

Abstract
During a day of skiing thousands of repeated contractions take place. Previous research on prolonged recreational alpine skiing show that physiological changes occur and hence some level of fatigue is inevitable. In the present paper the effect of prolonged skiing on the recruitment and coordination of the muscle activity was investigated. Six subjects performed 24 standardized runs. Muscle activity during the first two (PREskiing) and the last two (POSTskiing) runs was measured from the vastus lateralis (VL) and rectus femoris (RF) using EMG and quantified using wavelet and principal component analysis. The frequency content of the EMG signal shifted in seven out of eight cases significantly towards lower frequencies with highest effects observed for RF on outside leg. A significant pronounced outside leg loading occurred during POSTskiing and the timing of muscle activity peaks occurred more towards turn completion. Specific EMG frequency changes were observed at certain time points throughout the time windows and not over the whole double turn. It is suggested that general muscular fatigue, where additional specific muscle fibers have to be recruited due to the reduced power output of other fibers did not occur. The EMG frequency decrease and intensity changes for RF and VL are caused by altered timing (coordination) within the turn towards a most likely more uncontrolled skiing technique. Hence, these data provide evidence to suggest recreational skiers alter their skiing technique before a potential change in muscle fiber recruitment occurs.
Key points.
The frequency content of the EMG signal shifted in seven out of eight cases significantly towards lower frequencies with highest effects observed for RF.
General muscular fatigue, where additional specific fibers have to be recruited due to the reduced power output of other fibers, did not occur.
A modified skiing style towards a less functional and hence more uncontrolled skiing technique seems to be a key issue with respect to the influence on muscle recruitment for applied prolonged skiing session.
Key words: Muscle coordination, recruitment, electromyography, wavelet analysis
Introduction
During sustained recreational alpine skiing our bodies experience changes during the day that alter the way in which we perform. From a physiological point of view we can consider that recreational alpine skiing reaches a state of fatigue where physiological changes occur yet the activity can still be sustained. Those physiological processes are described within the skiing sports science literature for both, alpine ski racing (Turnbull et al., 2009) and recreational alpine skiing (Scheiber et al., 2009; Seifert et al., 2009; Tesch et al., 1978) as well. For recreational skiing fatigue is associated with increased risk of injury and reduced pleasure of the activity (Hunter, 1999). An increase in the number of injuries after 2-3 hours of skiing has been previously attributed to fatigue in adult skiers, with the incidence of injury increasing as the day progresses (Meyers et al., 2007). A more in-depth understanding of ongoing changes in muscle activity during a prolonged skiing session would provide an extension of primarily physiological measures previously reported in the literature. Knowing how skiers change their overall skiing technique based on shifts in control or altered recruitment within a muscle would serve useful information not only to improve the comfort and enjoyment, but also the safety in recreational alpine skiing (Hintermeister et al., 1997).
One consistent finding of the previous work is that there is a high level of effort by the knee extensor muscles during skiing. Numerous authors have reported that muscle contraction forces can reach upwards of 100-150% of maximal voluntary contraction (MVC) in the outside leg when making a turn (Berg et al., 1995, Hintermeister et al., 1997). Recently specific functional demands on the different knee extensor muscles in recreational skiers have been investigated (Kröll et al., 2010). Contrary to previously shown co-loading of the inside leg while skiing carving turns (Mueller and Schwameder, 2003), the results for the vastus lateralis muscle (VL) did not support the co-loading function for recreational skiing (Kröll et al., 2010). This muscle seems to be relatively inactive for the inside leg. However, they showed the functional demand for rectus femoris muscle (RF) of the inside leg is very high when skiing recreationally (Kröll et al., 2010). Assuming this is accurate, the demand on RF is greater compared to VL. This is most likely due to its dual function of influencing knee extension and hip flexion, for either the inside or outside leg.
Prolonged submaximal or maximal muscle contractions affect the capacity of muscles to generate forces (Smilios et al., 2009) and alters muscle activity as measured by surface electromyography (EMG) (Wakeling et al., 2001). These alterations occur and are quantified in three different domains:
EMG intensity domain: The integrated EMG signal during a moderate load muscular endurance session (squat exercise; 4 sets of 20 repetitions; 50% of one repetition maximum) increases within each set and from set to set (e.g.: squat exercise; 4 sets of 20 repetitions; 50% of one repetition maximum; Smilios et al., 2009). This increase has been attributed to the recruitment of the additional muscle fibers required to overcome fatigue and maintain power output.
EMG frequency domain: A decrease in the frequency content of the EMG signal at maximum workloads can be observed during fatigue (e.g. So et al., 2009). These changes have been related to different mechanisms. A decreased conduction velocity occurs with an increase of metabolites such as lactate (Brody et al., 1991). Recruitment adaptation within the muscle to maintain the power output is another possible reason since motor units with higher mean frequency tend to fatigue to a greater extent than those with relatively lower mean frequency (Moritani et al., 1982). Wakeling and Pascual (2001) reported a significant reduction in EMG intensity at low frequencies and a significant increase at high frequencies for submaximal running which is different to the fatigue related changes seen for maximal fatiguing contractions (So et al., 2009). However, they suggest that the pattern of muscle recruitment may change during sustained sub- maximal exercise.
EMG time domain (Coordination of the muscle, e.g. on/off): Although pedaling is standardized to a large extent since a bicycle constrains lower extremity movements, alterations in the coordination strategies during exhaustive pedaling exercise can be observed. (Dorel et al., 2009). Apriantono et al., 2006 showed for the instep kicking task in soccer that induced leg muscle fatigue disturbs kicking performance, leads to a less coordinated kicking motion, thereby making players more susceptible to injury. For a task such as alpine skiing, where the coordinative demand is very high, it can be assumed that alterations in coordination strategies may occur to a certain degree thus yielding an altered mechanical output that might also be less functional.
Hence, surface EMG requires analysis methods which permit the signal intensities to be simultaneously resolved in time and frequency domains to quantify alterations in muscle activity caused by fatigue. Among different time-frequency analysis methods, wavelet transform has been reported to be appropriate for analyzing the EMG signal during dynamic contractions (Karlsson et al., 2000; von Tscharner, 2000). The wavelet decomposition technique has been previously used for several dynamic tasks in non- fatigued (e.g. Wakeling et al., 2006), sub-maximal fatigued (von Tscharner, 2002, Wakeling et al., 2001) and maximal fatigued exercises (So et al., 2009). This method was previously applied to recreational alpine skiing to gain functional insights in the knee extensor muscles. (Kröll et al., 2010).
Consequently, the aim of the present study was to investigate how knee extensor muscles respond to sub maximal fatigue during recreational alpine skiing with respect to the recruitment and coordination of the muscle activity. As representative knee extensor muscles the one joint VL and two joint RF were investigated. We hypothesized that there would be changes in the intensity and frequency domain as well as in the general coordination pattern of muscle activity within the main knee extensor muscles during a recreational skiing session. Due to the higher functional demand on RF compared to VL (Kröll et al., 2010) the magnitude of these changes were expected to be larger on the biarticular RF.
