

Summary
Sigma receptors are nonopiate and nonphencyclidine binding sites that are thought to be neuroprotective due to modulation of NMDA receptors. Sigma receptor 1 expression has been demonstrated in numerous tissues including brain. Recently, studies using binding assays have demonstrated sigma receptor 1 in neural retina, however these studies did not demonstrate in which retinal cell type(s) sigma receptor 1 was present nor did they establish unequivocally the molecular identity of the receptor. The present study was designed to address these issues. RT-PCR analysis amplified sigma receptor 1 in neural retina, RPE/choroid complex, and lens isolated from mice. A similar RT-PCR product was amplified also in three cultured cell lines, rat Müller cells, rat ganglion cells and human ARPE-19 cells. In situ hybridization analysis revealed abundant sigma receptor 1 expression in ganglion cells, cells of the inner nuclear layer, inner segments of photoreceptor cells and retinal pigment epithelial (RPE) cells. Immunohistochemical studies detected the sigma receptor 1 protein in retinal ganglion, photoreceptor, RPE cells and surrounding the soma of cells in the inner nuclear layer. These data provide the first cellular localization of sigma receptor 1 in neural retina and establish the molecular identity of sigma receptor 1 in retinal cells. The demonstration that sigma receptor 1 is present in ganglion cells is particularly noteworthy given the well-documented susceptibility of these cells to glutamate toxicity. Our findings suggest that retinal ganglion cells may be amenable to the neuroprotective effects of sigma ligands under conditions of neurotoxicity such as occurs in diabetes.
Keywords: sigma receptor, ganglion cells, retina, RPE, ciliary body
Introduction
Sigma receptors are defined as nonopiate and nonphencyclidine binding sites that mediate the psychotomimetic actions of certain opioid derivatives [10,35]. Sigma receptors consist of several subtypes that are distinguishable by biochemical and pharmacological means [24]. Among these, the type 1 sigma receptor (sigma receptor 1) is the best characterized. The cDNA encoding sigma receptor 1 was cloned originally from guinea pig liver by Hanner and colleagues [12] and more recently from human, mouse and rat [11,15,23,31,32] in our laboratory. The sigma receptor 1 cDNA predicts a protein of 223 amino acids with a single putative transmembrane domain [11,12,15,23,24,31,32]. Sigma receptor 1 is believed to mediate the immunosuppressant, antipsychotic and neuroprotective effects elicited by sigma ligands such as haloperidol, ditolylguanidine and pentazocine [10,35]. Striking neuroprotective effects have been demonstrated with compounds such as ifenprodil, which has a high affinity for the sigma receptor 1 and produces complete protection against hypoxia-induced damage to neurons [14]. The neuroprotective effects of sigma receptor ligands are thought to be due to modulation of NMDA receptors as well as muscarinic receptors [4,21]. Lobner and Lipton [19] have shown that glutamate release from the hippocampus during ischemic insult is attenuated also by sigma receptor ligands suggesting additional mechanisms for the sigma receptor ligand-mediated neuroprotection against glutamate toxicity.
Sigma receptor expression has been demonstrated in a number of tissues (e.g. liver, spleen, gastrointestinal tract, adrenal gland, testis, ovary, placenta) [10,35]. Recently, the expression pattern of sigma receptor 1 was characterized in brain. In situ hybridization studies demonstrated that sigma receptor 1 mRNA is detectable primarily in the cerebral cortex, hippocampus, and Purkinje cells of the cerebellum [16,31]. It was suggested that the localization of sigma receptor 1 to these cells may be useful in modulation of sigma receptor 1-related brain functions. Immunohistochemical studies of sigma receptor in rat brain demonstrated high levels of immunostaining associated with neurons located in the granular layer of the olfactory bulb, various hypothalamic nuclei, the septum, the central gray matter, motor nuclei of the hindbrain and the dorsal horn of the spinal cord [1]. Less is known about the expression of sigma receptor 1 in the eye. Schoenwald et al [27] detected sigma receptor 1 in lacrimocytes isolated from rabbit lacrimal gland using binding assays. More recently, Bucolo et al [6] used binding assays to demonstrate the presence of sigma receptor 1 in iris-ciliary body isolated from rabbit. Their findings were particularly important as they showed a decrease of intraocular pressure when the sigma receptor ligands pentazocine and (+) NANM (N-allylnormetazocine) were applied topically. In retina, binding assays have demonstrated the presence of sigma receptor in bovine [28] and rat retina [29,37]. The density of sigma receptor was higher in retina than in adrenal medulla, lacrimocyte and brain. These investigators indicated that the retina has the highest density of sigma receptor 1 in central and peripheral tissue suggesting an important function for these receptors. There is evidence that amacrine cells express sigma receptor 1 based on observations that pentazocine and SA4503, two sigma receptor 1 agonists conferred protective effects against glutamate-induced damage of cultured amacrine cells [29]. It has not been demonstrated in which other cell types of the retina sigma receptor 1 is expressed. Given that sigma receptor 1 may offer neuroprotection by modulation of NMDA receptors and that ganglion cells are particularly vulnerable to glutamate toxicity [36], we were interested in determining whether ganglion cells, as well as other retinal cells, express sigma receptor 1. We addressed this question using molecular and immunohistochemical methods in intact retinal tissue from mice and in cells lines of three types of retinal cells (ganglion, Müller and epithelial cells). Our RT-PCR and in situ hybridization data show for the first time that sigma receptor 1 is expressed abundantly in ganglion cells in vivo and in vitro. Sigma receptor 1 is present also in Müller cells and retinal pigment epithelial (RPE) cells. Sigma receptor 1 mRNA is expressed in inner segments of photoreceptor cells and the protein is present in the cell soma. The presence of sigma receptor 1 in retina, particularly in ganglion cells, suggests that these cells may be amenable to the neuroprotective effects of sigma ligands under conditions of neurotoxicity such as occurs in diabetes [18].
