

Abstract
Infections with antibiotic-resistant bacteria (ARB) in hospitalized patients are becoming increasingly frequent despite extensive infection-control efforts. Infections with ARB are most common in the intensive care units of tertiary-care hospitals, but the underlying cause of the increases may be a steady increase in the number of asymptomatic carriers entering hospitals. Carriers may shed ARB for years but remain undetected, transmitting ARB to others as they move among hospitals, long-term care facilities, and the community. We apply structured population models to explore the dynamics of ARB, addressing the following questions: (i) What is the relationship between the proportion of carriers admitted to a hospital, transmission, and the risk of infection with ARB? (ii) How do frequently hospitalized patients contribute to epidemics of ARB? (iii) How do transmission in the community, long-term care facilities, and hospitals interact to determine the proportion of the population that is carrying ARB? We offer an explanation for why ARB epidemics have fast and slow phases and why resistance may continue to increase despite infection-control efforts. To successfully manage ARB at tertiary-care hospitals, regional coordination of infection control may be necessary, including tracking asymptomatic carriers through health-care systems.
Nosocomial infections with antibiotic-resistant bacteria (ARB) occur with alarming frequency (1), and new multidrug-resistant bacteria continue to emerge, including vancomycin-resistant enterococci (VRE), methicillin-resistant Staphylococcus aureus (MRSA), two recent but isolated cases of vancomycin-resistant S. aureus (2, 3), and multiple-drug resistance in Gram-negative bacteria. In response, hospitals have used a variety of infection-control measures, some of which are costly and difficult to implement (4). Despite efforts to reduce transmission of ARB within hospitals, the incidence of VRE, MRSA, and other antibiotic-resistant infections continues to increase (1).
An important distinction in the epidemiology of ARB is made between infection and colonization. Infection is characterized by serious illness when ARB contaminate wounds, the bloodstream, or other tissues. In contrast, colonization with ARB may occur in the gut, nasal cavities, or other body surfaces. Colonizing bacteria may persist for years without causing disease or harming their hosts (5, 6); we call such individuals carriers. These carriers increase colonization pressure; the number of patients who are shedding increases the risk that another patient becomes a carrier for ARB or acquires a resistant infection (7). Hospitals that reduce the incidence of resistance (the number of new cases) may see no reduction in overall prevalence (the fraction of patients with ARB), because these hospitals admit an increasing number of ARB carriers (8, 9). Patients infected with ARB generally remain hospitalized until the symptoms are cured, but they may continue to carry and shed ARB for months or years.
We show that the prevalence of ARB in hospitals approaches equilibrium rapidly because of the rapid turnover of patients; the average length of stay (LOS) is ≈5 days (10, 11). Moreover, prevalence changes rapidly in response to changes in hospital infection control (11-17), so slow and steady increases in resistance must be due to something else, such as increases in the proportion of carriers admitted from the catchment population of a hospital, defined as the population from which patients are drawn, including long-term care facilities (LTCFs), other hospitals, and the community (5, 18). The health-care institutions that serve a common catchment population vary substantially in their relative size, transmission rates, and average LOS. How do increases in the number of ARB carriers in the catchment population contribute to increases in infections by ARB in the hospital, and what can be done about it?
Mathematical models play an important role in understanding the spread of ARB (19). We have built on existing theory for the transmission dynamics of ARB developed for simple, well-mixed populations (12, 20), but we are focused on phenomena that occur at a large spatial scale, ignoring competition between sensitive and resistant bacteria and the biological cost of resistance (21) and the relationship between antibiotic use and the prevalence of ARB (11, 22, 23). We have developed mathematical models with multiple institutions connected by patient movement; such models are called “structured” populations or “metapopulations.” Thus, we are developing epidemiological models (20, 21, 24) focused specifically on the consequences of persistent colonization and population structure, applying existing theory for structured populations (20, 24-27).
Structured Population Models
We have adapted simple mathematical models that have been used to model the population dynamics of ARB. Our focus is on the dynamic consequences of persistent colonization and the movement of patients to and from the community and among health-care institutions. We assume the entire catchment population is subdivided into subpopulations (e.g., hospitals) that play a role in the transmission of ARB. The population may also be heterogeneous with respect to epidemiologically relevant parameters, especially the likelihood of hospitalization. We either consider homogeneous populations or subdivide the population into distinct groups, such as the elderly and nonelderly. We assume that the subpopulations are well mixed with respect to the transmission of ARB. We restrict our focus to increased prevalence of colonization with ARB and tacitly assume that infection represents some fraction of total transmission within hospitals.
Many factors may account for the differences in transmission rates among subpopulations, including antibiotic use, infection control, the ratio of nurses to patients, and the size of the hospital. We are focused on understanding the consequences of different transmission rates and lengths of stay (LOS), regardless of the mechanism that accounts for higher transmission. Thus, we assume that transmission differs among subpopulations without explicitly describing why. The transmission rate in the jth subpopulation is denoted βj.
The average LOS, 1/σg,j, may differ among groups, g, and subpopulations, j. In contrast, we assume that persistence times for ARB are identically distributed for all subpopulations and groups; the average persistence time is denoted 1/λ. We let mg,j = σg,j/(λ + σg,j) denote the probability that a carrier of a given group, g, will leave the jth subpopulation before “clearing” the ARB they are carrying. On discharge, each individual is replaced; the proportion of individuals of group g admitted to subpopulation j that come from subpopulation k is αg,j,k.
