

Abstract
Circadian misalignment affects total sleep time, but it may also affect sleep architecture. The objectives of this study were to examine intra-individual effects of circadian misalignment on sleep architecture and inter-individual relationships between sleep stages, cortisol levels and insulin sensitivity. Thirteen subjects (7 men, 6 women, age: 24.3±2.5 y; BMI: 23.6±1.7 kg/m2) stayed in a time blinded respiration chamber during three light-entrained circadian cycles (3x21h and 3x27h) resulting in a phase advance and a phase delay. Sleep was polysomnographically recorded. Blood and salivary samples were collected to determine glucose, insulin and cortisol concentrations. Intra-individually, a phase advance decreased rapid eye movement (REM) sleep and slow-wave sleep (SWS), increased time awake, decreased sleep and REM sleep latency compared to the 24h cycle. A phase delay increased REM sleep, decreased stage 2 sleep, increased time awake, decreased sleep and REM sleep latency compared to the 24h cycle. Moreover, circadian misalignment changed REM sleep distribution with a relatively shorter REM sleep during the second part of the night. Inter-individually, REM sleep was inversely associated with cortisol levels and HOMA-IR index. Circadian misalignment, both a phase advance and a phase delay, significantly changed sleep architecture and resulted in a shift in rem sleep. Inter-individually, shorter REM sleep during the second part of the night was associated with dysregulation of the HPA-axis and reduced insulin sensitivity.
Trial Registration: International Clinical Trials Registry Platform NTR2926 http://apps.who.int/trialsearch/
Introduction
Homeostatic and circadian processes control the quality of wakefulness and sleep. A primary role of the circadian clock is to promote wakefulness during the internal biological day, and to facilitate the consolidation of sleep during the internal biological night [1–3]. Consequently, misalignment between internal circadian time and wakefulness-sleep schedules leads to impaired wakefulness and sleep disturbance. Circadian misalignment may occur during shift work, jet lag or certain circadian rhythm disorders [4,5]. One of the effects of circadian misalignment is a reduction in total sleep time, but circadian misalignment may also affect sleep architecture. Human sleep is not a homogeneous state, but is composed of rapid eye movement (REM) sleep and non-REM sleep. The non-REM sleep can be further divided into 4 stages of progressively deeper sleep. Stages 3 and 4 of non-REM sleep are also named slow-wave sleep (SWS). The preferential distribution of REM sleep toward the latter part of the night is thought to be linked to a circadian oscillator. Contrarily, the preferential distribution of SWS in the beginning of a sleep episode is thought to be mediated by homeostatic processes, i.e. the length of prior wakefulness [6]. Thus, the circadian phase at which sleep occurs may affect the distribution of sleep stages. Therefore, the first aim of our study was to examine the different intra-individual effects of a phase advance and a phase delay on sleep architecture.
Previous studies have suggested that circadian misalignment may lead to adverse metabolic and cardiovascular consequences, which in turn may result in obesity, diabetes and cardiovascular disease [7–9]. In previous work, we showed that circadian misalignment, both a phase advance and a phase delay, resulted in a concomitant disturbance of the glucose-insulin metabolism and substrate oxidation [10], supporting this hypothesis. Additionally, we have shown in a study on sleep and metabolic consequences that inter-individual changes in sleep architecture, rather than total sleep time, are related to endocine and metabolic parameters [11]. The present study aims to establish if the metabolic consequences of circadian misalignment are connected with the effects of circadian misalignment on sleep architecture. Therefore, our second aim is to examine the inter-individual relationships between different sleep stages, cortisol levels as indicator of HPA-axis activity and HOMA-IR index as indicator of insulin sensitivity.
Methods
Ethics Statement
All procedures were carried out with adequate understanding and subjects provided written informed consent at the start of the first test day. The study was conducted according to guidelines laid down in the Declaration of Helsinki and the Medical Ethical Committee of Maastricht University Medical Center approved all procedures involving human subjects. The study was registered, ICTRP registration number is NTR2926. The protocol described here in this study deviates from the trial protocol approved by the Medical Ethical Committee of Maastricht University as it comprises only a part of the approved trial protocol. The protocol for this trial and supporting CONSORT checklist are available as supporting information; see Checklist S1 and Protocol S1.
Subjects
Thirteen healthy subjects (seven men, six women) with a mean age of 24.3 years (SD, 2.5) and with a mean BMI of 23.6 kg/m2 (SD, 1.7) participated in the present study (Figure 1 ). They were recruited via advertisements on notice boards at the Maastricht University. The subjects underwent an initial screening including measurements of body weight and height and completed a questionnaire related to health, use of medication, smoking behavior, alcohol consumption, physical activity, eating behavior and food allergies. All subjects were in good health, non-smokers, not using medication, and at most moderate alcohol consumers. Sleep characteristics were assessed using a questionnaire on habitual sleep duration, time of falling asleep, times woken up during the night, chronotype preference and the Epworth Sleepiness Scale. In general the subjects had no sleeping difficulties as they slept about 8h per night. Time to fall asleep was relatively short and times woken up during the night were low. Subject characteristics are presented in Table 1 .
