

Abstract
The purpose of this study was to evaluate whether Branched-chain amino acids (BCAAs) supplementation had any beneficial effects on growth and metabolic parameters of young rats submitted to chronic aerobic exercise. Thirty-two young rats (age: 21-d) were randomly assigned to four experimental groups (n = 8): Supplemented Trained (Sup/Ex), Control Trained (Ctrl/Ex), Supplemented Sedentary (Sup/Sed) and Control Sedentary (Ctrl/Sed). The trained groups underwent a five-week swimming protocol and received supplemented (45 mg BCAA/body weight/day) or control ration. Trained animals presented a lower body length and a higher cartilage weight, regardless of supplementation. Physical activity was responsible for a substantial reduction in proteoglycan synthesis in cartilage tissue, and BCAA supplementation was able to attenuate this reduction and also to improve glycogen stores in the liver, although no major differences were found in body growth associated to this supplementation.
Key points.
Cartilage proteoglycan synthesis was dramatically reduced in trained animals as a whole.
BCAA supplementation augmented liver glycogen stores and reduced proteolysis in our experimental conditions
Trained animals receiving BCAA supplementation featured increased proteoglycan synthesis compared to sedentary ones, probably because BCAA may have attenuated the negative effects of exercise on cartilage development.
BCAA supplementation was not capable of neutralizing directly the negative effects of long-term physical training and lower food intake in young male rats on body growth
Key words: Branched-chain amino acids, exercise, growth, metabolism, leucine, rat
Introduction
Many people from all age groups are currently involved in regular physical activity. Among them, children and adolescents deserve special attention (Ribeiro et al., 2010). Although moderate physical exercise has beneficial effect on growth and development, excessive exercise training could lead to opposite results (Caine et al., 2001; Georgepoulos et al., 2010).
Body growth processes (including bone, cartilage and muscle), demand a high amount of energy and nutrients. As such, nutritional deficits are expected to jeopardize growth as a whole. For instance, Yahya and Millward (1994) showed that short term protein deficits were capable of disbalancing the well-coordinated bone and muscle growth.
It is important to note that epiphysis cartilage development and growth consists of proteoglycan synthesis as well as cell multiplication (chondrogenesis). As such, proteoglycan synthesis is a measurement of cartilage and long bone growth. Indeed, Ribeiro et al., 2010 considered proteoglycan synthesis a biological marker of nutritional status.
Thinking about situations where the body faces energy/nutrient deficits, we can mention physical exercise. Exercise, single bout or regular training, characterizes a state of increased glucose uptake by skeletal muscle during (as an energy substrate) and after (for glycogen and protein synthesis) physical effort (Blomstrand and Saltin, 1999; Hargreaves, 1998; Tipton and Wolfe, 2001).
Considering the need for energy in protein synthesis, and consequently for body growth, there is a strict relationship between muscle protein synthesis and glycogen stores once glycogen is an available source of energy to the muscle (Newsholme and Leech, 1988). Muscle growth will occur adequately only if bone growth is guaranteed by an adequate nutrient supply (Yahya and Millward, 1994).
Branched-chain amino acids (BCAAs) are sources of nitrogen for the synthesis of non-essential amino acids such as glutamine and alanine (Tom and Nair, 2006). Leucine is a potent inhibitor of muscle protein degradation, enhances insulin secretion (Nicastro et al., 2011) and may also be an important precursor for gluconeogenesis in both liver and muscle (Araujo et al., 2006; Campos-Ferraz, data nonpublished).
To date, no studies have been conducted relating BCAAs supplementation (isoleucine, leucine and valine) on bone and cartilage development in growing trained animals. The few studies that used BCAA supplementation on cartilage development did not evaluate BCAA effects on training individuals. In one study with sedentary pigs, BCAA supplementation did not reduce severity of osteochondrosis lesions compared to the control group (Frantz et al., 2008).
With this in mind, we used an experimental model to reproduce the situation of young individuals submitted to a regular training in order to identify the effects of this practice in biomarkers of muscle and bone growth. Therefore, the aim of the present study was to evaluate if whether BCAA supplementation could feature any benefit regarding muscle and bone growth in growing rats submitted to aerobic exercise.
Methods
Animals
Male Wistar young rats (21 days old) were kept in individual cages on a 12 h dark/light cycle for 6 weeks. The animals were distributed into four experimental groups (n=8 each): supplemented trained (group Sup/Ex), control diet and trained (group Ctrl/Ex), supplemented diet and sedentary (group Sup/Sed) and control diet and sedentary (group Ctrl/Sed). The local Ethics Committee approved all experimental procedures, and the guidelines for animal care and use were followed (National Research Council, 1996). Both food intake and animal weight were measured three times a week. Protein ingestion was calculated based on total food intake (g) and protein content of the diet.
Diets
The control diet consisted of a standard ration designed for growing rodents by Reeves et al., 1993 and was prepared in our laboratory. It provided adequate levels of BCAA derived from casein, vitamins, minerals, fiber, carbohydrates and lipids. The protein content was assessed at 17% using the method provided by AOAC (1980). The supplemented diet was prepared using the same type of food as described above with the addition of BCAA (powdered, provided by Aji-No-Moto, Brazil) in order to reach 1.5 times the recommended level for each of the three amino acids (isoleucine, leucine and valine, 45 mg BCAA/kg body weight/day). The protein content of the supplemented diet was analyzed and resulted in 17.4% from the total energy content of the diet.