Let xg,j be the fraction of subpopulation j of group g that is colonized. Similarly, let Xj denote the proportion of all individuals in the jth subpopulation that are colonized. The changes in the fraction colonized is given by the system of equations:
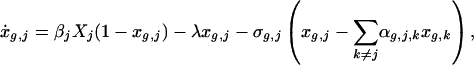 |
1 |
where k is summed across locations for each group, and the dot denotes a derivative with respect to time. We assume that each subpopulation has a constant size, and the proportion of each group is also constant (see Appendix). A diagram of the model is provided in Fig. 1.
Fig. 1.

A diagram of the general model. Individuals move among subpopulations, such as hospitals, LTCFs, and the community. The subpopulation is assumed to be well mixed with respect to the transmission of ARB. The population is also classified by group, based on some epidemiologically important difference. The size of the population at each location, Nj, and the proportion of each group, qg,j, are constant by assumption. The admission rate is equal to the discharge rate, σg,jqg, jNj. The portion of discharged individuals from subpopulation j that move to k is ωg,j,k. The portion of admitted individuals to subpopulation j that are from k is αg, j,k.
Threshold Criteria. A basis for threshold criteria in structured populations is given by the basic reproductive number, R0, defined as the number of new carriers generated by a typical carrier when resistance is otherwise absent. If each carrier generates more than one new carrier at low prevalence, R0 > 1, ARB spreads until an equilibrium is reached. In structured populations, carriers move among subpopulations, generating new cases for as long as an individual remains colonized. Because transmission rates and the LOS vary among subpopulations and groups, R0 is an average of the transmission rates in each location and the flow of carriers among institutions.
We define the “single-stay reproductive number” for each group in each subpopulation as Sg,j = βj/(λ + σg,j), the expected number of new cases generated by a carrier of a particular group g in subpopulation j during a single visit when resistance is rare. R0 in a structured population involves averaging of the number of cases per case in the next “generation,” for all groups and subpopulations, an expression involving Sg,j, mg,j, and the rates of movement among institutions (26).
Subpopulations may act as sources or sinks for ARB when the entire catchment population is considered, depending on whether they discharge more or fewer colonized patients than they admit. To identify sources and sinks, we define the “closed-population reproductive number,” Cj = βj/λ, as the reproductive number in subpopulation j when prevalence is low, if migration is ignored; i.e., we assume everybody stays in whatever population they start in. Because transmission rates do not vary by group, Cj is a property of the subpopulation. When resistance is rare, a location is a source if Cj > 1 and a sink otherwise.
ARB can persist in a catchment population if any subpopulation is a self-sustaining source; i.e., the single-stay reproductive number is >1, Sj > 1. If the LOS differs among groups, Sj for a subpopulation involves averaging (see Appendix). Self-sustaining sources with rapid turnover may also play the role of superspreaders (20, 24).
ARB may persist in a catchment population without a self-sustaining source, but with at least one dependent source, defined as a source with single-visit reproductive number <1. If all subpopulations are sinks, ARB cannot persist; whereas if all subpopulations are dependent sources, ARB will persist. If R0 > 1, prevalence will increase everywhere to a level where some subpopulations are converted from sources to sinks. If some subpopulations are sources and others are sinks, ARB may or may not persist, depending on the relative strength of these sources and sinks. Thus, locations that are sources (Cj > 1) should be focal points for control efforts, whether or not they are self-sustaining sources (Sj > 1).
Two-Population Model. An important example is the homogeneous two-subpopulation model; a single subscript is used to denote location (see Eq. 4). All individuals who leave one subpopulation simply move to the other; one may think of the two as a hospital and community. In this model, R0 is found by computing the dominant eigenvalue of a 2 × 2 “next-generation matrix” (26):
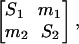 |
2 |
where Sj = βj/(λ+σj), mj = σj/(λ+σj). The basic reproductive number, R0, is the dominant eigenvalue of this matrix.
If Sj > 1 in either subpopulation, a single case will more than reproduce itself (at low prevalence) before migrating to another subpopulation. In this case, the subpopulation has a self-sustaining epidemic, and ARB will persist in both subpopulations.
An epidemic may occur, even if neither subpopulation is a self-sustaining source: i.e., R0 > 1 but Sj < 1 for both locations. Standard methods (28) can be used to express a threshold for invasion in a simpler way; persistence occurs if m1m2 > (1 - S1)(1 - S2) or equivalently m1m2 > (1 - C1(1 - m1))(1 - C2(1 - m2)). Thus, it is possible for the disease to be sustained by a combination of local transmission and migration. It is clear, however, that the disease can never spread if Cj < 1 for both subpopulations.
Equilibrium Prevalence. An important measure of risk is the equilibrium prevalence, the local, long-term average prevalence of ARB. Equilibrium prevalence in a subpopulation may be a complicated function. We illustrate some general principles in a homogeneous subpopulation model within a focal institution.
