

Abstract
The divergence of the two αβ T cell subsets defined by the mutually exclusive expression of CD4 and CD8 glycoproteins is an important event during the intrathymic differentiation of T lymphocytes. This reviews briefly summarizes the mechanisms that promote commitment to the CD4 or CD8 lineage in the thymus, and discusses the transcription factor circuits and epigenetic mechanisms that concur to maintain lineage integrity in post-thymic cells and how they adjust to allow expression of genes required for effector cell differentiation.
CD4 and CD8 T cells
The two T cell subsets defined by the expression of CD4 and CD8 form the bulk of T cells carrying an αβ TCR and recognizing peptides bound to class I or class II Major Histocompatibility Complex (MHC) molecules (MHC-I or MHC-II, respectively). The differentiation of these two subsets in the thymus has been particularly well studied [1-3] and has important characteristics. Its outcome is binary and defines two distinct lineages: mature T cells express CD4 or CD8 but not both, and their expression of either coreceptor is stable and propagated through cell division. CD4-CD8 differentiation matches MHC restriction, so that MHC II-restricted cells express CD4 and MHC I-restricted cells CD8. It also corresponds with functional differentiation, such that CD4 cells are precursors of helper or regulatory cells, whereas CD8 cells are cytotoxic precursors. Analyses in mice genetically engineered to generate T cells in which coreceptor gene expression is mismatched to MHC specificity have shown that function segregates with coreceptor gene expression rather than MHC specificity [4,5]. This indicates that the same differentiation process determines coreceptor expression and functional differentiation, and that this process is normally set to match lineage differentiation to MHC specificity. Because of this coordinate matching of lineage and functional differentiation to MHC specificity, CD4-CD8 lineage ‘choice’ is thought to be central to the function of the immune system.
CD4-CD8 lineage differentiation is a late event during the intrathymic development of αβ T cells (Fig. 1) [6]. It occurs in committed αβ precursors that have rearranged both genes encoding their TCR chains (TCRβ and TCRα) and started expression of both CD4 and CD8 coreceptors, thereby becoming ‘double positive’ (DP) thymocytes. In fact, most DP thymocytes carry TCRs unable to productively bind self-MHC peptide complexes expressed on the thymic stroma, and die in the thymic cortex by ‘neglect’ [7,8]. Only cells with appropriate avidity for such self ligands survive, a process called positive selection, and differentiate into mature T cells; the ‘choice’ of the CD4 and CD8 lineage is associated with that process. These selection and differentiation events complete the elaboration of the T cell repertoire, resulting in large populations of cells that are antigen specific, long lived and, for most of them, functionally dormant. Such cells populate peripheral lymphoid organs and are unique in that they are neither terminally differentiated nor post-mitotic. Upon antigen recognition, they proliferate and acquire effector properties that largely depend on environmental (notably cytokine) signals [9-11].
Figure 1. Outline of T cell differentiation.
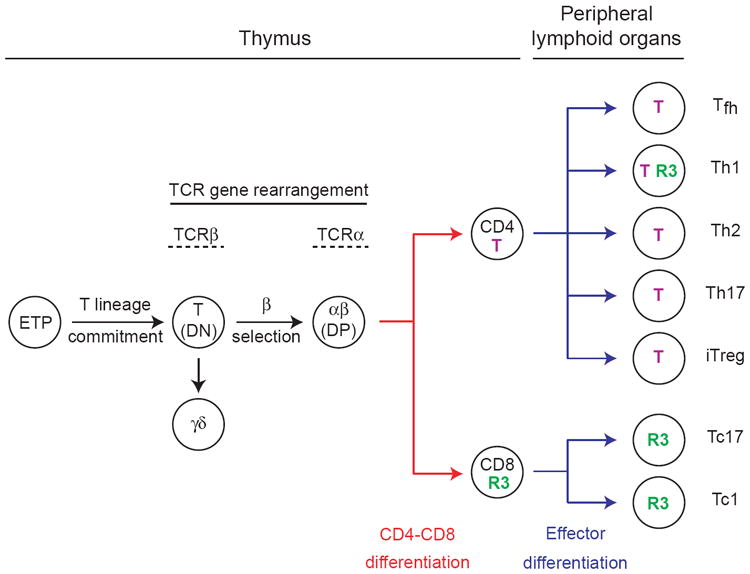
Key intermediates in T cell development are represented, including uncommitted thymic precursors (Early thymic progenitors, ETP), T-lineage committed cells (T) rearranging TCR genes prior to the onset of CD4 or CD8 expression (‘double negative’ cells, DN), αβ lineage precursors (DP thymocytes) and γδ T cells [for review, see Refs. 6,90,91]. Key developmental checkpoints are shown by arrows: commitment of early intrathymic precursors (at the DN2 stage) to the T lineage, commitment to DN cells to the αβ (β-selection) or γδ lineage, CD4-CD8 and effector differentiation. The developmental windows for TCRβ and TCRα rearrangement are indicated. The progeny of DP thymocytes, CD8 and CD4 cells are shown, as is a non-exhaustive list of the effector type they generate upon activation. Subsets with high-level expression of Thpok (T, purple) or Runx3 (R3, green) are schematically depicted. Effector CD8 cells that make IL-17 (Tc17) have been reported both in vitro and in vivo, and display reduced cytotoxic activity with reduced expression of cytotoxic genes perforin, granzyme B and of the transcription factor Eomes [92-94] but nonetheless continue expressing Runx3 (Y.X, L.W. and R.B, unpublished results).
The basic CD4-CD8 commitment machinery in the thymus
This review briefly surveys the transcriptional control of the differentiation of CD4 and CD8 lineages in the thymus, and discusses how transcriptional components that enact this ‘decision’ are subsequently re-used during the differentiation of CD4 and CD8 effector T cells. We will refer the reader to recent reviews [1-3] for a discussion of the extra-cellular signals and transcription factors that control CD4-CD8 choice, and focus on the lineage commitment machinery, of which two key components have been identified.
The zinc finger transcription factor Thpok (also called cKrox, and encoded by a gene named Zbtb7b that we will refer to as Thpok) is expressed by all CD4 cells and is critical for CD4 lineage commitment [12-15]. Thpok is normally not expressed by DP thymocytes, and is up-regulated, through mechanisms that have yet to be elucidated, in MHC II- but not MHC I-signaled thymocytes [12,16]. The role of Thpok in CD4 T cell differentiation is demonstrated in Thpok-deficient mice, in which MHC II-restricted T cells are in normal or subnormal numbers but differentiate into the CD8 rather than the CD4 lineage [13-15,17]. Both loss- and gain-of-function analyses have shown that Thpok is a repressor of CD8-lineage gene expression, including of Cd8 genes themselves [18,19] and of Runx3, a transcription critical for CD8-lineage differentiation (see below) [14,15,19]. Accordingly, enforced Thpok expression in developing MHC I-restricted thymocytes prevents their CD8-lineage differentiation [12,16]. Altogether, these studies indicate that Thpok, through its ability to repress CD8-lineage genes, is a key factor for CD4 commitment (biologically defined as the loss of CD8 developmental potential). Its contribution to the expression of CD4-lineage genes, which we will refer to its ‘specification’ function, remains to be fully evaluated.
On the other hand, Runx transcription factors are essential for CD8 T cell differentiation. In mammals, the Runx family includes three molecules (Runx1, Runx2, Runx3, of which Runx1 and Runx3 are expressed in T cells), with strong conservation across species and involvement in multiple differentiation processes [3,20]. Runx molecules can serve as transcriptional activators, often in synergy with other transcription factors, or as transcriptional repressors; in both instances, they bind DNA as obligate heterodimers with the unrelated factor Cbfβ.
The expression of Runx1 and Runx3 in T cells follows different patterns. Runx1 is expressed at most stages of T cell development. Its expression is lower in CD8- than in CD4-lineage cells [21-23]. accordingly, while disruption of Runx1 in the αβ lineage (i.e. at the DP stage using Cd4-Cre excision of a conditional Runx1 allele) has no known consequence on CD8 cell generation, it has pleiotropic effects in CD4 T cells [23,24]. In contrast, Runx3 expression in thymocytes is largely limited to CD8-lineage cells and ‘non conventional’ subsets (e.g. γδ or NK T cells) [15,23-26]. Runx3 disruption in the αβ T lineage impairs several aspects of CD8 T cell differentiation [23-25]. Runx3-deficient CD8-lineage cells are in reduced numbers and fail to properly silence Cd4, so that some appear CD4+CD8+ in peripheral lymphoid organs. Deletion of one Runx1 and both Runx3 alleles causes all remaining CD8 cells to express CD4, whereas complete disruption of Runx activity prevents CD8 T cell development [15,25,27]. This indicates that the subdued phenotype of Runx3-deficient cells is due to compensation by Runx1; indeed, the potential for functional redundancy between the two genes is increased by the enhanced expression of Runx1 in Runx3-deficient CD8 cells [23]. Altogether, these findings implicate Runx3 as critical for the generation of CD8 cells in the thymus.