Physical activity
The swimming training protocol was that described by LANCHA Jr. et al. (1995), in which 10 individual PVC pools were connected to a boiler to allow the circulation of water and to keep the water temperature constant at 32?C. The first week of exercise consisted of an adaptation period during which the animals swam for 20 minutes. After this period, the animals started to swim longer (30 min - week 2, 40 min - week 3, 50 min - week 4 and 60 min - week 5, respectively) with a 5% body weight load attached to the tail, five times a week, always between 8 and 11 am. On the last week, if the rats were not able to stand the 5% body weight load, it would be reduced to 4%, to assure they completed 60 min of exercise.
Euthanasia
Throughout the study, care was taken to avoid animals’ unnecessary suffering. After the last training session, which was performed on the day before euthanasia, animals were fasted overnight. The animals were euthanized by decapitation. Blood was collected and handled in heparin-stabilized tubes, centrifuged at 2500g for 10 min at 4?C, and plasma was stored at -80°C. Soleus and gastrocnemius muscles were weighed, immersed in liquid nitrogen and stored at -80°C for further analysis.
Biometric measurements
A few minutes before euthanasia, body weight and body length were measured using a digital scale and a graded wooden device, respectively (Hughes and Tanner, 1970). After euthanasia, muscle, liver, tibia bone, tibia cartilage and adipose tissue were collected and weighed.
Biochemical analysis
Glucose determination: Plasma glucose was determined according to Trinder, 1969 using colorimetric-enzymatic reactions. Basically, glucose is oxidized to gluconic acid and H2O2 by glucose-oxidase. H2O2, in the presence of peroxidase, catalizes phenol oxidation with 4-aminophenazon, giving a red color to the solution, whose peak absorbance is read at 505 nm.
Plasma aminogram: Plasma samples were deproteinized with 6% sulfosalicylic acid (1:5) and centrifuged at 9,000 g for 2 minutes, and the supernatant was diluted in NaS buffer, separated by ultra filtration and stored at -80°C. The concentrations of essential amino acids and ammonia were determined by reverse-phase HPLC (Busse and Carpenter, 1976).
Muscle protein and total RNA: Muscle protein content was determined according to Lowry et al., 1951. Protein was pretreated with copper (II) in a modified biuret reagent (alkaline copper solution stabilized with sodium potassium tartrate).The addition of Folin & Ciocalteu’s phenol reagent forms chromogens that give increasing absorbance between 550 nm and 750 nm. Normally, absorbance at the peak (750 nm) or shoulder (660 nm) is used to quantify protein concentrations between 1-100 μg/ml. In our case, bovine albumin was used to build the standard curve and read the samples at 660nm.
Total RNA concentration was determined in muscle samples as follows: protein in the homogenate was precipitated by the addition of an equal volume of 10% trichloroacetic acid (TCA) followed by two washings of the precipitate with 5% TCA to remove acid-soluble compounds. Lipids were removed by two washings with 95% ethanol: ether (3:1 v/v). The precipitate was suspended in 1.5 ml 0.5 N perchloric acid (PCA) and heated to 70C°for 20 minutes to hydrolyze DNA. The hydrolysis step was repeated with another 1.5 ml PCA. The samples were compared against a 2% perchloric acid solution as standard. For the calculation of RNA, each extinction degree at 260 nm was considered to be equal to a 32 μg RNA/ml solution (Munro and Fleck, 1966).
Plasma free fatty acid (FFA): FFA levels were determined using a modified method previously described by Regow et al. (1971). Plasma was treated with an organic solvent and CuNO2. Next, sodium diethyl-dithiocarbamate was used to produce a yellow color solution whose intensity was proportional to the FFA levels of the sample. The samples were read in a spectrophotometer at 435 nm.
Glycogen: Muscle and liver glycogen concentrations were determined according to Hassid and Abrahams (1957). Briefly, tissues were treated with 6 N KOH for 30 minutes in a 37°C water bath, and 70% ethanol was added to precipitate glycogen. Next, the solution was centrifuged for 20 minutes at 3,000 rpm and the pellet was diluted in hot deionized water. Anthrone diluted in sulfuric acid was then added to turn the solution green, and the reaction was analyzed with a spectrophotometer at 650 nm.
Proteoglycan synthesis: The proteoglycan synthesis was measured in the tibia epiphysis as the incorporation of 35S-sodium sulfate into acid-precipitable proteoglycan. The animals were injected i.p. with the 35SO4 in 0.9% (w/v) NaCl (0.5 mL/100 g, 40μCi/100 g) and were euthanatized 1 h after the injection. The epiphysis (including the growth plate) was cleaned of all visible traces of muscle and tendons, pulverized and homogenized in ice-cold 10% trichloroacetic acid (TCA), washed twice with 10% TCA and with perchloric acid to remove any residual free 35SO4 and solubilized in 2 ml 90% (w/v) formic acid, after incubation at 90°C overnight. 35S radioactivity was measured in a Beckman LS-150 liquid scintillation counter and expressed as d.p. m/mg tissue (Yahya et al., 1994). Proteoglycan synthesis was measured 23h after the last training session in order to analyze the differences among the different protocols of the study.
