

Abstract
The unicellular protist Blastocystis has long been an unsolved puzzle for taxonomists, microbiologists and clinicians. Over the years, the organism has been bounced on and off the different branches of the tree of life due the possession of unique phenotypic characters intermediary to different organisms. The organism is polymorphic with only few of forms such as vacuolar, granular, amoeboid, and the cyst form being commonly known. However it could exist in other forms much more frequently than the widely known forms which could be missed by the unaware observer. Certain older concepts in the life cycle of Blastocystis although has been proven wrong are still being followed in various textbooks and other trustworthy internet sources. The causal role of Blastocystis in human disease has long been a subject of controversy. It is widely believed that certain subtypes of the organism are virulent. But this is not so as other factors are also involved in the clinical outcome of the infection. In these contexts, this review intends to shed light on the past misconceptions and the recent findings on the taxonomy, biology and the virulence of this organism.
KEY WORDS: Amoeboid form, avacuolar form, Blastocystis, multi-vacuolar form, stramenophile, subtype
INTRODUCTION
Blastocystis is a unicellular, anaerobic, eukaryotic protist which lives in the intestinal tract of diverse hosts including humans. The organism exists in different morphological forms with each form showing considerable variations in size. It is one of the frequently encountered parasites in human faecal samples in the developing countries.[1] Currently it is estimated that more than 1 billion humans across the world would be colonized with Blastocystis (C. R. Stensvold, personal communication, March 28, 2013). Although, it was originally named Blastocystis hominis, recent phylogenetic studies suggest to limit the name to “Blastocystis species,” due to the genetic diversity exhibited by the members within the genus.[2] The organism was initially considered to be a commensal, but later observations and studies strongly suggest it to be a pathogen. Although this is supported by strong clinical and scientific evidences, the virulence factors, pathogenicity and other risk factors involved in disease manifestation are still obscure.[3] This review addresses the issues and discusses the recent advances in taxonomy, morphology, life cycle and pathogenic potential of Blastocystis.
TAXONOMY
The various observations made over the years to arrive at the present taxonomical status of Blastocystis would probably be one of the most intriguing stories for any taxonomist. The first observations of the organism were made independently by Brittan and Swayne while studying the infamous cholera epidemic of London in 1849. They had observed various then-unknown parasitic ova and cysts including Blastocystis and collectively tagged them as cholera bodies and suggested them to be the causative agents of cholera. It was only in the early 1900s that the organism was specifically observed by Alexeieff and Emile Brumpt independently, who proposed it to be harmless saprophytic yeast of the intestinal tract. The genus name “Blastocystis” was coined by Alexeieff while Brumpt provided the species name, “hominis.”[4] Over five decades later in 1967, Zierdt et al., reclassified the organism as a Protist based on its morphology and phenotypic properties such as the presence of one or more nuclei, cellular organelles like mitochondria, Golgi apparatus and endoplasmic reticulum, failure to grow on fungal media, resistance to antifungal agents and susceptibility to antiprotozoal drugs.[5]
With the advent of molecular tools, the task of finding a successful taxonomical home for this enigmatic organism was accomplished in the year 1996. By the molecular analysis of small sub-unit rRNA (SSU-rRNA) and elongation factor 1α, Silberman et al., placed the organism under the eukaryotic phylum, Heterokontophyta.[6] Although, this taxonomic status was initially considered controversial by other studies, which showed the resemblance of Blastocystis to Protists, later studies using multiple molecular sequence data confirmed it to be a heterokont, ruling out the taxonomic ambiguity.[7,8] There are over 100,000 members in the phylum Heterokontophyta commonly called as heterokonts or stramenophiles comprising of algae, diatoms, slime moulds and oomycetes, while Blastocystis forms the newer member of this complex group of “botanical Protists.”[1,9] Although this classification seems satisfactory at the molecular level, there is a discrepancy in morphology between Blastocystis and other stramenophiles. In general, the stramenophiles have flagella surrounded by lateral hair like mastigonemes, which is characteristically absent in Blastocystis. Hence there is a proposal to revise the current five kingdom classification and place this organism in a separate sixth kingdom named “Chromista.”[10]
Deducing the identity of the organism to the species level is still an unresolved challenge. Earlier, the species name was given based on the host from which it was isolated such as B. hominis from humans, B. ratti from rats etc., Subsequently, Blastocystis was isolated from diverse hosts as well as human-animal; animal-human transmission was noted.[11] These observations warranted a change in the specific names which prompted various reports attempting to classify the different species of Blastocystis, based on their ultra-structural electron microscopic morphology.[12,13] It was later found that the host specificity and the pathogenic potential of different isolates correlated with sequence variations in the SSU-rRNA. Based on these variations, the members of the genus are classified into several subtypes (ST), which could possibly be termed as species.[11] More recently, the small sub-unit rDNA (SSU-rDNA) was found to have a better correlation with ST and 17 ST have been reported till date.[14] At present it is appropriate to limit the nomenclature of the organism as “Blastocystis species” and it must further be characterized only by molecular subtyping of the SSU-rRNA or SSU-rDNA.[2]
BIOLOGY
Blastocystis is found to colonize the intestinal tract of a wide variety of hosts such as insects, reptiles, birds and mammals.[11] The host specificity seems to correlate with the ST. Among the 17 ST identified, ST1-8 can colonize/infect human as well as non-human hosts, ST9 is found to occur only in humans and ST10-17 are exclusive to non-human hosts.[14]
Blastocystis is a strict anaerobe. Although, numerous intracellular organelles resembling mitochondria are seen, they are completely devoid of cytochrome enzymes.[15] These organelles have the property of both the mitochondria of aerobes and the hydrogenosomes of anaerobes and are involved in various metabolic pathways such as amino acid metabolism, iron-sulfur cluster biogenesis and tricarboxylic acid cycle.[16,17] It is has also been observed that the organism is capable of synthesizing various essential cellular phospholipids and accumulate them within storage vacuoles.[18,19] The generation time of the organism observed in vitro in axenic media is 17-22 h; however, it varies on different media.[20] In cultures on xenic media followed by axenization the generation time is shorter, ranging 7-12 h.[21] Generation time is also found to vary among different strains or ST.[22]
The ability of the organism to undergo apoptosis under adverse conditions has been demonstrated.[23] This phenomenon has been observed when the cells are exposed to aerobic conditions and also in the presence of anti-parasitic agents such as metronidazole.[24,25] It has been postulated that apoptosis serves as a mechanism to increase the number of viable cells during stressed conditions.[26]
MORPHOLOGY
The baffling range of morphological forms of existence identified in Blastocystis species further mystifies the enigma surrounding the organism. It was only very recently identified that Blastocystis species produce cysts which could develop into vegetative forms.[27,28] Apart from the well characterized vacuolar, granular and amoeboid forms, other vegetative forms such as the avacuolar and the multi-vacuolar forms have also been identified.[29] The organism is also known to assume bizarre morphologies such as the medusa head form and chestnut burr cell on exposure to oxygen and in ageing cultures respectively.[4] Various microscopic techniques such as phase contrast microscopy of wet mounts, bright field microscopy of wet mount and stained smears and electron microscopy have been used to study the morphology. Considering the availability of detailed and extensive reviews on the morphology of Blastocystis,[4,30] only the salient features are described here [Figure 1].