Runx activity both promotes expression of genes characteristic of the CD8 lineage (a ‘specification’ function), including those encoding CD8α and CD8β subunits and cytotoxic enzymes such as perforin and Granzyme B [28], and the commitment of T cell precursors to the CD8 lineage. The committing function has been exceptionally well documented at the molecular level. Runx3 binds and represses the two defining CD4-lineage genes, Cd4 and Thpok. In the Cd4 locus, Runx-binding sites are critical components of the Cd4 silencer, an intronic element required to restrain the expression of Cd4 to CD4-lineage cells (Fig. 2) [24,29,30]. In addition to its effect on Cd4, Runx3 is thought to contribute to Thpok repression in CD8-differentiating thymocytes. The repressive effect of Runx3 on Thpok has similarities and differences with that on Cd4, and its analysis is also complicated by its functional redundancy with Runx1 [27]. Similar to Cd4, Runx binding sites have been identified within a silencer (itself part of a ‘Distal Regulatory Element’ [DRE]) located upstream of the Thpok promoter and needed to prevent Thpok gene expression in DP thymocytes and CD8 cells (Fig. 3)[27,31]. The current evidence indicates that Runx1 and Runx3 physiologically contribute to Thpok repression at distinct developmental stages. However, the developmental ‘timing’ of repression is different from Cd4: in DP thymocytes, Runx1 binds the Thpok silencer and this binding contributes to prevent premature transcription [27]. Biochemical and genetic evidence suggest that Runx3 performs the same function in CD8-differentiating thymocytes in a manner redundant with Runx1 (as only the complete disruption of Runx activity, not that of Runx1 or Runx3 separately) ‘redirects’ MHC I-restricted thymocytes into the CD4 lineage [23,24,27].
Figure 2. Thpok and Runx3 antagonism at the Cd4 locus.
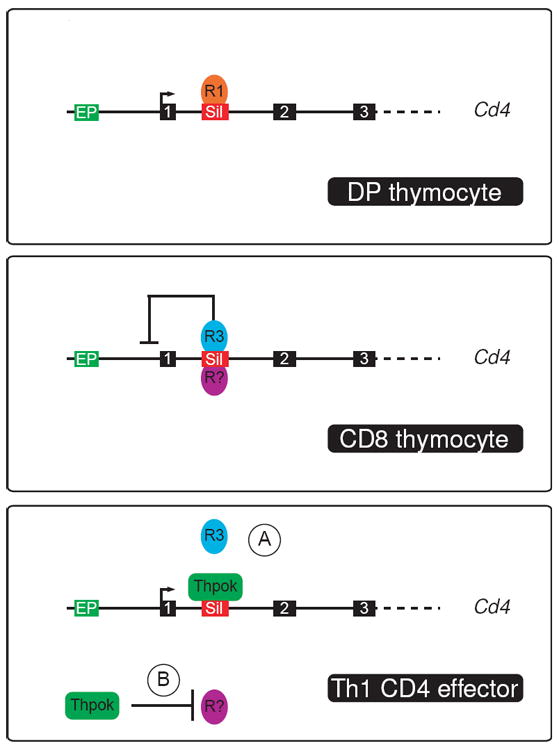
The Cd4 locus is schematically depicted with the transcription start site shown as an arrow and the first three exons as numbered black boxes. The Cd4 silencer (Sil) and proximal enhancer (EP) are shown as red and green boxes, respectively. In CD8-lineage thymocytes (middle), Runx3 (R3) and putative additional repressors (R?) binds the silencer, resulting in Cd4 repression and eventual silencing. In DP thymocytes (top), Runx1 (R1) binds the silencer, but fails to repress Cd4. Although this may be because of the absence of additional repressors (as depicted), enforced Runx3 expression in DP thymocytes similarly represses Cd4 repression, consistent with the possibility that the dosage of Runx proteins affects their ability to repress Cd4. In Th1 CD4 effectors (bottom), Runx3 fails to repress Cd4. Two mechanisms have been proposed to explain this effect: (A) Thpok binds the Cd4 silencer, suggesting that it could prevent recruitment of Runx3 (as depicted) or of required co-repressors, or antagonize their repressive activity; (B) Thpok could repress the expression of required co-repressors.
Figure 3. Transcriptional plasticity at the Thpok locus.
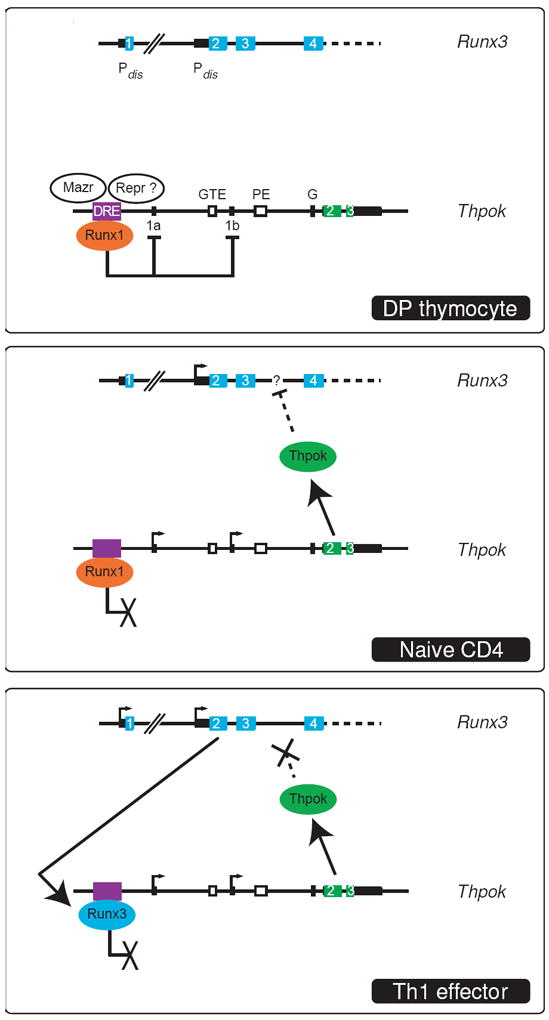
The Runx3 and Thpok loci are schematically represented in three separate boxes. For each gene, transcription start sites are shown as arrows and exons as numbered cyan or green boxes; non-coding exonic sequences are depicted as thinner black rectangles. Note the two alternative promoters and first exons (1a and 1b) of Thpok. Boxes (top) indicate the distal regulatory element (DRE, which includes or overlaps with the silencer [27,31]) the general T lymphoid element (GTE), the proximal enhancer (PE), and a conserved sequence (G) that includes a Gata3 binding site [13]. The distal (Pdis) and proximal (Pprox) Runx3 promoters are indicated upstream of exons 1 and 2, respectively. In DP thymocytes (top), Thpok is not transcribed because of reversible repression involving Runx1, Mazr and potentially unknown other repressors (Repr ?). The Runx3 distal promoter, that gives rise to protein-producing transcripts, is not active. In CD4 SP thymocytes and naïve CD4 T cells (middle), Thpok is expressed and represses Runx3 through so far unknown mechanisms (dotted line); note that this does not prevent transcription from the proximal promoter [23]. Although Runx1 is expressed and bound to the silencer, it does not repress Thpok, possibly because of the absence of the other repressors (see text for discussion). In Th1 effectors (bottom), Thpok is expressed but no longer represses the Runx3 distal promoter. Conversely, Runx3 fails to repress Thpok; it is shown as bound to the silencer by analogy with Runx1 in resting cells (see text for discussion). Drawings are not on scale.
These findings have led to a model in which the commitment engine in the thymus is a dual negative regulatory loop involving both Runx1 and Runx3, and Thpok (Fig. 3). One prominent characteristic of this loop is to produce a binary developmental outcome where cells become CD4 or CD8, respectively expressing either Thpok or Runx3 but not both, thereby preventing ‘partial’ differentiation status. Indeed, analyses in mice carrying hypomorphic Thpok alleles, that express reduced levels of a wild-type Thpok protein, indicate that even in such a situation the outcome of differentiation remains binary, with generation of cells expressing CD4 or CD8 but not both [15,19]. Similar regulatory modules have been shown to operate in other biological systems and have long been identified for their ability to unambiguously separate binary fates [32,33].
Important regulatory components are probably missing from this simple model. Notably, Runx proteins are necessary but not sufficient for Thpok repression as enforced expression of Runx3 in DP thymocytes fails to prevent Thpok expression and CD4 T cell differentiation [34]. Accordingly, genetic analyses of Thpok cis-regulatory elements have identified motifs aside from Runx-binding sites important to restrain Thpok expression to MHC II-restricted thymocytes [31]. This suggests that other factors cooperate with Runx molecules to promote Thpok repression. The BTB-POZ zinc finger transcription factor Mazr, recently shown to bind the Thpok silencer in DP thymocytes and to contribute to Thpok repression in DP thymocytes [35], is one such molecule, and others may remain to be discovered. Because Runx1 is normally expressed in CD4-differentiating thymocytes and post-thymic CD4 cells, it is likely that repressing the expression of such additional factors, or overcoming their activity, is essential for Thpok up-regulation during the differentiation of CD4 T cells in the thymus.