Methods
The current study is a part of a larger project in recreational alpine skiing. In Part One, the relationship and prediction of common physiological fatigue indices during downhill skiing (Seifert et al., 2009) were investigated. In Part Two, the functional aspects and muscle recruitment of selected knee extensor muscles were addressed (Kröll et al., 2010). This paper addresses the effect of prolonged skiing on the recruitment and coordination of muscle activity. As the data collection occurred simultaneously with Parts One and Two, some methodological details will be referred to from these previous publications.
Subjects
Ten healthy female subjects (22.7 yr ± 4.0 s.d.) provided informed consent to participate in this study. The local Ethics Committee of Salzburg approved the research protocol and all participants were fully acquainted with the nature of the study before they gave their informed written consent. To get a homogenous sample, subjects were selected according to their skiing ability and amount of skiing days per year. All subjects were of the intermediate level based on the Austrian Ski Teaching Concept (Woerndle, 2007). Intermediate level skiers are able to perform short and long radii turns on prepared terrains. In flat terrain, intermediate skiers are able to execute carved turns, but perform mostly skidded turns on steep terrain.
Experimental design
All subjects followed a standardized 15 minute warm-up on a cycle ergometer and two warm-up runs on the ski slope. Skiers were then instructed to perform two measurement runs to determine the PREskiing condition. Then the skiers had to perform 20 runs where no biomechanical data collection took place. Finally skiers performed two measurement runs to determine the POSTskiing condition. To standardize the length of turns and distance skied across the fall-line, subjects skied for all 24 runs through a standardized corridor. The skiing session lasted about three hours in which the total skiing time was about 40 minutes. Within each run, there were three pitch changes; for this study only the first one (top of run (TOP), 21°, on average 22 turns) and the last one (end of run (END), 13°, on average 28 turns) were considered. For more detailed information concerning the slope and the skis used see elsewhere (Kröll et al., 2010; Seifert et al., 2009).
Physiological measurements
Overall run time and heart rate (HR) at the end of each run was recorded (Polar, Finland). To reach a standardized load during the skiing session, verbal feedback on finishing time and HR were provided to each skier at the end of each run as absolute values. For further analysis percent of maximal HR (%HRmax) was calculated using 220 - age. Earlobe blood samples in the amount of 20 microliters were collected following the second (PREskiing) and immediately after the 24th (POSTskiing) run. The blood samples were analyzed for lactate (LA) (Biosen 5140, EKF-Diagnostic GmbH, Magdeburg, Germany). LA was considered as control parameter for interpretation of the shifts in EMG spectrum since the level of accumulated LA indicates a close relation to the shift in the EMG spectrum towards lower frequencies during fatigue (Horita and Ishiko, 1987; Tesch et al., 1983).
EMG and kinematics measurement
Data acquisition took place unilaterally on the right leg. Myoelectric activity was measured from the vastus lateralis (VL) and rectus femoris (RF) muscles using round bipolar surface electrodes (Ag/AgCL; 10mm diameter, 22mm spacing). The sensor placement procedure was based on the SENIAM recommendations (Hermens et al., 2000). The EMGs were amplified at source (bandwidth 10-500Hz, 3dB; Biovision, Wehrheim Germany) and recorded at 2000Hz (Daqcard-700, National Instruments, Austin, TX, USA; iPAQ H3800, Compaq, Houston, TX, USA). Simultaneously, a goniometer mounted on the knee joint measured the occurring knee angle (note that 180° was defined as full extension). The knee angle and myoelectric activity were recorded for the entire four measurement runs (two PREskiing / two POSTskiing).
Wavelet and principal component analysis of EMG during alpine skiing
Compared to a previous study (Kröll et al., 2010), methods used in the current study added a principal component analysis of the EMG spectra to quantify spectral shifts (Wakeling and Rozitis, 2004). The main information is derived by the first two principal components (PC I and PC II). Additional variations in the original spectra’s (e.g. movement artifacts and motor unit synchronisation) are partitioned into lower components and hence, do not obscure the main information of interest (Wakeling, 2009). Therefore it is suggested that the principal component technique is more sensitive to major changes in the EMG spectra that occur during movements and locomotion.
Wavelet transformation: The entire raw EMG signal (Figure 1A) was resolved with an EMG- specific wavelet transformation. The intensity was calculated using a filter-bank of wavelets following von Tscharner, 2000. Each wavelet has a frequency band and the time-varying intensity is calculated for this band. For each moment in time the EMG is thus represented by its intensity at a range of frequency bands (11- 432Hz), and this is termed the intensity spectrum.
Figure 1.
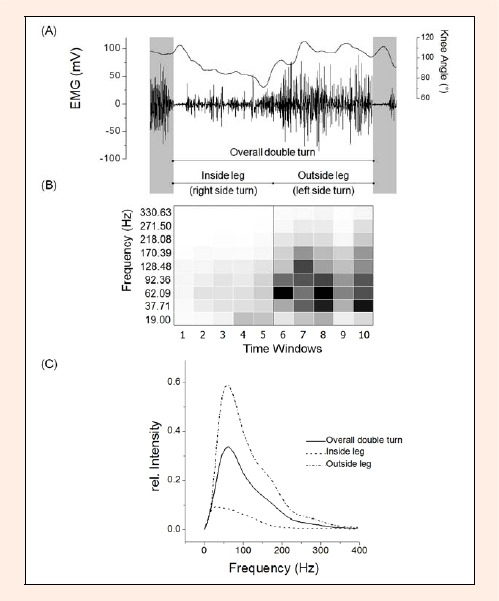
EMG trace and knee angle from the vastus lateralis (VL) during one double turn. (B) Myoelectric intensity of the turn is shown as a function of time and frequency, with high intensities denoted by dark shading. Ten time windows divide the overall double turn in different time frames. They are indicated by the vertical white lines. (C) Myoelectric intensity spectra for inside leg, outside leg and the overall double turn shown in (B)
Turn separation and time frame within a double turn: A combination of knee angle and raw EMG was used to determine the start and end of each double turn. A double turn was defined as where the right leg is the inside leg during the first turn and then is the outside leg during the second turn (Figure 1A). For 10 equal time windows along the double turn and each single wavelet the mean value of intensity was built to generate a 3-dimensional grid (X = Time; Y Frequency; Z = Intensity as shown in Figure 1B for one double turn). The mean intensity spectra was calculated for the overall double turn and was used for EMG intensity normalization (for details see Kröll et al., 2010). The overall inside leg and the overall outside leg are shown in Figure 1C. Furthermore the mean knee angle of the overall inside leg and the overall outside leg within the double turn was also calculated.
Elimination of artifacts and subjects: The same method for eliminating artifacts was used as previously described (Kröll et al., 2010). Compared to the paper with the functional analysis (n = 7), due to additional artifacts in the POSTskiing session one additional subject had to be excluded. This reduced the total number of subjects used in this analysis to six.