Figure 1.
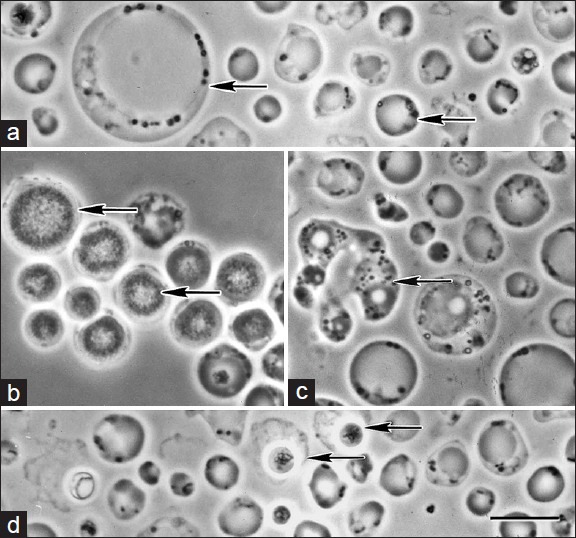
(a) Vacuolar forms of Blastocystis having a large centrally placed vacuole showing extensive variation in size (arrows), (b) Granular forms with distinct granules filling the central body, (c) Amoeboid form with characteristic pseudopodia, (d) Cyst forms. Note the smaller size and the characteristic refractile cyst wall surrounded by loose irregular outer coat. Bar-10 μm. Reproduced from Kevin S. W. Tan (reference 1) with permission
The vacuolar form comprises a large central vacuole which occupies most of the cell space, limiting the cytoplasm and other intracellular components to a thin peripheral rim. This morphological form shows extensive variations in size ranging from 3 μm to 120 μm in diameter [Figure 1a].[31] The size of these forms isolated from humans can range from 4 μm to 63 μm, with the majority of the isolates measuring 5-15 μm in diameter.[32,33] Initial reports identified this morphological form of Blastocystis by wet mount microscopy observing a central empty space and hence labeled it as vacuolar form. Subsequently it has been found that these so called vacuoles are actually membrane bound bodies containing unevenly distributed flocculent or fine granular material made up of carbohydrates and lipids.[32,34,35] Hence certain authors prefer to call it as the central body form, where the central body possibly functions as a storage organelle.[36] In addition to the storage function, studies have identified that the central body plays an integral role in the programmed cell death of the organism.[23]
The peripheral cytoplasmic rim of the central body form usually contains one or two nuclei and rarely more than two.[33] Mitochondria and other organelles are usually seen as rosettes surrounding the nucleus forming thickened pods in the cytoplasmic rim. These “nuclear-organelle pods” cause an inward bulge in the membrane of the central body.[4] Occasionally, the cytoplasm with or without organelles may extrude as membrane bound filaments into the central body. These invaginated cytoplasmic filaments are presumed to be involved in the apoptosis of the organism which is yet to be proven.[37] A surface coat or capsule of varying thickness is seldom seen surrounding the organism, especially in fresh clinical isolates and is lost on subculture. The surface coat serves to protect the organism from osmotic shock and is also postulated to be involved in trapping bacteria for nutrition.[13,38]
The granular form structurally resembles the vacuolar form except for the presence of granules in the central body and the cytoplasm [Figure 1b]. Electron microscopic studies have identified three types of granules namely metabolic, reproductory and lipid granules. The metabolic granules are exclusively found in the cytoplasm and are involved to carry out the various metabolic pathways of the organism. The reproductory granules are seen only within the central body and the suggestion of their role in schizogony is currently considered obsolete.[39,40] The lipid granules serve as storage bodies and are seen both in the cytoplasm and in the central body.[41] The granular forms show a lesser degree of pleomorphism as compared to the vacuolar forms. The average diameter of the granular forms ranges from 15 μm to 25 μm and the largest is reported to be 80 μm.[42,43]
The amoeboid form is comparatively less frequently identified than the vacuolar and granular forms. They are irregular in shape and usually measure around 10 μm in size [Figure 1c].[41] Although the possession of one or two pseudopodia is a characteristic feature, the amoeboid forms are non-motile.[1] The cytoplasm may either contain a single large vacuole as that of the central body form or have multiple smaller vacuoles.[44] It has been presumed that the central body form acquires an intermediate amoeboid form before conversion to cysts during which they feed on bacteria to acquire nutrition for encystment.[45] As these forms are increasingly being identified in symptomatic patients with diarrhea, recent reports suggest the possibility of amoeboid forms being pathogenic.[46,47] Due to their small size and resemblance to neutrophils and macrophages these forms are easily missed during stool examination. To differentiate them, Zierdt suggested performing a gram stain of air-dried unfixed smear where the amoeboid forms lyse on exposure to air while the leucocytes remain intact.[4]