Given the importance of the Thpok-Runx dual negative regulatory loop for CD4-CD8 commitment in the thymus, the question arises whether it continues to operate in post-thymic T cells. As we will see, although naïve post-thymic T cells express only either Thpok or Runx3, that is not the case of differentiating effector cells, which can express both factors and yet retain their defining Cd4 or Cd8 gene expression. These observations raise the question of how CD4-CD8 lineage integrity is maintained in post-thymic T cells, which we will now discuss.
Maintaining CD8 lineage integrity: the case for rigidity
Epigenetic silencing of CD4-lineage genes in CD8 cells
The studies that addressed how mature T cells maintain their CD4-CD8 lineage differentiation have reached contrasting results for CD4 and CD8 lineage cells. In CD8 cells, Runx3 is needed for cytotoxic effector differentiation, including expression of cytotoxic enzymes perforin and granzyme B, and of IFNγ, a prototypical CD8 cytokine [28]. In contrast, there is evidence that CD8 cells epigenetically silence CD4-lineage genes independently from Runx3 expression [36]. This idea first emerged from studies of Cd4 expression, using mice carrying a ‘floxed’ Cd4 silencer element, whose excision by the Cre recombinase leaves a ‘silencer-less’ Cd4 allele [37]. When the silencer is deleted in the germline or in DP thymocytes, all CD8 lineage cells maintain Cd4 expression and therefore appear as CD4+CD8+ in peripheral lymphoid organs [37,38]. This demonstrates the critical role of the silencer, as both CD4- and CD8-lineage cells express transcription factors required for Cd4 gene expression. In contrast, deletion of the silencer in post-thymic CD8 cells (that have developed with an intact silencer and are thus CD4−CD8+) fails to restore Cd4 expression [37]. The simplest interpretation of these results is that epigenetic modifications of the Cd4 locus, established during the late stages of intrathymic differentiation, irreversibly silence Cd4 expression. Such ‘silencing’ is maintained and propagated through cell division even in the absence of the transcription factors (and cis-regulatory elements) that installed it in developing thymocytes, and is therefore distinct of ‘active repression’ that requires the continued recruitment of repression factors to relevant cis-regulatory elements [30,37,39]. Further supporting the idea of a stable epigenetic repression of CD4-lineage genes, enforced expression of Thpok in CD8 cells, by retroviral transduction, fails to cause Cd4 gene reexpression, or expression of the CD4-lineage marker Cd40lg, although enforced expression of Thpok in thymocytes counteracts the Cd4-repressive function of Runx3 [18,40].
Whether Thpok is also subject to epigenetic silencing in CD8-lineage cells remains to be determined, although the current evidence suggests this would not be to the same extent as Cd4. While Thpok expression is not detected in ex-vivo CD8 cells (whether naïve or memory) [12-16], analyses of mice carrying a ‘silencer-less’ Thpok reporter allele have suggested that CD8 cells have all transcription factors necessary for Thpok expression [27]. However, it remains to be determined, using conditional deletion analyses, if this reflects true epigenetic silencing of Thpok, similar to that seen for Cd4, or active repression mediated by the Thpok silencer in mature CD8 cells. Supporting the second possibility, and suggesting greater plasticity at the Thpok than at the Cd4 locus, it was recently demonstrated that in vitro or in vivo activated CD8 cells express the Thpok gene [41,42]; this expression results in detectable Thpok protein expression and is important for the clonal expansion of CD8 effectors and their proper expression of cytotoxic enzymes upon rechallenge [41]. Thus, Cd4 and Thpok repression in CD8 cells appear to involve distinct mechanisms on which future studies will shed light.
Silencing mechanisms: facts and hypotheses
How epigenetic silencing is enforced remains enigmatic, and mechanistic investigations of Cd4 epigenetic silencing have yet to provide decisive clues. Imaging analyses of CD8-differentiating thymocytes have shown that Cd4 loci are relocated to nuclear heterochromatin, an area typically concentrating silenced genes [43,44]. Whether this is a cause or a consequence of Cd4 silencing remains to be determined; notably, Cd8 genes undergo a similar relocation to heterochromatin in CD4-differentiating thymocytes, even though they are not subject to the same epigenetic silencing as Cd4 (see below). In addition to subnuclear localization, covalent modifications of histones and DNA, namely methylation of histone H3 on lysine residues 9 and 27 (H3K9 and H3K27, respectively) and methylation of cytosines in CpG dinucleotides have been implicated in gene silencing [45-47]. Chromatin immunoprecipitation analyses have reported an enrichment in trimethyl H3K9 and H3K27 at the Cd4 promoter in CD8 cells relative to CD4−CD8− thymocytes (the precursors of DP thymocytes, in which transient silencer activity prevents premature Cd4 expression [30]). Because Cd4 is transiently repressed in CD4− CD8− thymocytes but irreversibly silenced in CD8 cells, this finding raised the hypothesis that silencing is caused by these histone modifications [48]. Multiple enzymes can contribute to H3K9 methylation, often in a stepwise fashion, complicating genetic analyses of this process; as a result, it is too early to formulate conclusions on its role in Cd4 silencing [49,50]. The role of H3K37 methylation has been evaluated by disrupting the gene encoding Ezh2, the enzymatic component of the Polycomb Repressive Complex 2 (PRC2), and main the H3K27 methyl transferase activity in T cells [51]. These analyses did not find any CD4 re-expression by Ezh2-deficient CD8 cells, although it cannot at present be excluded that this is due to redundancy with the related Ezh1 molecule [52,53] or to the extended half life of H3K27 trimethylation, that would long survive the disruption of the Ezh2 gene [54].
The role of DNA methylation in maintaining Cd4 silencing has been addressed genetically, in mice carrying a conditional allele of the gene encoding the DNA methyl transferase Dnmt1 that propagates CpG DNA methylation during cell division (but does not contribute to de novo DNA methylation) [55]. Disrupting Dnmt1 activity in mature T cells promotes expression of IL-4 in CD8 cells, but was not reported to promote their expression of CD4 [55,56]. Furthermore, inhibition of DNA methylation by the inhibitor 5 aza-deoxycytidine does not cause CD4 re-expression by CD8 cells made ‘silencer-less’ by post-thymic excision of the Cd4 silencer [37]. These results suggest that DNA methylation is not required to maintain Cd4 silencing. One limitation inherent to such experiments is that DNA methylation has pleiotropic effects, notably on cell proliferation and survival. Indeed, enforced expression of the anti-apoptotic molecule Bcl-xL rescues at least in part the development of thymocytes made Dnmt1-deficient before the DP stage of development [55]. As a result, it is conceivable that post-thymic cells made Dnmt1-deficient die prior to the point where removal of putative epigenetic marks on the Cd4 gene would result in re-expression.
In summary, although genetic studies offer strong support to the idea of epigenetic silencing of CD4-lineage genes in CD8 T cells, the mechanistic bases for such silencing remain unclear.
Post-thymic enforcement of commitment in CD4 cells: the case for plasticity
The Thpok-Runx3 circuitry in post-thymic CD4 cells
Do CD4 cells use similar epigenetic mechanisms to prevent expression of CD8-lineage genes ? Several observations suggest otherwise. First, although they do not re-express CD8, effectors CD4 cells can express genes typical of cytotoxic effectors such as perforin or granzyme B. While the expression of such genes is typically much lower than in CD8 effector cells, it is 100-1000 times higher than in naïve CD4 cells [19]. Furthermore, it can be induced in CD4 cells by enforced expression of Eomesodermin, a transcription factor important for the proper differentiation of CD8 cells into cytotoxic effector or memory cells [57-59], suggesting greater plasticity of gene expression in CD4 than in CD8 cells. Another important point came from studies of CD4 cell effector differentiation. CD4 T cells can adopt multiple effector profiles, characterized by specific cytokine production and transcription factor expression patterns (Fig. 1). Notably, they can differentiate into Th1 effectors that make IFNγ and promote responses against intra-cellular pathogens, or into Th2 cells (that express IL-4, IL-5 and IL-13 and are essential for defenses against multicellular parasites) [9]. Quite strikingly, Th1 effector differentiation requires Runx3, and differentiating Th1 effector express as much Runx3 protein as CD8 cells, despite continued expression of Thpok (and Cd4) [15,19,60,61].
Thus, whereas CD8 cells silence or keep at low level the expression of Thpok and genes typical of the CD4 lineage, at least some CD4 effector displays high-level expression of genes typical of the CD8 lineage. This raises two reciprocal questions, that we will now address: how do such Runx3-expressing CD4 cells maintain Cd4 and Thpok expression, and what role, if any, does Thpok have in post-thymic CD4 cells to control expression of Runx3 and CD8-lineage genes ?