Statistical analysis
The results are reported as mean ± standard deviation and were analyzed with the software SPSS 15.0 for Windows. A two-way analysis of variance (ANOVA) was used among all experimental groups, to determine any effect of supplementation and exercise on experimental parameters. If the two-way ANOVA was significant (p ≤ 0.05), individual differences between trained and untrained and between supplementation type were tested with a post-hoc test (Scheffé).
Results
To assess nutritional status, body weight variation throughout the experiment was measured. Table 1 shows the variation of body weight of the animals. Sedentary rats, regardless of their diet intake, were heavier than trained animals. This table also shows total food intake. A significant difference was observed between trained and sedentary rats, as the latter showed a higher food intake regardless of the diet. Absolute gastrocnemius weight, relative gastrocnemius (% of body weight) muscle weight and muscle/tibia ratio (according to Tirapegui et al. , 1994) are shown. Sedentary rats featured greater absolute gastrocnemius muscle weight compared to trained rats. Nevertheless, no significant differences were found in relative gastrocnemius weight and muscle/tibia ratio. Epydidimal adipose tissue relative weight was significantly higher in Ctrl/Sed animals, when compared to the Ctrl/Ex group.
Table 1.
Parameters related to food ingestion and body weight variation of the experimental animals. Values expressed as mean (± standard deviation).
| Groups | Δ body weight | Gastrocnemius weight (%) 1 |
Muscle/Tibia (mg/mm 3,85) |
Food Intake (g) |
Epydidimal Adipose Tissue weight (%) 1 |
|---|---|---|---|---|---|
| Sup/Ex | 4.0 (.5) | .55 (.07) | 1.96 (.50) | 600 (54) | 1.05 (.38) |
| Ctrl/Ex | 3.9 (.6) | .62 (.02) | 2.00 (.40) | 615 (98) | 1.26 (.20) |
| Sup/Sed | 5.2 (.3) † | .59 (.04) | 2.16 (.60) | 689 (29) † | 1.63 (.46) † |
| Ctrl/Sed | 5.3 (.8) * | .59 (.04) | 1.95 (.70) | 740 (62) ‡ | 1.58 (.49) ‡ |
Δ body weight: final weight-initial weight/initial weight
1 tissue weight in relation to total body weight
* p < 0.05 when compared to Ctrl/Sed
† p < 0.05 when compared to Sup/Ex
‡ p < 0.05 when compared to Ctrl/Ex.
Body growth was assessed by skull-tale length. Table 2 shows the skull-tale and tibia length; the first was significantly higher in sedentary rats; tibial length fetured no significant differences among the experimental groups. Muscle weight was examined between the different groups; however, absolute weight was higher in sedentary animals, irrespective of the diet. Epiphysis cartilage weight and sulfate incorporation are also present in this table. In trained rats, cartilage weight is increased compared to sedentary animals, however, proteoglycan synthesis is remarkably lower in trained compared to sedentary rats, albeit the Sup/Ex group featured increased sulfate incorporation compared to the Ctrl/Ex group.
Table 2.
Parameters of bone growth. Values expressed as mean (±standard deviation).
| Groups | Skull-tale length (cm) |
Tibia length (mm) |
Cartilage weight (% of body weight) |
Proteoglycan synthesis1 (cpm/100 mg tissue). |
|---|---|---|---|---|
| Sup/Ex | 36.07 (1.20) | 32.82 (1.68) | .039 (.001) § | 301.95 (81.50) ‡ |
| Ctrl/Ex | 35.92 (2.33) | 33.33 (2.39) | .041 (.003) †* | 218.52 (80.40) |
| Sup/Sed | 38.28 (.80) † | 34.12 (2.19) | .034 (.003) | 6395.95 (2066.20) † |
| Ctrl/Sed | 37.07 (1.30) ‡ | 35.03 (2.67) | .034 (.003) | 6284.27 (1877.30) ‡ |
1 by 35SO4 incorporation on cartilage
* p < 0.05 when compared to Ctrl/Sed
† p<0.05 when compared to Sup/Ex
‡ p<0.05 when compared to Ctrl/Ex
§ p<0.05 when compared to Sup/Sed.
Liver and muscle glycogen content are shown in Table 3. Liver glycogen levels were higher in supplemented rats, regardless of their physical activity (Sup/Ex and Sup/Sed). Muscle glycogen was significantly higher in the Ctrl/Ex group than in Ctrl/Sed.
Table 3.
Tissue glycogen content. Values expressed as mean (±standard deviation).
| Groups | Liver glycogen (mg%) | Muscle glycogen (mg%) |
|---|---|---|
| Sup/Ex | 16.2 (3.4) ‡ | 3.5 (.3) |
| Ctrl/Ex | 6.4 (1.6) | 4.2 (.4) * |
| Sup/Sed | 12.4 (4.8) * | 3.0 (1.1) |
| Ctrl/Sed | 5.7 (1.7) | 2.7 (.8) |
* p < 0.05 when compared to Ctrl/Sed
‡ p < 0.05 when compared to Ctrl/Ex.