The cyst form is the most recently described form of the parasite. They are spherical to ovoid in shape and smaller than the vacuolar and the granular forms [Figure 1d]. Cysts isolated from humans are usually 3-6 μm in diameter and do not exceed 10 μm while larger cysts have been isolated from animal hosts.[12,48] The distinct feature of the cyst form is the presence of a thick multi-layered cyst wall. In addition to the characteristic cyst wall, the newly formed cysts may in turn be surrounded by the surface coat of the vacuolar form from which they develop or they might lose the surface coat and appear as naked cysts.[49,50] The cytoplasm is condensed and contains varying numbers of mitochondria and small vacuoles made of lipids or glycogen. The cysts isolated from humans are commonly binucleate; however, the number of nuclei can vary from one to four.[27] The cyst form confers protection to the parasite during adverse conditions and are found to remain viable for up to 1 month at 25°C even on exposure to air. The cysts are now proven to be the transmissible infective forms, which on entering a suitable host develop into vacuolar forms.[28,51]
Studies have shown that each of the vegetative form can freely transform into other vegetative forms as a random, continuous process occurring in vivo. As a result of this transformation process, a plethora of intermediate forms are usually present in a fresh faecal specimen. But as they tend to assume various appearances, the morphology of these intermediate forms could not be well characterized and are hence often overlooked.[50]
Although the classically described vacuolar and granular forms are customarily used in the microscopic identification of the organism, recent studies have questioned their in vivo significance. By a study using supravital staining techniques, Vdovenko demonstrated that live organisms from fresh cultures were uniformly stained. Organisms stained after a time delay showed granular and vacuolar morphology with reduced intensity of staining over time. These observations suggest that the granular and vacuolar forms could indicate degenerative changes or fixation artifacts.[52] In this context, the other morphological forms such as the avacuolar and the multi-vacuolar forms are gaining importance. These forms are smaller, measuring 5-8 μm in diameter and extreme variations in size does not occur. The avacuolar form is devoid of the central body while the multi-vacuolar forms have multiple small vacuoles of different sizes interconnected to each other or lying discrete in the cytoplasm.[29] They are usually uninucleate but are occasionally found to be binucleate. The nuclei of the avacuolar forms are larger than that of any other morphological forms.[30] While the old school of thought considers these forms as intermediates during transformation of the common vegetative forms, recent observations suggest the avacuolar and the multi-vacuolar forms to be the predominant forms in vivo and are frequently missed during microscopic examination due to the lack of awareness about them.[28,29] The occurrence of the various morphological forms is summarized in Table 1.
Table 1.
Occurrence of the various morphological forms of Blastocystis in vivo and in vitro

LIFE CYCLE AND MODE OF REPRODUCTION
The life cycle of Blastocystis is not yet elucidated with surety, due to the lack of a proper animal model. However, some of the assumptions and misleading observations of the past have been proved wrong. Infectivity studies conducted on BALB/c mice and Wistar rats have proved that the cysts are the only transmissible forms of Blastocystis and are transmitted through the faeco-oral route.[51,62] Studies have revealed that suitable hosts could contract Blastocystis infection by drinking untreated water or eating raw aquatic plants contaminated with cysts.[63,64] Recent studies demonstrate that unclean hands can serve as fomites for transmission of cysts from infected individuals on direct contact or from contaminated soil.[53] Figure 2 depicts the most probable life cycle of Blastocystis agreed by general consensus.
Figure 2.
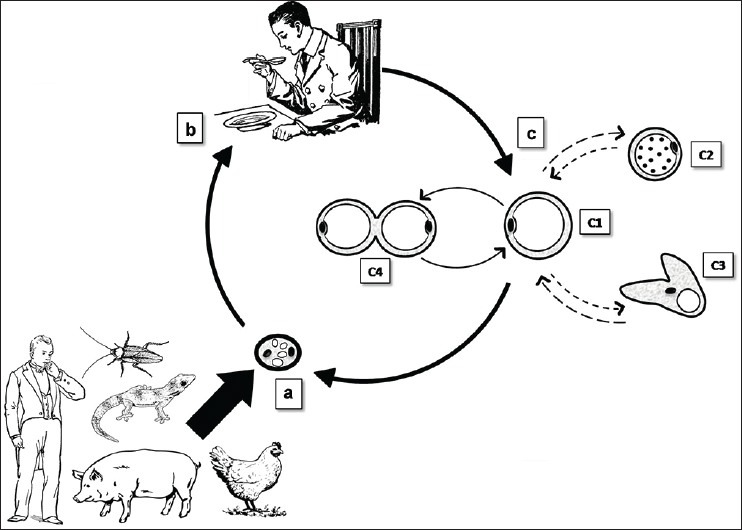
(a) Cyst forms are excreted in the faeces of infected hosts, (b) Man acquires infection by consumption of food/water contaminated with faeces containing cysts, (c) Excystation occurs in the large intestine to release the vacuolar form. The vacuolar form (c1) can transform into the granular form (c2) or the amoeboid form (c3) and vice versa. The vacuolar form multiplies by binary fission (other modes of reproduction such as plasmotomy and budding can also occur). The vacuolar form undergoes encystation in the lumen of the large intestine to produce the cyst form which is shed in the faeces
Upon ingestion, the cysts develop into vegetative forms only in the suitable host. The further continuation of the life cycle depends on the compatibility of the subtype with the host. The cyst form undergoes excystation in the large intestine to liberate the vacuolar form.[62] The vacuolar forms can transform into any of the other forms. Frequent observations of the amoeboid, avacuolar and multi-vacuolar forms in diarrhea suggest a possibility that these forms might play a role in the pathogenesis.[46] The vacuolar forms encyst in the intestinal lumen to form cysts which are shed in the faeces for further transmission.