Principal component analysis: For the analysis n = 2223 normalized intensity spectra from the different subjects (n = 6), turn side (inside leg = 1062, outside leg = 1161), conditions (PREskiing n = 1080, POSTskiing n = 1143), run part (TOP n = 984, END n = 1239), and muscle (VL n = 1111, RF n = 1112) were considered. Each of the intensity spectra contained 10 intensities corresponding to the wavelets 1-10 and a frequency band from 11- 432 Hz (Figure 1C). All of the 2223 spectra were compiled into a data matrix. The covariance matrix was calculated from the data matrix, and used to calculate the principal components (PC) (Wakeling and Rozitis, 2004). The principal components were calculated with no prior subtraction of mean data and describe the components of the entire signal (Wakeling and Rozitis, 2004). Each EMG intensity spectrum can then be reconstructed from the vector product of the PC weightings and the PC loading scores (Figure 2). The PCs are defined by the PC weightings that are the eigenvectors for the covariance matrix, and the PC loading scores that quantify the contribution of each eigen vector to the measured signal. The majority of the signal for any given myoelectric spectrum is defined by the first two PCs (PC I and PC II). The weighting for PC I has a shape similar to that for a myoelectric intensity spectrum (Wakeling and Rozitis, 2004). Furthermore it is expected that the myoelectric intensity is highly correlated with the PC I loading score (r2 > 0.95) (Wakeling et al., 2006). The relative loading scores of PC I and PC II give a measure of the frequency of the myoelectric signal (Wakeling, 2004). The angle θ was defined by the direction of the PC I-PC II loading score vector for each EMG intensity spectrum. Angle θ was then used as a measure of the mean myoelectric frequency for each EMG-intensity spectrum (for each turn), respectively (angleθ = arctan (PC I score/PC II score) (Wakeling, 2004).
Figure 2.
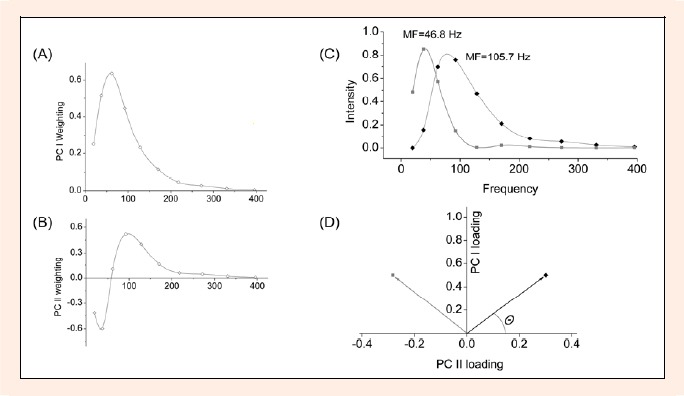
Principal component representation of EMG frequency spectra. Weighting for (A) the first principal component (PC I) and (B) the second principal component (PC II). (C) EMG-intensity spectra that can be reconstructed from vector products of the PC weightings shown in (A&B) with 0.5 PCI + 0.3 PCII shown in black and 0.5 PC I – 0.28 PC II shown in grey. The mean frequencies (MF) for the reconstructed spectra are shown. (D) Vector representation of the spectra in (C) following the same colors and symbols as in (C). The angle θ is a measure of the relative PC I and PC II loading scores and of the myoelectric frequency
Statistical analysis
Repeated measures (RM) analysis of variance (ANOVA) was used to test differences in PRE- to POSTskiing load (for overall run time, %HRmax and LA). Differences in PRE- to POSTskiing knee angle were tested with a 2(Time) x 2(Slope Part) RM-ANOVA which was performed for inside leg and outside leg separately.
The hypothesis that the myoelectric frequency differed between PRE- and POSTskiing due to the submaximal skiing session was tested with an analysis of covariance (ANCOVA). The response variable was θ. Subject code, time of measure, and slope part were used as factors in the test. The myoelectric intensity was used as a covariate. in order to establish whether the shifts in myoelectric frequency in this study were independent of variations in myoelectric intensity (Wakeling, 2009). The ANCOVA was repeated for both muscles (VL, RF) and both turn sides (inside leg, outside leg).
The hypothesis that myoelectric intensity between PRE- and POSTskiing differed was tested with a three factor ANOVA (subject code, time of measure, slope part). The response variable was the PC I loading score. The ANOVA was also repeated for both muscles and both turn sides. All data are presented as mean ± standard error of the mean (s.e.m) and statistical tests were deemed significant at α = 0.05 (tendency α = 0.1). Effect size measures (and interpretation) included partial eta squared (pη2) for all ANOVA.
Alterations in the specific coordination patterns of RF and VL (within each and between these two muscles) are described using 3-dimensional grids. Furthermore the course of the total intensity (calculated as the sum of the intensities within each spectrum) along the 10 time windows within each double turn are presented for PRE- and POSTskiing, each muscle and slope part. To get representative results for one subject the mean over all turns in a certain situation was calculated. To get a representative result for the group the same was done over all subjects.
Results
Physiological parameter and knee angle
Figure 3A depicts the overall run time for all skiers. The within subject range in finishing time was consistent and therefore no significant effects between PREskiing (100 ± 0.9 s), prolonged skiing session (103.2 ± 1.4 s) and POSTskiing (99.8 ± 1.2 s) were observed. Although the HR response showed pronounced variation between subjects, within subject range during the 24 runs was consistent (Figure 3B). PREskiing %HRmax (81.9 ± 3.9%), prolonged skiing session %HRmax (78.1 ± 4.2%) and POSTskiing %HRmax (82.3 ± 3. 6%) were not significantly different. Average blood LA concentration was 2.72 ± 0.53 mmol.L-1 after the second run. Blood LA concentration decreased significantly by the 24th run to 2.13 ± 0.46 mmol.L-1 (p < 0.035 pη2 = 0.814). Individual LA values ranged from 1.24-4.45 mmol.L-1 after the 2nd run and from 0.89-3.63 mmol.L-1 after the 24th run (Figure 3C).
Figure 3.
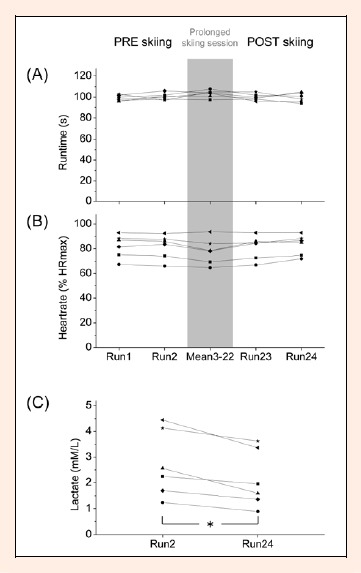
(A and B) Subjects’ overall run time and heart rate (%HRmax) in the PREskiing, prolonged skiing session (averaged) and POSTskiing condition. (C) Subjects individual blood lactate (LA) during PREskiing and POSTskiing
The average knee angle on the inside leg was relatively consistent between the two pitches and at the PREskiing and POSTskiing condition (PREskiing_TOP = 99 ± 6°, POSTskiing_TOP = 97 ± 5°, PREskiing_END = 96 ± 6° , POSTskiing_END = 92 ± 6°). Hence, the average knee angle of the inside leg was not significantly different between the PREskiing (97 ± 6°) and POSTskiing (95 ± 6°) condition. The average knee angle on the outside leg was also relatively consistent in the four situations (PREskiing_TOP = 115 ± 5°, POSTskiing_TOP = 114 ± 5°, PREskiing_END = 114 ± 4°, POSTskiing_END = 110 ± 4°) with no significant influence observed due to the prolonged skiing session (PREskiing 115 ± 4°; POSTskiing 112 ± 4°).