Let κ denote the proportion of hosts that is colonized on admission and m = σ/(λ + σ) the proportion that is discharged before ARB are cleared. The equilibrium prevalence in a subpopulation with immigration of colonized individuals is
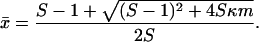 |
3 |
As long as κ > 0, it follows that x̄ > 0 and ARB are present in the subpopulation. Interestingly, the equilibrium depends only on two terms, the single-stay reproductive number S and the equilibrium proportion that would be maintained by immigration, clearance, and discharge in the absence of transmission in the subpopulation, mκ.
Fig. 2 shows the relationship between the single-stay reproductive number S and the proportion of individuals infected at equilibrium for a particular value of κm. Mathematically, we can distinguish three different regions on this graph. In the nonepidemic region (S « 1), the equilibrium prevalence is not limited by the number of susceptibles (i.e., noncarriers) in the subpopulation, but is determined mostly by the infectious potential of the immigrant cases; x̄ ≈ κm/(1 - S). In the epidemic region (S » 1), the disease will spread regardless of the immigration parameter, and equilibrium prevalence will be controlled by the depletion of noncarriers below the level needed for the equilibrium to continue spreading; x̄ ≈ 1 - (1 - κm)/S. Finally, there is an intermediate, quasiepidemic region (S ≈ 1), where immigration is important but the disease is magnified sufficiently that the depletion of susceptibles is also important; 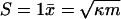 .
.
Fig. 2.

Colonization on admission interacts with local transmission dynamics to determine the equilibrium prevalence. We plotted the equilibrium prevalence of ARB (from Eq. 3) as a function of the single-stay reproductive number S when the proportion colonized without local transmission is κm = 0,0.04 (solid trace). We used three approximations to subdivide institutions into those that sustain internal epidemics (S » 1; dashed trace), for which prevalence is determined by patient-to-patient transmission; nonepidemics (S « 1; dotted trace), for which prevalence is determined by immigration of carriers; and quasiepidemics (S ≈ 1; dashed-dotted trace), for which prevalence is strongly influenced by both immigration and transmission. For example, for S = 1, prevalence is 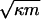 , either 0% or 20%.
, either 0% or 20%.
Fast and Slow Dynamics
In addition to threshold criteria and equilibrium prevalence, structured population models also describe the rate that prevalence changes in subpopulations when the LOS and transmission rates differ. To obtain parameter estimates for the relative size and average LOS in each subpopulation and group, we used U.S. health statistics. The distribution of persistence times for ARB has not been well characterized, even for clinically important species, such as VRE and MRSA. We used estimates for persistence that were consistent with available data. We have used a range of estimates for the single-stay reproductive numbers to illustrate the range of behavior possible from structured population models. Mathematical details for the examples are provided in Appendix.
Nosocomial and Community Transmission. The average LOS in a U.S. hospital is ≈5 days, and there are ≈1.6 occupied beds per 1,000 people (10). Put another way, the average LOS in the community is ≈620 times longer than in a hospital, ≈8.5 years.
The number of occupied hospital beds, the community size, and the single-stay reproductive numbers determine turnover and the rate of increase in prevalence. Fig. 3a shows a simulated epidemic for which neither the hospital nor the community is a self-sustaining source. The dynamics in the hospital and community are slow, reflecting the slow accumulation of carriers in both locations. The approach to equilibrium takes approximately three decades.
Fig. 3.

Structured population models may have fast and slow phases, depending on whether the hospital, the community, or neither is a self-sustaining source. (a) Carriers may accumulate slowly in the hospital population (solid trace) and community (dashed trace), even if neither one can sustain an internal epidemic. (b) When the hospital has a self-sustaining epidemic, the epidemic of ARB in the catchment population has fast and slow phases. Rapid early increases in prevalence are due to the epidemic in the hospital. Without colonization on admission, prevalence would rapidly reach a lower equilibrium (dashed trace and dashed-dotted trace). Slow increases after the initial epidemic reflect admission of carriers from the community. (c) The community may be a self-sustaining source, but prevalence increases slowly because resistance is initially rare and turnover is very slow. Prevalence remains higher in the hospital because daily transmission rates are higher. The role of community transmission may be underappreciated.
In contrast, when a hospital is a self-sustaining source, the fast turnover of hospitals implies that the dynamics will be much faster than the surrounding community. After an initial epidemic in the hospital, the slower approach to equilibrium reflects the readmission of carriers from the community who were recently hospitalized. The initial fast epidemic in the hospital provides the epidemic with a head start, and prevalence is near the equilibrium in about a decade for this set of parameters (Fig. 3b).
Despite the focus on hospitals, most people spend very little time there. Community transmission may play a very important role, even if it is very slow. We simulated another epidemic by using a set of parameters for which the community is a self-sustaining source (Fig. 3c). Because individuals stay longer in the community, it is possible for the community to have a higher single-stay reproductive number than the hospital (Sc > Sh), although the transmission rate and, thus, the closed-population reproductive number will generally be higher in the hospital (Cc < Ch). In this example, it is impossible to eliminate ARB by controlling incidence in the hospital alone. The importance of community transmission may not be appreciated, because prevalence is higher in the hospital and most infections (as opposed to colonization) happen there as well.
Frequently Hospitalized Patients. In the real world, risk is not distributed evenly across populations. We modified the above models to incorporate heterogeneity in hospitalization rates (Fig. 4). People >65 years of age account for ≈13% of the U.S. population, but nearly half of the total days of care (10). The average LOS for the elderly in hospitals is slightly longer than for the general population. Combining these statistics, we find that the average period between hospital visits for the elderly is ≈2.6 years, whereas the rest of the population is hospitalized about once every 15 years.