Protein and RNA concentrations in gastrocnemius muscle are shown in Table 4. A significant difference in protein concentration was only observed between the groups Sup/Sed and Sup/Ex, with reduced protein concentration in the Sup/Ex compared to the Sup/Sed group. An increase in RNA levels was observed for the Sup/Ex group compared to both groups Ctrl/Ex and Sup/Sed.
Table 4.
Muscle protein and RNA content. Values expressed as mean (±standard deviation).
| Groups | Protein (mg%) | RNA (ug/100 mg) |
|---|---|---|
| Sup/Ex | 3.92 (1.22) | 147.05 (19.98) ‡ § |
| Ctrl/Ex | 5.52 (2.14) | 124.00 (7.78) |
| Sup/Sed | 6.37 (.47) † | 118.66 (9.43) |
| Ctrl/Sed | 5.08 (1.91) | 128.86 (15.50) |
† p < 0.05 when compared to Sup/Ex
‡ p < 0.05 when compared to Ctrl/Ex
§ p < 0.05 when compared to Sup/Sed.
Plasma glucose concentration is shown in Table 5. It was significantly higher in Ctrl/Sed compared to Ctrl/Ex and Sup/Sed. In this case, plasma glucose concentration was similar for Sup/Ex and Ctrl/Sed. Regarding FFA (Table 1), the exercised groups (Sup/Ex and Ctrl/Ex), irrespective of their diets, had lower plasma FFA levels than the sedentary groups (Sup/Sed and Ctrl/Sed).
Table 5.
MBlood glucose and FFA. Values expressed as mean (±standard deviation).
| Groups | Glucose (mg.dL-1) | FFA (mEq.L-1) |
|---|---|---|
| Sup/Ex | 113 (14) § | .21 (.06) |
| Ctrl/Ex | 105 (8) | .37 (.09) |
| Sup/Sed | 101 (19) | .93 (.10) † |
| Ctrl/Sed | 117 (13) ‡ § | .94 (.20) ‡ |
† p < 0.05 when compared to Sup/Ex
‡ p < 0.05 when compared to Ctrl/Ex
§ p < 0.05 when compared to Sup/Sed.
Plasma amino acid concentrations are shown in Figure 1 and were also measured 24 h after the last workout session. Tyrosine, alanine and leucine concentrations were increased in the Ctrl/Sed group, compared to the Sup/Sed group.
Figure 1.
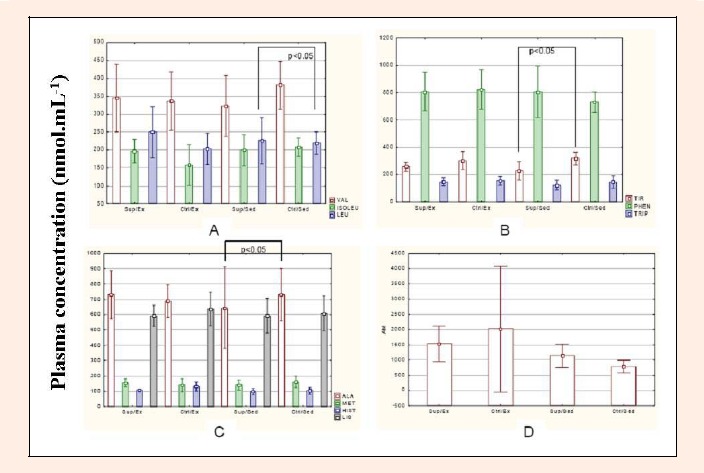
Plasma amino acid and ammonia concentration in the experimental animals. A = Branched Chain Amino Acids (valine, isoleucine, leucine); B = aromatic amino acids (tyrosine, phenylalanine, tryptophan); C = neutral and sulfur amino acids (alanine, methionine, histidine, lysine); D = ammonia. * =
Discussion
This study investigated whether BCAA supplementation had any beneficial effects on body growth, long bone and muscle growth in long-term exercise trained young male rats.
The main findings were as follows: physical exercise showed to be responsible for an impressive reduction of proteoglycan synthesis in cartilage tissue, and BCAA supplementation was capable of slightly attenuating this reduction. In addition, BCAA supplementation seemed to enhance glycogen stores in the liver, but not in the muscle. Exercise and BCAA supplementation together resulted in higher muscle total RNA and reduced muscle protein content compared to sedentary supplemented rats.
Food intake was lower in trained groups than in sedentary groups, especially among supplemented animals. This fact supports what has been observed in some studies, which have stated that exercise has an anorexic effect (Eckel and Moore, 2004). Also, authors such as Flier, 2006 have demonstrated that BCAA, especially leucine, can act on central receptors and signal anorexigen peptide synthesis, probably via mTOR. This could help explaining our results.
Nutrition and physical activity are known environmental factors that can influence body growth. Dietary protein has to be highlighted, as rats fed a 20% protein diet for 7 days had increased body growth, muscle growth, epiphyseal cartilage width and epiphysis length compared to a 5% protein diet group. Indeed, it is known that body growth and long bone growth are correlated (Yahya et al., 1994). According to these authors, in an unfavorable metabolic situation such as a low protein diet, the body tends to preserve long bone growth, although muscle and cartilage growth are compromised first.
