

Abstract
Botulinum neurotoxins (BoNTs), which are highly toxic proteins responsible for botulism, are produced by different strains of Clostridium botulinum. These various strains of bacteria produce seven distinct serotypes, labeled A-G. Once inside cells, the zinc-dependent proteolytic light chain (LC) degrades specific proteins involved in acetylcholine release at neuromuscular junctions causing flaccid paralysis, specifically SNAP-25 for BoNT/A.
BoNT endopeptidase assays using short substrate homologues have been widely used and developed because of their ease of synthesis, detection limits and cost. SNAPtide, a 13-amino acid FRET peptide, was used in this study as a SNAP-25 homologue for the endopeptidase kinetics study of BoNT/A LC. SNAPtide uses a FITC/DABCYL FRET pair to produce a signal upon substrate cleavage. Signal quenching can become an issue after cleavage since quencher molecules can quench cleaved fluorophore molecules in close proximity, reducing the apparent signal. This reduction in apparent signal provides an inherent error as SNAPtide concentrations are increased.
In this study, fluorescence internal quenching (FIQ) correction factors were derived using an unquenched SNAPtide peptide to quantify the signal quenching over a range of SNAPtide concentrations and temperatures. The BoNT/A LC endopeptidase kinetics at the optimally active temperature (37°C) using SNAPtide were studied and used to demonstrate the FIQ Correction Factors in this study. The FIQ Correction Factors developed provide a convenient method to allow for improved accuracy in determining and comparing BoNT/A LC activity and kinetics using SNAPtide over a broad range of concentrations and temperatures.
Keywords: Botulinum Neurotoxin, Clostridium, Internal Quenching, SNAPtide, Endopeptidase, Kinetics
Introduction
Botulinum neurotoxin (BoNT) is a member of a family of clostridial neurotoxins (CNTs) and is the most toxic substance known with a mouse LD50 on the order of 10 pg/kg.1,2,3 BoNTs include seven serotypes, A-G, which contain exclusive immunological characteristics and similar pharmacological properties.4 Each of these serotypes causes botulism; the result of intoxication is flaccid muscle paralysis by blocking the release of acetylcholine at neuromuscular junctions.5 Four of the serotypes (A, B, E and F) are responsible for botulism in humans.6 Because of this extreme toxicity, BoNTs are classified as a Category A bioterrorism agent by the Center for Disease Control.7 Despite the extreme toxicity, BoNTs are the subject of much interest as therapeutic agents for a wide range of neuromuscular disorders.8
BoNTs are approximately 150 kDa proteins, consisting of a 100 kDa heavy (H) chain and a 50 kDa light (L) chain linked through a disulfide bond. The H chain binds specifically to neuronal cells and assists L chains entry into the cell. The L chains are unique zinc-endopeptidases with remarkable substrate specificity. Substrate recognition requires a substantially long peptide sequence ranging from 16–50 amino acids, depending on the serotype.10 This is unique to BoNTs as other microbial metalloproteases can recognize sequences as short as a dipeptide.11 Serotypes A and E cleave SNAP-25 (25-kDa synaptosome associated protein), serotypes B, D, F, and G cleave synaptobrevin/VAMP (vesicle associated membrane protein) and serotype C cleaves SNAP-25 as well as syntaxin.12 While some substrates may be cleaved by more than one serotype, all serotypes cleave unique sites on the SNARE targets.
SNAPtide, a 13-amino acid peptide (sequence: (FITC)TRIDEANQRATK(DABCYL)M), has been used as a SNAP-25 homologue for the endopeptidase kinetics study of BoNT/A L chain (LCA). The intact SNAPtide is a fluorescence resonance energy transfer (FRET) based substrate where the N-terminal fluorophore, FITC, is quenched by DABCYL located on the C-terminal of the peptide. Once the peptide is cleaved, fluorescence is restored and can be measured spectroscopically.13
The SNAPtide assay is currently extensively used by many researchers in the field. However, there is a major technical problem in using this method for accurate kinetic analysis. Even after the SNAPtide is cleaved to release fluorescence (Figure 1), internal quenching can become an issue due to the presence of DABCYL on other intact SNAPtide molecules and DABCYL on cleaved SNAPtide fragments. The FITC portion of the cleaved SNAPtide should theoretically give full fluorescence; however, if FITC comes into close proximity to a DABCYL molecule, fluorescence would be decreased through quenching. This quenching problem becomes acute in enzyme kinetic analysis where varying concentrations of the substrate are used.