Figure 1. Flow diagram (CONSORT).
Table 1. Subject characteristics (n=13).
| Characteristic | Value |
|---|---|
| Age (y) | 24.3 ± 2.5 |
| Body weight (kg) | 70.4 ± 8.6 |
| Height (cm) | 172.5 ± 6.8 |
| Body mass index (kg/m2) | 24.3 ± 1.7 |
| Self-reported habitual sleep duration (h/night) | 8.2 ± 1.0 |
| Self-reported time to fall asleep | 16.7 ± 4.6 |
| Self-reported times woken up during the night | 0.6 ± 0.8 |
| Epworth Sleepiness Scale | 4.5 ± 2.5 |
| Chronotype preference (0=morning/1 = evening) | 0.8 ± 0.4 |
All values are means ± SDs.
On the basis of respiration chamber study by Westerterp-Plantenga et al [12], power analysis with the G*Power program (version 3.1; Heinrich- Heine-Universität, Düsseldorf, Germany) showed that with an α = 0.05 and β = 0.10(power = 1 -β = 0.90), ≥11 subjects were needed. The respiration chambers ensure a highly controlled situation in which subjects are in a stable environment.
Study design
The study had a randomized, single-blinded, crossover design. For the 21h and 27h condition, subjects stayed in the respiration chamber during three light-entrained circadian cycles (3 x 21h or 3 x 27h). The 21h condition lasted from 2100h until 1200h three days later. The 27h condition lasted from 2100h until 0600h four days later. In the 21h cycle subjects slept 7 hours and were awake for 14 hours, while in the 27h cycle they slept 9 hours and were awake for 18 hours. Subjects underwent the 21h and 27h conditions time-blinded in random order, separated by at least four weeks. Preceding the 21h and 27h cycles, subjects stayed in the respiration chamber for 24 hours, sleeping for 8 hours and being awake for 16 hours in the chamber (Figure 2 ). Subjects were confined in the chambers all the time. With the exception of partaking in strenuous exercise and sleeping, they were allowed to move freely during the wake phases. Two days before the experiment, subjects were asked to sleep according to their habitual sleep duration (8.2±1.0 hours, Table 1). During the stay in the respiration chamber polysomnography was used to monitor wake and sleep stages. Light entrainment was achieved by using daylight lamps during the waking hours (>400 lux, Energy Saver, Tornado E27, 900 lumen, Philips Lighting, Eindhoven, Netherlands) and black curtains during the sleeping hours. The endogenous melatonin rhythm, which is considered as a reliable marker of circadian timing, was used to confirm circadian misalignment. The maintenance of the 24-h melatonin secretion pattern, independent of the shifts, confirmed misalignment. This has been previously published in [10].
Figure 2. Study design.

The black bars represent the sleep episodes and the white bars represent the wake episodes. Blood sampling times, saliva-sampling times are indicated as respectively B and S. ↓indicates meal times.
Sleep monitoring
To measure wake and sleep stages, polysomnographic recordings were obtained continuously by using BrainRT (OSG BVBA, Rumst, Belgium). Before subjects entered the respiration chamber, surface electrodes for electroencephalogram (EEG), electromyogram (EMG), and electrooculogram (EOG) recording were applied according to standardized criteria [13]. All records were visually scored in 30-s epochs with standardized criteria by the same experienced person blind to the experimental condition [13]. The following sleep parameters were obtained for each subject: total sleep time (TST, consisting of all NREM+REM sleep), wake after sleep onset (WASO, min), sleep period time (defined as TST plus WASO), absolute and relative (% of sleep period time) amount of all sleep stages (Stage 1 sleep, Stage 2 sleep, SWS and REM sleep), sleep efficiency (%, defined as TST divided by time in bed), sleep latency (min, defined as the time to fall asleep) and REM sleep latency (min, defined as the time from sleep onset to the beginning of REM sleep). Daytime naps were not allowed. The sleep parameters were collected for the 24h condition and for each night of the misaligned conditions of 21h and 27h.