Overall inside leg and outside leg EMG results
The PC's were calculated from the matrix of spectra for the inside leg and outside leg EMG-intensity spectra. The first two principal components of the myoelectric intensity spectra described 97.3% of the signal. The PC I weighting (Figure 2A) took a similar form to an EMG power spectrum (Figure 1C). The myoelectric intensity for each turn correlated with the PC I loading score with a correlation coefficient of r=0.99. The PC II weighting shows negative and positive regions with a crossover at about 60 Hz (Figure 2B). Intensity spectra could be reconstructed from the vector product of the PC weightings and the PC loading scores. Reconstructed spectra with positive intensities across all frequencies (a physiological constraint) occur for a range of θ and result in two extreme spectra with mean frequencies of 46.8 Hz and 105.7 Hz (Figure 2C).
Plotting the mean scores for PC I and PC II for all analyzed situations (Figure 4), one can identify several distinct populations of activity. A distinct difference between inside leg (dashed circle) and outside leg (solid circle) is given for both muscles. Clearly higher intensities (= PC I loading) were observed for inside leg compared to outside leg for VL, while RF shows similar intensities on both turn sides. According to the angle θ (measure of myoelectric frequency) it was found that the value was smaller (=higher myoelectric frequency) compared to the outside leg for RF at all inside leg situations. The opposite was observed for VL, depicted by higher θ (=lower myoelectric frequency) for all inside leg situations.
Figure 4.
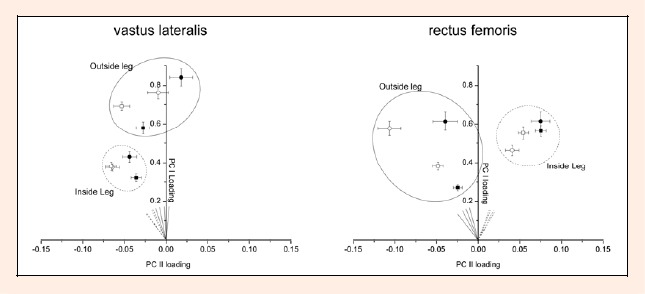
Principal component loading scores for PC I and PC II during recreational alpine skiing for m. vastus lateralis (VL, left graph) and for m. rectus femoris (RF, right graph). Each point shows the mean ± s.e.m. loading scores pooled from the six subjects. Data points within the solid circle represent outside leg while points within the dashed circle represent inside leg (solid symbols = PREskiing; open symbols = POSTskiing; circles = TOP of the run; squares = END of the run). The direction of the vectors in the PC I-PC II scoring plane represents angle θ and is visualized by solid (outside leg) and dashed lines (inside leg)
The mean scores for PC I and PC II are shown within each circle of Figure 4 for the TOP and the END of the run for PREskiing and POSTskiing, respectively. The prolonged skiing session caused a significant shift towards higher values for both muscles and both turn sides for angle θ. (Table 1). Furthermore post hoc tests on the angle θ depict overall significant/tendency effects towards higher values, except in the situation ‘RF- inside leg- TOP’. For angle θ, the highest effects (for global and post hoc test) were observed on RF outside leg (pη2 = 0.118; TOP pη2 = 0.196; END pη2 = 0.099; Table 1). For PC I loading the prolonged skiing session only caused a significant shift towards higher values in the RF of the outside leg (Table 1). However, post hoc analyses were performed for all situations. Those results show an inverse response during the prolonged skiing session depending whether skiing on TOP or at END of the run. On TOP, PC I scores decreased on the inside leg for both muscles. At the END of the run, PC I scores increased on the outside leg for both muscles (Table 1). The highest effect was observed again for RF outside leg (END pη2 = 0.104).
Table 1.
Probabilities from the analysis of covariance (ANCOVA) for the angle Θ and the analysis of variance (ANOVA) for the PC I loading score. The direction of changes is depicted by the arrows (↓=decrease; ↑=increase)
| Angle Θ | PC1 Loading | |||||||
|---|---|---|---|---|---|---|---|---|
| VL | RF | VL | RF | |||||
| Inside Leg | Outside Leg | Inside Leg | Outside Leg | Inside Leg | Outside Leg | Inside Leg | Outside Leg | |
| n | 520 | 591 | 542 | 570 | 520 | 591 | 542 | 570 |
| Subject | ||||||||
| p value | .000 | .000 | .000 | .000 | .000 | .000 | .000 | .000 |
| effect size pη2 | 0.811 | 0.808 | 0.45 | 0.826 | 0.374 | 0.323 | 0.149 | 0.472 |
| PRE to POST | ||||||||
| p value | .000 | .000 | .001 | .000 | .037 | |||
| increase / decrease | ↑ | ↑ | ↑ | ↑ | ↑ | |||
| effect size pη2 | 0.041 | 0.055 | 0.023 | 0.118 | 0.008 | |||
| Post Hoc: PRE to POST | ||||||||
| TOP | ||||||||
| p value | .000 | .000 | .000 | .035 | .024 | |||
| increase / decrease | ↑ | ↑ | ↑ | ↓ | ↓ | |||
| effect size pη2 | 0.076 | 0.064 | 0,196 | 0.019 | 0.022 | |||
| END | ||||||||
| p value | .066 (t) | .000 | .001 | .000 | .001 | .000 | ||
| increase / decrease | ↑ | ↑ | ↑ | ↑ | ↑ | ↑ | ||
| effect size pη2 | 0.013 | 0.054 | 0.039 | 0.099 | 0.031 | 0.104 | ||
Timing aspect within a double turn
Figure 5 contains descriptive information about changes caused by the prolonged skiing session with respect to timing aspects for the overall group. It can be noted that peaks in overall EMG activity occurred later in seven out of eight situations (turns) in the POSTskiing compared to the PREskiing situation (exception: VL inside leg TOP of run). Alterations within the different frequency band are shown by the grid plots. Alterations in RF are different between TOP and END of the run. A decrease in the higher frequency bands in time windows |2|3|7|8| and an increase in the lower frequency bands in time window |4|5|9|10| can be observed on TOP. A decrease in the higher frequency bands in time windows |2|3|4| and an increase in lower frequency bands on the whole outside leg (|6|7|8|9|) can be seen during END.
Figure 5.