Figure 1.
Overview of SNAPtide cleavage by BoNT/A LC. In-tact SNAPtide (top) produces no fluorescence since the FRET pair is in close proximity. Fluorescence signal of FITC is restored upon cleavage (bottom) where the distance between FITC and DABCYL is larger than approximately 100 Å.17
In this study, we have developed a fluorescence internal quenching (FIQ) correction factor using an unquenched cleaved SNAPtide to correct LCA endopeptidase kinetics at its optimally active temperature (37°C) This correction factor allows us to more accurately determine BoNT/A LC activity and kinetic parameters over a range of SNAPtide concentrations and temperatures. These FIQ Correction Factors can have a widespread benefit to studies using SNAPtide for inhibitor design such as by Boldt et al.14,15, residue substitution studies such as examined by Baldwin et al.13, BoNT/A detection as done by Bagramyan et al.16, and any studies requiring comparisons of multiple SNAPtide concentrations for endopeptidase activity or kinetics as studied by Baldwin et al.13.
Experimental Section
LCA Endopeptidase Substrate Calibration
A calibration curve for the SNAPtide used in the LCA endopeptidase kinetics was created using Product #528 (List Biological Labs), an unquenched SNAPtide peptide. The following concentrations of unquenched SNAPtide were prepared in 100.0 μL total volume using 20 mM HEPES, 0.1% v/v TWEEN-20, pH 7.6 (assay buffer): 0.0125, 0.0250, 0.0500, 0.100, 0.200, 0.400, 0.600 and 0.800 μM. Triplicate samples were carried out in a 96 well plate using a SpectraMax M5 plate reader at temperatures between 25–50°C and the fluorescence signal (490 nm excitation, 523 nm emission, 495 nm cutoff filter) was plotted vs. unquenched SNAPtide concentration. From the data, trendlines were calculated for each triplicate sample showing a slope with units of RFU/μM. An average of the inverse slope was calculated at each temperature.
Fluorescence Internal Quenching (FIQ) Correction Factor
A SNAPtide stock solution (2.5 mM in DMSO, List Biological Labs, Product #521) was diluted to 0.200 mM in the assay buffer before further dilutions. The unquenched SNAPtide stock solution (500 μM in DMSO) was diluted to 50.0 μM in assay buffer prior to further dilutions.
Fluorescence endpoint readings of each SNAPtide concentration used (0.00, 1.00, 1.50, 2.00, 4.00, 8.00, 10.00 μM) were taken in 99.0 μL assay buffer after an initial 15 minute incubation at the designated temperature. Afterward, the 50.0 μM unquenched SNAPtide was added to each SNAPtide concentration (1.00 μL, 0.500 μM final concentration), incubated at the designated temperature for 15 minutes with a second fluorescence endpoint reading taken.
The endpoint reading of 0.00 μM SNAPtide was subtracted from the endpoint reading of 0.500 μM unquenched peptide with 0.00 μM SNAPtide. This value gave the total signal of 0.500 μM unquenched SNAPtide.
Endpoint readings of each SNAPtide concentration in the absence and presence of 0.500 μM unquenched SNAPtide were observed. The signal of SNAPtide in the presence of unquenched SNAPtide was normalized by subtracting the signal of SNAPtide (concentration of interest) in the absence of unquenched SNAPtide giving the apparent signal of the 0.500 μM unquenched SNAPtide for all range of SNAPtide concentrations.