Fig. 4.

The elderly are frequently hospitalized, so they are more likely to be exposed and expose others. As a result of frequent hospitalization, ARB invade a catchment population more easily and prevalence is higher. One essential difference is that the period between hospital visits is shorter, so the likelihood of colonization on readmission is higher. The elderly population accounts for 12.6% of the population but about half of all days of care in the hospital. A greater portion of the elderly in each subpopulation are carriers (solid gray trace). Here, the daily transmission rates in the hospital and community are similar to those in Fig. 3a. The average prevalence in the hospital (solid black trace) and the community (dashed black trace) increases more rapidly and reaches a higher equilibrium than it would if the population were homogeneous.
The elderly illustrate how heterogeneity in the average LOS in various locations dramatically increases R0 for the entire population. Frequent hospitalization of the elderly implies that they are more frequently exposed, so they are more likely to be colonized. Moreover, because the average period between hospital visits is shorter, the elderly are more likely to remain colonized when they next visit a hospital. This set of simulations uses the same daily transmission rates as Fig. 3a, but the epidemic reaches a higher equilibrium, and the approach to equilibrium is faster.
LTCFs. LTCF residents are frequently hospitalized and many remain institutionalized after discharge from a hospital. Evidence suggests that prevalence of resistance in LTCFs is similar to the prevalence in surrounding hospitals (29). Patients in LTCFs receive a similar level and type of care compared with hospital patients, so daily transmission rates, thus Cj, in LTCFs is probably lower than but similar to hospitals. The slower turnover of patients in LTCFs thus implies that single-stay reproductive numbers may be much higher in LTCFs. It follows that LTCFs may play a very important role in spreading ARB.
By using parameters for average LOS and admissions from LTCFs, we simulated the spread of ARB in a system consisting of a LTCF, a hospital, and the community (Fig. 5). Although closed-population reproductive numbers for hospitals (Cj) may be similar to LTCFs, we have assumed the single-visit reproductive numbers, Sj, for the nonelderly in hospitals and LTCFs were identical. Thus, in these simulations, the closed population numbers are much lower in LTCFs compared with hospitals. For the hospital, the closed-population reproductive number is the same as Fig. 4.
Fig. 5.

LTCFs (dashed-dotted trace) may be the most important type of institution in health-care networks because LTCF patients are frequently hospitalized and receive a similar level and type of care as hospitalized patients. Single-stay reproductive numbers for the hospital and community are identical to those in Figs. 3a and 4. In this simulation, the single-stay reproductive numbers in the LTCF and hospital are identical, but the closed-population reproductive rate for the LTCF is much lower than the hospital because of the longer LOS. Compared with earlier simulations, prevalence increases faster and reaches a higher equilibrium in hospitals (solid trace) and the community (dashed trace).
Prevalence in LTCFs increases more slowly initially because of slower turnover. Later, prevalence at equilibrium is highest for LTCFs. Two contributing factors are that ≈95% of LTCF populations are elderly and 45% of LTCF admissions come from hospitals. Moreover, LTCFs further amplify resistance; compared with Fig. 4, prevalence is higher in hospitals and the community because of LTCFs. The relative importance of LTCFs versus hospitals as sources for resistance is an important issue in the epidemiology of ARB.
Discussion
When the average frequency of ARB in a population increases over time, resistance must be spreading somewhere faster than it is lost. An important component of the public health response to epidemics of ARB is to identify sources and sinks. Attention has tended to focus on the evolution of resistance in bacteria populations as a response to antibiotic use or nosocomial infections in hospital intensive care units, where the risk of infection is highest. But colonization through human-to-human transmission of ARB may be just as important to the increasing frequency of resistant infections. Each carrier in the unit increases the risk of infection for other patients (9); long-term increases in the risk of infection may reflect the increasing number colonized on admission. Because colonization is generally harmless, the importance of transmission and carriage among healthy people has been largely neglected.
Some information about what kinds of institutions are responsible for transmission may be found in examining the time course of an epidemic. Hospitals, LTCFs, and the community have different average LOS, which are reflected in the dynamics of spread. An epidemic triggered by high transmission in a hospital will generally cause a rapid increase in prevalence within that hospital, but this may be followed by slower increases in that hospital, other hospitals and institutions, and the community, as carriers accumulate in the catchment population. Prevalence of ARB may increase throughout the community for years or decades after a fast epidemic in a hospital with a self-sustaining epidemic. If the epidemic is triggered by changes in an LTCF, the initial epidemic will likely be slower and less likely to be accompanied by infection by ARB and, thus, less likely to be noticed. Epidemics triggered in the community will be still slower and less noticeable. Finally, if transmission increases gradually in several subpopulations, an epidemic of ARB may arise that cannot be attributed to any particular venue.
Fast and slow phases, such as we predict if both hospitals and the community are important, are consistent with reported epidemic patterns for ARB. For example, rapid increases in the prevalence of VRE in the late 1980s and early 1990s has been followed by slow and steady increases since then (1). MRSA, initially a problem in large research and teaching hospitals, spread later into smaller hospitals and the community. Now, MRSA is commonly acquired outside of hospitals. The changing epidemiology of MRSA may predict future changes in the epidemiology of VRE.
