

Abstract
This Event-Related Potential (ERP) study investigated whether components commonly measured at test, such as the FN400 and the parietal old/new components, could be observed during encoding and, if so, whether they would predict different levels of accuracy on a subsequent memory test. ERPs were recorded while subjects classified pictures of objects as man-made or natural. Some objects were only classified once while others were classified twice during encoding, sometimes with an identical picture, and other times with a different exemplar from the same category. A subsequent surprise recognition test required subjects to judge whether each probe word corresponded to a picture shown earlier, and if so whether there were two identical pictures that corresponded to the word probe, two different pictures, or just one picture. When the second presentation showed a duplicate of an earlier picture, the FN400 effect (a significantly less negative deflection on the second presentation) was observed regardless of subsequent memory response; however, when the second presentation showed a different exemplar of the same concept, the FN400 effect was only marginally significant. In contrast, the parietal old/new effect was robust for the second presentation of conceptual repetitions when the test probe was subsequently recognized, but not for identical repetitions. These findings suggest that ERP components that are typically observed during an episodic memory test can be observed during an incidental encoding task, and that they are predictive of the degree of subsequent memory performance.
Keywords: ERP, Event-Related Potential, FN400, LPC, Parietal Old/New, Dm effect, Subsequent Memory
1. Introduction
Event-Related Potential (ERP) studies of recognition memory at test have revealed two common components: the FN400 and the parietal old/new component (also called the Late Positive Component or LPC). The parietal old/new effect is a late positive deflection over parietal sites often studied in the context of recognition memory, and has been tied to recollection (e.g., Duarte et al., 2004; Rugg and Curran 2007; Voss, Lucas and Paller, 2010; Woodruff et al., 2006; see Yonelinas, 2002 for a review). Evidence for the parietal old/new effect as an index of recollection has primarily come from investigations of source memory. For instance, in listening tasks with male and female speakers, memory can be tested both for whether an item is old and whether the speaker’s gender is correctly identified; the parietal old/new effect is more positive for items where the speaker is correctly identified than for items judged old but with the incorrect source. (Wilding and Rugg, 1996; Senkfor and Van Petten, 1998). Likewise, when subjects are administered a drug that interferes with source memory more than familiarity, such as midazolam, the parietal old/new effect disappears for old items but the FN400 is not affected (Curran et al., 2006).
While there is considerable agreement that the parietal old/new effect is associated with recollection, there is less agreement concerning the interpretation of the FN400. The FN400 is a negative deflection over the frontal scalp that is more negative for items not seen before, and several studies have found evidence that suggest the FN400 is an index of familiarity used in episodic recognition (e.g., Curran, 2000; Curran and Cleary, 2003; Curran and Doyle, 2011; Duarte et al., 2004 Düzel et al., 1997; Ecker et al., 2007; Groh-Bordin et al., 2006; Rugg and Curran, 2007; Smith, 1993). Early evidence for this interpretation was found in studies of ‘lures’ in an old/new task. In these studies, the FN400 appeared for recognized old words and for false alarms for unstudied words that were similar to old words, such as words that differed only in their plurality from studied items Curran (2000). Studies of shallow versus deep encoding provide some converging evidence: while recollection is thought to suffer in the absence of deeper (semantic) encoding, familiarity tends to be relatively unaffected by “depth of encoding” (see Yonelinas, 2002 for a review). If the FN400 does measure familiarity, one should expect it to appear for items in both shallow and deep tasks while a component tied to recollection, like the parietal old/new, should be stronger following deep encoding. This is exactly what Rugg and colleagues (1998) found. Both this finding and the word plurality finding suggest that the FN400 is an index of familiarity ‘strength’.
On the other hand, Paller and colleagues (e.g., Yovel and Paller, 2004; Voss and Paller 2009) have disputed the notion that the FN400 is a ‘pure’ measure of familiarity. Rather, they propose that the FN400 is an index of conceptual priming. Evidence for this has come from studies that attempt to isolate familiarity from conceptual priming by using meaningless stimuli (e,g., kaleidoscope images). For these conceptually impoverished stimuli, judgments based on familiarity may not always lead to the typical FN400 effect (Paller, Boehm and Voss. 2007; Voss and Paller, 2009; Voss, Lucas and Paller, 2010; but see Groh-Bordin et al., 2006 for a counterexample). Voss and Paller, for instance, had subjects study kaleidoscope images, and then make a modified remember/know judgment. High Confidence ‘know’ responses, despite accuracy well above chance, did not elicit an FN400 significantly different from new items (Voss and Paller, 2009). By contrast, ‘know’ responses to more meaningful stimuli reliably show FN400s: (Woodruff et al, 2006).
The counter argument to FN400s being an index of conceptual priming has involved finding situations where conceptual priming should remain the same across conditions, but still leads to differing FN400 amplitudes. Manipulations of color is one such area: Groh-Bordin et al (2006) had drawings of objects in different colors at study, and then either held color constant or changed it at test; changing color led to reliably different FN400s. Similarly, Curran and Doyle (2011) found FN400 differences when manipulating modality from study to test. In their experiment, participants studied either words or pictures, and the representation of the concept could either match (word-word, picture-picture) or mismatch for the recognition test. FN400 differences were reliably smaller for switched modality, and this was not due to memory performance, as studied pictures had higher accuracy regardless of match or mismatch.
Despite this debate, most would agree that there exist two topographically and chronologically distinct ERP components, the FN400 and the parietal old/new effect, which index different processes observed at retrieval. However, no such distinction has been made during the encoding phases.
ERP studies that do focus on encoding have found an increased positivity, usually starting between 400 and 500 ms, for subsequently remembered items compared to items that were forgotten, often called ‘Dm’ (Paller et al., 1987; Yovel and Paller, 2004; Duarte et al., 2004; Friedman and Johnson, 2000). Paller coined the term Dm, for ‘differences due to subsequent memory’ to describe this posterior positivity. Dm studies are one of the few paradigms that allow incidental encoding to be partitioned based on memory strength, and therefore can show distinct effects for stronger versus weaker items at encoding. However, most Dm studies involve only a single presentation of an item, and the classic Dm effect has rarely shown distinctions past that of ‘remembered’ versus ‘forgotten’.