Various authors have claimed to have observed either of the modes of reproduction such as binary fission, budding, plasmotomy, multiple fission, endodyogeny and schizogony.[59] Binary fission of the vacuolar forms is the most commonly observed and well established mode of reproduction.[28] Recent studies have confirmed that budding and plasmotomy are the other mechanisms of reproduction in Blastocystis.[46] Earlier observations and assumptions of the existence of thick and thin walled cysts which undergo multiple fission to release the progeny are inconclusive due to the lack of scientific evidence.[1] The possible role played by the reproductory granules of the central body in endodyogeny and schizogony is elusive as these modes of reproduction have not been supported by further studies.[30]
VIRULENCE AND CLINICAL SIGNIFICANCE
The possibility of Blastocystis being a pathogen has long been a matter of debate. Numerous studies done across the world with a motive to identify the clinical significance of the organism have produced contradicting results, thereby adding to the existing controversy.[65] Although recent accumulation of clinical evidence suggests the pathogenic potential of Blastocystis, one must bear in mind that it is still not proven conclusively till date.[3]
At present, the most convincing yet unsatisfactory explanation to the pathogenicity of Blastocystis is the correlation of the ST with virulence.[66] Initial studies determining the subtype pathogenicity reported that ST3 had a strong correlation with symptomatic disease.[67] Subsequently the ST ST1, 2, 4 and 6 have also been isolated from symptomatic patients.[68,69] However, ST3 is the commonest subtype reported from symptomatic patients, followed by ST1 and ST2.[69,70] Intra subtype variations in pathogenicity has also been noted, that is, not all the strains of a particular subtype are pathogenic.[3] These observations suggest that subtyping alone does not predict the pathogenicity.[68]
The phenomenon of intra subtype variation in pathogenicity is currently explained by the phenotypic differences between the pathogenic and non-pathogenic members. The most studied phenotypic disparities of the pathogenic members are the predominance of amoeboid forms and the secretion of proteases.[47,71] The suspicion of the amoeboid forms being virulent had raised long ago when numerous of these forms were demonstrated in a patient with severe diarrhea.[72] Subsequent reports with similar findings seems to reinforce this further.[57] Tan and Suresh performed a comparative study by culturing Blastocystis isolates from symptomatic and asymptomatic individuals and observed the predominance of amoeboid forms in symptomatic patients.[47] An interesting observation supporting the virulence of amoeboid forms was made by Vassalos et al., while studying the intra subtype variations in ST3. In this study, one of the patients who was an asymptomatic carrier of vacuolar and granular forms of ST3 turned symptomatic within a short period, shedding amoeboid forms in his stool. Hence, the authors recommend performing a specific search for amoeboid forms while screening stool samples of symptomatic patients.[56]
Secretion of proteases and other hydrolytic enzymes by Blastocystis have been identified by polyacrylamide gel electrophoresis and attributed to be responsible for the pathogenesis of gastrointestinal symptoms.[71] Attempts have been made to deduce the functions of these extra-cellular virulence factors by in vitro studies. Blastocystis culture lysates have been found to produce cytoskeletal alterations and induce apoptosis in epithelial cells, which results in increased permeability.[73] Cysteine proteases secreted by the organism are known to stimulate the intestinal mucosal cell to produce interleukin-8.[74] These mechanisms are suggested to be responsible for the fluid loss and intestinal inflammation in affected individuals. Puthia et al., have observed the ability of a cysteine protease to cleave human secretory immunoglobulin A thereby helping in immune evasion and promoting parasite survival in vivo.[75]
Whole genome sequencing has been done for Blastocystis ST7, which has unraveled various known and unknown genes. Genes coding for proteins responsible for host protease inhibition have been identified and predicted to be secreted. These proteins are presumed to modulate the host protease activity thereby disturbing the intestinal homeostasis. Also recognized are the genes responsible for the production of non-ribosomal peptides and polyketides which are antibacterial and capable of causing dysbiosis of the protective intestinal microbiome.[76] Genetic targets coding for hydrolases have been identified, which on expression are capable of altering the protective mucus layer and exposing the surface of the intestinal epithelium for adhesion.[77] Expression of serine protease and glycosyltransferases is believed to disrupt the tight junctions of the intestinal mucosal epithelium resulting in increased permeability.[76] However, genomic studies must be followed up with protein expression studies to confirm the findings.
The parasitic molecules responsible for extra-intestinal manifestations are not yet identified. It has been hypothesized that like other parasitic antigens, the antigens of Blastocystis stimulate the T-Helper 2 Th2 cells producing immunoglobulin E mediated allergic response.[78] It is also speculated that Blastocystis activates the complement cascade to release anaphylatoxins which interact with the mast cells to convene skin related disorders.[79] The virulence factors of Blastocystis which cause iron deficiency anemia are not yet identified.[80]
Apart from these, the other features exclusive to virulent strains are large size, rough surface, slow growth rate and increased binding affinity to lectins such as concanavalin and Helix pomatia agglutinins.[67,81,82] Some of the reports attributing the causal role of Blastocystis in diseases have mentioned the direct correlation of infection density with severity of clinical symptoms and duration of illness.[81,83] Although, these are significant observations made, their relevance in pathogenicity must be substantiated by scientific evidence. Table 2 summarizes the possible clinical events which may occur with Blastocystis infection in humans.
Table 2.
The reported clinical manifestations of Blastocystis infection

CONCLUSIONS
Although Blastocystis was identified over a century ago, most of the details about it are still indistinct. The current taxonomical status assigned to the parasite seems to be appropriate for the time being, but is not completely agreeable. Various morphological forms of existence have been identified and described in detail, but their in vivo significance is still anonymous. The life cycle of the parasite has been proposed with bits and pieces of available facts and is yet to be elucidated with confirmation. Genomic studies have identified various putative genes responsible for the physiology and virulence of the organism, but this could only be speculative until confirmed by protein expression studies.[77]
Observations on the pathogenic potential of the organism have been constantly refuted by similar studies, which state the causal role to be statistically insignificant. Since the members of the genus exhibit a vast genetic diversity, several molecular modalities of subtyping have been devised. Numerous studies on assigning the pathogenic potential to the molecular ST are available, but they could not be compared or analyzed with an effective meta-analysis due to the different molecular techniques employed. At present, subtyping based on molecular differences between SSU-rRNA or SSU-rDNA has been uniformly accepted. But this method too has drawbacks while assigning virulence to specific ST, as intra subtype variations in virulence have been observed. Hence a fail proof molecular typing method, which correlates with the phenotype is still unavailable.[66]
Attempts to get a clear insight on various aspects of Blastocystis has been obscured due to the existence of vast intra generic genetic diversity, morphological heterogenicity, lack of a proper animal model and the tendency to associate the organism to an existing unanswered disease.[3,89] it is hoped that the ambiguity surrounding the organism would be cleared over the forthcoming years with sound scientific studies.