Materials and methods
Reagents
RPMI 1640 medium, TRIzol reagent, and penicillin-streptomycin were purchased from Life Technologies (Rockville, MD); fetal bovine serum from Sigma (St. Louis, MO). ARPE-19, a spontaneously arising human cell line established in the laboratory of Dr. Larry Hjelmeland (University of California, Davis), were obtained from American Type Culture Collection (Manassas, VA). The development of the rat ganglion cell line [17] and the rat Müller cell line have been described [25]. RNAWIZ reagent was purchased from Ambion (Austin, TX). RNA PCR core kit was from Perkin-Elmer (Boston, MA). Tissue-Tek OCT embedding compound was from Miles Laboratories (Elkhart, IN).
Cell Culture
The three cell types used in these studies, ARPE-19, ganglion and Müller cells were maintained at 37°C in a humidified chamber of 5% CO2. They were cultured in 75 cm2 flasks with Dulbecco's modified Eagle's medium:nutrient mixture F12 (DMEM:F12), supplemented with 10% fetal bovine serum, 100 U/ml penicillin, and 100 μg/ml streptomycin. The culture medium was replaced with fresh medium every other day. Upon confluency, cultures were passaged by dissociation in 0.05% (w/v) trypsin in phosphate-buffered saline (PBS).
Animals
C57BL/6 mice and albino ICR mice (Harlan-Sprague Dawley) were used in these experiments. The C57BL/6 pigmented mice were used for studies in which the retina was dissected from the RPE for RT-PCR analysis; the albino mice were used for the in situ hybridization and immunohistochemistry studies so that no pigment in the RPE layer would interfere with detection of positive colorometric signals. Mice were maintained in clear plastic cages and subjected to standard light cycles (12 hr light/12 hr dark). Light levels measured from the bottom of cages ranged from 1.2 to 1.5 foot candles (12.9 – 16.1 1x). Room temperature was 23 ± 1°C. Mice were fed Harlan's Teklad rodent diet #8604 (min. crude protein, 24.0%; min. crude fat, 4.0%; max. crude fiber, 4.5%). Care and use of the animals adhered to the principles set forth in the DHEW Publication, NIH 80-23, “The Guiding Principles in the Care and Use of Animals.”
RT-PCR analysis of sigma receptor 1 mRNA in mouse retina, RPE, lens and brain
C57BL/6 mice were killed by CO2 asphyxiation. The retina, lens and RPE/eyecup were dissected immediately from the animals following our published procedure [34]. Briefly, the eye was proptosed and the cornea slit which immediately released the lens (and vitreous). The retina was removed from the remaining RPE/choroid/eyecup complex. Tissues from six eyes were pooled for each analysis. Total RNA was isolated using the RNAWIZ reagent. Mouse brain was used as a positive control [31]. For analysis of brain, the frontal and a portion of the parietal lobes of mouse brain were taken for preparation of RNA using the TRIzol reagent. RT-PCR was carried out with RNA isolated from each of these tissues using primer pairs specific for mouse sigma receptor 1. The upstream primer 5′-TATCGCAGTGCTGATCCA-3′ and the downstream primer 5′-TACTCCACCATCCACGTGTT-3′ correspond to nucleotide positions 75 to 92 and 520 to 539, respectively, in the cloned mouse sigma receptor 1 cDNA [32]. PCR following reverse transcription was carried out for 32 cycles. The products were size-fractionated on an agarose gel. The expected size of the PCR product, predicted from the positions of the primers is 465 bp. The resulting RT-PCR products were used for restriction site analysis using PstI and StyI. The expected sizes of the digestion products were 280 and 185 bp for PstI and 285 and 180 for StyI.
RT-PCR analysis of sigma receptor 1 mRNA in rat Müller and ganglion cells
Total RNA was prepared from rat Müller and ganglion cells using the TRIzol reagent. RT-PCR was carried out using primer pairs specific for rat sigma receptor 1: 5′-GTTTCTGACTATTGTGGCGGTGCTG-3′ (sense) and 5′-CAAATGCCAGGGTAGACGGAATAAC-3′ (antisense) corresponding to nucleotide positions 80-104 and 567-591, respectively of the cloned rat sigma receptor 1 cDNA [30]. RT-PCR was carried out for 35 cycles. The products were size fractionated on an agarose gel. The expected size of the RT-PCR product, predicted from positions of the primers is 512 bp. The resulting RT-PCR products were used for restriction site analysis using PstI and NarI. The expected sizes of the digestion products were 315 and 197 bp for PstI and 282 and 230 for NarI.
RT-PCR analysis of sigma receptor 1 mRNA in human ARPE-19 cells
Total RNA was prepared from ARPE-19 cells using the TRIzol reagent. RT-PCR was carried out using primer pairs specific for human sigma receptor 1: 5′-AGCGCGAAGAGATAGC-3′ (sense) and 5′-AGCATAGGAGCGAAGAGT-3′ (antisense) corresponding to nucleotide positions 160-175 and 651-668, respectively, of the human sigma receptor 1 cDNA [15]. The RT-PCR was carried out for 32 cycles, with the denaturing phase of 30 sec at 94°C, annealing phase of 30 sec at 57.9°C, and an extension of 2 min at 72°C. The products were size fractionated on an agarose gel. The expected size of the RT-PCR product, predicted from positions of the primers is 509 bp. The resulting RT-PCR products were used for restriction site analysis using PstI and NarI. The expected sizes of the digestion products were 383 and 126 bp for PstI and 350 and 159 for NarI.
In situ hybridization
To localize the mRNA transcript encoding sigma receptor 1 in retina and other ocular tissues, in situ hybridization was performed on mouse eyes. For the preparation of the mouse sigma receptor 1-specific riboprobe, a 0.65 kbp fragment of the mouse sigma receptor 1 cDNA, obtained by the digestion of pSPORT mouse sigma receptor 1 cDNA by SalI/Sma I, was subcloned into pBluescript vector. The orientation of the cDNA insert in the pBluescript vector was determined by sequencing. Antisense and sense riboprobes were synthesized with T7 RNA polymerase or T3 RNA polymerase after linerization of the plasmid with appropriate restriction enzymes. The riboprobes were labeled using a digoxigenin-labeling kit (Boehringer-Mannheim, Indianapolis, IN, USA). Additional controls for the in situ hybridization included sense and antisense riboprobes specific for the housekeeping gene glyceraldehyde 3-phosphate dehydrogenase (GAPDH), as described previously [7].