Blood and saliva parameters
Every day before breakfast, fasting blood samples were collected for determination of fasting glucose and insulin concentrations (Figure 2). Furthermore, saliva samples were taken before and after each meal to determine cortisol levels (6 samples/day, Figure 2). Glucose, insulin and cortisol concentrations were determined according to a similar protocol used in previous studies conducted at the department of Human Biology, Maastricht University [14–16]. The homeostasis model assessment of insulin resistance (HOMA-IR) was calculated as follows [17]:
HOMA-IR = [fasting insulin (mU/L) x fasting glucose (mmol/L)]/22.5
Energy intake
Two days before the experiment and during their stay in the respiration chamber subjects were fed in energy balance. The energy content of the diet was tailored individually to the energy requirements of each subject based on basic metabolic rate (BMR) calculated with the Harris and Benedict equation [18]. To estimate the total energy requirement at home, BMR was multiplied with a physical activity index (PAL) of 1.75 estimated by means of a computer simulation program [19]. The total energy requirement in the respiration chamber was estimated by multiplying the sleeping metabolic rate (SMR) of the first night with a PAL of 1.35. Energy intake was divided over the meals as 20% for breakfast, 40% for lunch, and 40% for diner. The macronutrient composition of the diet was 12/55/33 En% (protein/carbohydrate/fat). Meals were served at time-points related to cycle duration. Participants were required to finish each meal within half an hour and were not allowed to eat additional food. Water was freely available during the whole experiment.
Statistical Analysis
Data are presented as means ± standard error of the mean (SEM), unless otherwise indicated. The area under the curve (AUC) across the day for cortisol was calculated using the trapezoidal method. ANOVA repeated measures were carried out to compare sleep parameters between the control night (24h cycle) and all nights of the 21h/27h cycle, as well as to compare sleep parameters between different nights of the 21h/27h cycle. Bonferroni corrections for multiple comparisons were applied. Linear regression analyses were completed to analyze the inter-individual relationship between different sleep stages and blood/saliva parameters. All tests were two-sided and differences at P < 0.05 were considered significant. Data were analyzed using SPSS version 18.0 for Macintosh, OS X (SPSS Inc., Chicago, IL, USA).
Results
Effects of sleep curtailment/extension
The control night (24h cycle), the first night of the 21h cycle and the first night of the 27h cycle started at 23h30 and differed only in the duration of sleep opportunity. As a consequence of this design, the effect of sleep duration on sleep architecture, independent of the circadian rhythm, could be determined by comparing the first night of the 21h and 27h cycle to the control night. Subjects slept on average 375.5±8.9min during the 21h cycle and 490.2±7.0min during the 27h cycle compared to 438.8±6.0min during the 24h cycle. Shorter sleep duration, independent of a circadian shift, resulted in a significant decrease of REM sleep, a significant decrease of stage 2 sleep and preservation of SWS. Longer sleep duration, independent of a circadian shift, resulted in a significant increase of REM sleep, stage 2 sleep and SWS (Table 2 ).
Table 2. Absolute (min) and relative (%) duration of sleep parameters during the 21h cycle and the 27h cycle compared to the control night (24h cycle). All values are means ± SEMs (n=13).
| Parameters | 24h night | 21h 1st night | 21h 2nd night | 21h 3rd night | 27h 1st night | 27h 2nd night | 27h 3rd night |
|---|---|---|---|---|---|---|---|
| WASO (min) | 9.6±3.2 | 20.5±8.0 | 11.0±5.1 | 34.3±6.3*, c | 17.0±4.9 | 24.3±7.3 | 18.0±1.9* |
| (%SPT) | 2.1±0.7 | 5.2±2.0 | 2.8±1.3 | 8.6±1.5*, c | 3.4±1.0 | 4.6±1.4 | 3.5±0.4 |
| TST (min) | 438.8±6.0 | 375.5±8.9* | 389.1±6.8* | 365.4±10.2* | 490.2±7.0* | 501.4±8.0* | 502.7±3.7* |
| Stage 1 (min) | 5.0±1.5 | 6.8±2.2 | 6.5±1.9 | 6.5±2.2 | 5.4±1.8 | 7.8±2.4 | 6.2±1.7 |
| (%SPT) | 1.1±0.3 | 1.7±0.6 | 1.7±0.5 | 1.6±0.4 | 1.1±0.4 | 1.5±0.4 | 1.2±0.3 |
| Stage 2 (min) | 214.9±10.5 | 177.2±9.8* | 194.3±6.8* | 174.3±8.3* | 244.8±11.1* | 234.6±7.7 | 220.8±4.4b |
| (%SPT) | 48.0±2.3 | 44.7±2.4 | 48.6±1.7 | 43.4±1.6c | 48.3±2.1 | 44.6±1.3 | 42.6±0.9*, b |
| SWS (min) | 125.7±13.1 | 125.4±13.5 | 108.9±9.2a | 108.2±7.6b | 131.9±14.2* | 125.3±11.6 | 136.8±9.7 |
| (%SPT) | 27.4±2.9 | 31.7±3.5 | 27.2±2.2a | 27.0±1.8b | 25.3±2.8 | 23.9±2.2* | 26.2±1.8 |