First, why is it that expression of Runx3 in Th1 CD4 cells (or more generally enforced Runx3 expression in CD4 cells [34,62]) fails to repress Cd4, whereas it does so in DP thymocytes [34,63])? To some extent, similar issues are raised from the persistent Runx1 expression in CD4-lineage thymocytes and T cells. Analyses in cell lines, and in vivo, have shown that in cells that co-express Thpok and Runx3, the former counteracts the Cd4 silencing effects of Runx3 and thereby ensures Cd4 expression [40]. Two hypotheses have been advanced to account for this observation (Fig. 2). First, Thpok has been shown to bind the Cd4 silencer, and it has been proposed that such binding opposes the recruitment of Runx molecules or their ability to mediate repression [14]. Analyses of Runx1 binding to the silencer are consistent with this idea: although Runx1 is expressed in both DP thymocytes and CD4 T cells, it associates with the Cd4 silencer in DP thymocytes that do not express Thpok, but not in mature CD4 T cells that do [48]. Second, experiments in cell lines assessing the activity of a Cd4 reporter construct indicated that, paradoxically, the ability of Thpok to counteract Runx3-mediated Cd4 repression required the activity of histone deacetylases, a family of enzymes that contribute to gene repression [40]. This suggests that Thpok counteracts Runx-mediated repression by inhibiting the expression of so far unknown genes whose products would be required for Runx3 to repress Cd4, so that their absence in Thpok-expressing CD4 cells would make Cd4 insensitive to Runx. Further work will be needed to distinguish between these two possibilities, which are not mutually exclusive.
Results suggesting that the Cd4 locus is subject to positive epigenetic control during positive selection shed yet another light on the persistent Cd4 expression in Th1 effectors despite high levels of Runx3 [36]. Conditional deletion of the Cd4 ‘proximal enhancer’ required for Cd4 expression in thymocytes (Fig. 2) demonstrates that post-thymic deletion of the enhancer does not affect Cd4 expression by effector T cells, suggesting that an ‘open’ epigenetic status was installed in the thymus; it is possible, although speculative at this point, that in this new epigenetic status Cd4 is no longer sensitive to Runx3 expression, even in the absence of Thpok. This idea would fit with the observation that activated CD4 cells maintain Cd4 expression despite Thpok disruption [19] (see below). However, there is evidence for differential control of Cd4 in activated and resting T cells [36,64]. Thus, it is possible that Thpok, even if not needed to maintain Cd4 expression in Th1 effectors, is necessary to protect Cd4 expression from Runx1-mediated repression in resting CD4 cells [14].
The issues raised by the lack of Thpok repression in Runx3-expressing Th1 effectors are fundamentally similar to those we just encountered with Cd4. However, a few differences are worth noting and point to the possibilities the Cd4 and Thpok silencer may not be regulated through the same mechanisms. Specifically, chromatin immunoprecipitation analyses in resting cells indicate that Runx complexes bind the Thpok silencer in both CD4 and CD8 T cells [27], and it seems reasonable to propose that the same would be true in Th1 effectors. Thus, even though Thpok molecules bind the Thpok silencer [14], such binding does not prevent the recruitment of Runx complexes, but would counteract their repressive activity. This interesting perspective raises the question of how Thpok, which belongs to a family of proteins primarily characterized as transcriptional repressors [65], could act to antagonize the repressive activity or Runx3 at the Thpok (or Cd4) silencer. Thpok was recently shown to bind and serve as a substrate for the p300 acetyl-transferase co-activator, which conceivably could convert Thpok into a transcriptional activator. However, acetylation was reported to increased Thpok stability and repressive activity [66], and further analyses will be needed to investigate how Thpok could antagonize gene repression.
Thpok represses CD8-lineage gene expression in post-thymic CD4 cells
Thus, the ability of Runx molecules to repress Thpok (and Cd4) is developmental stage- and cell type-dependent. On the other side of the same coin, the expression of Runx3 and cytotoxic genes in Th1 effectors suggests that Thpok does not serve to repress these genes in post-thymic CD4 cells, and that the antagonism between Thpok and CD8-lineage genes is unique to developing thymocytes. In fact, that is not the case, as demonstrated by eliminating Thpok from mature CD4 cells that have developed as Thpok-sufficient, using Cre-induced deletion of a conditional Thpok allele [19]. The most striking result of these experiments was that post-thymic expression of Thpok is required to prevent CD8 re-expression[19, and L.W. and R.B., unpublished data]. Reciprocally, gain-of-function analyses show that enforced Thpok expression in CD8 cells represses Cd8 expression and notably targets the activity of the Cd8 E8(I) enhancer, an element preferentially active in mature CD8 cells [18,39,67,68]. The repression of Cd8 by Thpok fits with the idea that this factor, similar to the related proteins Bcl6, PLZF or LRF, would primarily serve as a transcriptional repressor, possibly by recruiting chromatin modifying complexes [65].
These initial observation in Thpok-deficient CD4 cells observations make two points. First, unlike Cd4 which in CD8 cells is epigenetically silenced independently of Runx3, active repression is critical to maintain Cd8 repression in CD4 cells. Current investigations are exploring the mechanistic bases of Cd8 repression, and notably whether it involves direct Thpok recruitment to Cd8 genes. Indeed, the second point is that such active repression relies at least in part on Thpok. Additional investigations, in Thpok-conditional mice and in mice carrying hypomorphic Thpok alleles, have shed light on the range and mechanisms of this repression. Cd8 genes are not the only Thpok targets in CD4 cells: Thpok-deficient CD4 T cells express Runx3 [14,19], and specifically the isoform produced from the Runx3 distal promoter that is normally characteristic of CD8 cells [23]. Their expression of perforin and granzyme B genes was increased relative to that of wild-type CD4 cells [14,19]. Such cytotoxic gene expression is Runx-dependent, as up-regulation of granzyme B in Thpok-deficient CD4 cells is inhibited by a dominant-negative version of Runx3 [19]. This suggests that a key function of Thpok in post-thymic CD4 cells is to repress Runx3, and this conclusion is further supported by preliminary observations that the Cd8 re-expression by Thpok-deficient CD4 cells is partly dependent on Runx3 (L.W. and R.B., unpublished results). However, these observations should not mask the importance of Runx3-independent mechanisms as wild-type Th1 effectors do not re-express CD8, despite their high Runx3 expression, indicative of an Runx3-independent Thpok repression of Cd8.
In vitro analyses suggest that Thpok-disruption in CD4 cells has functional consequences aside from Cd8 or cytotoxic gene expression. Because Runx3 promotes Th1 at the expense of Th2 differentiation [60,61], the inappropriate expression of Runx3 by Thpok-deficient CD4 cells raised the possibility that their functional differentiation would be skewed towards the Th1 fate. Indeed, Thpok-deficient CD4 cells preferentially give rise to Th1 effectors when activated in vitro in the absence of polarizing cytokine signals [19]. Such Th1 differentiation is dependent on Runx activity. In addition, it is associated with (and largely dependent on) increased expression of the transcription factor Eomesodermin [19], which normally contributes to expression of IFNγ and of the cytotoxic program in CD8 effectors [57-59]. In contrast, it does not require T-bet, a transcription factor related to Eomesodermin and necessary to the Th1 differentiation of wild-type CD4 cells [69,70] (R.B. and L.W., unpublished observations). Such Th1 skewing does not result from the loss of Th2-differentiating potential, as, unlike CD8 T cells, Thpok-deficient CD4 cells become Th2 effectors if activated in the presence of the proper Th2-polarizing cytokine environment [19]. This finding, together with the persistent expression of CD4 and Cd40lg by Thpok-deficient CD4 cells [19, Y.X. and R.B., manuscript in preparation], are consistent with the hypothesis that the key function of Thpok in CD4 cells is to prevent inappropriate expression of Runx3 and CD8-lineage genes, rather than to promote expression of CD4-lineage specific genes. By constraining Runx3 expression, Thpok would prevent ‘preemptive’ Th1 effector differentiation upon activation, and therefore keep open a wider range of effector differentiation potential for CD4 cells than there is for CD8 cells.
Why plasticity ?
The distinct mechanisms used by CD4 and CD8 cells to maintain lineage integrity correlate with their distinct effector differentiation potentials, specifically with the need for Runx3 during Th1 effector differentiation of CD4 cells. Whether such need reflects truly distinct biochemical ‘functions’ of Runx3 that cannot be performed by Runx1 (such as distinct sets of interaction partners), or simply distinct expression patterns (e.g. that Runx1 expression cannot reach levels needed for Th1 differentiation) remains to be determined. Regardless, the need for Runx3 during Th1 differentiation implies that CD4 T cells do not irreversibly silence Runx3 during intrathymic CD4-lineage differentiation. In fact, plasticity is a recurring theme during CD4 effector cell differentiation, as highlighted by recent studies on the ‘reversibility’ of CD4 effector fates [11].