The signal of the unquenched SNAPtide in the presence of each SNAPtide concentration was divided by the total signal of unquenched SNAPtide. This gave a ratio of how much signal from the unquenched SNAPtide reaches the detector in the presence of each SNAPtide concentration (Fluorescence Internal Quenching Correction Factor).
FIQ Correction Factors were calculated for each SNAPtide concentration (1.00–10.00 μM) in each of the triplicate runs. The FIQ Correction Factor was plotted against SNAPtide concentrations and a trendline was calculated for each triplicate sample. The trendline slope for each sample was used to calculate the FIQ Correction Factors assuming 0.00 μM SNAPtide gave a FIQ Correction factor of 1.00 (Calculation 1). The average of these calculated FIQ Correction Factors were used to correct initial velocities observed for BoNT/A LC endopeptidase kinetics.
Calculation 1: Calculation of FIQ correction factor from trendline
The average slope calculated can be multiplied by the SNAPtide concentration used and then added to 1.00 to obtain the calculated FIQ Correction Factor for any SNAPtide concentration within the range tested experimentally. When compared to temperatures between 25–50°C, the correction factors are the same within the experimental error.
LCA Endopeptidase Kinetics
Two stock LCA solutions were used (2.0 mg/mL, purified November 2010 and 2.73 mg/mL, purified March 2011; both samples were stored at −80°C in a 50 mM Tris, 500 mM NaCl, pH 7.6 under 20% v/v glycerol). The stock solutions were diluted to 1000 nM (diluted in assay buffer). A SNAPtide stock solution (2.5 mM in DMSO) was diluted to 0.200 mM in assay buffer. Tests were carried out on a 96 well Costar clear bottom microplate using a SpectraMax M5 plate reader.
The LCA (100 nM) was incubated at 37°C for 30 min. The SNAPtide concentrations used were 0.00 μM, 1.00 μM, 1.50 μM, 2.00 μM, 4.00 μM, 8.00 μM, and 10.0 μM. After the addition of SNAPtide, the plate was read for fluorescence (490 nm excitation, 523 nm emission, 495 nm cutoff filter) at the designated temperature over a 5 min period in 30 second intervals on the microplate reader to obtain the initial velocity (Vo) in RFU/s. Triplicate samples for each SNAPtide concentration were performed.
The slope obtained from the substrate calibration was used to convert the Vo from the SoftMax Pro software from RFU/s to μM/s. The enzyme kinetic parameters (KM, kcat, and kcat/KM) were calculated based on a Lineweaver-Burk plot; these parameters were considered uncalibrated in terms of internal quenching. The Vo was then corrected using the FIQ Correction Factors found and the kinetic parameters above were again calculated, labeled as calibrated.
Results
Substrate Signal Calibration
Calibration curves of fluorescence signal (RFU) vs. Product #528 concentration (μM) were created at each temperature between 25–50°C and then analyzed with a trendline. Sample standard curves for each temperature examined are shown as Figures S1-S6 in the Supporting Information.
Initial velocities from the endopeptidase kinetics calculated by the SoftMax Pro software (in RFU/s) were multiplied by the inverse slope of the trendline from the designated temperature to obtain velocities in μM/s units. These velocities were used to create Michaelis-Menten and Lineweaver-Burk plots (Figures S-8 and S-9 in Supporting Information) and used to calculate kinetic parameters and are labeled as uncalibrated (Figure 3).
Figure 3.