A critical assumption is that people who are colonized with ARB may continue to carry the bacteria for months or years. Not all individuals who are colonized with ARB are persistently colonized; common wisdom holds that humans may be persistent, intermittent, or noncarriers of various strains of bacteria (30). Some qualitative information about persistence times are available for MRSA (31-34), VRE (5, 35-37), Escherichia coli, (38), Pseudomonas aeruginosa (39), Klebsiella pneumoniae (40, 41), and other enterobacteria (42). These studies suggest that carriage for months or years is not uncommon. Unfortunately, these studies measured mean or median persistence, not the portion of the population that carry ARB for years. Qualitative descriptions of persistence are no substitute for quantitative understanding of persistence times and the factors that cause persistent colonization.
Heterogeneity in the LOS in hospitals or heterogeneity in the frequency of hospitalization increases R0, much like certain other kinds of heterogeneity (43). Important groups can be identified a priori as those who are frequently hospitalized or who have long hospital visits. For example, the elderly play a critical role in the development of resistance, because they spend more time in high-transmission subpopulations, so they are more likely to be colonized and more likely to expose others. Identifying other groups that play a focal role and targeting control efforts may increase the efficiency of control efforts. Important groups may include long-stay patients, cancer patients, organ-transplant patients, burn patients, the mentally ill, and chronic hemodialysis patients (10, 44-48). Persistent colonization of hospital workers may also be a key factor contributing to an epidemic; these workers are in close contact with patients and they are in and out of the hospital every day (12, 49).
Daily transmission rates may be highest in institutions, and the response to intervention may be most efficient there, but the role of community transmission may be more important than it appears (21). Most people spend most of their time in the community, and the fact that individuals circulate between the hospital and the community makes the community a vital link in the chain of ARB transmission. About 80% of all antibiotics used by humans are outside of hospitals (50). High carriage rates of VRE in the community were linked to the use of antibiotics in agriculture in some regions of Europe (51, 52), with long-term consequences for hospital prevalence under quasiepidemic transmission (53, 54). Thus, any factor that contributes to increased prevalence of resistance in a catchment population must be considered in management plans for ARB.
The accumulation of carriers in a catchment population links all of the institutions that serve it. To protect patients, individual hospitals may isolate recently admitted patients who are likely to be carriers. Control efforts can be coordinated regionally by tracking persistent carriers, with active surveillance focused on important groups, such as health care workers; frequent, recent, and long-term hospital patients; and the elderly and their family members. A regional perspective guided a VRE control program in the Siouxland region of Iowa, Nebraska, and South Dakota that is credited with reversing an epidemic of VRE (55-57). The low prevalence of MRSA in the Netherlands is due to an aggressive “search and destroy” policy (58) that includes isolating patients who were admitted from hospitals outside of the Netherlands and closing intensive care units when MRSA transmission was suspected. The Dutch policy was expensive, but a recent analysis suggests it was also cost-effective (59). Similar policies may not be cost-effective for the less virulent VRE (60).
Hospital infection control is expensive, and the costs are shouldered entirely by hospitals. Hospitals may reduce the prevalence of ARB rapidly through improved hospital infection control to a certain point, but larger, long-term reductions in prevalence may require a reduction in the carriage rate in the catchment population. Such changes may require decades (11, 21). One reason for the success of the Dutch policy was that all of the hospitals in a region adopted the policy; most of the costs were generated when hospitals admitted carriers who had acquired MRSA elsewhere (59). The benefits of effective infection control and the costs of poor infection control extend beyond a single hospital. A successful public health response to the antibiotic resistance epidemic must recognize that colonization with ARB is a sort of pollution. Policies to control resistance should be coordinated regionally.
Acknowledgments
We thank Bruce Levin and an anonymous reviewer for their suggestions and Rebecca Freeman Grais and Ellis McKenzie for their advice and comments. This research was supported by grants from the Department of Veterans Affairs and the Fogarty International Center.
Appendix
Let Nj denote the total size of the subpopulation at the jth location, and let qg,j denote the fraction of the subpopulation at location j that is of group g. We assume Nj and qg,j are constant. The single-stay reproductive number for subpopulation j is Sj = Σgqg,jSg,j. The prevalence in subpopulation j is Xj = Σgqg,jxg,j. Given that αg,j,k represents the proportion of group g admitted to j from k, the sum over all sources is constrained such that Σkαg,j,k = 1. Let ωg,j,k be the proportion of the group g leaving subpopulation j that is admitted to location k; the sum over all destinations is constrained such that Σkωg,j,k = 1. Because subpopulation sizes are constant, admissions and discharges must be balanced: αg,j,kσg,jqg,jNj = ωg,j,kσg,kqg,kNk.
Nosocomial vs. Community Transmission. A simple model for nosocomial and community transmission is a special case of the general model with only two equations:
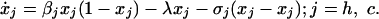 |
4 |
To compute the period between hospital visits, we note that σhNh = σcNc, 1/σh ≈ 5 days and Nc ≈ 620Nh.