Conversely, to our knowledge, encoding studies of repetition priming have not attempted to partition their trials by Dm analyses. Instead, much of this literature has focused on dissociations between implicit and explicit memory. In particular, studies that have contrasted ‘explicit’ and ‘implicit’ memory effects have not equated other factors, making it difficult to interpret the findings. For example, while analyses of explicit memory tasks invariably separate hit versus miss trials, or in some paradigms, Remember versus Know versus Miss trials, analyses of implicit memory tasks lump together both well-encoded and poorly encoded trials. In our view, implicit trials should also be partitioned based on memory strength. Of course, it is not possible to ask for memory judgments at the time of the implicit task, as that would make the task explicit. However, a Dm analysis should be a way to finesse this problem. The current ERP study involves up to two item presentations during encoding followed by a later explicit recognition test, thereby enabling back-sorting of encoding trials by subsequent memory. With such a design, one can examine encoding while controlling for the strength of the memory.
The encoding task involves presenting pictures of common objects one at a time that subjects must categorize as natural or man-made. Over the course of this categorization task, some of the repetitions involve the identical picture of an object, some involve showing a different exemplar (different picture) of the same object category while, for some objects, only one picture is presented.
After all pictures have been judged as man-made or natural, subjects are given a surprise recognition memory test in which they are presented with words that correspond to previously seen and unseen object categories. Subjects are asked to indicate whether they can recall seeing a picture that corresponds to the test word and, if so, whether they recall seeing two identical, two different, or just one picture. Their responses to the surprise memory test allow us to separately “bin” each encoding trial (first and second presentation, if there were two) based on its later memory status. This binning allows us to analyze ERP components as a function of the type of repetition (identical, conceptual or single), judgment accuracy and type of error. Critically, it also gives us a way to partition incidental effects by memory strength.
Previous work by Ecker and colleagues (Ecker and Zimmer 2009; Küper et al., 2012) used a similar paradigm to ours; they also presented subjects with identical repetitions or different exemplars. Critically, however, their effects were examined during an explicit memory test. In contrast, the current study examines the memory processes evoked during an incidental task (judging natural versus man-made on the second presentation). The advantage of the current study is that it allows us to ask whether the ERP memory components traditionally identified during an explicit memory test will only be observed when a subject engages in an explicit memory decision (and associated motor response) or whether these ERP memory components will also occur when a person is engaged in a task that does not require explicit memory. If the primary factors are memory strength and tacit recollection, then back-sorting by response at test should reveal these effects even in an implicit task.
Another benefit of our paradigm is that it provides a potential test of the familiarity versus conceptual priming debate for the FN400. Identical repetitions and exemplars from the same category should lead to roughly equivalent levels of conceptual priming but differing levels of familiarity. Thus, if different exemplars lead to reduced FN400 differences compared to identical repetitions, this would argue against conceptual priming as the sole factor mediating the FN400.
In summary, the questions we pursue are (1) will we find evidence that ERP components typically associated with explicit memory tasks can be observed during an incidental memory task, provided that we are able to back-sort the encoding trials as a function of subsequent memory? (2) If we do find support for (1), will the FN400 be stronger for identical repetitions than conceptual repetitions? Finally, (3) for those encoding pairs that are accurately remembered later, will we observe the parietal old/new effect for both conceptual and identical repetitions, neither of them, or just one type of repetition? Given that the parietal old/new is thought to represent semantic processing, we predict that we should find the effect for both types of repetition.
2. Results
The behavioral accuracy results focus primarily on performance on the final recognition test although we also examine behavioral priming effects (RT differences) during encoding. The data from the memory test are used primarily to allow us to partition the ERP encoding data based on subsequent memory. These encoding trials are categorized on several dimensions: (a) whether the trial was the first or second presentation of a pair; (b) whether the two presentations involved identical pictures or pictures of visually distinct exemplars from the same category; (c) whether the item was a single presentation, and if so, whether it was shown in the first or second half of the experiment. The single presentations allowed subjects to respond single if they did not remember both presentations but remembered seeing a picture that corresponded to the test probe.
These encoding trials were further classified based on their performance during the subsequent memory test. For example, if a particular item was shown only a single time during encoding and the subject responded that this item was shown only a single time at test, then the complete categorization of the corresponding encoding trial would be “Single, said Single.” If there were two different pictures of a shoe and the subject said that there were two different pictures that correct response would lead to the encoding trials being coded as “Different, said Different.” If the subject thought that there were two pictures of the same shoe but that they were actually different, the encoding trials would be scored as “Different, said Same.”
2.1 Behavioral Analyses
Figure 1 shows the distribution of test responses for each repetition type. The percent correct is the proportion of responses that match the correct answer. The figure also provides the distribution of erroneous responses for each condition. In general, subjects are highly accurate at selecting the correct answer for a given type of test probe. For example, subjects are far more likely to say “new” than anything else when no picture corresponded to a test probe; they are far more likely to respond “single” when there was only one picture that corresponded to the probe, and far more likely to respond “same” than any other respond when there were two identical pictures shown. The only condition that gave as many erroneous responses as correct responses was the “different” condition. Even in this condition, though, subjects were generally correct in that some form of the object had appeared twice at encoding.
Fig. 1.
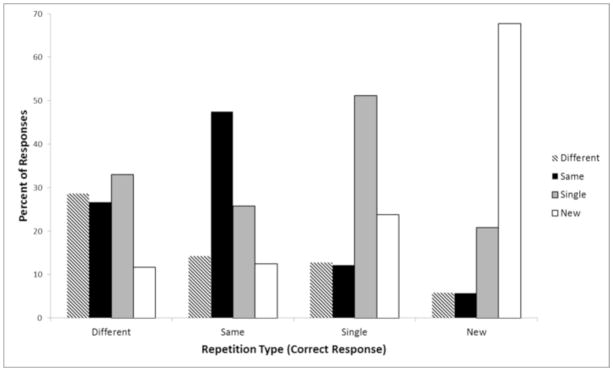
Proportions of correct and erroneous responses to memory probes as a function of correct category of response.