Our study showed that body growth has been compromised in response to physical training despite the fact that protein intake and long bone growth were preserved. These findings were in accordance with our previous study (Ribeiro et al., 2010)
The groups submitted to physical training showed higher cartilage weight. Nevertheless, to our surprise, sulphate incorporation, an indirect marker of proteoglycan synthesis (Yahya et al., 1994) was extremely reduced in those trained groups.
The importance of measuring proteoglycan synthesis relies on the fact that these molecules, together with a variety of others, are components of the extracellular matrix. The ECM turnover is influenced by physical activity. Both transcriptional and posttranslational modifications, as well as local and systemic release of growth factors, are enhanced following exercise (Kjaer, 2004).
On the other hand skeletal muscle is extremely sensitive to body growth, as it is the body’s greatest compartment of fat-free mass including protein. Thus, bone and muscle growth are intimately coupled. Endocrine and environmental stimuli allow bone growth, which stimulates muscle stretching in a passive way. Consequently, protein synthesis increases and growth occurs (Ribeiro and Tirapegui, 1996).
Cartilage weight, however, augmented with physical activity. As a nutritional marker, in our experimental conditions, we expected that it would be reduced in trained groups such as protein deficient rats in other studies (Yahya et al., 1994). This can mean that other mechanical factors (load) might be taking place, as further indicated.
In our study, proteoglycan synthesis was measured 24h after the last training session in order to analyze the chronic effects of regular physical training and BCAA supplementation. One of the limitations of this determination is that no analyzes were performed at the beginning of training (before the first week).
It is known that acute exercise performed by adult rats, especially exercise characterized by eccentric contraction, increases collagen synthesis, which is a component of the extracellular matrix, both in tendons and muscle (Heinemeier et al., 2007). However, in very young animals, the effect may be different. van Weeren et al., 2008 working with foals aged 10 days to 18 months, observed that they featured a reduction in glucosaminoglycan and augmentation collagen synthesis in the proximal articular surface of the proximal phalanx of the right hind limb when submitted to a 30% workload increase, while at the same time, posttranslational modifications of collagen were augmented. This author concluded that increased exercise in these very young animals contributed to advanced maturation of the cartilage extracellular matrix. This hypothesis could explain why in our case trained rats featured heavier cartilage with reduced proteoglycan synthesis.
From the comparison between trained groups (Sup/Ex and Ctrl/Ex), we can observe higher values of sulfate incorporation in Sup/Ex animals. The existence of proteoglycan leucine-rich molecules is known, and they have been studied not only for their role on bone and cartilage growth, but also for their role in many metabolic functions (Schaefer and Iozzo, 2008). From these observations, we can previously hypothesize that BCAA supplementation could be important to induce, at least in part, a higher production of proteoglycans in cartilage of young trained animals; thus neutralizing the effects of early cartilage maturation that seems to occur with exercise training at very young ages.
One of the limitations of this study is that collagen synthesis and other cartilage functional determinations were not measured, since this work aimed on more general growth parameters. This can also be a direction for future investigations.
Regarding glycogen synthesis, BCAA supplementation was much more effective in increasing glycogen reserves in the liver than in the muscle, considering that the animals were in 24h fasting and in resting state, and this is especially interesting if one considers that the BCAA dose in this study was not very high (45 mg/body weight/day). Our data support what others have observed with histidine (Tipton and Wolfe, 1998), glutamine, alanine and proline supplementation (Aoyama et al., 1993) and BCAA supplementation (Araujo et al., 2006). In the liver, several important pathways play a role in glucose homeostasis by maintaining a balance between glucose uptake and storage that is determined by glucogenesis and glycolysis, as well as glucose release that is regulated by glycogenolysis and gluconeogenesis (Baquet et al., 1990). Some key enzyme sites regulate the interaction between glycolytic and gluconeogenic pathways, especially during fasting (e.g., glucose 1,6 di-phosphate/fructose 1,6 bisphosphatase). Morifuji et al., 2005 trained and supplemented adult rats with whey proteins, which has plenty of BCAAs, and observed a higher activation of fructose 1,6 bisphosphatase and a lower activation of glucose-6-phosphatase in the liver, resulting in a significant increase in liver glycogen storage probably due to the glucose-alanine cycle.
Regarding muscle protein concentration and total RNA, trained supplemented rats were found to have lower protein levels than sedentary supplemented animals, while the opposite was observed for RNA concentrations. This effect on muscle RNA levels was not observed in trained rats that received a 21% protein diet (Ribeiro et al., 2010) and hence is especially due to BCAA supplementation.