The Elderly. A simple model that includes the elderly incorporates two institutions and two groups. Subscripts h and c denote the hospital and community; subscripts e and y denote elderly and nonelderly, respectively. We let qh,e denote the fraction of the hospital population that is composed of the elderly and qe,c the fraction of elderly in the community, and XH = qh,exh,e + (1 - qh,e)xh,y and XC = qc,e xc,e + (1 - qc,e)xc,y. The dynamics for the elderly are described by two equations: in hospitals ẋe,h = βHXH(1 - xe,h) - λxe,h - σe,h(xe,h - xe,c) and in the community ẋe,c = βCXC(1 - xe,c) - λxe,c - σe,c(xe,c - xe,h). The dynamics for the nonelderly are also described by two equations: in hospitals ẋy,h = βHXH(1 - xy,h) - λxy,h - σy,h(xy,h - xy,c), and in the community ẋy,c = βCXC(1 - xy,c) - λxy,c - σy,c(xy,c - xy,h). The average LOS for the elderly is ≈6 days compared with 4.9 days for the rest of the population. The total U.S. population is ≈281 million people, of which ≈.44 million are hospitalized. Because the elderly account for 12.6% of the total population (35.4 million) and 50% of all days of care, we get that Nr = Nh = 0.22 million, Ns = 35.2 million, and Nc = 245.4 million.
LTCFs. Models for LTCFs combine interinstitutional transfer and frequent hospitalization. This model extends the one for the elderly, adding one more location and the subscript f for the population in LTCFs. We assume the LTCF is composed entirely of the elderly. The dynamics are described by five equations. The notation and equations describing the nonelderly are taken from the previous model. Three equations now describe the elderly: in hospitals ẋe,h = βHXH(1 - xe,h) - λxe,h - σe,h(xe,h - αe,h,fxe,f - αe,h,cxe,c); in LTCFs ẋf,h = βfxf(1 - xe,f) - λxe,f - σe,f(xe,f - αe,j,hxe,h - αe,f,cxe,c); and in the community ẋe,c = βCXC(1 - xe,c) - λxe,c - σe,c(xe,c - αe,c,h xe,h - αe,c,fxe,f). By using statistics for the U.S. population, Nr = 0.22 million, Nf = 1.5 million, and Ns = 33.7 million. The average LOS for hospitals and LTCFs are 1/σr = 6 days, 1/σf = 60 days; ≈45% of nursing-home patients are admitted from the hospital (αe,f,r = 0.45), and are approximately equally likely to be discharged to the community or a hospital (ωr,f = 0.5). Solving for other parameters, we get 1/σs = 0.001, αe,s,r = 0.68, and αe,r,f = 0.33. For the elderly, average waiting time to a hospital visit is ≈4 years, and the average waiting time to a nursing home is ≈5.2 years.
Abbreviations: ARB, antibiotic-resistant bacteria; VRE, vancomycin-resistant enterococci; MRSA, methicillin-resistant Staphylococcus aureus; LOS, length of stay; LTCF, long-term care facility.
References
- 1.National Nosocomial Infections Surveillance (NNIS) System Report, Data Summary from January 1992-June 2001, issued August 2001 (2001) Am. J. Infect. Control 29, 404-421. [DOI] [PubMed] [Google Scholar]
- 2.Centers Disease Control and Prevention (2002) Morbid. Mortal. Wkly. Rep. 51, 565-567. [Google Scholar]
- 3.Centers Disease Control and Prevention (2002) Morbid. Mortal. Wkly. Rep. 51, 509. [Google Scholar]
- 4.Weinstein, R. A. (2001) Emerg. Infect. Dis. 7, 188-192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bonten, M. J., Hayden, M. K., Nathan, C., Rice, T. W. & Weinstein, R. A. (1998) J. Infect. Dis. 177, 378-382. [DOI] [PubMed] [Google Scholar]
- 6.Sanford, M. D., Widmer, A. F., Bale, M. J., Jones, R. N. & Wenzel, R. P. (1994) Clin. Infect. Dis. 19, 1123-1128. [DOI] [PubMed] [Google Scholar]
- 7.Bonten, M. J., Slaughter, S., Ambergen, A. W., Hayden, M. K., van Voorhis, J., Nathan, C. & Weinstein, R. A. (1998) Arch Intern. Med. (Moscow) 158, 1127-1132. [DOI] [PubMed] [Google Scholar]
- 8.Pelupessy, I., Bonten, M. J. & Diekmann, O. (2002) Proc. Natl. Acad. Sci. USA 99, 5601-5605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Bonten, M. J., Slaughter, S., Hayden, M. K., Nathan, C., van Voorhis, J. & Weinstein, R. A. (1998) Crit. Care Med. 26, 2001-2004. [DOI] [PubMed] [Google Scholar]
- 10.Hall, M. J & Owings, M. F. (2002) 2000 National Hospital Discharge Survey (Natl. Center for Health Statistics, Centers for Disease Control and Prevention, Atlanta), Adv. Data Vital Health Stat. No. 329.