Although it appears that the different condition is less accurate than the same condition, they only differ in response bias, not memory sensitivity. Subjects were far more likely to respond “same” when the correct answer was different than they were to respond “different” when the correct answer was same. To quantify potential differences in memory sensitivity and response bias, we calculated d′ and β for each subject for both types of pairs (Green and Swets, 1966). For the same repetition condition, a hit was defined as a trial where the subject responded “same” when the pictures were identical and a false alarm was responding “same” when the two pictures were different. For the different repetition condition, a hit was defined as a trial where the subject responded “different” when the two pictures were different, and a false alarm was defined as saying “different” when the two pictures were identical. The two conditions did not show reliable differences in discriminability, d′=.59 for different exemplars and .51 for identical repetitions, t=1.43 p > .1. On the other hand, there was a reliable difference in response bias, β= 2.12 for different exemplars and β=1.24 for identical repetitions, t=2.33, p < .05. As larger values of β reflect a more conservative response, this result suggests that subjects were simply much less likely to respond “different” when they were unsure than they were to respond “same” when they were unsure.1
Figure 2 plots mean response time (RT) data during encoding for correct classifications (natural, man-made) as a function of first versus second presentation. For single items, the mean RTs present are those from the first versus the second half of the encoding phase. By comparing RTs for “single” trials from the first half versus the second half of the encoding task, we have an estimate of the facilitation due to practice at the task. To quantify to what extent priming effects should be attributed to practice at the task, we used an ANOVA with presentation (half) (first versus second) and condition (same, different, single) as factors.
Fig. 2.
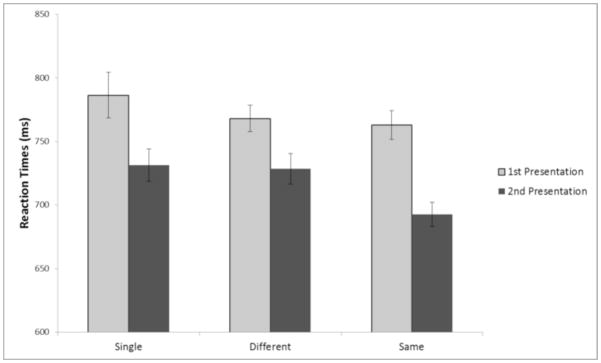
Mean correct reaction times during encoding for the first and second presentations of a “pair”, partitioned by type of “repetition.” Error Bars are ±1 standard error.
There was an expected main effect of first versus second half of the encoding task, such that RTs were reliably faster during the second half of the task, F(1,22) = 14.3, p < .01. There was also a main effect of condition (single, same, different), F(2,44) =10.5, p < .01, as well as a significant condition × presentation half (1st versus 2nd) interaction F(2,44) =4.7, p < .05. The only contrast that revealed a speed up significantly different from the single comparison was the identical condition F(1,22) =7.4, p < .05.2
2.2 ERP Analyses
All ERP analyses were done on the encoding trials recoded as described above in the behavioral results. Because the number of observations for each recoded group was limited by the response of the subject at test, some of the recoded categories had fewer observations per subject than others. As a consequence, we focused on the three categories of repetitions that have at least 15 observations/subject: the correct responses for different exemplars and identical repetitions, and different exemplars that were judged as the same.
In order to analyze the FN400 and parietal old/new ERP components, we focused on two clusters of electrodes, a frontal cluster (F3, Fz, and F4) and a parietal cluster (P3, Pz, P4), and two time windows, 300–450 ms for the FN400, and 450–700 ms for the parietal old/new effect; similar clusters of electrodes and times windows have been shown to elicit these components when recorded at test (e.g., Curran and Cleary, 2003; MacKenzie and Donaldson, 2007; Voss and Paller, 2009; Yovel and Paller 2004).3
Figure 3 shows topographic maps, based on all recorded electrodes, of the subsequent memory effects sorted by response-type. Amplitudes were compared at both time windows using separate repeated measures ANOVAs. Factors for the 2×2 ANOVAs were Presentation (first versus second) and electrode cluster (F3, Fz, F4 versus P3, Pz, P4), unless stated otherwise. For each effect examined, mean voltage amplitude was used as the dependent measure.
Fig. 3.
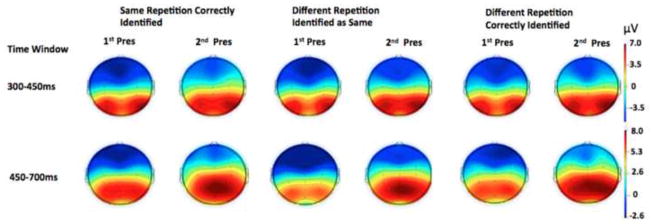
Head Plots illustrating the FN400 component (time window 300 – 450 ms) and the parietal old/new component (time window 450 – 700 ms) for first and second presentations as a function of type of repetition and subsequent memory response.
At the earlier time window, correctly judged identical repetitions showed an FN400, with a main effect of presentation (first versus second) F(1, 22) = 7.93, p < .05. For different exemplars correctly judged as “different”, however, the effect of presentation was only marginal F(1, 22) = 3.39, p < .08. Different exemplars judged as “same” did not show a reliable FN400 effect, p>.1, (see Figure 4). Figure 5 shows the averaged waveforms at the F3, Fz, and F4 electrodes.
Fig. 4.
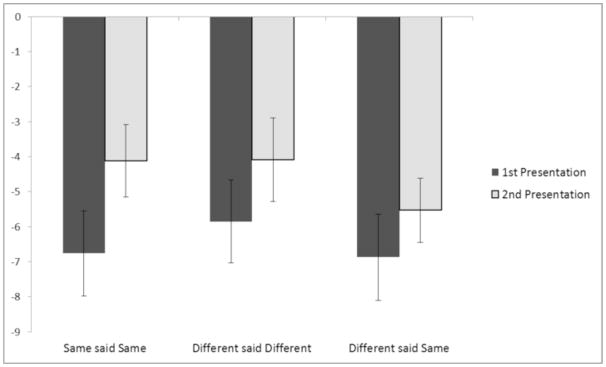
Mean amplitude in μV of the FN400 component using the F3, Fz, F4 cluster (time window 300 – 450 ms) at encoding, as a function of type of repetition and type of response. Error Bars are ±1 standard error.