Footnotes
Source of Support: Nil
Conflict of Interest: None declared
REFERENCES
- 1.Tan KS. New insights on classification, identification, and clinical relevance of Blastocystis spp. Clin Microbiol Rev. 2008;21:639–65. doi: 10.1128/CMR.00022-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Stensvold CR, Suresh GK, Tan KS, Thompson RC, Traub RJ, Viscogliosi E, et al. Terminology for Blastocystis subtypes: A consensus. Trends Parasitol. 2007;23:93–6. doi: 10.1016/j.pt.2007.01.004. [DOI] [PubMed] [Google Scholar]
- 3.Scanlan PD. Blastocystis: Past pitfalls and future perspectives. Trends Parasitol. 2012;28:327–34. doi: 10.1016/j.pt.2012.05.001. [DOI] [PubMed] [Google Scholar]
- 4.Zierdt CH. Blastocystis hominis: Past and future. Clin Microbiol Rev. 1991;4:61–79. doi: 10.1128/cmr.4.1.61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Zierdt CH, Rude WS, Bull BS. Protozoan characteristics of Blastocystis hominis. Am J Clin Pathol. 1967;48:495–501. doi: 10.1093/ajcp/48.5.495. [DOI] [PubMed] [Google Scholar]
- 6.Silberman JD, Sogin ML, Leipe DD, Clark CG. Human parasite finds taxonomic home. Nature. 1996;380:398. doi: 10.1038/380398a0. [DOI] [PubMed] [Google Scholar]
- 7.Ho LC, Armiugam A, Jeyaseelan K, Yap EH, Singh M. Blastocystis elongation factor-1alpha: Genomic organization, taxonomy and phylogenetic relationships. Parasitology. 2000;121:135–44. doi: 10.1017/s0031182099006113. [DOI] [PubMed] [Google Scholar]
- 8.Arisue N, Hashimoto T, Yoshikawa H, Nakamura Y, Nakamura G, Nakamura F, et al. Phylogenetic position of Blastocystis hominis and of stramenopiles inferred from multiple molecular sequence data. J Eukaryot Microbiol. 2002;49:42–53. doi: 10.1111/j.1550-7408.2002.tb00339.x. [DOI] [PubMed] [Google Scholar]
- 9.Riisberg I, Orr RJ, Kluge R, Shalchian-Tabrizi K, Bowers HA, Patil V, et al. Seven gene phylogeny of heterokonts. Protist. 2009;160:191–204. doi: 10.1016/j.protis.2008.11.004. [DOI] [PubMed] [Google Scholar]
- 10.Cavalier-Smith T. A revised six-kingdom system of life. Biol Rev Camb Philos Soc. 1998;73:203–66. doi: 10.1017/s0006323198005167. [DOI] [PubMed] [Google Scholar]
- 11.Noël C, Dufernez F, Gerbod D, Edgcomb VP, Delgado-Viscogliosi P, Ho LC, et al. Molecular phylogenies of Blastocystis isolates from different hosts: Implications for genetic diversity, identification of species, and zoonosis. J Clin Microbiol. 2005;43:348–55. doi: 10.1128/JCM.43.1.348-355.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Stenzel DJ, Lee MG, Boreham PF. Morphological differences in Blastocystis cysts-an indication of different species? Parasitol Res. 1997;83:452–7. doi: 10.1007/s004360050279. [DOI] [PubMed] [Google Scholar]
- 13.Cassidy MF, Stenzel DJ, Boreham PF. Electron microscopy of surface structures of Blastocystis sp. from different hosts. Parasitol Res. 1994;80:505–11. doi: 10.1007/BF00932698. [DOI] [PubMed] [Google Scholar]
- 14.Stensvold CR. Blastocystis-genetic diversity and molecular methods for diagnosis and epidemiology. Trop Parasitol. 2013;3:26–34. doi: 10.4103/2229-5070.113896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Zierdt CH. Cytochrome-free mitochondria of an anaerobic protozoan: Blastocystis hominis. J Protozool. 1986;33:67–9. doi: 10.1111/j.1550-7408.1986.tb05559.x. [DOI] [PubMed] [Google Scholar]
- 16.Stechmann A, Hamblin K, Pérez-Brocal V, Gaston D, Richmond GS, van der Giezen M, et al. Organelles in Blastocystis that blur the distinction between mitochondria and hydrogenosomes. Curr Biol. 2008;18:580–5. doi: 10.1016/j.cub.2008.03.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Tsaousis AD, Ollagnier de Choudens S, Gentekaki E, Long S, Gaston D, Stechmann A, et al. Evolution of Fe/S cluster biogenesis in the anaerobic parasite Blastocystis. Proc Natl Acad Sci U S A. 2012;109:10426–31. doi: 10.1073/pnas.1116067109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Keenan TW, Zierdt CH. Lipid biosynthesis by axenic strains of Blastocystis hominis. Comp Biochem Physiol Biochem Mol Biol. 1994;107:525–31. doi: 10.1016/0305-0491(94)90180-5. [DOI] [PubMed] [Google Scholar]
- 19.Zierdt CH. Blastocystis hominis, a long-misunderstood intestinal parasite. Parasitol Today. 1988;4:15–7. doi: 10.1016/0169-4758(88)90049-x. [DOI] [PubMed] [Google Scholar]
- 20.Irikov OA, Antokhin AI, Romanov YA. Study of the dynamics of Blastocystis hominis reproduction in vitro. Bull Exp Biol Med. 2009;148:99–102. doi: 10.1007/s10517-009-0651-7. [DOI] [PubMed] [Google Scholar]
- 21.Lanuza MD, Carbajal JA, Villar J, Borrás R. Description of an improved method for Blastocystis hominis culture and axenization. Parasitol Res. 1997;83:60–3. doi: 10.1007/s004360050209. [DOI] [PubMed] [Google Scholar]
- 22.Zierdt CH, Swan JC. Generation time and growth rate of the human intestinal parasite Blastocystis hominis. J Protozool. 1981;28:483–5. doi: 10.1111/j.1550-7408.1981.tb05324.x. [DOI] [PubMed] [Google Scholar]