- 11.Lipsitch, M., Bergstrom, C. T. & Levin, B. R. (2000) Proc. Natl. Acad. Sci. USA 97, 1938-1943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Austin, D. J., Bonten, M. J. M., Weinstein, R. A., Slaughter, S. & Anderson, R. M. (1999) Proc. Natl. Acad. Sci. USA 96, 6908-6913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Cosseron-Zerbib, M., Roque Afonso, A., Naas, T., Durand, P., Meyer, L., Costa, Y., el Helali, N., Huault, G. & Nordmann, P. (1998) J. Hosp. Infect. 40, 225-235. [DOI] [PubMed] [Google Scholar]
- 14.Boyce, J. M., Landry, M., Deetz, T. R. & DuPont, H. L. (1981) Infect. Control 2, 110-116. [DOI] [PubMed] [Google Scholar]
- 15.Dunkle, L. M., Naqvi, S. H., McCallum, R. & Lofgren, J. P. (1981) Am. J. Med. 70, 455-458. [DOI] [PubMed] [Google Scholar]
- 16.Lilly, H. A & Lowbury, E. J. (1978) J. Antimicrob. Chemother. 4, 545-550. [DOI] [PubMed] [Google Scholar]
- 17.Franco, J. A., Eitzman, D. V. & Baer, H. (1973) Am. J. Dis. Child. 126, 318-321. [DOI] [PubMed] [Google Scholar]
- 18.Bonilla, H. F., Zervos, M. A., Lyons, M. J., Bradley, S. F., Hedderwick, S. A., Ramsey, M. A., Paul, L. K. & Kauffman, C. A. (1997) Infect. Control Hosp. Epidemiol. 18, 333-339. [DOI] [PubMed] [Google Scholar]
- 19.Bonten, M. J., Austin, D. J. & Lipsitch, M. (2001) Clin. Infect. Dis. 33, 1739-1746. [DOI] [PubMed] [Google Scholar]
- 20.Austin, D. J & Anderson, R. M. (1999) J. Infect. Dis. 179, 883-891. [DOI] [PubMed] [Google Scholar]
- 21.Levin, B. R. (2001) Clin. Infect. Dis. 33, S161-S169. [DOI] [PubMed] [Google Scholar]
- 22.Austin, D. J., Kakehashi, M. & Anderson, R. M. (1997) Proc. R. Soc. Lond. B 264, 1629-1638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Austin, D. J., Kristinsson, K. G. & Anderson, R. M. (1999) Proc. Natl. Acad. Sci. USA 96, 1152-1156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Austin, D. J & Anderson, R. M. (1999) Philos. Trans. R. Soc. Lond. B 354, 721-738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Sattenspiel, L. & Simon, C. P. (1988) Math Biosci. 90, 341-366. [Google Scholar]
- 26.Diekmann, O., Heesterbeek, J. A. P. & Metz, J. A. J. (1990) J. Math Biol. 28, 365-382. [DOI] [PubMed] [Google Scholar]
- 27.Sattenspiel, L. & Dietz, K. (1995) Math Biosci. 128, 71-91. [DOI] [PubMed] [Google Scholar]
- 28.Jury, E. I. (1971) IEEE Trans Autom. Control AC-16, 233-240. [Google Scholar]
- 29.Elizaga, M. L., Weinstein, R. A. & Hayden, M. K. (2002) Clin. Infect. Dis. 34, 441-446. [DOI] [PubMed] [Google Scholar]
- 30.Kluytmans, J., van Belkam, A. & Verbrugh, H. (1997) Clin. Microbiol. Rev. 10, 505-520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Scanvic, A., Denic, L., Gaillon, S., Giry, P., Andremont, A. & Lucet, J. C. (2001) Clin. Infect. Dis. 32, 1393-1398. [DOI] [PubMed] [Google Scholar]
- 32.Harbarth, S., Liassine, N., Dharan, S., Herrault, P., Auckenthaler, R. & Pittet, D. (2000) Clin. Infect. Dis. 31, 1380-1385. [DOI] [PubMed] [Google Scholar]
- 33.Perl, T. M., Cullen, J. J., Wenzel, R. P., Zimmerman, M. B., Pfaller, M. A., Sheppard, D., Twombley, J., French, P. P., Herwaldt, L. A., et. al. (2002) N. Engl. J. Med. 346, 1871-1877. [DOI] [PubMed] [Google Scholar]
- 34.Lee, Y. L., Cesario, T., Gupta, G., Flionis, L., Tran, C., Decker, M. & Thrupp, L. (1997) Am. J. Infect. Control 25, 312-321. [DOI] [PubMed] [Google Scholar]
- 35.Henning, K. J., Delencastre, H., Eagan, J., Boone, N., Brown, A., Chung, M., Wollner, N. & Armstrong, D. (1996) Pediatr. Infect. Dis. J. 15, 848-854. [DOI] [PubMed] [Google Scholar]
- 36.Baden, L. R., Thiemke, W., Skolnik, A., Chambers, R., Strymish, J., Gold, H. S., Moellering, R. C. J. & Eliopoulos, G. M. (2001) Clin. Infect. Dis. 33, 1654-1660. [DOI] [PubMed] [Google Scholar]
- 37.Wong, M. T., Kauffman, C. A., Standiford, H. C., Linden, P., Fort, G., Fuchs, H. J., Porter, S. B. & Wenzel, R. P. (2001) Clin. Infect. Dis. 33, 1476-1482. [DOI] [PubMed] [Google Scholar]
- 38.Mereghetti, L., Tayoro, J., Watt, S., Lanotte, P., Loulergue, J., Perrotin, D. & Quentin, R. (2002) J. Hosp. Infect. 52, 43-51. [DOI] [PubMed] [Google Scholar]
- 39.Blanc, D. S., Petignat, C., Janin, B., Bille, J. & Francioli, P. (1998) Clin. Microbiol. Infect. 4, 242-247. [DOI] [PubMed] [Google Scholar]
- 40.Bonten, M. J., Bergmans, D. C., Speijer, H. & Stobberingh, E. E. (1999) Am. J. Respir. Crit. Care Med. 160, 1212-1219. [DOI] [PubMed] [Google Scholar]
- 41.Maroncle, N., Balestrino, D., Rich, C. & Forestier, C. (2002) Infect. Immun. 70, 4729-4734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.De Champs, C., Sauvant, M. P., Chanal, C., Sirot, D., Gazuy, N., Malhuret, R., Baguet, J. C. & Sirot, J. (1989) J. Clin. Microbiol. 27, 2887-2890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Dushoff, J. & Levin, S. (1995) Math Biosci. 128, 25-40. [DOI] [PubMed] [Google Scholar]
- 44.Hall, M. J. & Kozak, L. H. (1993) Long-Stay Patients in Short-Stay Hospitals (Natl. Center for Health Statistics, Centers for Disease Control and Prevention, Atlanta), Adv. Data Vital Health Stat. No. 229.