Fig. 5.
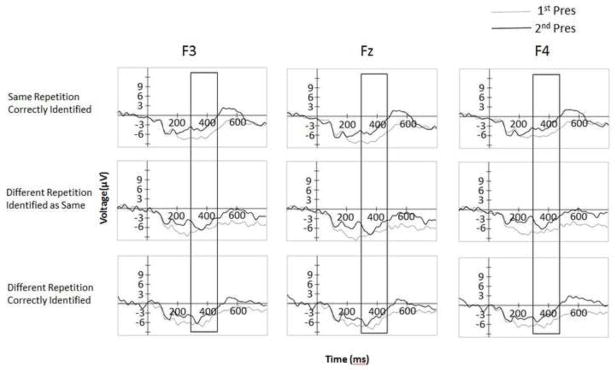
Stimulus-locked waveforms in μV at F3, Fz, and F4 electrodes for the first and second presentation as a function of type of repetition and type of response.
In contrast to the FN400, at the later time window there were parietal old/new components for both types of different exemplars analyzed (see Figure 6), with main effects of presentation for both different exemplars correctly judged “different” F(1, 22) = 5.97, p < .05, and for different exemplars judged as “same” F(1, 22) = 7.77, p < .01. A direct comparison between these response categories indicates that different exemplars that were later erroneously judged as identical presentations were more negative overall F(1, 22) = 4.43, p < .05 (Figure 6). Identical repetitions correctly judged “same” showed a marginal effect of presentation, F(1, 22) = 3.98, p = .058; however, this was qualified by an interaction with electrode site F(1, 22) = 9.49, p < .01, such that these differences appeared to be driven more by the frontal than the parietal cluster. Follow-up tests confirmed that at the parietal cluster alone this effect does not reach significance (p>.1) although numerically it was in the right direction4, while the frontal cluster showed a reliable difference between first and second presentation, F(1, 22) = 9.28, p < .01. Figure 7 shows the averaged waveforms at the P3, Pz, and P4 electrodes.
Fig. 6.
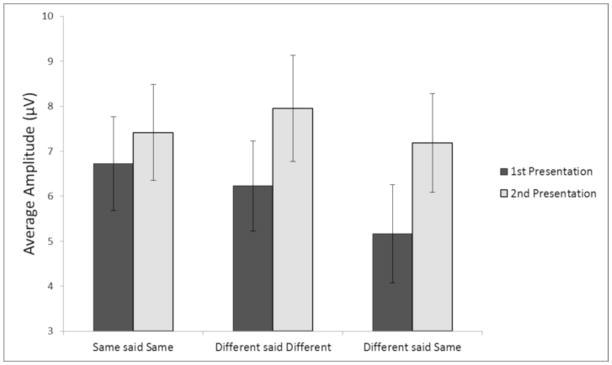
Mean amplitude in μV of the parietal old/new component using the P3, Pz, P4 cluster (time window 450 – 700 ms) at encoding, as a function of type of repetition and type of response. Error Bars are ±1 standard error.
Fig. 7.
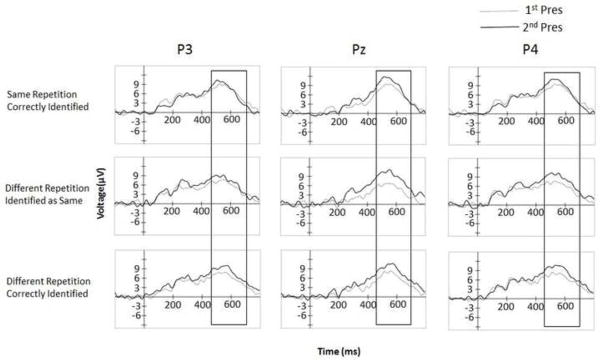
Stimulus-locked waveforms in μV at P3, Pz, and P4 electrodes for the first and second presentation at encoding, as a function of type of repetition and type of response.
2.3 Analyses of ERP Results without Back-Sorting
To test whether these effects were in fact due to differences in subsequent memory, the same analyses were conducted on all encoding trials regardless of the accuracy of the memory test response. For the FN400, the observed pattern matched that found when encoding trials were back-sorted by subsequent memory. When comparing first versus second presentation without partitioning based on subsequent memory, only identical repetitions showed a reliable FN400 effect, F(1, 22) = 14.89, p < .01. Different exemplars again showed only a marginal effect when not back-sorted, F(1, 22) = 3.2, p < .09.
For the parietal old/new effect, when there was no back-sorting based on subsequent memory, both identical and different exemplars showed effects of repetition, F(1, 22) = 9.43, p < .01; F(1, 22) = 6.58, p < .05, respectively, but these were qualified by interactions with electrode site, such that these differences were greater at the frontal cluster than at the parietal cluster. Follow up tests for identical repetitions and different exemplars at only the parietal cluster showed no reliable effects, all ps > .1).
3. Discussion
Our results indicate that the FN400 and the parietal old/new effects are observed not only during explicit memory tasks, as previous research has noted, but also during performance on an incidental task that does not require an explicit memory judgment. When memory strength is controlled for by back-sorting these incidental encoding trials based on subsequent memory, we found that both ERP components showed robust effects. Furthermore, the different patterns of the parietal old/new effect were found to predict the accuracy of the response for different types of stimulus pairs. Despite the incidental nature of the task, neither the FN400 nor the parietal old/new effect showed a pattern that is distinguishable, in terms of time window, size of effect, or scalp localization, from its corresponding component seen during a typical recognition experiment.
Past research has suggested that the parietal old/new is less strong (or absent) for implicit tasks (Rugg et al. 1998; Küper et al., 2012). We think the reason for the discrepancy from our findings involves the lack of back-sorting based on subsequent memory. In the Rugg et al. (1998) study, after the initial encoding that involved using a word in a sentence, subjects made one of two types of judgments for each item previously encoded: either a recognition test or an animacy categorization task (similar to our encoding task). Rugg et al. found that, while there were FN400 and parietal old/new effects during the old/new test for the recognition group, these effects were not evident in the data from the animacy categorization task. Instead, the categorization data showed an ERP pattern similar to that found for unrecognized words.