- 23.Nasirudeen AM, Tan KS, Singh M, Yap EH. Programmed cell death in a human intestinal parasite, Blastocystis hominis. Parasitology. 2001;123:235–46. doi: 10.1017/s0031182001008332. [DOI] [PubMed] [Google Scholar]
- 24.Tan KS, Howe J, Yap EH, Singh M. Do Blastocystis hominis colony forms undergo programmed cell death? Parasitol Res. 2001;87:362–7. doi: 10.1007/s004360000364. [DOI] [PubMed] [Google Scholar]
- 25.Nasirudeen AM, Hian YE, Singh M, Tan KS. Metronidazole induces programmed cell death in the protozoan parasite Blastocystis hominis. Microbiology. 2004;150:33–43. doi: 10.1099/mic.0.26496-0. [DOI] [PubMed] [Google Scholar]
- 26.Dhurga DB, Suresh KG, Tan TC, Chandramathi S. Apoptosis in Blastocystis spp. is related to subtype. Trans R Soc Trop Med Hyg. 2012;106:725–30. doi: 10.1016/j.trstmh.2012.08.005. [DOI] [PubMed] [Google Scholar]
- 27.Stenzel DJ, Boreham PF. A cyst-like stage of Blastocystis hominis. Int J Parasitol. 1991;21:613–5. doi: 10.1016/0020-7519(91)90069-j. [DOI] [PubMed] [Google Scholar]
- 28.Moe KT, Singh M, Howe J, Ho LC, Tan SW, Chen XQ, et al. Development of Blastocystis hominis cysts into vacuolar forms in vitro. Parasitol Res. 1999;85:103–8. doi: 10.1007/s004360050517. [DOI] [PubMed] [Google Scholar]
- 29.Stenzel DJ, Boreham PF, McDougall R. Ultrastructure of Blastocystis hominis in human stool samples. Int J Parasitol. 1991;21:807–12. doi: 10.1016/0020-7519(91)90149-2. [DOI] [PubMed] [Google Scholar]
- 30.Stenzel DJ, Boreham PF. Blastocystis hominis revisited. Clin Microbiol Rev. 1996;9:563–84. doi: 10.1128/cmr.9.4.563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Lee MG, Stenzel DJ. A survey of Blastocystis in domestic chickens. Parasitol Res. 1999;85:109–17. doi: 10.1007/s004360050518. [DOI] [PubMed] [Google Scholar]
- 32.Dunn LA, Boreham PF, Stenzel DJ. Ultrastructural variation of Blastocystis hominis stocks in culture. Int J Parasitol. 1989;19:43–56. doi: 10.1016/0020-7519(89)90020-9. [DOI] [PubMed] [Google Scholar]
- 33.MacPherson DW, MacQueen WM. Morphological diversity of Blastocystis hominis in sodium acetate-acetic acid-formalin-preserved stool samples stained with iron hematoxylin. J Clin Microbiol. 1994;32:267–8. doi: 10.1128/jcm.32.1.267-268.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Yoshikawa H, Kuwayama N, Enose Y. Histochemical detection of carbohydrates of Blastocystis hominis. J Eukaryot Microbiol. 1995;42:70–4. doi: 10.1111/j.1550-7408.1995.tb01542.x. [DOI] [PubMed] [Google Scholar]
- 35.Yoshikawa H, Satoh J, Enose Y. Light and electron microscopic localization of lipids in Blastocystis hominis. J Electron Microsc (Tokyo) 1995;44:100–3. [PubMed] [Google Scholar]
- 36.Stenzel DJ, Dunn LA, Boreham PF. Endocytosis in cultures of Blastocystis hominis. Int J Parasitol. 1989;19:787–91. doi: 10.1016/0020-7519(89)90067-2. [DOI] [PubMed] [Google Scholar]
- 37.Tan KS, Nasirudeen AM. Protozoan programmed cell death: Insights from Blastocystis deathstyles. Trends Parasitol. 2005;21:547–50. doi: 10.1016/j.pt.2005.09.006. [DOI] [PubMed] [Google Scholar]
- 38.Zaman V, Howe J, Ng M. Observations on the surface coat of Blastocystis hominis. Parasitol Res. 1997;83:731–3. doi: 10.1007/s004360050329. [DOI] [PubMed] [Google Scholar]
- 39.Suresh K, Howe J, Ng GC, Ho LC, Ramachandran NP, Loh AK, et al. A multiple fission-like mode of asexual reproduction in Blastocystis hominis. Parasitol Res. 1994;80:523–7. doi: 10.1007/BF00932701. [DOI] [PubMed] [Google Scholar]
- 40.Tan KS, Stenzel DJ. Multiple reproductive processes in Blastocystis: Proceed with caution. Trends Parasitol. 2003;19:290–1. doi: 10.1016/s1471-4922(03)00119-3. [DOI] [PubMed] [Google Scholar]
- 41.Tan HK, Zierdt CH. Ultrastructure of Blastocystis hominis. Z Parasitenkd. 1973;42:315–24. doi: 10.1007/BF00328892. [DOI] [PubMed] [Google Scholar]
- 42.Zierdt CH. Studies of Blastocystis hominis. J Protozool. 1973;20:114–21. doi: 10.1111/j.1550-7408.1973.tb06013.x. [DOI] [PubMed] [Google Scholar]
- 43.Zierdt CH, Williams RL. Blastocystis hominis: Axenic cultivation. Exp Parasitol. 1974;36:233–43. doi: 10.1016/0014-4894(74)90062-9. [DOI] [PubMed] [Google Scholar]
- 44.Tan TC, Suresh KG. Amoeboid form of Blastocystis hominis-A detailed ultrastructural insight. Parasitol Res. 2006;99:737–42. doi: 10.1007/s00436-006-0214-z. [DOI] [PubMed] [Google Scholar]
- 45.Singh M, Suresh K, Ho LC, Ng GC, Yap EH. Elucidation of the life cycle of the intestinal protozoan Blastocystis hominis. Parasitol Res. 1995;81:446–50. doi: 10.1007/BF00931510. [DOI] [PubMed] [Google Scholar]
- 46.Zhang X, Zhang S, Qiao J, Wu X, Zhao L, Liu Y, et al. Ultrastructural insights into morphology and reproductive mode of Blastocystis hominis. Parasitol Res. 2012;110:1165–72. doi: 10.1007/s00436-011-2607-x. [DOI] [PubMed] [Google Scholar]
- 47.Tan TC, Suresh KG, editors. Predominance of amoeboid forms of Blastocystis hominis in isolates from symptomatic patients. Parasitol Res. 2006;98:189–93. doi: 10.1007/s00436-005-0033-7. [DOI] [PubMed] [Google Scholar]