Eyes from albino mice were enucleated, frozen immediately in Tissue-Tek OCT sectioned at 10 μm thickness, and fixed in 4% paraformaldehyde. Following our published protocol [7], sections were rinsed in ice-cold PBS and treated with active 1% diethylpyrocarbonate prepared in PBS to facilitate penetration of the labeled probes. Sections were permeabilized further with proteinase K (1 μg/ml) in PBS for 4 minutes. The proteinase K activity was stopped by rinsing the slides in glycine (2 mg/ml) in PBS. Sections were washed in PBS, equilibrated in 5 × SSC, and were prehybridized for 2 h at 58°C in 50% (v/v) formamide, 5 × SSC, 2% (w/v) blocking reagent (provided with the DIG Nucleic Acid Detection Kit), 0.1% wt/vol N-lauroylsarcosine, and 0.02% (w/v) sodium dodecyl sulfate. Sections were hybridized with the probes (1 μg/ml) and were incubated overnight at 58°C. They were washed twice in 2 × SSC at room temperature, twice in 1 × SSC at 55°C, and twice in 0.1 × SSC at 37°C. For immunologic detection of the probe, sections were washed in a buffer containing 0.1 M maleic acid and 0.15 M NaCl (pH 7.5) and were blocked with the same buffer containing 1% blocking reagent. The anti-DIG-AP was diluted 1:5000, and slides were incubated with this antibody for 2 h at room temperature. Sections were washed in the preceding wash buffer containing levamisol (200 μg/ml) twice for 10 minutes and were equilibrated with a buffer containing 100 mM Tris-HCl (pH 9.5), 100 mM NaCl, and 50 mM MgCl2. The color reaction was developed in NBT/BCIP. Slides were washed in distilled water coverslipped, but not counterstained so that the purplish-red colored precipitate, indicative of a positive reaction, could be visualized in the sections.
Immunohistochemistry
Immunohistochemical methods were used to localize sigma receptor 1 in retina of intact mouse eyes. Cryosections of mouse eyes were fixed with ice-cold acetone, blocked with 10% normal goat serum and were incubated 3 h at room temperature with the sigma receptor 1 specific monoclonal antibody [13] at a dilution of 1:50 followed by an overnight incubation at 4°C. Incubation with 0.1% normal rabbit serum or with buffer only served as negative controls. After rinsing, all sections were incubated overnight at 4°C with a fluorescein thiocyanate (FITC)-conjugated AffiniPure goat anti-rabbit IgG at a dilution of 1:100. Sections were examined using a Zeiss Axioplan 2 flourescent microscope (Carl Zeiss Inc., West Germany) equipped with a Spot Camera and Spot Software version 2.2 (Diagnostic Instruments, Inc., Sterling Heights, MI).
Results
RT-PCR analysis of sigma receptor 1 mRNA in mouse retina, RPE, lens and brain
After extraction of total RNA from mouse retina, lens and the RPE/choroid complex, RT-PCR was used to establish whether sigma receptor 1 mRNA transcripts were present in these tissues. Mouse brain was used as a positive control. As shown in figure 1A, RT-PCR of the three eye tissues, neural retina, RPE, and lens amplified a product (465 bp) that was similar to that amplified from mouse brain. The RT-PCR products from these tissues were gene-cleaned and subjected to restriction site analysis using two different enzymes (Pst I and Sty I). The restriction pattern with the two enzymes was identical for the RT-PCR products and was exactly as expected from the known restriction map of mouse sigma receptor 1 cDNA (data shown for retina in Fig. 1B, data for lens and RPE not shown). These data confirm that the RT-PCR product from mouse retina, RPE and lens is identical to sigma receptor 1.
Figure 1. RT-PCR analysis of sigma receptor 1-specific mRNA transcript in mouse tissues.

(A) Total RNA was isolated from mouse brain (positive control), lens, RPE/choroid and neural retina. RT-PCR was carried out with primers specific for mouse sigma receptor 1 mRNA. As a negative control, the brain sample was run through the RT-PCR procedure without reverse transcriptase (- RT). (B) Restriction analysis of RT-PCR products. Shown here, the RT-PCR product from retina was gene cleaned and used for restriction analysis with PstI and StyI. (Restriction analysis for other tissues not shown). DNA standards (75 bp – 12.2 kbp) were run on the gel in parallel to determine the size of the RT-PCR products.
RT-PCR analysis of sigma receptor 1 mRNA in rat ganglion and Müller cells and human RPE cells
The data from RT-PCR analysis presented above indicated that sigma receptor 1 mRNA was expressed in mouse retina. The limitations of the dissection procedure do not reveal which neural retinal cells contain the mRNA encoding sigma receptor 1. To address this, we used cell lines of two retinal cell types and carried out additional RT-PCR analysis to examine the expression of sigma receptor 1. We used a rat ganglion cell line [17] and a rat Müller cell line [25], cultured them as described above and extracted total RNA from them. Using primers specific for rat sigma receptor 1, we amplified a 512 bp RT-PCR product from Müller cells and ganglion cells. In addition to these two cell types from neural retina, we cultured a human RPE cell line, ARPE-19 and examined the expression of sigma receptor 1 mRNA using primers specific for human sigma receptor 1 by RT-PCR. As shown in Fig. 2, an RT-PCR product of approximately 509 bp was amplified. The molecular identity of the RT-PCR products from rat ganglion and Müller cell lines and from human ARPE-19 cells was established by restriction analysis (data not shown).
Figure 2. RT-PCR analysis of sigma receptor 1-specific mRNA transcript in retinal cell lines.