Küper et al. (2012) used a design more similar to ours. They contrasted two ‘sessions’ of an animacy task with an animacy task followed by an old/new test. They also compared identical versus ‘different’ pictures, and unlike Rugg et al. (1998), did show marginally increased parietal activation from 500–700 in the implicit condition for identical repetitions. However, they found no significant effects for ‘different’ exemplars.
As alluded to above, the greatest difference between our study and those of Rugg et al. (1998) and Küper et al. (2012) is that we included a final memory test that enabled back-sorting as in the Dm studies. Because the other two studies did not back-sort the encoding trials, their repeated items involved a mixture of well-encoded items and those encoded more weakly, which would obscure potential memory effects. In the present study, for instance, the parietal old/new effects we found largely disappeared when encoding trials were analyzed without back-sorting.
In our view it is not surprising that these parietal old/new effects typically seen in an explicit memory test would occur in an incidental task when the trials that are analyzed (based on back-sorting) exclude explicit memory failures. This levels the playing field, so to speak. The question then becomes whether these observed effects derive from an implicit memory system or an explicit memory system; our answer is that the proposed distinction of implicit and explicit memory systems is not valid (see Reder et al., 2009 for a comprehensive review). Instead, we believe that implicit memory and explicit memory should refer to tasks, not memory systems. Given that both tasks tap the same memory representations, then when the strength of a memory trace is equated as it was in our study it is not surprising that the ERP patterns are so similar.
The current findings also shed some light on the debate concerning the functional significance of the FN400. As previously discussed, there has been disagreement about the extent of perceptual versus conceptual influences on the FN400 effect. The current results provide evidence that perceptual influences matter for the FN400 effect: at encoding, the FN400 was weaker for conceptual repetitions compared to perceptually identical repetitions. However, while perceptual influences seem to matter given conceptual priming, Paller’s initial claim was more specific: that, in the absence of conceptual priming, the typical FN400 would not occur. Paller’s “absence of conceptual priming” seems to refer to situations where there should be no existing representation in semantic (long term) memory for the stimuli (e.g., novel kaleidoscope images) and thus no representation to prime. While we agree with that assessment, the current study was not designed to test that meaning of conceptual priming. Our goal was to see whether both the FN400 and parietal old/new effects could appear in a repetition priming task. Thus our design intended to reinstate the concept, as this would be necessary for recollection to occur for different exemplars.
For FN400 effects in the absence of conceptual priming, the evidence seems at best mixed. Yovel and Paller (2004) had found no FN400 for nonfamous faces; however, FN400 effects have later been found for this type of stimuli (Curran and Hancock, 2007; but see also MacKenzie and Donaldson, 2007). Likewise, more direct tests using potentially less meaningful stimuli such as squiggles, blobs, or kaleidoscope images have been similarly divided (Voss and Paller, 2009 showed no FN400 effect, but see Groh-Bordin et al., 2006; Curran et al., 2002). The current study shows FN400 effects can reliably appear at encoding; at the least, this widens the field for future investigations on the issue.
Interestingly, the FN400 effect also did not appear to be influenced by back-sorting, as the magnitude of the effects for both conceptual and identical repetitions seemed little affected by subsequent memory. One of our interests was not only whether the traditional recognition test effects could be found during incidental encoding, but whether they could be used to predict later performance. We found that the parietal old/new effect was predictive and familiarity was not; however, it bears mentioning that our task required discrimination that went beyond familiarity -- to confidently distinguish between types of repetitions at test one needed to recollect both presentations. Therefore, it is possible that had we asked just for old/new, the FN400 might have predicted subsequent memory as well. Indeed, previous research suggests that the FN400 might predict performance given a more lenient test. In a regular old/new test, FN400s can distinguish missed old items from correctly identified old items in the absence of strong parietal old/new effects (Rugg et al., 1998), and in a midazolam study, reliable FN400 effects (but no parietal old/new) were found for old items at test, with performance still well above chance (Curran et al., 2006). The absence of FN400 differences between back-sorted and non-back-sorted items in our results may be a byproduct of the fact that, even when not back-sorted, identical repetitions were rarely classified as ‘new’ (only about 10% of the time).
Reliable parietal old/new effects were found for different exemplars when subjects’ judgments were accurate, and interestingly, were also significant for different exemplars that were later identified as “same”. This result seems sensible when considering that the parietal old/new effect is considered an indication of recollection. If a conceptual repetition is noted, then a parietal old/new effect should occur at the second presentation. In addition to the parietal old/new effect, different exemplars later recollected as identical were also less positive than the different exemplars correctly identified as “Different”. Conceivably, this smaller positivity represents something like the standard ‘Dm’ effect, in which less positive amplitudes are associated with stimuli that are less strongly encoded. That is, subjects notice there is a repetition while doing the task, but the stimuli are not encoded as strongly as correctly identified different exemplars, and thus at test they could not remember whether or not the pictures were identical.
It is worth noting that correctly identified ‘same’ repetitions showed only a weak parietal old/new effect. Given that recollection at the second encoding was incidental to the task, recollection has to occur in a high percentage of trials to show a reliable effect. However, in the current study, subjects showed a bias to respond “same” when unsure of the type of repetitions, so correct responses for identical repetitions are likely to have fewer recollections and more ‘lucky guesses’ compared to different pairs that were correctly identified. This mixture of guesses in the identical condition may have obscured the parietal old/new effect by watering it down. We had attempted to reduce that bias by including the single condition, thereby allowing subjects to respond “single” if they did not remember two presentations. In a previous version of this paradigm (e.g., Manelis et al., 2011; Manelis et al., in press) we did not include a single condition and the bias to respond “same” when unsure was very strong. Nevertheless, this bias to assume that two repetitions are identical still occurred in the present study as shown by the much more liberal β for same than different pairs.
Prior to the current study, studies of subsequent memory in ERP were confined to studying the standard Dm effect. The current study suggests that the components typically studied at test can be observed during encoding, and may be similarly predictive. In this case, the parietal old/new predicted the type of answer for different exemplars, and the effect combined with a greater positivity predicted the correct answer. A limitation of using the standard Dm paradigm for ERP is that it is insufficiently precise to enable discrimination between better versus worse responses; for instance, Dm studies of remember/know often have trouble distinguishing between ‘know’ and ‘new’ responses (Friedman and Trott, 2000; Yovel & Paller 2004). The current study offers a method of finessing these problems, as the effects found for different exemplars can distinguish not only between correct and incorrect, but also better and worse guesses.