| REM sleep (min) | 93.1±8.2 | 66.1±5.4* | 79.3±4.9a | 76.3±4.7 | 108.1±7.6* | 133.7±9.5*, a | 138.9±8.5*, b |
| 1st part of the night | 22.5±5.2 | 19.9±4.1 | 33.0±2.6a | 31.9±6.7 | 32.5±5.6* | 46.2±5.3*, a | 62.2±5.3*, b, c |
| 2nd part of the night | 70.6±7.6 | 46.2±3.7* | 46.3±4.4* | 44.4±4.6* | 75.7±7.1 | 87.5±7.4 | 76.7±5.8 |
| (%SPT) | 20.6±1.7 | 16.7±1.3* | 19.8±1.2a | 19.3±1.4 | 21.2±1.4 | 25.4±1.8*, a | 26.7±1.6*, b |
| Sleep latency (min) | 31.4±5.7 | 23.0±4.0 | 16.9±3.1 | 9.5±1.3*, b, c | 31.4±5.7 | 12.7±2.4*, a | 8.5±2.1*, b, c |
| REM sleep latency (min) | 114.3±15.5 | 110.1±15.5 | 65.0±9.5*, a | 82.2±15.4 | 114.3±15.5 | 83.7±11.8*, a | 93.1±13.8 |
| Sleep efficiency (%) | 91.4±1.2 | 89.4±2.1 | 92.6±1.6 | 87.0±2.4 | 90.8±1.3 | 92.8±1.5 | 93.1±0.7 |
WASO: Wake After Sleep Onset; TST: Total Sleep Time; SWS: slow-wave sleep; REM: rapid eye movement. * Differences to control night. a Within-cycle differences between 1st and 2nd night. b Within-cycle differences between 1st and 3rd night. c Within-cycle differences between 2nd and 3rd night.
Intra-individual effects of shifted sleep on sleep architecture
A phase advance resulted in a significantly increased time awake (WASO) during the third night compared to the second night. Moreover, during the third night of the phase advance sleep latency was significantly decreased compared to the first and second night. Stage 2 sleep stayed significantly decreased during all three nights compared to the control night. SWS decreased significantly during the second night and stayed decreased during the third night compared to the first night. Although REM sleep during the first night was significantly decreased compared to the control night, REM sleep showed a rebound during the second and third night (Table 2).
A phase delay resulted in a significant increased time awake (WASO) during the third night compared to the control night. Moreover, during the third night of the phase delay sleep latency was significantly decreased compared to the first and second night. Stage 2 sleep significantly decreased again during the third night. Although SWS during the first night was significantly increased compared to the control night, there was no significant difference in SWS between the three nights of the 27h cycle. REM sleep further increased significantly during the second and third night (Table 2).
A different role of REM sleep in the first part of the night compared to the latter part of the night has been observed previously [20]. Therefore, we further investigated the distribution of REM sleep over the night in relation to circadian misalignment (Table 2). Subjects’ time in bed was respectively 7, 8 or 9 hours, so REM sleep during the first part of the night is calculated as REM sleep during the first respectively 3.5, 4 or 4.5 hours of time in bed. A phase advance resulted in a significant decrease in REM sleep in the second part of the night (calculated as REM sleep during the last 3.5 hours of time in bed) compared to the control night. The REM rebound in the second night is observed as a significantly increased REM sleep in the first part of the night. Consequently, a phase advance resulted in significantly decreased REM sleep latency.
A phase delay resulted in a significant increase in REM sleep in the first part of the night compared to the control night. The further increase in REM sleep in the second and third night is observed as a significantly increased REM sleep in the first part of the night. Consequently, a phase delay resulted in significantly decreased REM sleep latency. Thus, a phase advance as well as a phase delay resulted in a change in distribution of REM sleep over the night (Table 2).
Inter-individual relationships between sleep architecture, cortisol levels and HOMA-IR index
Possible changes in cortisol concentrations, as an indicator of HPA-axis activity were observed as follows. Linear regression analyses showed that during phase delay cortisol concentrations across the day were inversely related to the amount of REM sleep (R2=0.328, P=0.041). More specifically, the cortisol concentrations in response to lunch were inversely related to the total amount of REM sleep (R2=0.542, P=0.004) and to the amount of REM sleep during the second part of the night (R2=0.334, P=0.021, Figure 3A). This suggests that subjects with less total REM sleep during the second part of the night had higher cortisol concentrations.
Figure 3. Relationship between REM sleep during the 2nd part of the night and cortisol concentrations in response to lunch (nmol/L) during phase delay.