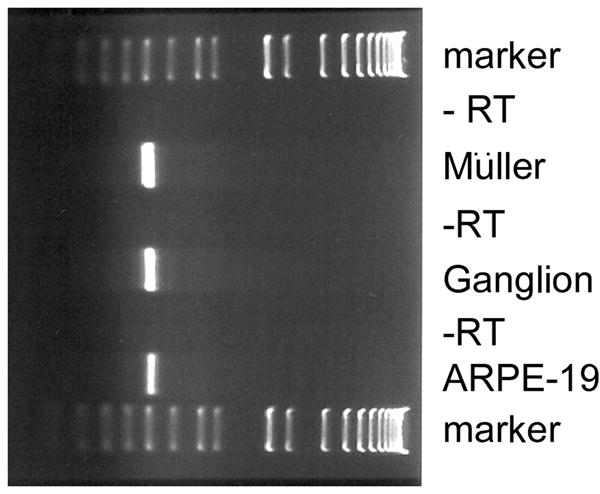
(A) Total RNA was isolated from rat Müller cells, rat ganglion cells and human RPE (ARPE-19) cells. RT-PCR was carried out with primers specific for rat sigma receptor 1 mRNA in the case of the Müller and ganglion cells and human sigma receptor 1 mRNA in the case of the ARPE-19 cells. As negative controls, the samples were run through the RT-PCR procedure without reverse transcriptase (- RT). DNA standards (75 bp – 12.2 kbp) were run on the gel in parallel to determine the size of the RT-PCR products.
In situ hybridization analysis of sigma receptor 1 in mouse eyes
In situ hybridization analysis was performed on cryosections of adult albino mouse eyes (n = 5) to determine which cells of the retina expressed the mRNA encoding the sigma receptor 1 and which additional neuronal and non-neuronal ocular tissues also expressed this transcript. The studies were performed using digoxigenin-labeled riboprobes. The in situ hybridization data for the retina are shown in figure 3. An H & E stained section of the retina is shown in figure 3A for comparison to panels B,C and D. The sigma receptor 1 mRNA transcripts were detected in the cells of the ganglion cell layer which includes ganglion cells, but may include also an occasional amacrine cell. The labeling of this cell layer is shown at higher magnification in 1C and all cells were positive. The sigma receptor 1 mRNA was expressed also in the inner segments of photoreceptor cells and in the RPE cells (3B and higher power 3D). There was slight, albeit detectable expression of sigma receptor 1 mRNA in cells of the inner portion of the inner nuclear layer. This labeling could represent labeling of amacrine cells and/or labeling of Müller cells. As shown in panel 3E, there was considerable expression of sigma receptor 1 mRNA in oligodendrocytes of the optic nerve. Hybridization of cryosections with the sense probe of the sigma receptor 1 showed no positive signal (Fig. 3F).
Figure 3. Distribution of sigma receptor 1-specific mRNA transcript in adult mouse retina as assessed by in situ hybridization.

(A) Hematoxylin and eosin stained section for comparison to adjacent unstained retinal sections. Several layers of the retina are indicated (gc = ganglion cell layer, inl = inner nuclear layer, onl = outer nuclear layer, is = inner segment, rpe = retinal pigment epithelium). (B) Mouse retina probed with the antisense digoxigenin-labeled sigma receptor 1 riboprobe showing positive reaction in ganglion cell, inner segment and RPE layers. The intense purple stain indicates a positive reaction. (C) Higher magnification of inner portion of retina in panel “B” showing intense labeling of ganglion cells (arrows). (D) Higher magnification of outer portion of retina in panel “B” showing intense labeling of inner segments and RPE layer. (E) Sigma receptor 1 mRNA detected in optic nerve. (F) Mouse retina probed with sense (negative control) digoxigenin-labeled sigma receptor 1 riboprobe. No specific signal is detected with the sense probe. (Magnifications: A, B, F = X200; C,D, E = X400).
Non-neuronal tissues of the eye also expressed the mRNA encoding sigma receptor 1. In figure 4, the left panel photomicrographs (A,C,E) are H & E stained sections for comparison to the right panel of sections on which the in situ hybridization assays were performed. In the cornea, the columnar cells of the stratified epithelium (4B) were positive for sigma receptor 1 whereas the anterior epithelium and corneal stroma (substantia propria) were negative. In the ciliary body and iris, the epithelium was positive (Fig. 4D). It should be pointed out that the dark purple positive reaction is not to be confused with pigmentation of these epithelial cells (nor in the RPE shown in fig. 3D) as albino mice were used for the analysis. In the lens, the epithelial cells of the intermediate zone and lens equator were positive for the sigma receptor 1 mRNA as were the nucleated lens fibers (4F). The remaining lens fibers were negative.
Figure 4. Distribution of sigma receptor 1-specific mRNA transcript in non-retinal adult mouse eye as assessed by in situ hybridization.