Lastly, while our primary concern has been whether the FN400 and parietal old/new effect would appear in incidental tasks if we sort by memory strength, it is also worth discussing what these components might tell us about the encoding process. While both the FN400 and the parietal old/new effect reliably appeared at encoding, only the parietal old/new, which is thought to index recollection, predicted better performance. As our method used Dm, literally ‘differences due to subsequent memory’, we have focused on how back-sorting might let us predict these effects. However, studies of learning are sometimes concerned with whether recollection at encoding causes better memory. The so-called test effect popularized by Roediger and Karpicke (2006) demonstrates that retention is greater when subjects are asked to recall material rather than merely passively study it. While our subjects were not required to recall or recollect, the parietal old/new data suggest that they did. Further, the repetitions where this effect was strongest were those that demonstrated better encoding: the correct response or the next best answer (correct in terms of two presentations). We had used the parietal old/new as a marker of these better encoded trials, but it could well be that the incidental retrieval, indexed by this component, is responsible for the ultimate better encoding of these trials, not the other way around.
Theories of explicit and implicit memory have considered retrieval intention to be a primary way to distinguish between types of memories. However, many of these studies have not equated the memory strength of the items being compared. When memory strength was equated in the current study, the parietal old/new effect was found to be a reliable Dm component, predicting better performance at test. More importantly, the current study shows that when the items of an incidental task are partitioned by memory strength, the two ERP signatures of ‘intentional’ retrieval can appear in an incidental task. These data support the view that implicit and explicit memory rely on the same underlying memory system, and that distinctions between ‘explicit’ and ‘implicit’ might be better thought of as differences in task, not memory itself.
4. Methods
4.1 Subjects
Twenty eight Carnegie Mellon University students (mean age: 19.6, 16 males) participated in this study to fulfill a research requirement for their undergraduate psychology course. Five subjects were excluded from the subsequent analyses. One subject was excluded because of an equipment malfunction. Four additional subjects were excluded because artifact rejection resulted in losing more than two-thirds of each subjects’ trials. All analyses were conducted on the remaining twenty-three subjects.
4.2 Materials and Design
The picture stimuli consisted of color pictures of common man-made (e.g., tea kettle) and natural (e.g., apple) objects. Objects were chosen by first selecting concrete words from the MRC database (Kucera and Francis, 1967) that described either man-made or natural objects. Then two different images were selected from the internet, using the corresponding word as the search cue. Two different pictures of each object were selected that were not visually similar. Subjects from the lab who had not selected the images had to correctly label all pictures before we used them in the final stimulus pool.
For both types of stimuli, pictures were edited using Photoshop to ensure that the nature of the background (all white versus extraneous details) was not confounded with type of stimulus (i.e., pictures of natural objects were not more likely to have a background scene than pictures of man-made objects). There were two different pictures that corresponded to each word/concept so that each word was randomly assigned to repetition condition for each subject. For identical repetitions, one of the two pictures was randomly selected from the pair. For the different repetition condition, the order of presentation of the two pictures was randomly determined for each subject.
There were two phases to the study: an incidental encoding task and a surprise recognition test based on the preceding phase. Trials from the encoding phase formed a 2 (object type: man-made versus natural) × 3 (type of repetition: repeated identical, repeated different or single presentation) within subjects design. Subjects were to judge whether each sequentially presented image was a natural or man-made object. For each subject, stimuli were randomly assigned to type of repetition with the constraint that there was an equal number of each object (natural and man-made) per condition. The presentation order of stimuli within the encoding phase was randomly determined for each subject with the constraint that an equal number of singles appeared within the first and second half of the list and that the repetition pairs (identical or conceptual/different exemplar) were not presented in the same half of the 300 trials. In other words, the first half contained the first presentation of all stimuli that were to be repeated and half of the stimuli that were to be presented only once. There were 50 pictures per repetition type per half of the experiment. The entire encoding phase consisted of 300 trials. During the test phase, subjects were presented with words that corresponded to an object in the encoding phase. Additionally, foil word probes were shown that were selected from a list of potential stimuli that had not been used during the encoding phase for that particular subject. There was a target word for every picture shown and there were half as many foil word probes giving 270 total test trials. Order of presentation was randomly determined for each subject.
4.3 ERP Methods
EEG waveforms were recorded using 32 Ag-AgCL sintered electrodes (10–20 system) and a bio-amplification system (Neuroscan Inc., Sterling, VA). Neuroscan was used in conjunction with E-Prime (which recorded all behavioral data) to present the experiment and record behavioral and EEG measures. E-Prime was also used to insert markers into the waveform recording when behavioral events happened such as a key press for a response.
Participants sat in a Faraday Cage (an electrically shielded booth), looking through a window of radio-frequency shielded glass at stimuli presented on a computer display that was placed approximately one inch behind the window. Impedances were adjusted to be less than 5 kΩ. Data were sampled at a rate of 1 kHz with a band pass filter of 0.1–200 Hz.
In order to monitor and account for eye movements, electrodes were placed above and below the orbit of the left eye to record vertical eye movements. Horizontal eye movements were monitored with an additional pair of electrodes at the external canthi. The right mastoid served as the reference electrode, and scalp recordings were algebraically re-referenced offline to the average of the right and left mastoids.
Using EEGLab (Delorme and Makeig, 2004), a toolbox of Matlab, data were corrected for ocular artifacts using ICA. Epochs of 1000 ms (including a 200 ms baseline) were extracted from the continuous recording and corrected over the pre-stimulus interval. Trials contaminated with voltages above 75 μV or below −75 μV were excluded from the analysis. The segmented data were then averaged across trials within each subject for each condition. For plotting, data were first smoothed using a 30 Hz low pass filter.