Conversely, the presence of Runx3 molecules in Th1 CD4 effectors requires active mechanisms to counteract their repression of genes essential for CD4 cells, including Cd4 and Thpok, and its potential positive effect on Cd8 genes. Both in resting cells and during effector differentiation, Thpok is an essential contributor to such mechanisms, but it is not the only one. Notably, the transcription factor Gata3, which is essential for Th2 differentiation, counteracts the effects of Runx3 during the differentiation of Th2 cells and specifically prevents its activation of IFNγ and Eomes expression [71].
A similar situation where cells epigenetically silence genes that have become useless (or potentially detrimental) and actively repress those for which there is a future need is encountered at the lineage separation checkpoint between T and NK cells [72]. These two lineages share many properties, notably the cytotoxicity and effector cytokine production patterns common to NK and CD8 T cells. Thus, it was of particular interest that the transcriptional repressor Bcl11b, recently shown to be required for T cell lineage commitment in the thymus, notably serves to avoid ‘drifting’ towards NK-like gene expression [73-75] (even though it is also important for CD8 cells to undergo cytotoxic effector differentiation [76]). That is, disruption of Bcl11b in committed αβ T cells (from the DN3 stage to post-thymic cells) results not only in a reduced expression of T cell specific genes, but in the appearance of NK-specific genes and the generation of NK-like cells with cytotoxic function [75]. In contrast, although T cell precursors maintain the potential to revert to a myeloid fate until the Bcl11b-operated commitment checkpoint, disruption of Bcl11b in committed T cells did not give rise to re-expression of typical myeloid genes, including PU.1 and C/EBPα [73,74]. These findings are consistent with the possibility that T cells maintain an active and reversible repression of genes characteristic of the NK program, as some of these are part of the cytotoxic effector program used by CD8 effector T cells, whereas genes characteristic of the myeloid lineage would be epigenetically silenced.
Conclusions and perspectives
Three conclusions emerge from this comparison of processes that establish and maintain the divergence of CD4 and CD8 lineages. First, CD4 and CD8 cells use distinct mechanisms to maintain their lineage integrity, understood as the persistent repression of genes characteristic of the other lineage. In CD4 cells, the same molecule, Thpok, is instrumental to establish commitment (in the thymus) and maintain lineage integrity (in post-thymic cells). To the contrary, whereas CD8 cells use Runx3 to establish commitment in the thymus, they seem to rely on Runx3-independent epigenetic silencing to repress CD4-lineage genes. Second, the co-expression of Runx3 with Cd4 and Thpok in Th1 CD4 effectors indicates that, although CD4-lineage integrity is maintained by an active Thpok-based circuitry, this circuitry differs from that operating in CD4-differentiating thymocytes. Third, the control of Thpok and Cd4 expression is unlikely to follow similar rules as Thpok but not Cd4 is expressed by activated CD8 cells, whereas Cd4 but not Thpok is expressed in DP thymocytes. In addition, although both genes have silencer elements that recruit Runx proteins and can bind Thpok, Runx complexes bind the Thpok silencer in both immature thymocytes and mature T cells, whereas they bind the Cd4 silencer in preselection thymocytes but not in CD4 cells.
The opposition between active repression and epigenetic silencing should not be overemphasized, as epigenetic mechanisms and active repression can cooperate to control gene expression. To stay within the scope of this review, Cd8 repression in CD4 cells seems to epitomize such cooperation, as re-expression of Cd8 by Thpok-deficient cells is variegated, suggesting an additional layer of epigenetic control [19]. However, the fact that no Cd8 re-expression is observed in Thpok-sufficient cells indicates that such epigenetic control is subordinate to Thpok activity. More broadly, although there is evidence for self maintained epigenetic silencing or activation that has become independent of cis-regulatory elements that installed them in the first place, there is also evidence of ‘epigenetic’ marks that are removed in response to differentiation signals [77-79]. This suggests that in many circumstances there is an interplay between active transcription factors networks and epigenetic marks. One attractive possibility is that signals remove methylation marks by recruiting histone de-methylases [80-82], whereas in the absence of such signals methylation marks are self propagated.
Understanding which chromatin modifications promote epigenetic silencing and activation, and how they are installed in cells that undergo lineage commitment, are important challenges for current research. It is possible that transcriptional repressors recruit epigenetic marking complexes, including histone deacetylases and methylases, and thereby contribute to install epigenetic silencing [83], but studies in yeast and higher eukaryotes suggest additional mechanisms [84] Notably, the possibility that non-coding RNAs would be involved in gene silencing (and more generally in the control of gene expression) has recently attracted much interest [85,86]. Analyses in several systems, including X-chromosome inactivation, Hox gene expression and cell-cycle related genes have pointed to the ability of non-coding RNAs to repress transcription both in cis (on the gene they are transcribed from) and in trans [87,88]. Several mechanisms have been proposed to underlie such effects, including the repression of sense transcription by anti-sense transcription non-coding transcripts. In the case of the tumor-suppressor p15 gene, encoding a cell-cycle inhibitor, gain-of-function analyses suggest that non-coding anti-sense transcripts give rise to epigenetic silencing presumably mediated by heterochromatin formation [89]. Indeed, non-coding RNAs have been shown to recruit repressive or activating chromatin-modifying complexes [87], and future investigations will lift the veil on this important, yet poorly understood, aspect of epigenetic gene expression control.
Acknowledgments
We thank Yasmine Belkaid, Avinash Bhandoola and Jinfang Zhu for stimulating discussions. Research work in the authors’ laboratory is supported by the Intramural Research Program of the National Cancer Institute, Center for Cancer Research, NIH.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Singer A, Adoro S, Park JH. Lineage fate and intense debate: myths, models and mechanisms of CD4- versus CD8-lineage choice. Nat Rev Immunol. 2008;8:788–801. doi: 10.1038/nri2416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Wang L, Bosselut R. CD4-CD8 lineage differentiation: Thpok-ing into the nucleus. J Immunol. 2009;183:2903–2910. doi: 10.4049/jimmunol.0901041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Collins A, Littman DR, Taniuchi I. RUNX proteins in transcription factor networks that regulate T-cell lineage choice. Nat Rev Immunol. 2009;9:106–115. doi: 10.1038/nri2489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Corbella P, Moskophidis D, Spanopoulou E, Mamalaki C, Tolaini M, Itano A, Lans D, Baltimore D, Robey E, Kioussis D. Functional commitment to helper T cell lineage precedes positive selection and is independent of T cell receptor MHC specificity. Immunity. 1994;1:269–276. doi: 10.1016/1074-7613(94)90078-7. [DOI] [PubMed] [Google Scholar]
- 5.Matechak EO, Killeen N, Hedrick SM, Fowlkes BJ. MHC class II-specific T cells can develop in the CD8 lineage when CD4 is absent. Immunity. 1996;4:337–347. doi: 10.1016/s1074-7613(00)80247-2. [DOI] [PubMed] [Google Scholar]