- 48.Zaman V, Howe J, Ng M. Ultrastructure of Blastocystis hominis cysts. Parasitol Res. 1995;81:465–9. doi: 10.1007/BF00931787. [DOI] [PubMed] [Google Scholar]
- 49.Zaman V, Howe J, Ng M. Scanning electron microscopy of Blastocystis hominis cysts. Parasitol Res. 1998;84:476–7. doi: 10.1007/s004360050432. [DOI] [PubMed] [Google Scholar]
- 50.Zhang X, Qiao JY, Dong XH, Li YQ, Li XQ, Li C. Study on morphology of Blastocystis hominis in culture and from diarrhea patients. Zhongguo Ji Sheng Chong Xue Yu Ji Sheng Chong Bing Za Zhi. 2003;21:116–8. [PubMed] [Google Scholar]
- 51.Yoshikawa H, Yoshida K, Nakajima A, Yamanari K, Iwatani S, Kimata I. Fecal-oral transmission of the cyst form of Blastocystis hominis in rats. Parasitol Res. 2004;94:391–6. doi: 10.1007/s00436-004-1230-5. [DOI] [PubMed] [Google Scholar]
- 52.Vdovenko AA. Blastocystis hominis: Origin and significance of vacuolar and granular forms. Parasitol Res. 2000;86:8–10. doi: 10.1007/pl00008506. [DOI] [PubMed] [Google Scholar]
- 53.Anuar TS, Ghani MK, Azreen SN, Salleh FM, Moktar N. Blastocystis infection in Malaysia: Evidence of waterborne and human-to-human transmissions among the Proto-Malay, Negrito and Senoi tribes of Orang Asli. Parasit Vectors. 2013;6:40. doi: 10.1186/1756-3305-6-40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Roberts T, Barratt J, Harkness J, Ellis J, Stark D. Comparison of microscopy, culture, and conventional polymerase chain reaction for detection of Blastocystis sp. in clinical stool samples. Am J Trop Med Hyg. 2011;84:308–12. doi: 10.4269/ajtmh.2011.10-0447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Zhang X, Qiao J, Wu X, Da R, Zhao L, Wei Z. In vitro culture of Blastocystis hominis in three liquid media and its usefulness in the diagnosis of Blastocystosis. Int J Infect Dis. 2012;16:e23–8. doi: 10.1016/j.ijid.2011.09.012. [DOI] [PubMed] [Google Scholar]
- 56.Vassalos CM, Spanakos G, Vassalou E, Papadopoulou C, Vakalis N. Differences in clinical significance and morphologic features of Blastocystis sp subtype 3. Am J Clin Pathol. 2010;133:251–8. doi: 10.1309/AJCPDOWQSL6E8DMN. [DOI] [PubMed] [Google Scholar]
- 57.Katsarou-Katsari A, Vassalos CM, Tzanetou K, Spanakos G, Papadopoulou C, Vakalis N. Acute urticaria associated with amoeboid forms of Blastocystis sp. subtype 3. Acta Derm Venereol. 2008;88:80–1. doi: 10.2340/00015555-0338. [DOI] [PubMed] [Google Scholar]
- 58.Tan SW, Singh M, Thong KT, Ho LC, Moe KT, Chen XQ, et al. Clonal growth of Blastocystis hominis in soft agar with sodium thioglycollate. Parasitol Res. 1996;82:737–9. doi: 10.1007/s004360050194. [DOI] [PubMed] [Google Scholar]
- 59.Zhang X, Qiao JY, Zhou XJ, Yao FR, Wei ZC. Morphology and reproductive mode of Blastocystis hominis in diarrhea and in vitro. Parasitol Res. 2007;101:43–51. doi: 10.1007/s00436-006-0439-x. [DOI] [PubMed] [Google Scholar]
- 60.Suresh K, Ng GC, Ramachandran NP, Ho LC, Yap EH, Singh M. In vitro encystment and experimental infections of Blastocystis hominis. Parasitol Res. 1993;79:456–60. doi: 10.1007/BF00931582. [DOI] [PubMed] [Google Scholar]
- 61.Silard R, Burghelea B. Ultrastructural aspects of Blastocystis hominis strain resistant to antiprotozoal drugs. Arch Roum Pathol Exp Microbiol. 1985;44:73–85. [PubMed] [Google Scholar]
- 62.Moe KT, Singh M, Howe J, Ho LC, Tan SW, Chen XQ, et al. Experimental Blastocystis hominis infection in laboratory mice. Parasitol Res. 1997;83:319–25. doi: 10.1007/s004360050256. [DOI] [PubMed] [Google Scholar]
- 63.Lee LI, Chye TT, Karmacharya BM, Govind SK. Blastocystis sp.: Waterborne zoonotic organism, a possibility? Parasit Vectors. 2012;5:130. doi: 10.1186/1756-3305-5-130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Li LH, Zhou XN, Du ZW, Wang XZ, Wang LB, Jiang JY, et al. Molecular epidemiology of human Blastocystis in a village in Yunnan province, China. Parasitol Int. 2007;56:281–6. doi: 10.1016/j.parint.2007.06.001. [DOI] [PubMed] [Google Scholar]
- 65.Clark CG. Extensive genetic diversity in Blastocystis hominis. Mol Biochem Parasitol. 1997;87:79–83. doi: 10.1016/s0166-6851(97)00046-7. [DOI] [PubMed] [Google Scholar]
- 66.Tan KS, Mirza H, Teo JD, Wu B, Macary PA. Current Views on the Clinical Relevance of Blastocystis spp. Curr Infect Dis Rep. 2010;12:28–35. doi: 10.1007/s11908-009-0073-8. [DOI] [PubMed] [Google Scholar]
- 67.Tan TC, Suresh KG, Smith HV. Phenotypic and genotypic characterisation of Blastocystis hominis isolates implicates subtype 3 as a subtype with pathogenic potential. Parasitol Res. 2008;104:85–93. doi: 10.1007/s00436-008-1163-5. [DOI] [PubMed] [Google Scholar]
- 68.Nagel R, Cuttell L, Stensvold CR, Mills PC, Bielefeldt-Ohmann H, Traub RJ. Blastocystis subtypes in symptomatic and asymptomatic family members and pets and response to therapy. Intern Med J. 2012;42:1187–95. doi: 10.1111/j.1445-5994.2011.02626.x. [DOI] [PubMed] [Google Scholar]