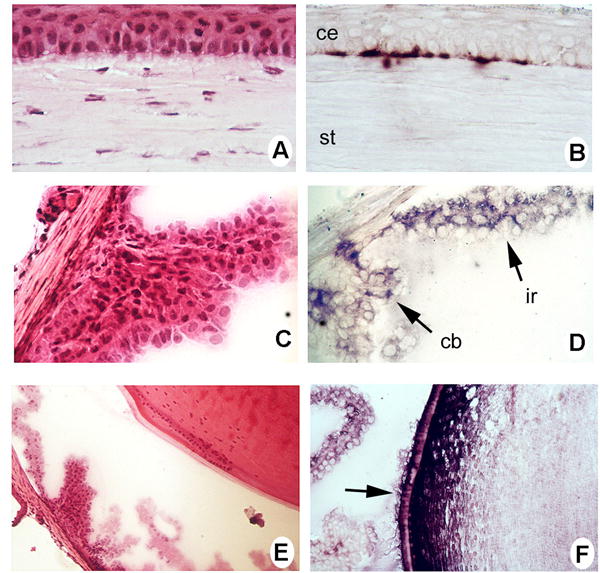
(A) Hematoxylin and eosin (H&E) stained section of the cornea for comparison to adjacent section. (B) Mouse cornea probed with the antisense digoxigenin-labeled sigma receptor 1 riboprobe showing positive reaction (purple color) in the basal epithelial cells of cornea (ce) but not staining in the stroma (st). (C) H & E stained section of the ciliary body and iris. (D) Mouse ciliary body (cb) and iris (ir) probed with the antisense digoxigenin-labeled sigma receptor 1 riboprobe showing positive reaction (purple color) in these structures. (E) H & E stained section of the lens and adjacent ciliary body. (F) Mouse lens probed with the antisense digoxigenin-labeled sigma receptor 1 riboprobe showing positive reaction (purple color) in cells at the equator of the lens (arrow). (Magnification: A, B = X 400; C,D,E,F = X200)
Immunolocalization of sigma receptor 1 in mouse eyes
Our RT-PCR analysis (Figs. 1 and 2) demonstrated the presence of sigma receptor 1 mRNA in retina, lens and RPE and the in situ hybridization supported this observation. To localize the sigma receptor 1 protein itself, we used a monoclonal antibody against sigma receptor 1 and performed immunohistochemical analysis in cryosections of mouse eye. Figure 5A shows the immunolabeling of sigma receptor 1 in retina at low magnification. Sigma receptor 1 was detected in cells of the ganglion layer and surrounding soma of cells in the inner nuclear layer. Sigma receptor 1 was present also around the soma of photoreceptor cells, and in the RPE layer. A higher magnification of the inner retina is shown in fig. 5B in which the intense labeling of ganglion cells and cells throughout the inner nuclear layer is observed. Fig. 5C shows the intense labeling of cells of oligodendrocytes and the external mantle of the optic nerve. In non-retinal tissues, sigma receptor 1 was detected in corneal epithelial cells (Fig. 5D), in the epithelial cells of the ciliary body (Fig. 5E) and in the equatorial cells of the lens (Fig. 5F).
Figure 5. Light microscopic immunolocalization of sigma receptor 1 in intact mouse eye.

Mouse cryosections were incubated with antibody against sigma receptor 1 followed by incubation with FITC-labeled secondary antibody as described in the text. (A) Immunolabeling of mouse retina: sigma receptor 1 is present in the ganglion cells (gc), around soma of cells in the inner nuclear layer (inl), cell bodies of photoreceptor cells (onl), and cells in the RPE layer (rpe). (B) Higher magnification of panel “A” showing intense immunolabeling of ganglion cells (arrow) and cells of the inner nuclear layer. (C) Optic nerve. (D) Cornea. Note sigma receptor 1 immunolabeling of the epithelial cells (ce) of the cornea and some labeling of the corneal stroma (st). (E) Iris-ciliary complex. Note intense labeling of cells of the ciliary body (cb). (F) Lens. Note immunolabeling of cells along the equator of the lens (arrow). (Magnification: A = X200; B,C = X 400; D,E, F = X100)
Discussion
In the eye, receptor binding assays have provided pharmacological evidence for the presence of sigma receptor 1 in lacrimal gland [26,27], iris-ciliary body [6] and retina [28,29]. These data are extremely important as sigma receptor 1 ligands may have potential therapeutic use. For example, lacrimal secretion has been stimulated with sigma receptor 1 ligands [33], making these compounds of possible therapeutic use in dry eye syndrome [27]. Topical administration of sigma receptor 1-site agonists lowered intraocular pressure, hence they may offer a novel class of agents effective in control of ocular hypertension [6]. In retina, sigma receptor 1 binding sites have been demonstrated in bovine [28] and rat retinal membranes [29]. Though these studies established the presence of sigma receptor in neural retina, they did not demonstrate in which cell type(s) sigma receptor 1 was present nor did they establish unequivocally the molecular identity of the receptor. The present study was designed to address these issues.
In the present experiments, we determined which retinal cells express sigma receptor 1 using semi-quantitative RT-PCR, in situ hybridization and immunohistochemical methods. We used intact retinal tissue from mice as well as three well-characterized retinal cell lines. The RT-PCR analysis of intact tissue amplified sigma receptor 1 in neural retina, RPE/choroid complex, and the lens. While the dissection of the lens yields a purified cell population, the retina/RPE dissection does not permit dissection of individual cell types. Therefore, we used ganglion, Müller, and RPE cell lines to determine whether sigma receptor 1 mRNA was expressed in these specific retinal cell types. The rat ganglion cell line RGC-5 is phenotypically similar to normal ganglion cells and expresses several ganglion cell-specific markers including Thy-1 and Brn-3C. RGC-5 cells are considered a useful model to study various aspects of retinal ganglion cell biology [17]. The rat Müller cell line has many characteristic features of normal Müller cells including expression of GFAP and CRALBP [25]. The human ARPE-19 cells have been used extensively for studies of RPE as they have many characteristic features of normal RPE cells including formation of tight junctions, development of microvillous processes and the ability to phagocytose shed photoreceptor outer segments disks [8]. In all three cases, the RT-PCR amplified a product of appropriate size and restriction digestion with appropriate enzymes confirmed that sigma receptor 1 was expressed in RPE, Müller and ganglion cells. These data establish unequivocally the molecular identity of sigma receptor 1 in these cell types. Furthermore, our in situ hybridization studies using an antisense probe specific for sigma receptor 1 localized for the first time the mRNA encoding this protein. Sigma receptor 1 mRNA was expressed in a variety of retinal cell types including ganglion cells, the innermost cells of the inner nuclear layer, inner segments of photoreceptor cells and RPE cells. Given that several cell types comprise the inner nuclear layer, the cells positive for sigma receptor 1 probe could include Müller, amacrine, bipolar or horizontal cells. Based on our studies with cell lines, we can only say definitively that in the inner nuclear layer, Müller cells express the sigma receptor 1 transcript. Senda and colleagues studied sigma receptor 1 binding in primary cultures of fetal rat retina and manipulated the cultures to favor growth of amacrine cells [29]. Thus, amacrine cells may also be one of the cell types that express sigma receptor 1. The development of an amacrine line would be useful in resolving this question. The immunohistochemical analysis localized the sigma receptor 1 protein to ganglion cells, soma of cells throughout the inner nuclear layer, soma of photoreceptor cells, and RPE cells. While the gene expression colocalizes for the most part with the protein in the retina, there are two notable exceptions. In the inner nuclear layer, the mRNA encoding sigma receptor 1 is limited to the innermost cells of the layer, while the protein is expressed abundantly throughout this layer. The explanation for this discrepancy is not known. In the photoreceptor cells, the mRNA expression is restricted to the inner segments because the endoplasmic reticulum is sequestered in this portion of the cell. The sigma receptor 1 protein is localized to the soma that occupy the outer nuclear layer of the retina.