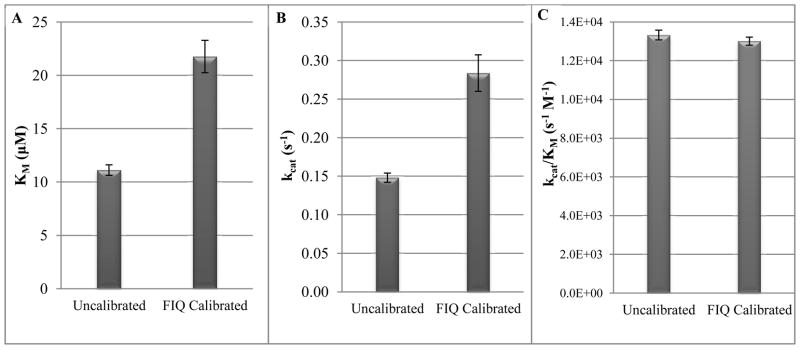
A. Michaelis constant (KM), B. catalytic constant (kcat), and C. catalytic efficiency (kcat/KM) comparisons of BoNT/A LC endopeptidase kinetics data (100 nM LCA) before and after calibration by the 37°C FIQ Correction Factors. Standard deviations of the mean are shown as error bars.
Fluorescence Internal Quenching (FIQ) Correction Factors
FIQ Correction Factors calculated experimentally (in triplicate, 1.00–10.0 μM SNAPtide concentration) for each temperature between 25–50°C were plotted vs. SNAPtide concentration then analyzed with a trendline. The slopes from the trendlines (FIQ Correction Factor/μM SNAPtide) are shown in Table 1 with the visual depiction seen as Figure S-7 in the Supporting Information and were later used to calculate FIQ Correction Factors for each SNAPtide concentration at each temperature (Calculation 1).
Table 1.
Slope of each trendline showing FIQ Correction Factor/μM SNAPtide calculated for each temperature. These slopes can be used to calculate FIQ Correction Factors at SNAPtide concentrations between 0.00–10.0 μM and at reaction temperatures between 25–50°C if the 0.0 μM SNAPtide sample is assumed to have an FIQ Correction Factor of 1.00(Calculation 1). Standard deviations of the mean for the triplicate samples are shown.
| Temperature | 25°C | 30°C | 37°C | 40°C | 45°C | 50°C |
|---|---|---|---|---|---|---|
| FIQ Correction Factor/μM SNAPtide Slope | −0.0264 ± 0.0013 | −0.0255 ± 0.0017 | −0.0247 ± 0.0007 | −0.0232 ± 0.0022 | −0.0255 ± 0.0015 | −0.0258 ± 0.0036 |
FIQ Correction Factors were calculated for each SNAPtide concentration using the slope at the designated temperature from Table 1. The FIQ Correction Factor values are similar, within experimental error, at each concentration over the 25–50°C span (shown in Table 2 and Figure 2).
Table 2.
Calculated Fluorescence Internal Quenching Correction Factor for each SNAPtide concentration at each temperature used for enzyme kinetics studies. Standard deviations of the mean for the triplicate samples are shown.
| SNAPtide Concentration (μM) | 25°C | 30°C | 37°C | 40°C | 45°C | 50°C |
|---|---|---|---|---|---|---|
| 0.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| 1.00 | 0.982 ± 0.001 | 0.982 ± 0.002 | 0.984 ± 0.001 | 0.983 ± 0.002 | 0.983 ± 0.002 | 0.984 ± 0.004 |
| 1.50 | 0.960 ± 0.002 | 0.962 ± 0.003 | 0.963 ± 0.001 | 0.964 ± 0.003 | 0.962 ± 0.002 | 0.961 ± 0.005 |
| 2.00 | 0.947 ± 0.003 | 0.949 ± 0.003 | 0.951 ± 0.001 | 0.952 ± 0.004 | 0.949 ± 0.003 | 0.948 ± 0.007 |
| 4.00 | 0.895 ± 0.005 | 0.898 ± 0.001 | 0.902 ± 0.003 | 0.903 ± 0.009 | 0.898 ± 0.006 | 0.897 ± 0.014 |
| 8.00 | 0.789 ± 0.011 | 0.796 ± 0.013 | 0.805 ± 0.006 | 0.806 ± 0.018 | 0.796 ± 0.012 | 0.794 ± 0.029 |
| 10.0 | 0.736 ± 0.013 | 0.745 ± 0.017 | 0.756 ± 0.007 | 0.758 ± 0.022 | 0.745 ± 0.015 | 0.742 ± 0.036 |
Figure 2.