- 69.Dogruman-Al F, Kustimur S, Yoshikawa H, Tuncer C, Simsek Z, Tanyuksel M, et al. Blastocystis subtypes in irritable bowel syndrome and inflammatory bowel disease in Ankara, Turkey. Mem Inst Oswaldo Cruz. 2009;104:724–7. doi: 10.1590/s0074-02762009000500011. [DOI] [PubMed] [Google Scholar]
- 70.Moosavi A, Haghighi A, Mojarad EN, Zayeri F, Alebouyeh M, Khazan H, et al. Genetic variability of Blastocystis sp. isolated from symptomatic and asymptomatic individuals in Iran. Parasitol Res. 2012;111:2311–5. doi: 10.1007/s00436-012-3085-5. [DOI] [PubMed] [Google Scholar]
- 71.Abdel-Hameed DM, Hassanin OM. Proteaese activity of Blastocystis hominis subtype 3 in symptomatic and asymptomatic patients. Parasitol Res. 2011;109:321–7. doi: 10.1007/s00436-011-2259-x. [DOI] [PubMed] [Google Scholar]
- 72.Zierdt CH, Tan HK. Ultrastructure and light microscope appearance of Blastocystis hominis in a patient with enteric disease. Z Parasitenkd. 1976;50:277–83. doi: 10.1007/BF02462972. [DOI] [PubMed] [Google Scholar]
- 73.Puthia MK, Sio SW, Lu J, Tan KS. Blastocystis ratti induces contact-independent apoptosis, F-actin rearrangement, and barrier function disruption in IEC-6 cells. Infect Immun. 2006;74:4114–23. doi: 10.1128/IAI.00328-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Puthia MK, Lu J, Tan KS. Blastocystis ratti contains cysteine proteases that mediate interleukin-8 response from human intestinal epithelial cells in an NF-kappaB-dependent manner. Eukaryot Cell. 2008;7:435–43. doi: 10.1128/EC.00371-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Puthia MK, Vaithilingam A, Lu J, Tan KS. Degradation of human secretory immunoglobulin A by Blastocystis. Parasitol Res. 2005;97:386–9. doi: 10.1007/s00436-005-1461-0. [DOI] [PubMed] [Google Scholar]
- 76.Denoeud F, Roussel M, Noel B, Wawrzyniak I, Da Silva C, Diogon M, et al. Genome sequence of the stramenopile Blastocystis, a human anaerobic parasite. Genome Biol. 2011;12:R29. doi: 10.1186/gb-2011-12-3-r29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Poirier P, Wawrzyniak I, Vivarès CP, Delbac F, El Alaoui H. New insights into Blastocystis spp.: A potential link with irritable bowel syndrome. PLoS Pathog. 2012;8:e1002545. doi: 10.1371/journal.ppat.1002545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Pasqui AL, Savini E, Saletti M, Guzzo C, Puccetti L, Auteri A. Chronic urticaria and blastocystis hominis infection: A case report. Eur Rev Med Pharmacol Sci. 2004;8:117–20. [PubMed] [Google Scholar]
- 79.Valsecchi R, Leghissa P, Greco V. Cutaneous lesions in Blastocystis hominis infection. Acta Derm Venereol. 2004;84:322–3. doi: 10.1080/00015550410025949. [DOI] [PubMed] [Google Scholar]
- 80.Yavasoglu I, Kadikoylu G, Uysal H, Ertug S, Bolaman Z. Is Blastocystis hominis a new etiologic factor or a coincidence in iron deficiency anemia? Eur J Haematol. 2008;81:47–50. doi: 10.1111/j.1600-0609.2008.01080.x. [DOI] [PubMed] [Google Scholar]
- 81.Hegazy MM, Maklouf LM, El Hamshary EM, Dawoud HA, Eida AM. Protein profile and morphometry of cultured human Blastocystis hominis from children with gastroenteritis and healthy ones. J Egypt Soc Parasitol. 2008;38:453–64. [PubMed] [Google Scholar]
- 82.Vassalos CM, Vakalis N, Papadopoulou C. Blastocystis and its pathogenic potential: latest aspects. Rev Med Microbiol. 2008;19:87–97. [Google Scholar]
- 83.Giacometti A, Cirioni O, Antonicelli L, D’Amato G, Silvestri C, Del Prete MS, et al. Prevalence of intestinal parasites among individuals with allergic skin diseases. J Parasitol. 2003;89:490–2. doi: 10.1645/0022-3395(2003)089[0490:POIPAI]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- 84.Cekin AH, Cekin Y, Adakan Y, Tasdemir E, Koclar FG, Yolcular BO. Blastocystosis in patients with gastrointestinal symptoms: A case-control study. BMC Gastroenterol. 2012;12:122. doi: 10.1186/1471-230X-12-122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Gupta R, Parsi K. Chronic urticaria due to Blastocystis hominis. Australas J Dermatol. 2006;47:117–9. doi: 10.1111/j.1440-0960.2006.00244.x. [DOI] [PubMed] [Google Scholar]
- 86.Kick G, Rueff F, Przybilla B. Palmoplantar pruritus subsiding after Blastocystis hominis eradication. Acta Derm Venereol. 2002;82:60. doi: 10.1080/000155502753600948. [DOI] [PubMed] [Google Scholar]
- 87.Micheloud D, Jensen J, Fernandez-Cruz E, Carbone J. Chronic angioedema and blastocystis hominis infection. Rev Gastroenterol Peru. 2007;27:191–3. [PubMed] [Google Scholar]
- 88.El deeb H, Khodeer S, Blastocystis SP. Frequency and subtype distribution in iron deficiency anemic versus non-anemic subjects from Egypt. J Parasitol. 2013 doi: 10.1645/12-80.1. In Press. [DOI] [PubMed] [Google Scholar]
- 89.Tan KS, Singh M, Yap EH. Recent advances in Blastocystis hominis research: Hot spots in terra incognita. Int J Parasitol. 2002;32:789–804. doi: 10.1016/s0020-7519(02)00005-x. [DOI] [PubMed] [Google Scholar]