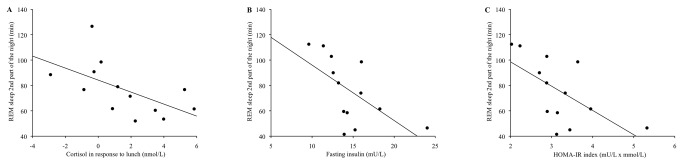
R2=0.334, P<0.05, n=13 (A). Relationship between REM sleep during the 2nd part of the night and fasting insulin concentrations (mU/L) during phase delay. R2=0.374, P<0.05, n=13 (B). Relationship between REM sleep during the 2nd part of the night and HOMA-IR index (mU/L x mmol/L) during phase delay. R2=0.382, P<0.05, n=13 (C).
Possible changes in fasting insulin concentrations and HOMA-IR index, as indicators of insulin sensitivity, were observed as follows. During phase delay, shorter TST was associated with higher fasting insulin levels (R2=0.575, P=0.003) and a higher HOMA-IR index (R2=0.49, P=0.008). In addition, shorter REM sleep was associated with higher fasting insulin levels (R2=0.375, P=0.026) and a higher HOMA-IR index (R2=0.363, P=0.029). Particularly the amount of REM sleep during the second part of the night was inversely related to fasting insulin levels (R2=0.374, P=0.026, Figure 3B) and to the HOMA-IR index (R2=0.382, P=0.024, Figure 3C). This suggests that subjects with less total REM sleep during the second part of the night had higher fasting insulin levels and a higher HOMA-IR index.
In the control and the phase-advanced condition, no relations were found for cortisol, fasting insulin concentrations and HOMA-IR index with sleep parameters.
Discussion
In the present study, the intra-individual effects of circadian misalignment on sleep architecture were examined. Moreover, inter-individual relationships between different sleep stages and cortisol concentrations, as an indicator of HPA-axis activity, and the fasted HOMA-IR index, as an indicator of insulin sensitivity, were investigated.
Intra-individual analyses showed that circadian misalignment, both a phase advance and a phase delay of the sleep period, resulted in disruption of the normal phase distribution between SWS and REM sleep in that REM sleep was relatively phase advanced to SWS and sleep onset. This abnormal circadian sequencing results in shortening of REM sleep latency. This short latency to REM sleep is typical of narcoleptic and depressive patients [21]. In addition to the change in distribution of REM sleep over the night, a phase advance resulted in increased time awake after sleep onset, which is in agreement with the study of Wyatt et al, showing a significant effect of circadian phase on time awake after sleep onset [22]. Consequently, phase advancing the time to go to bed results in diminished sleep continuity. In addition, a phase advance resulted in decreased SWS, which is consistent with the dominant homeostatic modulation of SWS found by Wyatt et al. [22]. During the 21h cycle, subjects are awake for 14h compared to 16h during a 24h day. It may be that during this 14h less homeostatic sleep pressure is built up before sleep, which may explain the reduced amount of SWS during the phase advance. Furthermore, advancing the sleep period phase resulted in a REM sleep rebound since REM sleep was decreased due to decreased time in bed (comparison 7h vs. 8h time in bed). During the second night of the phase advance shift REM sleep latency significantly decreased. This decrease in REM sleep latency may be caused by the REM sleep rebound following REM sleep deprivation [23]. Similarly to the phase advance, the phase delay resulted in increased time awake during the night resulting in decreased sleep continuity. Moreover, phase delaying the time to go to bed significantly increased REM sleep with a decreased REM sleep latency, which is consistent with the pronounced circadian modulation of REM sleep found by Dijk et al. [24]. REM sleep is normally concentrated in the second half of the night due to a circadian disposition for REM sleep to occur at this particular time of the day [25]. Consequently, delaying the phase of full-length sleep period results in a higher percentage of REM sleep compared to advancing the phase of a full-length sleep period.
Inter-individual analyses showed that subjects with relatively less REM sleep, particularly during the second part of the night, showed higher cortisol concentrations and a higher HOMA-IR index. In concordance with a study by Knutson et al. who found that shorter sleep duration measured using wrist actigraphy was associated with higher fasting insulin levels and a higher HOMA-IR index [26], our study indicates a negative correlation between total sleeping time and fasting insulin concentrations and between total sleeping time and the HOMA-IR index. In addition, shorter REM sleep especially during the second part of the night was associated with higher fasting insulin concentrations and higher HOMA-IR index. Koren et al. found positive associations between SWS duration and insulin secretory measures in obese adolescents [27], while in the present study no correlations between SWS and the HOMA-IR index were observed. Koren et al. studied individuals with a mean age of 14.4 years, while in the present study the mean age was 24.3 years, this may explain the discrepancy. Moreover, the discrepancy may be explained by the differences in sleep architecture between obese and non-obese subjects. Obese adolescents show an abnormal sleep pattern with above average SWS but below normal REM sleep percentage [28]. The association between REM sleep and cortisol levels is in accordance with the results described by Van Cauter et al., who observed an inverse relationship between nadir cortisol levels and REM sleep [29]. It is striking that especially REM sleep during the second part of the night was negatively associated with cortisol levels. Wu et al. already showed that sleep at the 03:00-06:00 period, during the circadian nadir, is important in protecting normal physiological rhythms and function of the HPA-axis [30].