- 6.Carpenter AC, Bosselut R. Decision checkpoints in the thymus. Nat Immunol. 2010;11:666–673. doi: 10.1038/ni.1887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Starr TK, Jameson SC, Hogquist KA. Positive and negative selection of T cells. Annu Rev Immunol. 2003;21:139–176. doi: 10.1146/annurev.immunol.21.120601.141107. [DOI] [PubMed] [Google Scholar]
- 8.von Boehmer H, Melchers F. Checkpoints in lymphocyte development and autoimmune disease. Nat Immunol. 2010;11:14–20. doi: 10.1038/ni.1794. [DOI] [PubMed] [Google Scholar]
- 9.Zhu J, Paul WE. CD4 T cells: fates, functions, and faults. Blood. 2008;112:1557–1569. doi: 10.1182/blood-2008-05-078154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Zhou L, Chong MM, Littman DR. Plasticity of CD4+ T cell lineage differentiation. Immunity. 2009;30:646–655. doi: 10.1016/j.immuni.2009.05.001. [DOI] [PubMed] [Google Scholar]
- 11.O’Shea JJ, Paul WE. Mechanisms underlying lineage commitment and plasticity of helper CD4+ T cells. Science. 2010;327:1098–1102. doi: 10.1126/science.1178334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.He X, He X, Dave VP, Zhang Y, Hua X, Nicolas E, Xu W, Roe BA, Kappes DJ. The zinc finger transcription factor Th-POK regulates CD4 versus CD8 T-cell lineage commitment. Nature. 2005;433:826–833. doi: 10.1038/nature03338. [DOI] [PubMed] [Google Scholar]
- 13.Wang L, Wildt KF, Zhu J, Zhang X, Feigenbaum L, Tessarollo L, Paul WE, Fowlkes BJ, Bosselut R. Distinct functions for the transcription factors GATA-3 and ThPOK during intrathymic differentiation of CD4(+) T cells. Nat Immunol. 2008;9:1122–1130. doi: 10.1038/ni.1647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Muroi S, Naoe Y, Miyamoto C, Akiyama K, Ikawa T, Masuda K, Kawamoto H, Taniuchi I. Cascading suppression of transcriptional silencers by ThPOK seals helper T cell fate. Nat Immunol. 2008;9:1113–1121. doi: 10.1038/ni.1650. [DOI] [PubMed] [Google Scholar]
- 15.Egawa T, Littman DR. ThPOK acts late in specification of the helper T cell lineage and suppresses Runx-mediated commitment to the cytotoxic T cell lineage. Nat Immunol. 2008;9:1131–1139. doi: 10.1038/ni.1652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Sun G, Liu X, Mercado P, Jenkinson SR, Kypriotou M, Feigenbaum L, Galera P, Bosselut R. The zinc finger protein cKrox directs CD4 lineage differentiation during intrathymic T cell positive selection. Nat Immunol. 2005;6:373–381. doi: 10.1038/ni1183. [DOI] [PubMed] [Google Scholar]
- 17.Keefe R, Dave V, Allman D, Wiest D, Kappes DJ. Regulation of lineage commitment distinct from positive selection. Science. 1999;286:1149–1153. doi: 10.1126/science.286.5442.1149. [DOI] [PubMed] [Google Scholar]
- 18.Jenkinson SR, Intlekofer AM, Sun G, Feigenbaum L, Reiner SL, Bosselut R. Expression of the transcription factor cKrox in peripheral CD8 T cells reveals substantial postthymic plasticity in CD4-CD8 lineage differentiation. J Exp Med. 2007;204:267–272. doi: 10.1084/jem.20061982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Wang L, Wildt KF, Castro E, Xiong Y, Feigenbaum L, Tessarollo L, Bosselut R. The zinc finger transcription factor Zbtb7b represses CD8-lineage gene expression in peripheral CD4+ T cells. Immunity. 2008;29:876–887. doi: 10.1016/j.immuni.2008.09.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Speck NA, Gilliland DG. Core-binding factors in haematopoiesis and leukaemia. Nat Rev Cancer. 2002;2:502–513. doi: 10.1038/nrc840. [DOI] [PubMed] [Google Scholar]
- 21.Lorsbach RB, Moore J, Ang SO, Sun W, Lenny N, Downing JR. Role of RUNX1 in adult hematopoiesis: analysis of RUNX1-IRES-GFP knock-in mice reveals differential lineage expression. Blood. 2004;103:2522–2529. doi: 10.1182/blood-2003-07-2439. [DOI] [PubMed] [Google Scholar]
- 22.Sato T, Ohno S, Hayashi T, Sato C, Kohu K, Satake M, Habu S. Dual functions of Runx proteins for reactivating CD8 and silencing CD4 at the commitment process into CD8 thymocytes. Immunity. 2005;22:317–328. doi: 10.1016/j.immuni.2005.01.012. [DOI] [PubMed] [Google Scholar]
- 23.Egawa T, Tillman RE, Naoe Y, Taniuchi I, Littman DR. The role of the Runx transcription factors in thymocyte differentiation and in homeostasis of naive T cells. J Exp Med. 2007;204:1945–1957. doi: 10.1084/jem.20070133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Taniuchi I, Osato M, Egawa T, Sunshine MJ, Bae SC, Komori T, Ito Y, Littman DR. Differential requirements for Runx proteins in CD4 repression and epigenetic silencing during T lymphocyte development. Cell. 2002;111:621–633. doi: 10.1016/s0092-8674(02)01111-x. [DOI] [PubMed] [Google Scholar]
- 25.Woolf E, Xiao C, Fainaru O, Lotem J, Rosen D, Negreanu V, Bernstein Y, Goldenberg D, Brenner O, Berke G, Levanon D, Groner Y. Runx3 and Runx1 are required for CD8 T cell development during thymopoiesis. Proc Natl Acad Sci U S A. 2003;100:7731–7736. doi: 10.1073/pnas.1232420100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Liu X, Bosselut R. Duration of TCR signaling controls CD4-CD8 lineage differentiation in vivo. Nat Immunol. 2004;5:280–288. doi: 10.1038/ni1040. [DOI] [PubMed] [Google Scholar]
- 27.Setoguchi R, Tachibana M, Naoe Y, Muroi S, Akiyama K, Tezuka C, Okuda T, Taniuchi I. Repression of the transcription factor Th-POK by Runx complexes in cytotoxic T cell development. Science. 2008;319:822–825. doi: 10.1126/science.1151844. [DOI] [PubMed] [Google Scholar]
- 28.Cruz-Guilloty F, Pipkin ME, Djuretic IM, Levanon D, Lotem J, Lichtenheld MG, Groner Y, Rao A. Runx3 and T-box proteins cooperate to establish the transcriptional program of effector CTLs. J Exp Med. 2009;206:51–59. doi: 10.1084/jem.20081242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Sawada S, Scarborough JD, Killeen N, Littman DR. A lineage-specific transcriptional silencer regulates CD4 gene expression during T lymphocyte development. Cell. 1994;77:917–929. doi: 10.1016/0092-8674(94)90140-6. [DOI] [PubMed] [Google Scholar]
- 30.Taniuchi I, Sunshine MJ, Festenstein R, Littman DR. Evidence for distinct CD4 silencer functions at different stages of thymocyte differentiation. Mol Cell. 2002;10:1083–1096. doi: 10.1016/s1097-2765(02)00735-9. [DOI] [PubMed] [Google Scholar]
- 31.He X, Park K, Wang H, He X, Zhang Y, Hua X, Li Y, Kappes DJ. CD4-CD8 lineage commitment is regulated by a silencer element at the ThPOK transcription-factor locus. Immunity. 2008;28:346–358. doi: 10.1016/j.immuni.2008.02.006. [DOI] [PubMed] [Google Scholar]
- 32.Monod J, Jacob F. Teleonomic mechanisms in cellular metabolism, growth, and differentiation. Cold Spring Harb Symp Quant Biol. 1961;26:389–401. doi: 10.1101/sqb.1961.026.01.048. [DOI] [PubMed] [Google Scholar]
- 33.Ferrell JEJ. Self-perpetuating states in signal transduction: positive feedback, double-negative feedback and bistability. Curr Opin Cell Biol. 2002;14:140–148. doi: 10.1016/s0955-0674(02)00314-9. [DOI] [PubMed] [Google Scholar]
- 34.Grueter B, Petter M, Egawa T, Laule-Kilian K, Aldrian CJ, Wuerch A, Ludwig Y, Fukuyama H, Wardemann H, Waldschuetz R, Moroy T, Taniuchi I, Steimle V, Littman DR, Ehlers M. Runx3 Regulates Integrin {alpha}E/CD103 and CD4 Expression during Development of CD4-/CD8+ T Cells. J Immunol. 2005;175:1694–1705. doi: 10.4049/jimmunol.175.3.1694. [DOI] [PubMed] [Google Scholar]
- 35.Sakaguchi S, Hombauer M, Bilic I, Naoe Y, Schebesta A, Taniuchi I, Ellmeier W. The zinc-finger protein MAZR is part of the transcription factor network that controls the CD4 versus CD8 lineage fate of double-positive thymocytes. Nat Immunol. 2010;11:442–448. doi: 10.1038/ni.1860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Chong MM, Simpson N, Ciofani M, Chen G, Collins A, Littman DR. Epigenetic propagation of CD4 expression is established by the Cd4 proximal enhancer in helper T cells. Genes Dev. 2010;24:659–669. doi: 10.1101/gad.1901610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Zou YR, Sunshine MJ, Taniuchi I, Hatam F, Killeen N, Littman DR. Epigenetic silencing of CD4 in T cells committed to the cytotoxic lineage. Nat Genet. 2001;29:332–336. doi: 10.1038/ng750. [DOI] [PubMed] [Google Scholar]
