tumor immunosurveillance and immunotherapy. Nat Immunol 2: 293–299. [DOI] [PubMed] [Google Scholar]
- 23. Rosenberg SA, Yang JC, Sherry RM, Hwu P, Topalian SL, et al. (2003) Inability to immunize patients with metastatic melanoma using plasmid DNA encoding the gp100 melanoma-melanocyte antigen. Hum Gene Ther 14: 709–714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Fogler WE, Volker K, McCormick KL, Watanabe M, Ortaldo JR, et al. (1996) NK cell infiltration into lung, liver, and subcutaneous B16 melanoma is mediated by VCAM-1/VLA-4 interaction. J Immunol 156: 4707–4714. [PubMed] [Google Scholar]
- 25. Gong JH, Maki G, Klingemann HG (1994) Characterization of a human cell line (NK-92) with phenotypical and functional characteristics of activated natural killer cells. Leukemia 8: 652–658. [PubMed] [Google Scholar]
- 26. Tonn T, Becker S, Esser R, Schwabe D, Seifried E (2001) Cellular immunotherapy of malignancies using the clonal natural killer cell line NK-92. J Hematother Stem Cell Res 10: 535–544. [DOI] [PubMed] [Google Scholar]
- 27. Tam YK, Maki G, Miyagawa B, Hennemann B, Tonn T, et al. (1999) Characterization of genetically altered, interleukin 2-independent natural killer cell lines suitable for adoptive cellular immunotherapy. Hum Gene Ther 10: 1359–1373. [DOI] [PubMed] [Google Scholar]
- 28. Nagashima S, Mailliard R, Kashii Y, Reichert TE, Herberman RB, et al. (1998) Stable transduction of the interleukin-2 gene into human natural killer cell lines and their phenotypic and functional characterization in vitro and in vivo. Blood 91: 3850–3861. [PubMed] [Google Scholar]
- 29. Moretta A, Bottino C, Vitale M, Pende D, Cantoni C, et al. (2001) Activating receptors and coreceptors involved in human natural killer cell-mediated cytolysis. Annu Rev Immunol 19: 197–223. [DOI] [PubMed] [Google Scholar]
- 30. Lanier LL (2008) Up on the tightrope: natural killer cell activation and inhibition. Nat Immunol 9: 495–502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Moretta L, Moretta A (2004) Killer immunoglobulin-like receptors. Curr Opin Immunol 16: 626–633. [DOI] [PubMed] [Google Scholar]
- 32. Zamai L, Ahmad M, Bennett IM, Azzoni L, Alnemri ES, et al. (1998) Natural killer (NK) cell-mediated cytotoxicity: differential use of TRAIL and Fas ligand by immature and mature primary human NK cells. J Exp Med 188: 2375–2380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Xu X, Rao GS, Groh V, Spies T, Gattuso P, et al. (2011) Major histocompatibility complex class I-related chain A/B (MICA/B) expression in tumor tissue and serum of pancreatic cancer: role of uric acid accumulation in gemcitabine-induced MICA/B expression. BMC Cancer 11: 194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Boelaert K, Franklyn JA (2003) Sodium iodide symporter: a novel strategy to target breast, prostate, and other cancers? Lancet 361: 796–797. [DOI] [PubMed] [Google Scholar]
- 35. Kogai T, Schultz JJ, Johnson LS, Huang M, Brent GA (2000) Retinoic acid induces sodium/iodide symporter gene expression and radioiodide uptake in the MCF-7 breast cancer cell line. Proc Natl Acad Sci U S A 97: 8519–8524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Dohan O, De la Vieja A, Carrasco N (2006) Hydrocortisone and purinergic signaling stimulate sodium/iodide symporter (NIS)-mediated iodide transport in breast cancer cells. Mol Endocrinol 20: 1121–1137. [DOI] [PubMed] [Google Scholar]
- 37. Fortunati N, Catalano MG, Marano F, Mugoni V, Pugliese M, et al. (2010) The pan-DAC inhibitor LBH589 is a multi-functional agent in breast cancer cells: cytotoxic drug and inducer of sodium-iodide symporter (NIS). Breast Cancer Res Treat 124: 667–675. [DOI] [PubMed] [Google Scholar]