In addition to the expression of sigma receptor 1 in retina, the use of intact mouse ocular tissue permitted in situ hybridization and immunohistochemical analyses of other non-retinal ocular structures. The expression of sigma receptor 1 in ciliary body supports the binding studies observations by Buccolo and co-workers [6] that iris-ciliary body has sigma receptor 1 and thus may be amenable targets for sigma receptor 1 ligands. Our observations that sigma receptor 1 was expressed in lens and cornea may suggest uses for sigma receptor 1 ligands in disorders of these tissues as well. Interestingly, while the expression patterns of sigma receptor 1 mRNA were similar to immunostaining patterns in ciliary body and lens, this was not the case with the cornea. The in situ hybridization analysis showed expression in cells at the basal layer of the corneal epithelium, while remaining corneal epithelial and stromal cells were negative. Immunohistochemical analysis, however, demonstrated widespread presence of the protein throughout the corneal epithelium as well as in distinct fibers of the corneal stroma.
For the purposes of our future work, we were most intrigued by the expression of sigma receptor 1 in ganglion cells; particularly as this might relate to alterations of ganglion cells in diabetes. Diabetic retinopathy is the leading cause of blindness in the United States. The vascular changes that accompany diabetic retinopathy have been studied extensively and include hemorrhages, microaneurysms, sharply defined waxy deposits and vessel proliferation [9]. These vascular alterations set in after 10-15 years of diabetes in humans and after more than a year in rat models of diabetes. Unlike these late-onset vascular changes, degenerative processes begin in the neural retina shortly after the onset of diabetes. Loss of color and contrast sensitivity are early signs of neural retinal dysfunction in humans that occur within 2 years of diabetes. Alterations of the b-wave of the electroretinogram occur after only a few years of the onset of diabetes in human patients and within 2 weeks in rats. It has been proposed by Bresnick [5] that diabetic retinopathy is a primary neurosensory disorder that precedes vasculopathy by many years. Very recently, Barber and co-workers [3] documented increased apoptosis of neural retinal cells in experimental diabetes in rats and diabetes mellitus in humans. At particular risk were retinal ganglion cells, which demonstrated a 10% decrease in cell number in streptozotocin-induced diabetic rats compared with controls. Though present day treatment of diabetic retinopathy focuses on the vascular pathology of this disease through photocoagulation therapy of leaky blood vessels [20], additional therapeutic strategies to protect retinal neuronal cells are sorely needed.
The data presented in this study demonstrating the expression of sigma receptor 1 in ganglion cells are relevant to the death of these cells in diabetes. There is evidence that the ganglion cell death in diabetic retinopathy occurs via glutamate-mediated toxicity [18]. Within 3 months of the onset of diabetes in the streptozotocin-induced rat model, levels of glutamate increase significantly. Undiluted vitreous samples from diabetic patients who had undergone pars plana vitrectomy had levels of glutamate that were 2.5 times greater than non-diabetic persons [2]. The toxic effects of glutamate on the retina, particularly the retinal ganglion cells are well established [22,36]. The neurotoxicity is caused by excessive stimulation of receptors for excitatory amino acids predominantly through the N-methyl-D aspartate (NMDA) subtype of glutamate receptor. NMDA-receptor-mediated toxicity in retinal ganglion cells is dependent on the influx of extracellular Ca2+. The increase in intracellular Ca2+ concentration acts as a second messenger that sets in motion the cascade leading to eventual cell death. Glutamate further stimulates its own release in a positive feedback loop by its interaction with the non-NMDA receptor subtypes. Ca2+-induced Ca2+ release and further influx of Ca2+ through voltage-gated Ca2+ channels after glutamate-induced depolarization contribute to glutamate toxicity. Given that the neuroprotective effects of sigma receptor ligands are thought to be due to modulation of NMDA receptors [4,21], and that these receptors are present on ganglion cells, we speculate that apoptotic ganglion cell death characteristic of diabetic retinopathy may be amenable to neuroprotection by these ligands. Future studies in our lab will explore the use of sigma receptor 1 ligands in treatment of ganglion cell death in diabetic retinopathy.
Acknowledgments
This work was supported by National Institutes of Health Grants EY12830, EY13089, and DA10065, Fight for Sight-Prevent Blindness America, an unrestricted award from Research to Prevent Blindness, Inc. to the Department of Ophthalmology, Medical College of Georgia, the Medical College of Georgia Research Institute and the American Health Assistance Foundation – National Glaucoma Foundation.