- 38.Leung RK, Thomson K, Gallimore A, Jones E, Van dB M, Sierro S, Alsheikhly AR, McMichael A, Rahemtulla A. Deletion of the CD4 silencer element supports a stochastic mechanism of thymocyte lineage commitment. Nat Immunol. 2001;2:1167–1173. doi: 10.1038/ni733. [DOI] [PubMed] [Google Scholar]
- 39.Taniuchi I, Ellmeier W, Littman DR. The CD4/CD8 lineage choice: new insights into epigenetic regulation during T cell development. Adv Immunol. 2004;83:55–89. doi: 10.1016/S0065-2776(04)83002-5. [DOI] [PubMed] [Google Scholar]
- 40.Wildt KF, Sun G, Grueter B, Fischer M, Zamisch M, Ehlers M, Bosselut R. The transcription factor zbtb7b promotes CD4 expression by antagonizing runx-mediated activation of the CD4 silencer. J Immunol. 2007;179:4405–4414. doi: 10.4049/jimmunol.179.7.4405. [DOI] [PubMed] [Google Scholar]
- 41.Setoguchi R, Taniuchi I, Bevan MJ. ThPOK derepression is required for robust CD8 T cell responses to viral infection. J Immunol. 2009;183:4467–4474. doi: 10.4049/jimmunol.0901428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Wang L, Carr T, Xiong Y, Wildt KF, Zhu J, Feigenbaum L, Bendelac A, Bosselut R. The sequential activity of Gata3 and Thpok is required for the differentiation of CD1d-restricted CD4(+) NKT cells. Eur J Immunol. 2010;40:2385–2390. doi: 10.1002/eji.201040534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Merkenschlager M, Amoils S, Roldan E, Rahemtulla A, O’connor E, Fisher AG, Brown KE. Centromeric repositioning of coreceptor loci predicts their stable silencing and the CD4/CD8 lineage choice. J Exp Med. 2004;200:1437–1444. doi: 10.1084/jem.20041127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Delaire S, Huang YH, Chan SW, Robey EA. Dynamic repositioning of CD4 and CD8 genes during T cell development. J Exp Med. 2004;200:1427–1435. doi: 10.1084/jem.20041041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Wilson CB, Makar KW, Shnyreva M, Fitzpatrick DR. DNA methylation and the expanding epigenetics of T cell lineage commitment. Semin Immunol. 2005;17:105–119. doi: 10.1016/j.smim.2005.01.005. [DOI] [PubMed] [Google Scholar]
- 46.Barski A, Cuddapah S, Cui K, Roh TY, Schones DE, Wang Z, Wei G, Chepelev I, Zhao K. High-resolution profiling of histone methylations in the human genome. Cell. 2007;129:823–837. doi: 10.1016/j.cell.2007.05.009. [DOI] [PubMed] [Google Scholar]
- 47.Wilson CB, Rowell E, Sekimata M. Epigenetic control of T-helper-cell differentiation. Nat Rev Immunol. 2009;9:91–105. doi: 10.1038/nri2487. [DOI] [PubMed] [Google Scholar]
- 48.Yu M, Wan M, Zhang J, Wu J, Khatri R, Chi T. Nucleoprotein structure of the CD4 locus: implications for the mechanisms underlying CD4 regulation during T cell development. Proc Natl Acad Sci U S A. 2008;105:3873–3878. doi: 10.1073/pnas.0800810105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Lehnertz B, Northrop JP, Antignano F, Burrows K, Hadidi S, Mullaly SC, Rossi FM, Zaph C. Activating and inhibitory functions for the histone lysine methyltransferase G9a in T helper cell differentiation and function. J Exp Med. 2010;207:915–922. doi: 10.1084/jem.20100363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Thomas LR, Miyashita H, Cobb RM, Pierce S, Tachibana M, Hobeika E, Reth M, Shinkai Y, Oltz EM. Functional analysis of histone methyltransferase g9a in B and T lymphocytes. J Immunol. 2008;181:485–493. doi: 10.4049/jimmunol.181.1.485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Su IH, Dobenecker MW, Dickinson E, Oser M, Basavaraj A, Marqueron R, Viale A, Reinberg D, Wulfing C, Tarakhovsky A. Polycomb group protein ezh2 controls actin polymerization and cell signaling. Cell. 2005;121:425–436. doi: 10.1016/j.cell.2005.02.029. [DOI] [PubMed] [Google Scholar]
- 52.Margueron R, Li G, Sarma K, Blais A, Zavadil J, Woodcock CL, Dynlacht BD, Reinberg D. Ezh1 and Ezh2 maintain repressive chromatin through different mechanisms. Mol Cell. 2008;32:503–518. doi: 10.1016/j.molcel.2008.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Shen X, Liu Y, Hsu YJ, Fujiwara Y, Kim J, Mao X, Yuan GC, Orkin SH. EZH1 mediates methylation on histone H3 lysine 27 and complements EZH2 in maintaining stem cell identity and executing pluripotency. Mol Cell. 2008;32:491–502. doi: 10.1016/j.molcel.2008.10.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Tarakhovsky A. Tools and landscapes of epigenetics. Nat Immunol. 2010;11:565–568. doi: 10.1038/ni0710-565. [DOI] [PubMed] [Google Scholar]
- 55.Lee PP, Fitzpatrick DR, Beard C, Jessup HK, Lehar S, Makar KW, Perez-Melgosa M, Sweetser MT, Schlissel MS, Nguyen S, Cherry SR, Tsai JH, Tucker SM, Weaver WM, Kelso A, Jaenisch R, Wilson CB. A critical role for Dnmt1 and DNA methylation in T cell development, function, and survival. Immunity. 2001;15:763–774. doi: 10.1016/s1074-7613(01)00227-8. [DOI] [PubMed] [Google Scholar]
- 56.Makar KW, Perez-Melgosa M, Shnyreva M, Weaver WM, Fitzpatrick DR, Wilson CB. Active recruitment of DNA methyltransferases regulates interleukin 4 in thymocytes and T cells. Nat Immunol. 2003;4:1183–1190. doi: 10.1038/ni1004. [DOI] [PubMed] [Google Scholar]
- 57.Pearce EL, Mullen AC, Martins GA, Krawczyk CM, Hutchins AS, Zediak VP, Banica M, DiCioccio CB, Gross DA, Mao CA, Shen H, Cereb N, Yang SY, Lindsten T, Rossant J, Hunter CA, Reiner SL. Control of effector CD8+ T cell function by the transcription factor Eomesodermin. Science. 2003;302:1041–1043. doi: 10.1126/science.1090148. [DOI] [PubMed] [Google Scholar]
- 58.Intlekofer AM, Takemoto N, Wherry EJ, Longworth SA, Northrup JT, Palanivel VR, Mullen AC, Gasink CR, Kaech SM, Miller JD, Gapin L, Ryan K, Russ AP, Lindsten T, Orange JS, Goldrath AW, Ahmed R, Reiner SL. Effector and memory CD8+ T cell fate coupled by T-bet and eomesodermin. Nat Immunol. 2005;6:1236–1244. doi: 10.1038/ni1268. [DOI] [PubMed] [Google Scholar]
- 59.Intlekofer AM, Banerjee A, Takemoto N, Gordon SM, Dejong CS, Shin H, Hunter CA, Wherry EJ, Lindsten T, Reiner SL. Anomalous type 17 response to viral infection by CD8+ T cells lacking T-bet and eomesodermin. Science. 2008;321:408–411. doi: 10.1126/science.1159806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Djuretic IM, Levanon D, Negreanu V, Groner Y, Rao A, Ansel KM. Transcription factors T-bet and Runx3 cooperate to activate Ifng and silence Il4 in T helper type 1 cells. Nat Immunol. 2007;8:145–153. doi: 10.1038/ni1424. [DOI] [PubMed] [Google Scholar]
- 61.Naoe Y, Setoguchi R, Akiyama K, Muroi S, Kuroda M, Hatam F, Littman DR, Taniuchi I. Repression of interleukin-4 in T helper type 1 cells by Runx/Cbf beta binding to the Il4 silencer. J Exp Med. 2007;204:1749–1755. doi: 10.1084/jem.20062456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Telfer JC, Hedblom EE, Anderson MK, Laurent MN, Rothenberg EV. Localization of the domains in Runx transcription factors required for the repression of CD4 in thymocytes. J Immunol. 2004;172:4359–4370. doi: 10.4049/jimmunol.172.7.4359. [DOI] [PubMed] [Google Scholar]
- 63.Kohu K, Sato T, Ohno S, Hayashi K, Uchino R, Abe N, Nakazato M, Yoshida N, Kikuchi T, Iwakura Y, Inoue Y, Watanabe T, Habu S, Satake M. Overexpression of the Runx3 transcription factor increases the proportion of mature thymocytes of the CD8 single-positive lineage. J Immunol. 2005;174:2627–2636. doi: 10.4049/jimmunol.174.5.2627. [DOI] [PubMed] [Google Scholar]
- 64.Manjunath N, Shankar P, Stockton B, Dubey PD, Lieberman J, von Andrian UH. A transgenic mouse model to analyze CD8(+) effector T cell differentiation in vivo. Proc Natl Acad Sci U S A. 1999;96:13932–13937. doi: 10.1073/pnas.96.24.13932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Bilic I, Ellmeier W. The role of BTB domain-containing zinc finger proteins in T cell development and function. Immunol Lett. 2007;108:1–9. doi: 10.1016/j.imlet.2006.09.007. [DOI] [PubMed] [Google Scholar]