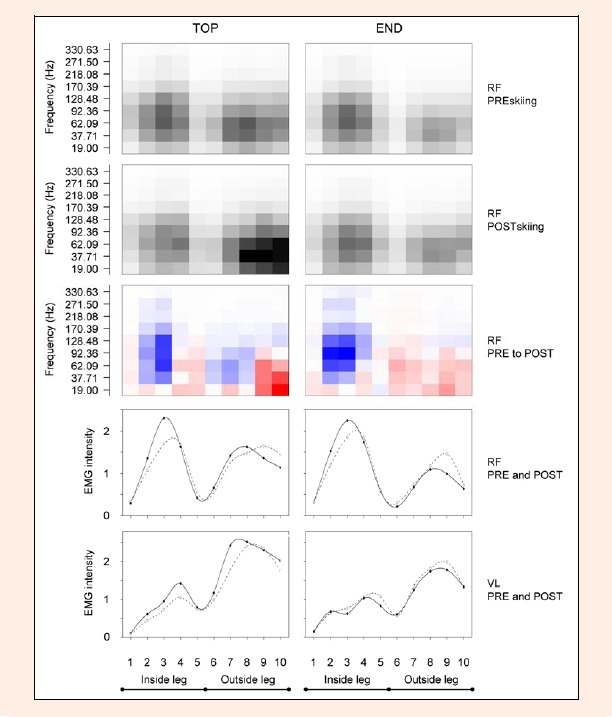
Alterations in timing aspects from PREskiing to POSTskiing of m. rectus femoris (RF) and m. vastus lateralis (VL). Left column of diagrams represent results for TOP of the run while right column represent END of the run. From top to bottom: PREskiing grid plot for RF, POSTskiing grid plot for RF (high intensities = dark shading); shift from PRE to POST for RF (blue = decrease, white = unchanged, red = increase); course of total EMG intensity for RF, course of total EMG intensity for VL (PREskiing = solid, POSTskiing = dashed)
Discussion
If classic fatigue exercises are defined as activities that cannot be sustained, then a normal recreational alpine skiing session (and the majority of daily tasks) does not reach a fatigued state. This study did not use a classic fatigue exercise protocol where the activity could no longer be sustained. Rather, this study employed a typical 3 hour skiing session at an intensity where physiological changes toward a fatigued state have been reported to occur (Seifert et al., 2009, Tesch et al., 1978) and which is indicative of typical recreational skiing.
The blood lactate level, which was recently reported to be a significant predictor for chronic stress (Seifert et al., 2009), was similar to a previous study on recreational skiers (Scheiber et al., 2009). With verbal feedback on overall run times and HR, skiers were able to maintain a constant load. A protocol identical to the present study reported base line values increased for salivary cortisol (16%) and creatine kinase levels (42%) (Seifert et al., 2009). The same study showed that muscle performance was influenced by the fact that isometric knee extension endurance decreased by 12%, but peak force was not different from PREskiing to POSTskiing. Hence, the notion that fatigue identifiable at sub maximal levels can occur within 24 runs of recreational skiing seems reasonable.
Distinct differences between inside leg and outside leg were observed in the same manner as seen previously in recreational alpine skiing (Kröll et al., 2010). Co-loading of the VL on inside leg was not observed while RF showed pronounced activity on both turn sides and the frequency content in RF of the inside leg points out a pronounced involvement of fast fibers. Furthermore, the current results demonstrate that the spectral properties and the overall intensities of muscle activity are affected by the prolonged three hour skiing session, and that the greatest effects can be observed in RF which supports our assumptions based on its functional demand (Kröll et al., 2010).
Changes in the myoelectric spectra caused by the prolonged skiing session
A shift towards lower frequencies was observed for POSTskiing for the mean frequency. Even though our results are consistent with general fatigue effects (decreased EMG frequency) there are several mechanisms which can shape myoelectric intensity and determine the myoelectric frequency (Wakeling et al., 2001). Possible mechanisms will be discussed, with particular focus on whether these can explain the changes in frequencies that were measured in this study.
Influence of muscle length
To discuss the changes in mean frequency of the EMG signal during a prolonged skiing session, it is necessary to estimate changes in muscle length since Doud and Walsh, 1995 reported decreased frequency is caused by increased muscle length (20% increase in length with an 18% decrease in EMG frequency). To estimate muscle fiber behavior solely from observation of joint performance is difficult since the fascicle does not behave exactly like the muscle-tendon unit (Ishikawa et al., 2003). Nevertheless, in a previous alpine skiing study (Kröll et al., 2010) muscle length changes were estimated based on the knee-hip-joint kinematics during recreational skiing linked with a worst case scenario calculation for differences in muscle tendon unit and fascicle behavior in a closely related movement (Chleboun et al., 2008). Although the average range of motion was about 30°, the estimated length changes were not pronounced enough to explain the changes in mean frequency from the inside leg to the outside leg of RF (mean frequency increase by 16%) and VL (mean frequency increase by 11%) (Kröll et al., 2010). The length changes for the biarticular RF which occur during a 30° knee angle decrease were less than -1% for RF while the length changes for VL were approximately +4% (Kröll et al., 2010). It was found in the current study that the influence of a prolonged skiing session on the knee angle is in general very small and not significant. Hence, the length changes in the current study seem to be negligible for RF and VL.
Consequently, the lower frequency during the POSTskiing for RF and VL cannot be explained by changes in muscle length in the present study. According to the observation by Doud and Walsh, 1995 the altered muscle length can account for at least a part of the frequency shift since changes in knee angle were minor.
Influence of lactate
Theoretical calculations have shown that the EMG frequency spectrum shifts to higher frequencies as a result of an increase in the motor unit action potential (MUAP) conduction velocity (Lindstrom et al., 1970). Hence, much of the variation in the frequency component of the EMG has been attributed to changes in MUAP conduction velocity. A decrease of pH results in a decrease of MUAP conduction velocity, and as a consequence, a decrease of EMG mean frequency (Brody et al., 1991). As metabolites such as lactates decrease pH it has been suggested that the shift in the EMG spectrum towards lower frequencies during fatigue indicates a close relation to the level of accumulated lactate within the muscle (Horita and Ishiko, 1987, Tesch et al., 1983). The lactate level in the current study differed between PREskiing and POSTskiing, but this level decreased significantly during the prolonged skiing session (Figure 3C). A shift to lower frequencies was observed for the frequency content of the EMG signal due to the prolonged skiing session. This is opposite to what the lactate effect should cause according to Horita and Ishiko, 1987 or Tesch and Komi (1983). For this reason we consider that lactate changes were not responsible for the observed changes in the intensity spectrum seen here.
Influence of muscle temperature
Next to altered muscle length or altered metabolites some of the variation in EMG frequency may be due to differences in the temperature of the muscle. The conduction velocity of nerve and muscle action potentials is a function of tissue temperature. It increases by ~5% per degree C as the temperature of the nerve increases from 29° to 38° (Kiernan et al., 2001). During an isometric contraction of the biceps brachii at different force levels it has been demonstrated that a 10° reduction in temperature results in a 32Hz reduction in centre frequency (Petrofsky and Laymon, 2005). It is therefore suggested to ensure that skin temperature is within defined limits before clinical measurements (Kiernan et al., 2001). In a complex field study like the present skiing study, environmental conditions for testing cannot be absolutely controlled. However a window of acceptable temperatures can be determined as was done in this study. Muscle temperature may vary greatly in an outdoor study due to the influence of previous exercise, clothing, and daily rhythm of environmental temperature. With respect to the previous exercise aspect, an adequate warm up process was completed which included two runs of skiing and a 15min ride on a cycle ergometer. Hence it was assumed that the muscle temperature during the PREskiing runs was similar to that during the prolonged skiing session and POSTskiing. Accordingly, subjects wore current state of the art clothing made of breathable material with the aim of minimizing heat accumulation. The change of environmental temperature due to the daily fluctuation is less pronounced on slopes with a north exposure because of the shadow effect. Consequently a slope with north exposure was selected for the experiment with an additional advantage of consistent snow conditions during the day. Nevertheless a slight increase in environmental temperature during the day occurred and could be assumed that muscle temperature increased slightly compared to the PREskiing situation. An increased muscle temperature would result in increased mean frequency (Kiernan et al., 2001, Petrofsky and Laymon, 2005). Our results depict a shift to lower frequencies due to the prolonged skiing session and this is opposite to what the temperature effect should be. For this reason we consider that increased temperature was not the mechanism underlying the observed changes in the intensity spectrum, but we estimate that if anything, the temperature effect reduced the amount of the observed changes.