References
- 1.Alonso G, Phan V, Guillemain I, Saunier M, Legrand A, Anoal M, Maurice T. Immunocytochemical localization of the sigma(1) receptor in the adult rat central nervous system. Neuroscience. 2000;97:155–170. doi: 10.1016/s0306-4522(00)00014-2. [DOI] [PubMed] [Google Scholar]
- 2.Ambati J, Chalam KV, Chawla DK, D'Angio CT, Guillet EG, Rose SJ, Vanderlinde RE, Ambati BK. Elevated gamma-aminobutyric acid, glutamate, and vascular endothelial growth factor levels in the vitreous of patients with proliferative diabetic retinopathy. Arch Ophthalmol. 1997;115:1161–1166. doi: 10.1001/archopht.1997.01100160331011. [DOI] [PubMed] [Google Scholar]
- 3.Barber AJ, Lieth E, Khin SA, Antonetti DA, Buchanan AG, Gardner TW. The Penn State Retina Research Group, Neural apoptosis in the retina during experimental and human diabetes: early onset and effect of insulin. J Clin Invest. 1998;102:783–791. doi: 10.1172/JCI2425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bowen WD, Kirschner BN, Newmen AH, Rice KC. Sigma receptors negatively modulate agonist-stimulated phosphoinositide metabolism in rat brain. Eur J Pharmacol. 1988;149:399–400. doi: 10.1016/0014-2999(88)90678-4. [DOI] [PubMed] [Google Scholar]
- 5.Bresnick GJ. Diabetic retinopathy viewed as a neurosensory disorder. Arch Ophthalmol. 1986;104:989–990. doi: 10.1001/archopht.1986.01050190047037. [DOI] [PubMed] [Google Scholar]
- 6.Bucolo C, Campana G, Di Toro R, Cacciaguerra S, Spampinato S. Sigma 1 recognition sites in rabbit iris-ciliary body: topical sigma1-site agonists lower intraocular pressure. J Pharmacol Exp Ther. 1993;289:1362–1369. [PubMed] [Google Scholar]
- 7.Chancy CD, Kekuda R, Huang W, Prasad PD, Kuhnel JM, Sirotnak FM, Roon P, Ganapathy V, Smith SB. Expression and Differential Polarization of the Reduced-Folate Transporter-1 and the Folate Receptor alpha in Mammalian Retinal Pigment Epithelium. J Biol Chem. 2000;275:20676–20684. doi: 10.1074/jbc.M002328200. [DOI] [PubMed] [Google Scholar]
- 8.Dunn KC, Aotaki-Keen AE, Putkey FR, Hjelmeland LM. ARPE-19, a human retinal pigment epithelial cell line with differentiated properties. Exp Eye Res. 1996;62:155–169. doi: 10.1006/exer.1996.0020. [DOI] [PubMed] [Google Scholar]
- 9.Engerman RL, Kern TS. Retinopathy in animal models of diabetes. Diabetes Metab Rev. 1995;11:109–120. doi: 10.1002/dmr.5610110203. [DOI] [PubMed] [Google Scholar]
- 10.Ferris CD, Hirsch DJ, Brooks BP, Snyder SH. Sigma Receptors: from molecule to man. J Neurochem. 1991;57:729–737. doi: 10.1111/j.1471-4159.1991.tb08213.x. [DOI] [PubMed] [Google Scholar]
- 11.Ganapathy ME, Prasad PE, Huang W, Seth P, Leibach FH, Ganapathy V. Molecular and Ligand-Binding Characterization of the sigma-receptor in the Jurkat Human T Lymphocyte Cell Line. J Pharmacol Exp Ther. 1999;289:251–260. [PubMed] [Google Scholar]
- 12.Hanner M, Moebius FF, Flandorfer A, Knaus HG, Striessnig J, Kempner E, Glossmann H. Purification, molecular cloning, and expression of the mammalian sigma 1-binding site. Proc Natl Acad Sci U S A. 1996;93:8072–8077. doi: 10.1073/pnas.93.15.8072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Jbilo O, Vidal H, Paul R, De Nys N, Bensaid M, Silve S, Carayon P, Davi D, Galiegue S, Bourrie B, Guillemot JC, Ferrara P, Loison G, Maffrand JP, Le Fur G, Casellas P. Purification and characterization of the human SR 31747A-binding protein. A nuclear membrane protein related to yeast sterol isomerase. J Biol Chem. 1997;272:27107–27115. doi: 10.1074/jbc.272.43.27107. [DOI] [PubMed] [Google Scholar]
- 14.Karbon EW, Patch RJ, Pontecorvo MJ, Ferkany JW. Ifenprodil potently interacts with [3H](+)-3-PPP-labeled sigma binding sites in guinea pig brain membranes. Eur J Pharmacol. 1990;176:247–248. doi: 10.1016/0014-2999(90)90538-h. [DOI] [PubMed] [Google Scholar]
- 15.Kekuda R, Prasad PD, Fei YJ, Leibach FH, Ganapathy V. Cloning and functional expression of the human type 1 sigma receptor (hSigmaR1) Biochem Biophys Res Commun. 1996;229:553–558. doi: 10.1006/bbrc.1996.1842. [DOI] [PubMed] [Google Scholar]
- 16.Kitaichi K, Chabot J, Moebius FF, Flandorfer A, Glossmann H, Quirion R. Expression of the purported sigma(1) (sigma(1)) receptor in the mammalian brain and its possible relevance in deficits induced by antagonism of the NMDA receptor complex as revealed using an antisense strategy. J Chem Neuroanat. 2000;20:375–387. doi: 10.1016/s0891-0618(00)00106-x. [DOI] [PubMed] [Google Scholar]
- 17.Krishnamoorthy RR, Agarwal P, Prasanna G, Vopat K, Lambert W, Sheedlo HJ, Pang IH, Shade D, Wordinger RJ, Yorio T, Clark AF, Agarwal N. Characterization of a transformed rat retinal ganglion cell line. Mol Brain Res. 2001;86:1–12. doi: 10.1016/s0169-328x(00)00224-2. [DOI] [PubMed] [Google Scholar]
- 18.Lieth E, Barber AJ, Xu B, Dice C, Ratz MJ, Tanase D, Strother JM. Glial reactivity and impaired glutamate metabolism in short-term experimental diabetic retinopathy. Diabetes. 1998;47:815–820. doi: 10.2337/diabetes.47.5.815. [DOI] [PubMed] [Google Scholar]