Our results indicate that despite having a higher capacity for synthesizing protein in skeletal muscle, BCAA supplementation did not result in higher muscle protein concentration in trained rats. This is in accordance with another study, in which oral administration of 270 mg leucine to rats enhanced phosphorylation of the eukaryotic initiation factor (eIF) (4F-BP1), but no differences in protein synthesis were found in the liver, which led them to conclude that the translation of mRNA is disjointed from the rates of total protein synthesis but is related to the degree of SK61 phosphorylation (Anthony et al., 2001). Tipton & Sharp (2005) demonstrated that the addition of insulin or BCAA to perfused muscle increased polypeptide initiation and the protein synthesis ratio, concluding that the acute responses of protein synthesis to various types of low activity exercise in slow fiber muscles seemed to be under translational control, while pretranslational control was more evident in chronic alterations caused by muscle activity.
In our experimental protocol of moderate intensity physical activity, all metabolic parameters were measured in order to characterize the chronic effects of exercise, which might highlight the final mechanisms of protein synthesis. In addition to the effects of training per se, some authors have suggested that there is either an enhancing effect on the synthesis or an inhibitory effect on the proteolysis caused by ingestion of either BCAA or leucine alone (Zanchi et al., 2008; Norton and Layman, 2006).
According to our results, BCAA supplementation increased the availability of plasma glucose when accompanied by a systematic training protocol. As it is known, both glycolysis and oxidative phosphorylation provide the energy bone and cartilage cells need; and recent evidence suggests that other amines (creatine) also play an energetic role in these tissues (Gualano et al., 2010). Thus we can consider the possibility that BCAA in this case may have contributed to an increased energy generation in this site due to their capacity of ATPs generation from their carbon skeletons.
On the other hand, a lower plasma FFA level was observed in trained rats (Sup/Ex and Ctrl/Ex) compared to sedentary animals (Sup/Sed and Ctrl/Sed). This parameter results from both the release of FFA from adipose tissue and liver as well as FFA uptake by muscle, indicating that exercise has enhanced these metabolic reactions, as expected.
Plasma tyrosine, alanine and leucine levels presented significant changes after intervention. Plasma tyrosine concentration was significantly lower in Sup/Sed animals than in the Ctrl/Sed group. Plasma alanine levels were higher in Sup/Ex than in Sup/Sed, while leucine levels were higher in the latter group (Sup/Sed) compared to Ctrl/Sed. Some interesting trends can be observed. First, in the Sup/Sed group, BCAA supplementation reduced the plasma concentration of tyrosine, an essential aromatic amino acid, which is used to indicate protein catabolism, since it is not generated or utilized by any other oxidation pathway (Libby and Goldberg, 1978).
Another relevant aspect, regarding the plasma amino acid profile, is the reduction of alanine in the Sup/Sed group compared to Sup/Ex and Ctrl/Sed groups. The role of alanine in the glucose-alanine cycle as an energy source for skeletal muscle has been well established (Pascoe and Gladden, 1996). In the present study, alanine concentrations were reduced in the Sup/Sed group compared to the other groups, indicating that alanine is less available for glucose synthesis. In the same group, plasma glucose was reduced compared to Ctrl/Sed. Considering the trained rats regardless of their diet, plasma alanine levels were the same, although a reduction in this parameter would be expected in the supplemented groups (Sup/Sed and Sup/Ex). This result suggests that BCAA supplementation during exercise may optimize alanine use to maintain plasma glucose levels and reduce proteolysis caused by an unfavorable metabolic situation for optimal growth.
Conclusion
From our results, we can conclude that BCAA supplementation did not have major effects on improving growth on trained rats, but features mild results in enhancing liver glycogen deposition, reducing proteolysis and attenuating proteoglycan synthesis reduction associated to physical training.
More studies are necessary to identify the mechanisms underlying cartilage maturation, body growth and BCAA supplementation associated to exercise.
Acknowledgments
This research was supported by FAPESP (Sao Paulo State Research Support Foundation) and CNPq (National Council of Development and Research). We also thank Dr. Úrsula Lanfer and Dr. Rosa Maria Cerdeiros for helping in the plasma aminogram analyses
Biographies
Patricia Lopes de Campos-Ferraz
Employment
Postdoctoral Fellow at University of Sao Paulo.
Degree
PhD.
Research interests
Protein metabolism, obesity and exercise.
E-mail: patlcamp@yahoo.com.br
Sandra Maria Lima Ribeiro
Employment
Professor at University of Sao Paulo, School of Arts, Science and Humanities.
Degree
PhD.
Research interests
Nutrition, aging and exercise.
E-mail: smlribeiro@usp.br
Silmara dos Santos Luz
Employment
Applied Human Nutrition at the University of Sao Paulo.
Degree
PhD.
Research interests
Nutrition, exercise and eating disorders, protein metabolism and growth.
E-mail: silmaral@yahoo.com
Antonio Herbert Lancha Jr.
Employment
Professor at School of Physical Education and Sport of the University of Sao Paulo.
Degree
PhD.
Research interests
Protein metabolism, amino acid supplementation and exercise.
E-mail: lanchajr@usp.br
Julio Tirapegui
Employment
Professor at Faculty of Pharmaceutical Sciences of the University of São Paulo.
Degree
PhD.
Research interests
Protein metabolism and exercise.