Effect of recruitment patterns
The pattern of motor unit recruitment is an important factor in shaping the myoelectric intensity spectrum. Action potentials from faster fibers travel at higher conduction velocities and thus have higher mean frequencies, so an EMG frequency spectra provides information about motor unit recruitment strategies (Solomonow et al., 1990). Recently it was shown by theoretical calculations, that recruitment strategies resulting in a greater proportion of faster muscle units being active had a significantly higher mean frequency (Wakeling, 2009). Although other physiological factors can bias the EMG frequency, careful experimental and statistical design can account for such bias, and so distinct high- and low- frequency components of the EMG have been reported for a range of in vivo activities (Wakeling and Rozitis, 2004; Wakeling et al., 2006, von Tscharner, 2002). It has been suggested, these spectral characteristics are the result of altered recruitment patterns between different motor units. It does not appear that the changes in myoelectric intensity spectra that occurred during the submaximal skiing session can be explained by the length of the muscle fiber or by the decrease in physiological changes (pH level, muscle temperature). The most plausible explanation for the observed changes in myoelectric signal is that the pattern of motor unit recruitment was altered during the 3 hour skiing session.
In the classical explanation of fatigue, it would be expected that in order to maintain muscle power output required for skiing at a constant pace, more muscle fibers would have to be recruited or that they are excited at higher frequencies as they become fatigued (Wakeling et al., 2001). Fast twitch fibers are more susceptible to fatigue than the slow muscle fibers (Komi and Tesch, 1979; Moritani et al., 1982) and increased recruitment should occur for these fibers in particular. If such a fatigue effect occurs during a prolonged alpine skiing session one could assume that an increased recruitment in high frequencies should be observable along all time windows of a double turn. Figure 5 does not depict such a uniform alteration at higher frequency band. The general decrease in mean frequency combined with inconsistent changes within the time windows is contrary to the assumption, that additional fast fibers are recruited due to the reduced power output of other fibers.
From a physiological point of view those results correspond with findings on glycogen depletion during alpine skiing (Nygaard et al., 1978; Tesch et al., 1978). For most recreational downhill skiers, the major glycogen loss occurs in the slow fibers. This suggests that this fiber type produces the predominant proportion of tension development during a day of alpine skiing (Nygaard et al., 1978). On the other hand highly skilled (racers) and unskilled (beginners) skiers, which were not investigated in our experiment, also showed particular responses on fast fibers (Nygaard et al., 1978). Hence, with respect to our findings and the earlier studies on glycogen depletion we suggest that the fast fibers do not reach a state of fatigue in terms of generally reduced power output as they are not used to a large extend during recreational skiing. This argument is furthermore supported by the results of earlier published isometric peak force data which were not different from PREskiing to POSTskiing (Seifert et al., 2009).
In earlier studies it was also suggested, that glycogen depletion toward the end of the day, especially in the slow fibers, could contribute to the injury pattern which peaks toward the end of the ski day (Seifert et al., 2009).
A generally reduced power output of the slow fibers should be reflected in the EMG signal in the way that additional fibers are recruited along the double turn. But similar to the fast components, no uniform alteration along the 10 time windows in the slow frequencies can be observed (Figure 5).
Altered timing of muscle activity during a prolonged skiing session and implication on functional aspects of the skiing technique.
We suggest that the observed shifts in frequencies towards lower values in the present study are not caused by a general substitution of fibers with a concomitant reduction in power output. The most plausible explanation for the reduced frequencies is altered skiing style with modified timing of muscle activity. Depending on the turning side (inside leg vs. outside leg) different alterations occur and the shift to the lower frequencies in the POSTskiing can be explained in several ways. For example RF on TOP showed a clear decrease of the high frequency components during the earlier phases of the inside leg (Figure 5; time window 2|3). Conversely, a clear increase in the low frequency components specifically can be observed during the last part of the turn on the outside leg for RF (Figure 5; time window 9|10).
This level of detail has been made possible by the use of wavelet analysis on the myoelectric signal. The mean frequency of the power spectrum, calculated by a Fourier transformation, was previously used to evaluate fatigue during alpine skiing (Ushiyama et al., 2005). The results showed a decreasing trend in mean frequencies which appeared to be more pronounced in the afternoon compared to that in the morning. However, a discussion of this shift in frequencies was not possible in terms of an altered timing of muscles (skiing style) due to the collapse in temporal aspects of the signal. Hence, interpretation of these data is limited.
It was shown recently (Kröll et al., 2010) that the functional importance of RF during recreational skiing in the inside leg is very high. From a coordinative point of view, recreational alpine skiers should try to obtain an adequate bilateral loaded skiing technique (Mueller and Schwameder, 2003). To accomplish this, the skier has to shift weight to the inside leg by knee extension, while being careful to avoid too high of loading of the inside leg by hip flexion. The situation dependent repetitive loading (RF as knee extensor) and unloading (RF as hip flexor) activities are crucial and are reflected in the EMG signal by the involvement of more fast twitch fibers throughout the muscle contraction in the inside leg compared to the outside leg (Kröll et al., 2010).
After the prolonged skiing session, there was a decrease in high frequency components which could be a result of a reduction in the recruitment of fast components at the TOP and at the END of the run in the inside leg. The corresponding overall EMG intensities decreased on TOP for the inside leg, but increased at the END on the outside leg. Therefore, the situation dependent repetitive loading (RF as knee extensor) and unloading (RF as hip flexor) activities seem to be reduced and skiers ski more unilaterally on the outside leg. Even though the functional capacity would be enough to maintain the original skiing style, recreational skiers could preferentially ski more unilaterally, on the outside leg, in order to reduce the load on the inside leg. One could speculate that this occurs due to the necessity to have a recovery phase during each inside leg phase.
Following the biomechanical distinction of the carving technique and the traditional parallel technique (Mueller and Schwameder, 2003) we suggest that the prolonged submaximal skiing session leads to a shift in skiing style towards the traditional parallel technique. This alteration in skiing style is furthermore supported by the overall intensity results of VL which changed in the same way as the RF (Table 1). The observation that activity peaks occur later within the turn for almost all situations (in seven cases out of eight, Figure 5), after the prolonged skiing session, is another indicator of a shift to traditional parallel technique since the duration of the initiation phase for the following turn is thereby reduced (Mueller and Schwameder, 2003).