- 19.Lobner D, Lipton P. Sigma-ligands and non-competitive NMDA antagonists inhibit glutamate release during cerebral ischemia. Neurosci Lett. 1990;117:169–174. doi: 10.1016/0304-3940(90)90139-z. [DOI] [PubMed] [Google Scholar]
- 20.Menchini U, Lanzetta P, Soldano F, Ferrari E, Virgili G. Continuous wave Nd:YAG laser photocoagulation in proliferative diabetic retinopathy. Brit J Ophthamol. 1995;79:642–645. doi: 10.1136/bjo.79.7.642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Monnet FP, Debonnel G, Fournier A, De Montigny C. Neuropeptide Y potentiates the N-methyl-D-aspartate response in the CA3 dorsal hippocampus. II. Involvement of a subtype of sigma receptors. J Pharmacol Exp Ther. 1992;263:1219–1225. [PubMed] [Google Scholar]
- 22.Moore P, El-Sherbeny A, Roon P, Schoenlein PV, Ganapathy V, Smith SB. Apoptotic retinal ganglion cell death is induced in vivo by the excitatory amino acid homocysteine. Exp Eye Res. 2001;73:45–57. doi: 10.1006/exer.2001.1009. [DOI] [PubMed] [Google Scholar]
- 23.Prasad PD, Li HW, Fei YJ, Ganapathy ME, Fujita T, Plumley LH, Yang-Feng TL, Leibach FH, Ganapathy V. Exon-intron structure, analysis of promoter region, and chromosomal localization of the human type 1 sigma receptor gene. J Neurochem. 1998;70:443–451. doi: 10.1046/j.1471-4159.1998.70020443.x. [DOI] [PubMed] [Google Scholar]
- 24.Quirion R, Bowen WD, Itzhak Y, Junien JL, Musacchio JM, Rothman RB, Su TP, Tam SW, Taylor DP. A proposal for the classification of sigma binding sites. Trends Pharmacol Sci. 1992;13:85–86. doi: 10.1016/0165-6147(92)90030-a. [DOI] [PubMed] [Google Scholar]
- 25.Sarthy VP, Brodjian SJ, Dutt K, Kennedy BN, French RP, Crabb JW. Establishment and characterization of a retinal Müller cell line. Invest. Ophthalmol Vis Sci. 1998;39:212–216. [PubMed] [Google Scholar]
- 26.Schoenwald RD, Barfknecht CF, Shirolkar S, Xia E, Ignace CC. Identification of sigma receptors in lacrimocytes and their therapeutic implication in dry eye syndrome. Adv Exp Med Biol. 1994;350:141–146. doi: 10.1007/978-1-4615-2417-5_24. [DOI] [PubMed] [Google Scholar]
- 27.Schoenwald RD, Barfknecht CF, Xia E, Newton RE. The presence of σ receptor in the lacrimal gland. J Ocular Pharmacol. 1993;9:125–139. doi: 10.1089/jop.1993.9.125. [DOI] [PubMed] [Google Scholar]
- 28.Senda T, Matsuno K, Mita S. The presence of σ receptor subtypes in bovine retinal membranes. Exp Eye Res. 1997;64:857–860. doi: 10.1006/exer.1996.0272. [DOI] [PubMed] [Google Scholar]
- 29.Senda T, Mita S, Kaneda K, Kikuchi M, Akaike A. Effect of SA4503, a novel σ1 receptor agonist, against glutamate neurotoxicity in cultured rat retinal neurons. Eur J Pharmacol. 1998;342:105–111. doi: 10.1016/s0014-2999(97)01450-7. [DOI] [PubMed] [Google Scholar]
- 30.Seth P, Fei YJ, Li HW, Huang W, Leibach FH, Ganapathy V. Cloning and functional characterization of a σ receptor from rat brain. J Neurochem. 1998;70:922–931. doi: 10.1046/j.1471-4159.1998.70030922.x. [DOI] [PubMed] [Google Scholar]
- 31.Seth P, Ganapathy ME, Conway SJ, Chancy CD, Smith SB, Casellas P, Ganapathy PV. Expression pattern of the type 1 sigma receptor in the brain and identity of critical anionic amino acid residues in the ligand binding domain of the receptor. Biochim Biophys Acta. 2001;1540:59–67. doi: 10.1016/s0167-4889(01)00117-3. [DOI] [PubMed] [Google Scholar]
- 32.Seth P, Leibach FH, Ganapathy V. Cloning and structural analysis of the cDNA and the gene encoding the murine Type 1 sigma receptor. Biochem Biophys Res Comm. 1997;241:535–540. doi: 10.1006/bbrc.1997.7840. [DOI] [PubMed] [Google Scholar]
- 33.Shirolkar S, Schoenwald RD, Barfknecht CF, Xia E, Cheng B, Iwai Y, Ignace CC, Vidvauns S, Newton RE. Lacrimal secretion stimulants: sigma receptors and drug implications. J Ocul Pharmacol. 1993;9:211–227. doi: 10.1089/jop.1993.9.211. [DOI] [PubMed] [Google Scholar]
- 34.Smith SB, Duncan T, Kutty G, Kutty RK, Wiggert B. Increase in retinyl palmitate concentration in eyes and livers and the concentration of interphotoreceptor retinoid-binding protein in eyes of vitiligo mutant mice. Biochem J. 1994;300:63–68. doi: 10.1042/bj3000063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Su TP. Sigma receptors. Putative links between nervous, endocrine and immune systems. Eur J Biochem. 1991;200:633–642. doi: 10.1111/j.1432-1033.1991.tb16226.x. [DOI] [PubMed] [Google Scholar]
- 36.Sucher NJ, Lipton SA, Dreyer EB. Molecular basis of glutamate toxicity in retinal ganglion cells. Vis Res. 1997;37:3483–3493. doi: 10.1016/S0042-6989(97)00047-3. [DOI] [PubMed] [Google Scholar]
- 37.Zuo P, Ogita K, Yoneda Y. Presence of the binding of a variety of ligands related to ionotropic excitatory amino acid receptors in rat retina. Brain Res. 1992;576:168–172. doi: 10.1016/0006-8993(92)90626-k. [DOI] [PubMed] [Google Scholar]