The circadian system, however, does not only causes circadian modulation of sleep stages in particular REM sleep, but in parallel it also causes modulation of fasting glucose, insulin and cortisol concentrations [10]. In previous work, phase advancing the sleep period resulted in an increased insulin response to food intake and glucose secretion increased when the sleep period was delayed [10]. Moreover, circadian misalignment caused flattening of the cortisol-secretion pattern [10]. Therefore, the reported inter-individual correlations may also be explained by parallel changes in the circadian phases of REM sleep, fasting insulin and cortisol concentrations, which are all under circadian control.
Finally, the present study showed the effects of sleep duration on sleep architecture. Shortening sleep duration to 7h of time in bed decreased the amount of stage 2 and REM sleep, while SWS was preserved compared to the control night of 8h of sleep. These findings correspond with other studies investigating sleep restriction to only 4h or 5.5h time in bed [31–34]. When the total time in bed is reduced, the sleep system responds by primarily decreasing stage 2 sleep and REM sleep while SWS does not seem to be affected. The preservation of SWS during sleep restriction supports the presumption that SWS is the most restorative part of sleep [25]. With respect to our results on longer sleep duration, the absolute amount of stage 2 sleep, SWS and REM sleep was increased but the percentage of the sleep period spent in these stages remained constant.
Important to note is that all significant findings in this study were observed when considering the absolute values. Despite the difference in time in bed between conditions, it is crucial to compare the absolute duration of sleep stages and not the relative duration; in that case percentages of sleep stages would seem higher as the denominator is reduced sleep time, while absolutely it has not increased at all.
In the present study, subjects are submitted to a sudden advance or delay in their dark-light, rest-activity, sleep-wake cycle, similar to what occurs in jet lag or shift work rotations. Such a protocol allows for the effects of circadian modulation to be observed in the absence of sleep and for the effects of sleep to be observed at an unusual circadian time. In contrast to a forced desynchrony protocol, we were not able to distinguish between the independent effects of the circadian system and behavioral cycles. Possible limitations of the present study are the lack of an adaptation night and that the 24h condition each time took place before the 21h and 27h conditions. However, there were no significant differences in sleep efficiency between different nights diminishing the possible sequence effect (Table 2).
Taken together, intra-individually we found a REM sleep rebound during the phase advance and increased REM sleep during the phase delay moreover REM sleep distribution over the night changed with a relatively shorter REM sleep duration during the second part of the night. Inter-individually we found that shorter REM sleep duration during the second part of the night was correlated with higher cortisol concentrations and a higher HOMA-IR index. This may also explain why not all people are sensitive for circadian misalignment and its negative metabolic consequences.
In conclusion, circadian misalignment, both a phase advance and a phase delay, resulted in a significant change in sleep architecture, especially a shift in REM sleep. Inter-individually, shorter REM sleep during the second part of the night was associated with dysregulation of the HPA-axis, as indicated by increased cortisol concentrations, and reduced insulin sensitivity. Dysregulation of the HPA-axis and insulin resistance are hallmarks of several metabolic diseases such as type-2 diabetes and obesity, which are known to be associated with circadian misalignment.
Supporting Information
Trial Protocol.
(PDF)
CONSORT Checklist.
(DOC)
Funding Statement
The authors have no funding or support to report.