Mueller and Schwameder, 2003 suggested when skiers use carving skis with adequate technique, they maintain better sagittal balance and have improved edge steering ability that help them remain centrally positioned over their skis. The enhanced steering ability when using a co-loading technique was recently described by theoretical calculations (Heinrich et al., 2009). Furthermore, an increase in force does not occur as quickly with co-loading as with traditional parallel turn technique (Mueller and Schwameder, 2003). We speculate that the altered skiing style towards the traditional parallel turn counteracts the functional properties of carving skis. This could increase the potential risk for over edging and consequently for falls as the saggital balance and the edge steering behavior of the equipment is altered. From a skiing safety perspective, the more rapid force increase on the outside leg compared to the bilateral steered (more controlled) technique at the beginning of the skiing session may be important. It seems to be plausible that quicker and more uncontrolled force increases in combination with the self steering abilities of carving skis may increase the risk of potential falls due to sudden, unanticipated edging. We suggest that recreational skiers should be intentionally instructed to maintain a situation dependent repetitive loading (RF as knee extensor) and unloading (RF as hip flexor) activity of the inside leg to reduce the shift towards a traditional parallel technique during a prolonged skiing session.
Conclusion
Based on our interpretation of these data, we propose that the frequency decrease and intensity changes for RF and VL are caused by altered timing (coordination) within the turn. Furthermore, general muscular fatigue, where additional fibers have to be recruited due to reduced power output of specific motor units, likely does not occur. In other words, these data provide evidence to suggest recreational skiers alter their skiing technique before a potential change in muscle fiber recruitment occurs. It is important to note that the current study did not employ an extreme exercise, but used a 3 hour skiing session at an intensity which is indicative of recreational skiing. Seifert et al (2009) using slightly longer run times reported similar HR data but greater changes in physiological stress markers compared to the current study. Perhaps the actual methods used, combined with using longer ski durations or greater intensities, might identify general muscle fatigue during skiing in a way that additional specific fibers have to be recruited to prevent a reduction in power output.
For elite competitors in alpine ski racing, investigators reported heart rate (HR) and oxygen consumption (VO2) achieve or exceed maximal values during competitive alpine skiing (Tesch, 1995) and glycogen depletion of fast fibers, in particular, was identified (Nygaard et al., 1978). Hence, for alpine ski racing an increased recruitment could occur in fast twitch fibers since these fibers are susceptible to fatigue (Komi and Tesch, 1979; Moritani et al., 1982). To suggest general muscle fatigue does generally not occur in alpine skiing would exceed the scope of this study, but could be investigated using this same method with more intensive skiing applications (aim for future projects). For the applied prolonged skiing session in recreational skiers in this study the modified skiing style towards a less functional and hence more uncontrolled skiing technique seems to be a key issue with respect to the influence on muscle recruitment.
Acknowledgments
The authors would like to thank Atomic Skis for support, to all our co workers of the Department of Sport Science and Kinesiology for their technical assistance and especially to our subjects for their efforts and cooperation.
Biographies
Josef Kröll
Employment
Research assistant and PhD candidate at the Department of Sport Science and Kinesiology, and CD-Laboratory “Biomechanics in Skiing”, University of Salzburg, Austria.
Degree
MSc
Research interests
Applied biomechanics, muscle function, motor learning.
E-mail: josef.kroell@sbg.ac.at
Erich Müller
Employment
Head of the Department of Sport Science and Kinesiology, and CD-Laboratory “Biomechanics in Skiing”, University of Salzburg, Austria.
Degree
PhD, Professor.
Research interests
Biomechanics; training and coaching, motor learning.
E-mail: erich.mueller@sbg.ac.at
John G. Seifert
Employment
Associate Professor of Exercise Science at Montana State University in Bozeman, Montana, USA.
Degree
PhD
Research interests
Alpine skiing and the effects of nutrition supplements on exercise performance.
E-mail: john.seifert@montana.edu
James M. Wakeling
Employment
Director of Neuromuscular Mechanics Laboratory, School of Kinesiology,
Simon Fraser University, Vancouver, Canada.
Degree
PhD, Associate Professor.
Research interests
Mechanics of whole muscles: activity patterns, structural arrangements, power output.
E-mail: wakeling@sfu.ca
References
- Apriantono T., Nunome H., Ikegami Y., Sano S. (2006) The effect of muscle fatigue on instep kicking kinetics and kinematics in association football. Journal of Sports Sciences 24, 951-960 [DOI] [PubMed] [Google Scholar]
- Berg H.E., Eiken O., Tesch P.A. (1995) Involvement of eccentric muscle actions in giant slalom racing. Medicine and Science in Sports and Exercise 27, 1666-1670 [PubMed] [Google Scholar]
- Brody L.R., Pollock M.T., Roy S.H., De Luca C.J., Celli B. (1991) pH-induced effects on median frequency and conduction velocity of the myoelectric signal. Journal of Applied Physiology 71, 1878-1885 [DOI] [PubMed] [Google Scholar]
- Chleboun G.S., Harrigal S.T., Odenthal J.Z., Shula-Blanchard L.A., Steed J.N. (2008) Vastus lateralis fascicle length changes during stair ascent and descent. Journal of Orthopaedic and Sports Physical Therapy 38, 624-631 [DOI] [PubMed] [Google Scholar]
- Dorel S., Drouet J.M., Couturier A., Champoux Y., Hug F. (2009) Changes of pedaling technique and muscle coordination during an exhaustive exercise. Medicine and Science in Sports and Exercise 41, 1277-1286 [DOI] [PubMed] [Google Scholar]
- Doud J.R., Walsh J.M. (1995) Muscle fatigue and muscle length interaction: effect on the EMG frequency components. Electromyography and Clinical Neurophysiology 35, 331-339 [PubMed] [Google Scholar]
- Heinrich D., Mossner M., Kaps P., Nachbauer W. (2009) Calculation of the contact pressure between ski and snow during a carved turn in Alpine skiing. Scandinavian Journal of Medicine and Science in Sports 20(3), 485-492 [DOI] [PubMed] [Google Scholar]
- Hermens H.J., Freriks B., Disselhorst-Klug C., Rau G. (2000) Development of recommendations for SEMG sensors and sensor placement procedures. Journal of Electromyography and Kinesiology 10, 361-374 [DOI] [PubMed] [Google Scholar]