E-mail: tirapegu@usp.br
References
- Anthony T.G., Anthony J.C., Yoshizawa F., Kimball S.R., Jefferson L.S. (2001) Oral administration of leucine stimulates ribosomal Protein mRNA translation but not global rates of protein synthesis in the liver of rats. Journal Nutrition 131, 1171-1176 [DOI] [PubMed] [Google Scholar]
- Aoyama Y., Tsuda T., Hitomi-Ohmura E., Yoshida A. (1993) Activities of some regulatory enzymes of carbohydrate metabolism in the liver of rats fed a histidine-excess diet. Comparative Biochemical Physiology - Comparative Physiology 104, 381-388 [PubMed] [Google Scholar]
- Araujo Jr J., Falavigna G., Rogero M.M., Pires I.S.O., Pedrosa R.G., Castro I.A., Donato J., Jr, Tirapegui J. (2006) Effect of chronic supplementation with branched-chain amino acids on the performance and hepatic and muscle glycogen content in trained rats. Life Sciences 79, 1343-1348 [DOI] [PubMed] [Google Scholar]
- Association of Official Analytical Chemists Official Methods of Analysis of AOAC (1980). National Academic Press, Washington [Google Scholar]
- Baquet A., Hue L., Meijer A.J., van Voerkom G.M., Plomp P.J. (1990) Swelling of rat hepatocytes stimulates glycogen synthesis. Journal Biological Chemistry 265, 955-959 [PubMed] [Google Scholar]
- Blomstrand E., Saltin B. (1999) Effect of muscle glycogen on glucose, lactate and amino acid metabolism during exercise and recovery in human subjects. Journal Physiology 514(1), 293-302 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Busse W.D., Carpenter F.H. (1976) Synthesis and properties of carbonyl bis (methionyl) insulin, a pro-insulin analogue which is convertible to insulin by cyanogen bromide cleavage. Biochemistry 20, 1649-1657 [DOI] [PubMed] [Google Scholar]
- Caine D., Lewis R., O'Connor P., Howe W., Bass S. (2001) Does gymnastics training inhibit growth of females? Clinical Journal Sports Medicine 11, 260-270 [DOI] [PubMed] [Google Scholar]
- Eckel L.A., Moore S.R. (2004) Diet-induced hyperphagia in rat is influenced by sex and exercise. American Journal Physiology - Regulatory Integration Comparative Physiology 287, R1080-R1085 [DOI] [PubMed] [Google Scholar]
- Flier J.S. (2006) Neuroscience: Regulating energy balance: the substrate strikes back. Science 3312(5755), 861-864 [DOI] [PubMed] [Google Scholar]
- Frantz N.Z., Nelssen J.L., Andrews G.A., Tokach M.D., Goodband R.D., DeRouchey J.M., Dritz S.S. (2008) The effects of dietary nutrients on osteochondrosis lesions and cartilage properties in pigs. American Journal of Veterinary Research 69, 617-624 [DOI] [PubMed] [Google Scholar]
- Georgopoulos N.A., Roupas N.D., Theodoropoulou A., Tsekouras A., Vagenakis A.G., Markou K.B. (2010) The influence of intensive physical training on growth and pubertal development in athletes. Annals New York Academy of Sciences 1205, 39-44 [DOI] [PubMed] [Google Scholar]
- Gualano B., Artioli G.G., Poortmans J.R., Lancha Jr A.H. (2010) Exploring the therapeutic role of creatine supplementation. Amino Acids 38, 31-44 [DOI] [PubMed] [Google Scholar]
- Hargreaves M. (1998) Skeletal muscle glucose metabolism during exercise: implications for health and performance. Journal Science Medicine Sport 1, 195-202 [DOI] [PubMed] [Google Scholar]
- Hassid W.Z., Abraham S. (1957) Chemical procedures for analysis of polysaccharides. Methods Enzymology 3, 34-50 [Google Scholar]
- Heinemeier K.M., Olesen J.L., Haddad F., Langberg H., Kjaer M., Baldwin K.M., Schjerling P. (2007) Expression of collagen and related growth factors in rat tendon and skeletal muscle in response to specific contraction types. Journal Physiology 582, 1303-1316 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hughes P.C.R., Tanner J.M. (1970) A longitudinal study of the growth of the black-hooded rat: methods of measurements and rates of growth for skull, limbs, pelvis, nose-rump and tail lengths. Journal Anatomy 106, 34-370 [PMC free article] [PubMed] [Google Scholar]
- Kjær M. (2004) Role of extracellular matrix in adaptation of tendon and skeletal muscle to mechanical loading. Physiology Reviews 84, 649-698 [DOI] [PubMed] [Google Scholar]
- Lancha A.H., Jr, Recco M.B., Abdalla D.S.P., Curi R. (1995) Effect of aspartate, asparagine and carnitine supplementation in the diet on metabolism of skeletal muscle during a moderate exercise. Physiology Behavior 57, 367-371 [DOI] [PubMed] [Google Scholar]
- Libby P., Goldberg A.L. (1978) Leupeptin, a protease inhibitor, decreases protein degradation in normal and diseased muscles. Science 199, 534-536 [DOI] [PubMed] [Google Scholar]