4.4 Procedure
Subjects sat in a Faraday cage approximately 70 cm away from a computer monitor and were instructed that the task was to judge each picture as natural or man-made, pressing the 7 key for natural and 2 key for man-made. They were told to keep their fingers resting of the response keys and to respond as quickly as possible while retaining high accuracy. Each trial began with a fixation cross that lasted between 1 and 1.2 seconds, and varied in duration from trial to trial. The trials in the encoding phase were self-paced, with each object being displayed until the subject made a response. In all, the encoding phase lasted approximately 10 minutes.
After subjects completed the encoding phase, they took a break for 5 minutes and then proceeded to a surprise recognition test. Subjects were shown words one at a time that described a particular category of natural or man-made items. The word either referred to one of the pictures that was shown during encoding or was a novel category word (there was no picture that corresponded to the word). Subjects pressed one of four keys, indicating whether or not they thought a picture had been seen corresponding to the word probe. The four possible responses were new (K key), single (H key), different (D key) or same (A key). Here too, subjects were instructed to respond as quickly as possible while retaining high accuracy. Before each trial, there was a fixation cross that varied in duration between 1 and 1.2 seconds. The word displays in the test phase were also self-paced. No feedback was provided.
Supplementary Material
In ERP, Dm dissociations have not been as successful as those for the FN400 and LPC
An incidental task explores whether these test components can be found at encoding
We backsort ‘conceptual’ and identical repetitions for potential Dm effects
The FN400 and LPC were robust at encoding, and predict performance at test
The FN400 was more reliably found for identical, not conceptual repetitions
Acknowledgments
The authors wish to thank Matthew Walsh for programming and technical assistance and Lindsay Victoria for help with data collection, stimulus generation and comments on earlier drafts of this manuscript. This project was supported by the NIH training grant T32MH019983, and by author LR’s NIH grant 5R01MH052808. Additionally, author AK was supported by the NIH training grant R90DA023426.
Footnotes
While there were two other potential responses, “Single” and “New”, the (error) response rate to these two potential responses for same and different pairs was indistinguishable (see Figure 1) so those response categories should not affect our calculations.
It is tempting to conclude based on these results that there is only priming for identical repetitions; however, the story may be more complicated than that. It is important to note that, although the different presentations with correct responses show the same level of priming as single presentations, numerically both same and different repetitions produce more priming when subjects respond “single” than when they remember that there were two presentations (see Fig. S1). Our explanation is that there is a mixture of facilitation from repetition (priming) and a cost for noticing the repetition. That is, when subjects are reminded on the second presentation of the same or similar exemplar shown earlier, their response times are somewhat slowed from this “double-take.” On those trials where they fail to recollect the earlier presentation, there is still priming but no added cost of the “double-take.” We have replicated this pattern in three other experiments using a related paradigm (Manelis et al., 2011; Manelis et al., in press; Victoria et al., 2009).
To check for potential laterality effects that may have been obscured by this clustering, we ran these same electrodes with laterality as an additional factor (left lateral, center, right lateral) and found no significant interactions with laterality. We also ran an initial set of exploratory analyses on just the Fz and Pz electrodes, as these had appeared to have the strongest differences from visual inspection. The pattern of effects found were the same, although amplitude differences were slightly stronger.
An initial set of exploratory analyses had compared second correct second presentations to ‘singles’ instead of their first presentation at only the Pz electrode (which seemed to be where parietal old/new effects were strongest for Sames). In this case, the effect was reliable at alpha = .05, (i.e., with no correction for multiple comparisons).
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Curran T. Brain potentials of recollection and familiarity. Memory & Cognition. 2000;28:923–938. doi: 10.3758/bf03209340. [DOI] [PubMed] [Google Scholar]
- Curran T, Cleary AM. Using ERPs to dissociate recollection from familiarity in picture recognition. Cognitive Brain Research. 2003;15:191–205. doi: 10.1016/s0926-6410(02)00192-1. [DOI] [PubMed] [Google Scholar]
- Curran T, Debuse C, Woroch B, Hirshman E. Combined pharmacological and electrophysiological dissociation of familiarity and recollection. The Journal of Neuroscience. 2006;26(7):1979–1985. doi: 10.1523/JNEUROSCI.5370-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Curran T, Doyle J. Picture superiority doubly dissociates the ERP correlates of recollection and familiarity. Journal of Cognitive Neuroscience. 2011;23(5):1247–1262. doi: 10.1162/jocn.2010.21464. [DOI] [PubMed] [Google Scholar]
- Curran T, Hancock J. The FN400 indexes familiarity-based recognition of faces. NeuroImage. 2007;36:464–471. doi: 10.1016/j.neuroimage.2006.12.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Curran T, Tanaka JW, Weiskopf DM. An electrophysiological comparison of visual categorization and recognition memory. Cognitive, Affective, & Behavioral Neuroscience. 2002;2:1–18. doi: 10.3758/cabn.2.1.1. [DOI] [PubMed] [Google Scholar]
- Delorme A, Makeig S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. Journal of Neuroscience Methods. 2004;134:9–21. doi: 10.1016/j.jneumeth.2003.10.009. [DOI] [PubMed] [Google Scholar]
- Duarte A, Ranganath C, Winward L, Hayward D, Knight RT. Dissociable neural correlates for familiarity and recollection during the encoding and retrieval of pictures. Cognitive Brain Research. 2004;18:255–272. doi: 10.1016/j.cogbrainres.2003.10.010. [DOI] [PubMed] [Google Scholar]