References
- 1. Czeisler CA, Weitzman Ed, Moore-Ede MC, Zimmerman JC, Knauer RS (1980) Human sleep: its duration and organization depend on its circadian phase. Science 210: 1264-1267 doi:10.1126/science.7434029. PubMed: 7434029. [DOI] [PubMed]
- 2. Dijk DJ, Czeisler CA (1994) Paradoxical timing of the circadian rhythm of sleep propensity serves to consolidate sleep and wakefulness in humans. Neurosci Lett 166: 63-68. doi:10.1016/0304-3940(94)90841-9. PubMed: 8190360. [DOI] [PubMed] [Google Scholar]
- 3. Morris CJ, Aeschbach D, Scheer FA (2012) Circadian system, sleep and endocrinology. Mol Cell Endocrinol 349(1): 91-104. doi:10.1016/j.mce.2011.09.003. PubMed: 21939733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Akerstedt T, Wright KP Jr. (2009) Sleep Loss and Fatigue in Shift Work and Shift Work Disorder. Sleep Med Clin 4: 257-271. doi:10.1016/j.jsmc.2009.03.001. PubMed: 20640236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Reid KJ, Zee PC (2009) Circadian rhythm disorders. Semin Neurol 29: 393-405. doi:10.1055/s-0029-1237120. PubMed: 19742414. [DOI] [PubMed] [Google Scholar]
- 6. Carskadon MA, Dement WC (2011) Monitoring and staging human sleep. Principles and practice of sleep medicine. 5th edition ed. St. Louis: Elsevier Saunders; pp. 16-26. [Google Scholar]
- 7. Roenneberg T, Allebrandt KV, Merrow M, Vetter C (2012) Social jetlag and obesity. Curr Biol 22: 939-943. doi:10.1016/j.cub.2012.03.038. PubMed: 22578422. [DOI] [PubMed] [Google Scholar]
- 8. Hampton SM, Morgan LM, Lawrence N, Anastasiadou T, Norris F et al. (1996) Postprandial hormone and metabolic responses in simulated shift work. J Endocrinol 151: 259-267. doi:10.1677/joe.0.1510259. PubMed: 8958786. [DOI] [PubMed] [Google Scholar]
- 9. Scheer FA, Hilton MF, Mantzoros CS, Shea SA (2009) Adverse metabolic and cardiovascular consequences of circadian misalignment. Proc Natl Acad Sci U S A 106: 4453-4458. doi:10.1073/pnas.0808180106. PubMed: 19255424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Gonnissen HK, Rutters F, Mazuy C, Martens EA, Adam TC et al. (2012) Effect of a phase advance and phase delay of the 24-h cycle on energy metabolism, appetite, and related hormones. Am J Clin Nutr 96: 689-697. doi:10.3945/ajcn.112.037192. PubMed: 22914550. [DOI] [PubMed] [Google Scholar]
- 11. Rutters F, Gonnissen HK, Hursel R, Lemmens SG, Martens EA et al. (2012) Distinct associations between energy balance and the sleep characteristics slow wave sleep and rapid eye movement sleep. Int J Obes (Lond) 36: 1346-1352. doi:10.1038/ijo.2011.250. PubMed: 22234280. [DOI] [PubMed] [Google Scholar]
- 12. Westerterp-Plantenga MS, Lejeune MP, Smeets AJ, Luscombe-Marsh ND (2009) Sex differences in energy homeostatis following a diet relatively high in protein exchanged with carbohydrate, assessed in a respiration chamber in humans. Physiol Behav 97: 414-419. doi:10.1016/j.physbeh.2009.03.010. PubMed: 19318111. [DOI] [PubMed] [Google Scholar]
- 13. Rechtschaffen A, Kales A (1968) A Manual of Standardized Terminology, Techniques and Scoring System for Sleep Stages of Human Subjects. Bethesda, MD: US National Institute of Neurological Diseases and Blindness. [Google Scholar]
- 14. Lemmens SG, Born JM, Martens EA, Martens MJ, Westerterp-Plantenga MS (2011) Influence of consumption of a high-protein vs. high-carbohydrate meal on the physiological cortisol and psychological mood response in men and women. PLOS ONE 6: e16826. doi:10.1371/journal.pone.0016826. PubMed: 21304815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Veldhorst MA, Nieuwenhuizen AG, Hochstenbach-Waelen A, van Vught AJ, Westerterp KR et al. (2009) Dose-dependent satiating effect of whey relative to casein or soy. Physiol Behav 96: 675-682. doi:10.1016/j.physbeh.2009.01.004. PubMed: 19385022. [DOI] [PubMed] [Google Scholar]
- 16. Lemmens SG, Martens EA, Kester AD, Westerterp-Plantenga MS (2011) Changes in gut hormone and glucose concentrations in relation to hunger and fullness. Am J Clin Nutr 94: 717-725. doi:10.3945/ajcn.110.008631. PubMed: 21795439. [DOI] [PubMed] [Google Scholar]