- Lowry D.H., Rosebrough N.J., Farr A.L., Rendall R.J. (1951) Protein measurement with Folin phenol reagent. Journal Biological Chemistry 193, 266-275 [PubMed] [Google Scholar]
- Morifuji M., Sakai K., Sugiura K. (2005) Dietary whey protein modulates liver glycogen level and glycoregulatory enzyme activities in exercise-trained rats. Experimental Biological Medicine 230, 23-30 [DOI] [PubMed] [Google Scholar]
- Munro H.N., Fleck A. (1966) The determination of nucleic acids. Methods Biochemical Analyses 14, 113-176 [DOI] [PubMed] [Google Scholar]
- National Research Council Commission on Life Sciences Institute of Laboratory Animal Resources (1996) Guide for the Care and Use of Laboratory Animals. (USA), National Academy Press, Washington, D.C. [Google Scholar]
- Newsholme E.A., Leech A.R. (1988) Biochemistry for the medical sciences. 2ndedition John Willey, New York [Google Scholar]
- Nicastro H., Artioli G.G., Costa Ados S., Solis M.Y., da Luz C.R., Blachier F., Lancha A.H., Jr. (2011) An overview of the therapeutic effects of leucine supplementation on skeletal muscle under atrophic conditions. Amino Acids 40(2), 287-300 [DOI] [PubMed] [Google Scholar]
- Norton L.E., Layman D.K. (2006) Leucine regulates initiation of protein synthesis in skeletal muscle after exercise. Journal Nutrition 136, S533-S537 [DOI] [PubMed] [Google Scholar]
- Pascoe D.D., Gladden L.B. (1996) Muscle glycogen resynthesis after short term, high intensity exercise and resistance exercise. Sports Medicine 2, 98-118 [DOI] [PubMed] [Google Scholar]
- Reeves PG., Nielsen F.H., Fahey G.C., Jr (1993) AIN-93 purified diets for laboratory rodents: final report of the American Institute of Nutrition, Ad Hoc Writing Committee on the Reformulation of the AIN-76 Rodent Diet. Journal Nutrition 123, 1939-1951 [DOI] [PubMed] [Google Scholar]
- Regow B.J.M., Cornelissen P.J.H.C., Helder R.A.P., Spijkers J.B.F., Weber Y.M.M. (1971) Specific determination of free fatty acid in plasma. Clinical Chemical Acta 31, 187-195 [DOI] [PubMed] [Google Scholar]
- Ribeiro S.M.L., Tirapegui J. (1996) Insulin-like growth factor-1.Some relations among body and tissue growth, physical exercise and diet. Cadernos de Nutrição 10, 30-47 [Google Scholar]
- Ribeiro S L., Rogero M.M., Bacurau R.F.P., Campos P.L., Luz S.S., Lancha A.H., Jr, Tirapegui J. (2010) Effects of different levels of protein intake and physical training on growth and nutritional status of young rats. Journal of Nutritional Vitaminology and Science 57, 177-184 [PubMed] [Google Scholar]
- Schaefer L., Iozzo R.V. (2008) Biological functions of the small leucine-rich proteoglycans: from genetic to signal transduction. Journal Biological Chemistry 283, 21305-21309 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tipton K.D., Wolfe R.R. (2001) Exercise, protein metabolism and muscle growth. International Journal Sport Nutrition Exercise Metabolism 11, 109-132 [DOI] [PubMed] [Google Scholar]
- Tipton K.D., Wolfe R.R. (1998) Exercise induced changes in protein metabolism. Acta Physiologica Scandinavica 162, 377-387 [DOI] [PubMed] [Google Scholar]
- Tipton K., Sharp C.P. (2005) The response of intracellular signaling and muscle-protein metabolism to nutrition and exercise. European Journal Sport Science 5, 107-201 [Google Scholar]
- Tirapegui J., Yahya Z.A.H., Bates P.C., Millward D.J. (1994) Dietary protein, glucocorticoids and the regulation of muscle growth in the rat. Clinical Science 87, 599-606 [DOI] [PubMed] [Google Scholar]
- Tom A., Nair K.S. (2006) Assessment of Branched-Chain Amino Acid Status and Potential for Biomarkers. Journal of Nutrition 136, S324-330 [DOI] [PubMed] [Google Scholar]
- Trinder P. (1969) Determination of blood glucose using 4-aminophenazone as oxygen acceptor. Journal Clinical Pathology 22, 246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Weeren P.R., Firth E.C., Bromer B., Hyttinen M. M., Helminen A.E., Rogers C.W., Degroot J., Brama P.A. (2008) Early exercise advances the maturation of glucosaminoglycans and collagen in the extracellular matrix of articular cartilage in the horse. Equine Veterinary Journal 40, 128-35 [DOI] [PubMed] [Google Scholar]
- Yahya Z.A., Tirapegui J.O., Bates P.C., Millward D.J. (1994) Influence of dietary protein, energy and corticosteroids on protein turnover, proteoglycan sulfation and growth of long bone and skeletal muscle in the rat. Clinical Science 87, 607-618 [DOI] [PubMed] [Google Scholar]
- Zanchi N.E., Nicastro H., Lancha A.H., Jr (2008) Potential antiproteolitic effects of L-leucine: observations of in vitro and in vivo studies. Nutrition & Metabolism 5, 20. [DOI] [PMC free article] [PubMed] [Google Scholar]