- Hintermeister R.A., O'Connor D.D., Lange G.W., Dillman C.J., Steadman J.R. (1997) Muscle activity in wedge, parallel, and giant slalom skiing. Medicine and Science in Sports and Exercise 29, 548-553 [DOI] [PubMed] [Google Scholar]
- Horita T., Ishiko T. (1987) Relationships between muscle lactate accumulation and surface EMG activities during isokinetic contractions in man. European Journal of Applied Physiology and Occupational Physiology 56, 18-23 [DOI] [PubMed] [Google Scholar]
- Hunter R.E. (1999) Skiing injuries. American Journal of Sports Medicine 27, 381-389 [DOI] [PubMed] [Google Scholar]
- Ishikawa M., Finni T., Komi P.V. (2003) Behaviour of vastus lateralis muscle-tendon during high intensity SSC exercises in vivo. Acta Physiologica Scandinavica 178, 205-213 [DOI] [PubMed] [Google Scholar]
- Karlsson S., Yu J., Akay M. (2000) Time-frequency analysis of myoelectric signals during dynamic contractions: a comparative study. IEEE Transactions on Biomedical Engineering 47, 228-238 [DOI] [PubMed] [Google Scholar]
- Kiernan M.C., Cikurel K., Bostock H. (2001) Effects of temperature on the excitability properties of human motor axons. Brain 124, 816-825 [DOI] [PubMed] [Google Scholar]
- Komi P.V., Tesch P. (1979) EMG frequency spectrum, muscle structure, and fatigue during dynamic contractions in man. European Journal of Applied Physiology and Occupational Physiology 42, 41-50 [DOI] [PubMed] [Google Scholar]
- Kröll J., Wakeling J.M., Seifert J.G., Müller E. (2010) Quadriceps muscle function during recreational alpine skiing. Medicine and Science in Sports and Exercise 42, 1545-1556 [DOI] [PubMed] [Google Scholar]
- Lindstrom L., Magnusson R., Petersen I. (1970) Muscular fatigue and action potential conduction velocity changes studied with frequency analysis of EMG signals. Electromyography 10, 341-356 [PubMed] [Google Scholar]
- Meyers M.C., Laurent C.M., Jr, Higgins R.W., Skelly W.A. (2007) Downhill ski injuries in children and adolescents. Sports Medicine 37, 485-99 [DOI] [PubMed] [Google Scholar]
- Moritani T., Nagata A., Muro M. (1982) Electromyographic manifestations of muscular fatigue. Medicine and Science in Sports and Exercise 14, 198-202 [PubMed] [Google Scholar]
- Mueller E., Schwameder H. (2003) Biomechanical aspects of new techniques in alpine skiing and ski-jumping. Journal of Sports Science 21, 679-692 [DOI] [PubMed] [Google Scholar]
- Nygaard E., Andersen P., Nilsson P., Eriksson E., Kjessel T., Saltin B. (1978) Glycogen depletion pattern and lactate accumulation in leg muscles during recreactional downhill skiing. European Journal of Applied Physiology and Occupational Physiology 38, 261-269 [DOI] [PubMed] [Google Scholar]
- Petrofsky J., Laymon M. (2005) Muscle temperature and EMG amplitude and frequency during isometric exercise. Aviation Space and Environmental Medicine 76, 1024-1030 [PubMed] [Google Scholar]
- Scheiber P., Krautgasser S., von Duvillard S.P., Muller E. (2009) Physiologic responses of older recreational alpine skiers to different skiing modes. European Journal of Applied Physiology and Occupational Physiology 105, 551-558 [DOI] [PubMed] [Google Scholar]
- Seifert J., Kroell J., Mueller E. (2009) The relationship of heart rate and lactate to cumulative muscle fatigue during recreational alpine skiing. Journal of Strength and Conditioning Research 23, 698-704 [DOI] [PubMed] [Google Scholar]
- Smilios I., Hakkinen K., Tokmakidis S.P. (2009) Power output and electromyographic activity during and after a moderate load muscular endurance session. Journal of Strength and Conditioning Research 24, 2122-2131 [DOI] [PubMed] [Google Scholar]
- So R.C., Ng J.K., Lam R.W., Lo C.K., Ng G.Y. (2009) EMG wavelet analysis of quadriceps muscle during repeated knee extension movement. Medicine and Science in Sports and Exercise 41, 788-796 [DOI] [PubMed] [Google Scholar]
- Solomonow M., Baten C., Smit J., Baratta R., Hermens H., D'Ambrosia R., Shoji H. (1990) Electromyogram power spectra frequencies associated with motor unit recruitment strategies. Journal of Applied Physiology 68, 1177-1185 [DOI] [PubMed] [Google Scholar]
- Tesch P., Larsson L., Eriksson A., Karlsson J. (1978) Muscle glycogen depletion and lactate concentration during downhill skiing. Medicine and Science in Sports 10, 85-90 [PubMed] [Google Scholar]
- Tesch P.A. (1995) Aspects on muscle properties and use in competitive Alpine skiing. Medicine and Science in Sports and Exercise 27, 310-314 [PubMed] [Google Scholar]
- Tesch P.A., Komi P.V., Jacobs I., Karlsson J., Viitasalo J.T. (1983) Influence of lactate accumulation of EMG frequency spectrum during repeated concentric contractions. Acta Physiologica Scandinavica 119, 61-67 [DOI] [PubMed] [Google Scholar]
- Turnbull J. R., Kilding A. E., Keogh J. W. (2009) Physiology of alpine skiing. Scandinavian Journal of Medicine and Science in Sports 19, 146-155 [DOI] [PubMed] [Google Scholar]
- Ushiyama Y., Chigira T., Murayama T., Kiryu T. (2005) Muscle fatigue evaluation system applicable while skiing using EMG and knee joint angles. Transactions of the Japanese Society for Medical and Biological Engineering 43, 616-622 [Google Scholar]
- von Tscharner V. (2000) Intensity analysis in time-frequency space of surface myoelectric signals by wavelets of specified resolution. Journal of Electromyography and Kinesiology 10, 433-445 [DOI] [PubMed] [Google Scholar]
- von Tscharner V. (2002) Time-frequency and principal-component methods for the analysis of EMGs recorded during a mildly fatiguing exercise on a cycle ergometer. Journal of Electromyography and Kinesiology 12, 479-492 [DOI] [PubMed] [Google Scholar]
- Wakeling J.M. (2004) Motor units are recruited in a task-dependent fashion during locomotion. Journal of Experimental Biology 207, 3883-3890 [DOI] [PubMed] [Google Scholar]
- Wakeling J.M. (2009) Patterns of motor recruitment can be determined using surface EMG. Journal of Electromyography and Kinesiology 19, 199-207 [DOI] [PubMed] [Google Scholar]
- Wakeling J.M., Pascual S.A., Nigg B.M., von Tscharner V. (2001) Surface EMG shows distinct populations of muscle activity when measured during sustained sub-maximal exercise. European Journal of Applied Physiology and Occupational Physiology 86, 40-47 [DOI] [PubMed] [Google Scholar]
- Wakeling J.M., Rozitis A.I. (2004) Spectral properties of myoelectric signals from different motor units in the leg extensor muscles. Journal of Experimental Biology 207, 2519-2528 [DOI] [PubMed] [Google Scholar]
- Wakeling J.M., Uehli K., Rozitis A.I. (2006) Muscle fibre recruitment can respond to the mechanics of the muscle contraction. Journal of the Royal Society, Interface 3, 533-544 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woerndle W. (2007) Alpiner Skilauf. In: Snowsport Austria - Die österreichische Skischule (engl.: Snowsport Austria - The Austrian Ski School). Walter R.3 rd edition Purkersdorf: Hollinek; 23-128 (In German) [Google Scholar]