- 45.Chadwick, P. R., Oppenheim, B. A., Fox, A., Woodford, N., Morgenstern, G. R. & Scarffe, J. H. (1996) J. Hosp. Infect. 34, 171-182. [DOI] [PubMed] [Google Scholar]
- 46.Patel, R., Allen, S. L., Manahan, J. M., Wright, A. J., Krom, R. A., Wiesner, R. H., Persing, D. H., Cockerill, F. R. & Thompson, R. L. (2001) Liver Transpl. 7, 27-31. [DOI] [PubMed] [Google Scholar]
- 47.Phillips, L. G., Heggers, J. P. & Robson, M. (1992) J. Burn Care Rehabil. 13, 293-297. [DOI] [PubMed] [Google Scholar]
- 48.Tokars, J. I., Gehr, T., Jarvis, W. R., Anderson, J., Armistead, N., Miller, E. R., Parrish, J., Qaiyumi, S., Arduino, M., Holt, S. C., Tenover, F. C., Westbrook, G. & Light, P. (2001) Kidney Int. 60, 1511-1516. [DOI] [PubMed] [Google Scholar]
- 49.D'Agata, E. M., Gautam, S., Green, W. K. & Tang, Y. W. (2002) Clin. Infect. Dis. 34, 167-172. [DOI] [PubMed] [Google Scholar]
- 50.Hoiby, N. (2000) J. Antimicrob. Chemother. 46, 59-62. [PubMed] [Google Scholar]
- 51.Wegener, H. C., Aarestrup, F. M., Jensen, L. B., Mammerum, A. M. & Bager, F. (1999) Emerg. Infect. Dis. 5, 329-335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Bonten, M. J., Willems, R. & Weinstein, R. A. (2001) Lancet 1, 314-325. [DOI] [PubMed] [Google Scholar]
- 53.Smith, D. L., Harris, A. D., Johnson, J. A., Silbergeld, E. K. & Morris, J. G. (2002) Proc. Natl. Acad. Sci. USA 99, 6434-6439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Smith, D. L., Johnson, J. A., Harris, A. D., Furuno, J. P., Perencevich, E. N. & Morris, J. G., Jr. (2003) Lancet Infect. Dis. 3, 241-249. [DOI] [PubMed] [Google Scholar]
- 55.Ostrowsky, B. E., Trick, W. E., Sohn, A. H., Quirk, S. B., Holt, S., Carson, L. A., Hill, B. C., Arduino, M. J., Kuehnert, M. J. & Jarvis, W. R. (2001) N. Engl. J. Med. 344, 1427-1433. [DOI] [PubMed] [Google Scholar]
- 56.Sohn, A. H., Ostrowsky, B. E., Sinkowitz-Cochran, R. L., Quirk, S. B. & Jarvis, W. R. (2001) Am. J. Infect. Control 29, 53-57. [DOI] [PubMed] [Google Scholar]
- 57.Trick, W. E., Kuehnert, M. J., Quirk, S. B., Arduino, M. J., Aguero, S. M., Carson, L. A., Hill, B. C., Banerjee, S. N. & Jarvis, W. R. (1999) J. Infect. Dis. 180, 391-396. [DOI] [PubMed] [Google Scholar]
- 58.Verhoef, J., Beaujean, D., Blok, H., Baars, A., Meyler, A., van der Werken, C. & Weersink, A. (1999) Eur. J. Clin. Microbiol. Infect. Dis. 18, 461-466. [DOI] [PubMed] [Google Scholar]
- 59.Vriens, M., Blok, H., Fluit, A., Troelstra, A., van der Werken, C. & Verhoef, J. (2002) Eur. J. Clin. Microbiol. Infect. Dis. 21, 782-786. [DOI] [PubMed] [Google Scholar]
- 60.Ridwan, B., Mascini, E., van Der Reijden, N., Verhoef, J. & Bonten, M. (2002) BMJ 324, 666-668. [DOI] [PMC free article] [PubMed] [Google Scholar]