- Düzel E, Yonelinas AP, Mangun GR, Heinze HJ, Tulving E. Event-related brain potential correlates of two states of conscious awareness in memory. Proceedings of the National Academy of Sciences of the USA. 1997;94(11):5973–5978. doi: 10.1073/pnas.94.11.5973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ecker UKH, Zimmer HD, Groh-Bordin C. Color and context: An ERP study on intrinsic and extrinsic feature binding in episodic memory. Memory & Cognition. 2007;35(6):1483–1501. doi: 10.3758/bf03193618. [DOI] [PubMed] [Google Scholar]
- Ecker UKH, Zimmer HD. ERP Evidence for flexible adjustment of retrieval orientation and its influence on familiarity. Journal of Cognitive Neuroscience. 2009;21(10):1907–1919. doi: 10.1162/jocn.2009.21135. [DOI] [PubMed] [Google Scholar]
- Friedman D, Johnson R. Event-related potential (ERP) studies of memory and retrieval: A selective review. Microscopy Research and Technique. 2000;51:6–28. doi: 10.1002/1097-0029(20001001)51:1<6::AID-JEMT2>3.0.CO;2-R. [DOI] [PubMed] [Google Scholar]
- Friedman D, Trott C. An event-related potential study of encoding in young and older adults. Neuropsychologia. 2000;38:542–557. doi: 10.1016/s0028-3932(99)00122-0. [DOI] [PubMed] [Google Scholar]
- Green DM, Swets JA. Signal detection theory and psychophysics. Vol. 1974. New York: Wiley; 1966. [Google Scholar]
- Groh-Bordin C, Zimmer HD, Ecker UKH. Has the butcher on the bus dyed his hair? When color changes modulate ERP correlates of familiarity and recollection. NeuroImage. 2006;32:1879–1890. doi: 10.1016/j.neuroimage.2006.04.215. [DOI] [PubMed] [Google Scholar]
- Kučera H, Francis WN. Computational analysis of present day American English. Providence, RI: Brown University Press; 1967. [Google Scholar]
- Küper K, Groh-Bordin C, Zimmer HD, Ecker UKH. Electrophysiological correlates of exemplar-specific processes in implicit and explicit memory. Cognitive, Affective & Behavioral Neuroscience. 2012;12:52–64. doi: 10.3758/s13415-011-0065-7. [DOI] [PubMed] [Google Scholar]
- MacKenzie G, Donaldson DI. Dissociating recollection from familiarity: electrophysiological evidence that familiarity for faces is associated with a posterior old/new effect. NeuroImage. 2007;36:454–463. doi: 10.1016/j.neuroimage.2006.12.005. [DOI] [PubMed] [Google Scholar]
- Manelis A, Paynter CA, Wheeler ME, Reder LM. Repetition related changes in activation and functional connectivity in hippocampus predict subsequent memory. Hippocampus. doi: 10.1002/hipo.22053. (in press) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manelis A, Wheeler ME, Paynter CA, Storey L, Reder LM. Opposing patterns of neural priming in same-exemplar versus different-exemplar repetition predict subsequent memory. NeuroImage. 2011;55(2):763–772. doi: 10.1016/j.neuroimage.2010.12.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paller KA, Kutas M, Mayes AR. Neural correlates of encoding in an incidental learning paradigm. Electroencephalography and Neurophysiology. 1987;67:360–371. doi: 10.1016/0013-4694(87)90124-6. [DOI] [PubMed] [Google Scholar]
- Paller KA, Boehm SG, Voss JL. Validating neural correlates of familiarity. Trends in Cognitive Sciences. 2007;11:243–250. doi: 10.1016/j.tics.2007.04.002. [DOI] [PubMed] [Google Scholar]
- Reder LM, Park H, Kieffaber PD. Memory systems do not divide on consciousness: Reinterpreting memory in terms of activation and binding. Psychological Bulletin. 2009;135(1):23–49. doi: 10.1037/a0013974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roediger HL, Karpicke JD. The power of testing memory: Basic research and implications for educational practice. Perspectives on Psychological Science. 2006;1:181–210. doi: 10.1111/j.1745-6916.2006.00012.x. [DOI] [PubMed] [Google Scholar]
- Rugg MD, Curran T. Event-related potentials and recognition memory. Trends in Cognitive Sciences. 2007;11(5):251–257. doi: 10.1016/j.tics.2007.04.004. [DOI] [PubMed] [Google Scholar]
- Rugg MD, Mark RE, Walla P, Schloerscheidt AM, Birch CS, Allan K. Dissociation of the neural correlates of implicit and explicit memory. Nature. 1998;392:595–598. doi: 10.1038/33396. [DOI] [PubMed] [Google Scholar]
- Senkfor AJ, Van Petten C. Who said what? An event- related potential investigation of source and item memory. Journal of Experimental Psychology: Learning Memory, and Cognition. 1998;24:1005–1025. doi: 10.1037//0278-7393.24.4.1005. [DOI] [PubMed] [Google Scholar]
- Smith ME. Neurophysiological manifestations of recollective experience during recognition memory judgments. Journal of Cognitive Neuroscience. 1993;5(1):1–13. doi: 10.1162/jocn.1993.5.1.1. [DOI] [PubMed] [Google Scholar]
- Wilding EL, Rugg MD. An event-related potential study of recognition memory with and without retrieval of source. Brain. 1996;119:889–905. doi: 10.1093/brain/119.3.889. [DOI] [PubMed] [Google Scholar]
- Woodruff CC, Hayama HR, Rugg MD. Electrophysiological dissociation of the neural correlates of recollection and familiarity. Brain Research. 2006;1100:125–136. doi: 10.1016/j.brainres.2006.05.019. [DOI] [PubMed] [Google Scholar]
- Victoria LW, Reder LM, Dutcher JM, Aizenstein HJ, Quinlan JJ, Gyulai F. Recognition and priming under midazolam. Poster presented at Psychonomics; Boston, MA. 2009. [Google Scholar]
- Voss JL, Paller KA. Fluent conceptual processing and explicit memory for faces are electrophysiologically distinct. Journal of Neuroscience. 2006;26(3):926–933. doi: 10.1523/JNEUROSCI.3931-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Voss JL, Paller KA. Remembering and knowing: electrophysiological distinctions at encoding but not retrieval. NeuroImage. 2009;46:280–289. doi: 10.1016/j.neuroimage.2009.01.048. [DOI] [PubMed] [Google Scholar]
- Voss JL, Lucas HD, Paller KA. Conceptual priming and familiarity: Different expressions of memory during recognition testing with distinct neurophysiological correlates. Journal of Cognitive Neuroscience. 2010;22:2638–2651. doi: 10.1162/jocn.2009.21341. [DOI] [PubMed] [Google Scholar]
- Yonelinas AP. The nature of recollection and familiarity: a review of 30 years of research. Journal of Memory and Language. 2002;46:441–517. [Google Scholar]
- Yovel G, Paller KA. The neural basis of the butcher-on-the-bus phenomenon: when a face seems familiar but is not remembered. NeuroImage. 2004;21:789–800. doi: 10.1016/j.neuroimage.2003.09.034. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.