Visual depiction of the calculated FIQ Correction Factor vs. SNAPtide concentration. FIQ Correction Factors for 37°C depicted. Standard deviations of the mean for the triplicate samples are shown as error bars.
Endopeptidase Kinetics of BoNT/A Light Chain
These calculated FIQ Correction Factors were applied to the initial velocity obtained after the substrate signal calibration by dividing the initial velocity by the appropriate FIQ Correction Factor. The corrected velocities were plotted in a Michaelis-Menten plot and Lineweaver-Burk plot (Figures S-8 and S-9 in the Supporting Information).
Initial velocities at 8.00 and 10.0 μM show the most notable change, as expected. The internal quenching of SNAPtide at these concentrations was at a maximum for this study and had the highest correction factor necessary. The increase to these velocities resulted in a significant change to enzyme kinetic parameters calculated from the Lineweaver-Burk plot.
While the slope of the Lineweaver-Burk plot remains similar after correction with the FIQ Correction Factors (2.2% increase), the y-intercept decreases by approximately half (52.0%). Since the y-intercept is inversely proportional to the maximum velocity, the decrease in y-intercept produces a corrected Vmax of over double the uncalibrated Vmax.
Using the Lineweaver-Burk plots, the KM, kcat, and kcat/KM were calculated and a comparison between using uncalibrated and corrected initial velocities is shown for each parameter (Figure 3).
Correction using the FIQ Correction Factors resulted in a 1.96-fold increase in KM, a 2.12-fold increase in kcat, and essentially no change to the kcat/KM. While calibration increases the error in KM and kcat values, the error remains about the same in kcat/KM since the ratio of the parameters remains similar in the triplicate experiments. The substantial increase in kcat provides strong support for the necessity of a correction factor when using SNAPtide.
Discussion
Improved analysis of assays using SNAPtide would have a wide-reaching benefit to studies involving BoNT/A detection, inhibitor design, BoNT/A residue mutation studies, and BoNT/A endopeptidase activity and kinetics under different conditions such as pH, temperature, and buffer. Significant corrections are necessary to the kinetic parameters and endopeptidase activity of BoNT/A LC when using SNAPtide due to the signal quenching present after cleavage.
We were able to derive correction factors for a range of SNAPtide concentrations at different temperatures to eliminate the effect of internal quenching of SNAPtide when used for endopeptidase studies with BoNT/A LC. The application of these factors to BoNT/A LC endopeptidase kinetics proved to have a significant impact on the kinetic parameters, highlighted by a 1.96-fold increase in KM and 2.12-fold increase in kcat while maintaining a similar kcat/KM.
While these factors were used to correct initial velocities measured in 20 mM HEPES buffer at pH 7.6 in the presence of 0.1% v/v TWEEN-20 for use in enzyme kinetics for this study, the same factors would also be useful under the following conditions:
When using different SNAPtide concentrations for endpoint fluorescence readings in place of kinetic rates
Over a physiological range of pH as well as in different buffers assuming similar ionic strength and viscosity of the buffer used. The fluorescence signal value would likely be altered in different buffers or pH conditions; however, the percentage of quenching used for calculations should remain constant. In contrast, changes to the viscosity or ionic strength would alter the rate of interactions between the FITC/DABCYL and therefore the percentage of quenching.
Furthermore, we noted that the use of a microplate for fluorescence signal monitoring facilitated rapid determination of accurate enzyme activity and kinetics.
Supplementary Material
Acknowledgments
Funding Sources
The work was in part supported by a NIH grant (1U01A1078070-02) and DoD contract (W911QY-10-C-0005).
Authors are grateful to discussion with Dr. Raj Kumar for analyzing the data.