- 17. Adam TC, Hasson RE, Lane CJ, Davis JN, Weigensberg MJ et al. (2011) Fasting indicators of insulin sensitivity: effects of ethnicity and pubertal status. Diabetes Care 34: 994-999. doi:10.2337/dc10-1593. PubMed: 21357795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Harris JA, Benedict FG (1918) A Biometric Study of Human Basal Metabolism. Proc Natl Acad Sci U S A 4: 370-373. doi:10.1073/pnas.4.12.370. PubMed: 16576330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Westerterp KR, Donkers JH, Fredrix EW, Boekhoudt P (1995) Energy intake, physical activity and body weight: a simulation model. Br J Nutr 73: 337-347. doi:10.1079/BJN19950037. PubMed: 7766558. [DOI] [PubMed] [Google Scholar]
- 20. Horne J (2009) REM sleep, energy balance and 'optimal foraging'. Neurosci Biobehav Rev 33: 466-474. doi:10.1016/j.neubiorev.2008.12.002. PubMed: 19133292. [DOI] [PubMed] [Google Scholar]
- 21. Lee ML, Swanson BE, de la Iglesia HO (2009) Circadian timing of REM sleep is coupled to an oscillator within the dorsomedial suprachiasmatic nucleus. Curr Biol 19: 848-852. doi:10.1016/j.cub.2009.03.051. PubMed: 19375313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Wyatt JK, Ritz-De Cecco A, Czeisler CA, Dijk DJ (1999) Circadian temperature and melatonin rhythms, sleep, and neurobehavioral function in humans living on a 20-h day. Am J Physiol 277: R1152-R1163. PubMed: 10516257. [DOI] [PubMed] [Google Scholar]
- 23. Brass SD, Auerbach S (2009) A sleepy patient with REM rebound. J Clin Sleep Med 5: 386-389. PubMed: 19968020. [PMC free article] [PubMed] [Google Scholar]
- 24. Dijk DJ, Czeisler CA (1995) Contribution of the circadian pacemaker and the sleep homeostat to sleep propensity, sleep structure, electroencephalographic slow waves, and sleep spindle activity in humans. J Neurosci 15: 3526-3538. PubMed: 7751928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Tilley AJ, Wilkinson RT (1984) The effects of a restricted sleep regime on the composition of sleep and on performance. Psychophysiology 21: 406-412. doi:10.1111/j.1469-8986.1984.tb00217.x. PubMed: 6463172. [DOI] [PubMed] [Google Scholar]
- 26. Knutson KL, Van Cauter E, Zee P, Liu K, Lauderdale DS (2011) Cross-sectional associations between measures of sleep and markers of glucose metabolism among subjects with and without diabetes: the Coronary Artery Risk Development in Young Adults (CARDIA) Sleep Study. Diabetes Care 34: 1171-1176. doi:10.2337/dc10-1962. PubMed: 21411507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Koren D, Levitt Katz LE, Brar PC, Gallagher PR, Berkowitz RI et al. (2011) Sleep architecture and glucose and insulin homeostasis in obese adolescents. Diabetes Care 34: 2442-2447. doi:10.2337/dc11-1093. PubMed: 21933909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Willi SM, Oexmann MJ, Wright NM, Collop NA, Key LL Jr. (1998) The effects of a high-protein, low-fat, ketogenic diet on adolescents with morbid obesity: body composition, blood chemistries, and sleep abnormalities. Pediatrics 101: 61-67. doi:10.1542/peds.101.1.61. PubMed: 9417152. [DOI] [PubMed] [Google Scholar]
- 29. Van Cauter E, Leproult R, Plat L (2000) Age-related changes in slow wave sleep and REM sleep and relationship with growth hormone and cortisol levels in healthy men. JAMA 284: 861-868. doi:10.1001/jama.284.7.861. PubMed: 10938176. [DOI] [PubMed] [Google Scholar]
- 30. Wu H, Stone WS, Hsi X, Zhuang J, Huang L et al. (2010) Effects of different sleep restriction protocols on sleep architecture and daytime vigilance in healthy men. Physiol Res 59: 821-829. PubMed: 20406032. [DOI] [PubMed] [Google Scholar]
- 31. Akerstedt T, Gillberg M (1986) A dose-response study of sleep loss and spontaneous sleep termination. Psychophysiology 23: 293-297. doi:10.1111/j.1469-8986.1986.tb00635.x. PubMed: 3749409. [DOI] [PubMed] [Google Scholar]
- 32. Brunner DP, Dijk DJ, Borbély AA (1993) Repeated partial sleep deprivation progressively changes in EEG during sleep and wakefulness. Sleep 16: 100-113. PubMed: 8446828. [DOI] [PubMed] [Google Scholar]
- 33. Nedeltcheva AV, Kilkus JM, Imperial J, Kasza K, Schoeller DA et al. (2009) Sleep curtailment is accompanied by increased intake of calories from snacks. Am J Clin Nutr 89: 126-133. PubMed: 19056602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Nedeltcheva AV, Kilkus JM, Imperial J, Schoeller DA, Penev PD (2010) Insufficient sleep undermines dietary efforts to reduce adiposity. Ann Intern Med 153: 435-441. doi:10.7326/0003-4819-153-7-201010050-00006. PubMed: 20921542. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Trial Protocol.
(PDF)
CONSORT Checklist.
(DOC)