- 66.Zhang M, Zhang J, Rui J, Liu X. p300-mediated acetylation stabilizes the Th-inducing POK factor. J Immunol. 2010;185:3960–3969. doi: 10.4049/jimmunol.1001462. [DOI] [PubMed] [Google Scholar]
- 67.Hostert A, Tolaini M, Roderick K, Harker N, Norton T, Kioussis D. A region in the CD8 gene locus that directs expression to the mature CD8 T cell subset in transgenic mice. Immunity. 1997;7:525–536. doi: 10.1016/s1074-7613(00)80374-x. [DOI] [PubMed] [Google Scholar]
- 68.Ellmeier W, Sunshine MJ, Losos K, Hatam F, Littman DR. An enhancer that directs lineage-specific expression of CD8 in positively selected thymocytes and mature T cells. Immunity. 1997;7:537–547. doi: 10.1016/s1074-7613(00)80375-1. [DOI] [PubMed] [Google Scholar]
- 69.Szabo SJ, Sullivan BM, Peng SL, Glimcher LH. Molecular mechanisms regulating Th1 immune responses. Annu Rev Immunol. 2003;21:713–758. doi: 10.1146/annurev.immunol.21.120601.140942. [DOI] [PubMed] [Google Scholar]
- 70.Szabo SJ, Sullivan BM, Stemmann C, Satoskar AR, Sleckman BP, Glimcher LH. Distinct effects of T-bet in TH1 lineage commitment and IFN-gamma production in CD4 and CD8 T cells. Science. 2002;295:338–342. doi: 10.1126/science.1065543. [DOI] [PubMed] [Google Scholar]
- 71.Yagi R, Junttila IS, Wei G, Urban JFJ, Zhao K, Paul WE, Zhu J. The transcription factor GATA3 actively represses RUNX3 protein-regulated production of interferon-gamma. Immunity. 2010;32:507–517. doi: 10.1016/j.immuni.2010.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Klein Wolterink RG, Garcia-Ojeda ME, Vosshenrich CA, Hendriks RW, Di Santo JP. The intrathymic crossroads of T and NK cell differentiation. Immunol Rev. 2010;238:126–137. doi: 10.1111/j.1600-065X.2010.00960.x. [DOI] [PubMed] [Google Scholar]
- 73.Ikawa T, Hirose S, Masuda K, Kakugawa K, Satoh R, Shibano-Satoh A, Kominami R, Katsura Y, Kawamoto H. An essential developmental checkpoint for production of the T cell lineage. Science. 2010;329:93–96. doi: 10.1126/science.1188995. [DOI] [PubMed] [Google Scholar]
- 74.Li L, Leid M, Rothenberg EV. An early T cell lineage commitment checkpoint dependent on the transcription factor Bcl11b. Science. 2010;329:89–93. doi: 10.1126/science.1188989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Li P, Burke S, Wang J, Chen X, Ortiz M, Lee SC, Lu D, Campos L, Goulding D, Ng BL, Dougan G, Huntly B, Gottgens B, Jenkins NA, Copeland NG, Colucci F, Liu P. Reprogramming of T cells to natural killer-like cells upon Bcl11b deletion. Science. 2010;329:85–89. doi: 10.1126/science.1188063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Zhang S, Rozell M, Verma RK, Albu DI, Califano D, VanValkenburgh J, Merchant A, Rangel-Moreno J, Randall TD, Jenkins NA, Copeland NG, Liu P, Avram D. Antigen-specific clonal expansion and cytolytic effector function of CD8+ T lymphocytes depend on the transcription factor Bcl11b. J Exp Med. 2010;207:1687–1699. doi: 10.1084/jem.20092136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Wei G, Wei L, Zhu J, Zang C, Hu-Li J, Yao Z, Cui K, Kanno Y, Roh TY, Watford WT, Schones DE, Peng W, Sun HW, Paul WE, O’Shea JJ, Zhao K. Global mapping of H3K4me3 and H3K27me3 reveals specificity and plasticity in lineage fate determination of differentiating CD4+ T cells. Immunity. 2009;30:155–167. doi: 10.1016/j.immuni.2008.12.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.De Santa F, Narang V, Yap ZH, Tusi BK, Burgold T, Austenaa L, Bucci G, Caganova M, Notarbartolo S, Casola S, Testa G, Sung WK, Wei CL, Natoli G. Jmjd3 contributes to the control of gene expression in LPS-activated macrophages. EMBO J. 2009 doi: 10.1038/emboj.2009.271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Barradas M, Anderton E, Acosta JC, Li S, Banito A, Rodriguez-Niedenfuhr M, Maertens G, Banck M, Zhou MM, Walsh MJ, Peters G, Gil J. Histone demethylase JMJD3 contributes to epigenetic control of INK4a/ARF by oncogenic RAS. Genes Dev. 2009;23:1177–1182. doi: 10.1101/gad.511109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Lan F, Nottke AC, Shi Y. Mechanisms involved in the regulation of histone lysine demethylases. Curr Opin Cell Biol. 2008;20:316–325. doi: 10.1016/j.ceb.2008.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Miller SA, Weinmann AS. Molecular mechanisms by which T-bet regulates T-helper cell commitment. Immunol Rev. 2010;238:233–246. doi: 10.1111/j.1600-065X.2010.00952.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Satoh T, Takeuchi O, Vandenbon A, Yasuda K, Tanaka Y, Kumagai Y, Miyake T, Matsushita K, Okazaki T, Saitoh T, Honma K, Matsuyama T, Yui K, Tsujimura T, Standley DM, Nakanishi K, Nakai K, Akira S. The Jmjd3-Irf4 axis regulates M2 macrophage polarization and host responses against helminth infection. Nat Immunol. 2010;11:936–944. doi: 10.1038/ni.1920. [DOI] [PubMed] [Google Scholar]
- 83.Reed-Inderbitzin E, Moreno-Miralles I, Vanden-Eynden SK, Xie J, Lutterbach B, Durst-Goodwin KL, Luce KS, Irvin BJ, Cleary ML, Brandt SJ, Hiebert SW. RUNX1 associates with histone deacetylases and SUV39H1 to repress transcription. Oncogene. 2006;25:5777–5786. doi: 10.1038/sj.onc.1209591. [DOI] [PubMed] [Google Scholar]
- 84.Grewal SI. RNAi-dependent formation of heterochromatin and its diverse functions. Curr Opin Genet Dev. 2010;20:134–141. doi: 10.1016/j.gde.2010.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Wilusz JE, Sunwoo H, Spector DL. Long noncoding RNAs: functional surprises from the RNA world. Genes Dev. 2009;23:1494–1504. doi: 10.1101/gad.1800909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Hung T, Chang HY. Long noncoding RNA in genome regulation: Prospects and mechanisms. RNA Biol. 2010;7 doi: 10.4161/rna.7.5.13216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Tsai MC, Manor O, Wan Y, Mosammaparast N, Wang JK, Lan F, Shi Y, Segal E, Chang HY. Long noncoding RNA as modular scaffold of histone modification complexes. Science. 2010;329:689–693. doi: 10.1126/science.1192002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Gupta RA, Shah N, Wang KC, Kim J, Horlings HM, Wong DJ, Tsai MC, Hung T, Argani P, Rinn JL, Wang Y, Brzoska P, Kong B, Li R, West RB, van de Vijver MJ, Sukumar S, Chang HY. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature. 2010;464:1071–1076. doi: 10.1038/nature08975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Yu W, Gius D, Onyango P, Muldoon-Jacobs K, Karp J, Feinberg AP, Cui H. Epigenetic silencing of tumour suppressor gene p15 by its antisense RNA. Nature. 2008;451:202–206. doi: 10.1038/nature06468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Rothenberg EV, Moore JE, Yui MA. Launching the T-cell-lineage developmental programme. Nat Rev Immunol. 2008;8:9–21. doi: 10.1038/nri2232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Chi AW, Bell JJ, Zlotoff DA, Bhandoola A. Untangling the T branch of the hematopoiesis tree. Curr Opin Immunol. 2009;21:121–126. doi: 10.1016/j.coi.2009.01.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Kondo T, Takata H, Matsuki F, Takiguchi M. Cutting edge: Phenotypic characterization and differentiation of human CD8+ T cells producing IL-17. J Immunol. 2009;182:1794–1798. doi: 10.4049/jimmunol.0801347. [DOI] [PubMed] [Google Scholar]
- 93.Yen HR, Harris TJ, Wada S, Grosso JF, Getnet D, Goldberg MV, Liang KL, Bruno TC, Pyle KJ, Chan SL, Anders RA, Trimble CL, Adler AJ, Lin TY, Pardoll DM, Huang CT, Drake CG. Tc17 CD8 T cells: functional plasticity and subset diversity. J Immunol. 2009;183:7161–7168. doi: 10.4049/jimmunol.0900368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Curtis MM, Way SS, Wilson CB. IL-23 promotes the production of IL-17 by antigen-specific CD8 T cells in the absence of IL-12 and type-I interferons. J Immunol. 2009;183:381–387. doi: 10.4049/jimmunol.0900939. [DOI] [PMC free article] [PubMed] [Google Scholar]