Abbreviations
- BoNT/A
Botulinum Neurotoxin Type A
- LCA
Light Chain A
- FRET
Fluorescence Resonance Energy Transfer
- FIQ
Fluorescence Internal Quenching
- DABCYL
4-((4-(dimethylamino)phenyl)azo) benzoic Acid
- FITC
Fluorescein isothiocyanate
- SNAP-25
Synaptosomal-associated protein 25
- CNT
Clostridial Neurotoxins
Footnotes
Author Contributions
TMF and BRS conceived the idea, TMF carried out the experiments and analyzed the data. BRS evaluated the data and discussed modifications to experimental plans and data presentation. TMF and BRS wrote the manuscript.
Supporting Information. Additional information as noted in the text. This material is available free of charge via the Internet at http://pubs.acs.org.
References
- 1.Binz T, Sikorra S, Mahrhold S. Toxins. 2010;2:665–682. doi: 10.3390/toxins2040665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Cai S, Kukreja R, Shoesmith S, Chang TW, Singh BR. Protein J. 2006;25(7–8):455–462. doi: 10.1007/s10930-006-9028-1. [DOI] [PubMed] [Google Scholar]
- 3.Cai S, Sarkar HK, Singh BR. Biochem. 1999;38:6903–6910. doi: 10.1021/bi990086c. [DOI] [PubMed] [Google Scholar]
- 4.Sharma SK, Singh BR. Biochem. 2004;43:4791–4798. doi: 10.1021/bi0355544. [DOI] [PubMed] [Google Scholar]
- 5.Singh BR. Nature. 2000;7(8):617–619. doi: 10.1038/77900. [DOI] [PubMed] [Google Scholar]
- 6.Montal M. Annu Rev Biochem. 2010;79:591–617. doi: 10.1146/annurev.biochem.051908.125345. [DOI] [PubMed] [Google Scholar]
- 7.Binz T, Sikorra S, Mahrhold S. Toxins. 2010;2:665–682. doi: 10.3390/toxins2040665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Weingart OG, Schreiber T, Mascher C, Pauly D, Dorner MB, Berger TFH, Egger C, Gessler F, Loessner MJ, Avondet MA, Dorner BG. Appl Environ Microbiol. 2010;76(10):3293–3300. doi: 10.1128/AEM.02937-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Wang J, Zurawski TH, Meng J, Lawrence G, Olango WM, Finn DP, Wheeler L, Dolly JO. J Biol Chem. 2011;286(8):6375–6385. doi: 10.1074/jbc.M110.181784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Segelke B, Knapp M, Kadkhodayan S, Balhorn R, Rupp B. PNAS. 2004;101(18):6888–6893. doi: 10.1073/pnas.0400584101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Silvaggi NR, Boldt GE, Hixon MS, Kennedy JP, Tzipori S, Janda K, Allen K. Chem Bio. 2007;14:533–542. doi: 10.1016/j.chembiol.2007.03.014. [DOI] [PubMed] [Google Scholar]
- 12.Kukreja RV, Sharma S, Cai S, Singh BR. Biochim Biophys Acta. 2007;1774:213–222. doi: 10.1016/j.bbapap.2006.11.007. [DOI] [PubMed] [Google Scholar]
- 13.Baldwin MR, Bradshaw M, Johnson EA, Barbieri JT. Protein Expression Purif. 2004;37:187–195. doi: 10.1016/j.pep.2004.05.009. [DOI] [PubMed] [Google Scholar]
- 14.Boldt GE, Eubanks LM, Janda KD. Chem Comm. 2006:3063–3065. doi: 10.1039/b603099h. [DOI] [PubMed] [Google Scholar]
- 15.Boldt GE, Kennedy JP, Hixon MS, McAllister LA, Barbieri JT, Tzipori S, Janda KD. J Comb Chem. 2006;8:513–521. doi: 10.1021/cc060010h. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Bagramyan K, Barash JR, Arnon SS, Kalkum M. PLoS ONE. 2008;3(4):e2041. doi: 10.1371/journal.pone.0002041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Liu Y, Kati W, Chen CM, Triptathi R, Molla A, Kohlbrenner W. Anal Biochem. 1999;267:331–335. doi: 10.1006/abio.1998.3014. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.