

Abstract
In the recent years, there has been an increase in the number of metabolomic approaches used, in parallel with proteomic and functional genomic studies. The wide variety of chemical types of metabolites available has also accelerated the use of different techniques in the investigation of the metabolome. At present, metabolomics is applied to investigate several human diseases, to improve their diagnosis and prevention, and to design better therapeutic strategies. In addition, metabolomic studies are also being carried out in areas such as toxicology and pharmacology, crop breeding, and plant biotechnology. In this review, we emphasize the use and application of metabolomics in human diseases and plant research to improve human health.
1. Introduction
Metabolomics is the comprehensive, nonbiased, high throughput analysis of complex metabolite mixtures allowing ideally the identification and quantification of every individual metabolite. Metabolomics has emerged as a functional genomics methodology that contributes to our understanding of the complex molecular interactions in biological systems [1]. As such, metabolomics represents the logical progression from large-scale analysis of RNA and proteins at the systems level [2, 3] but unlike transcripts and proteins, the highly diverse molecular identity of metabolites relies on sophisticated instrumentation such as mass spectrometry (MS) and nuclear magnetic resonance spectroscopy (NMR) [4] to be determined.
Metabolites are the end products of cellular regulatory processes, and their levels can be regarded as the ultimate response of biological systems to genetic or environmental changes [5]. Metabolites are also highly dynamic in time and space and show an immense range of structures raising the challenges for analytical procedures in their measurement [6].
Metabolomics allows for a global assessment of a cellular state within the context of the immediate environment, taking into account gene expression, genetic regulation, altered kinetic activity and regulation of enzymes, and changes in metabolic reactions [7–9]. Thus, metabolomic studies compared with genomics or proteomics reflect changes in phenotype and function of a particular tissue, or organism. Thus, the omic sciences are used in a complementary manner to carry out studies in order to characterize a phenotype. The change in the expression of genes and proteins will surely produce changes in the metabolic profile of a cell, tissue or organism [7, 10]. Moreover, metabolomic determinations offer the same advantages as proteomic and transcriptomic techniques: they all have the ability to assay biofluids and are relatively inexpensive, rapid, and automated techniques once start-up costs are taken into account.
Metabolomics was first applied to the study of toxicology and pharmacology, inborn metabolic errors, and nutrition [10]. There is currently great interest in the application of metabolomics to characterize different pathological states of human diseases like cancer, diabetes, autoimmune, and coronary diseases. Moreover, metabolomics can provide valuable tools in a wide range of applications, including microbial biotechnology, food technology, pharmacology, toxicology, enzyme discovery, systems biology, and plant biotechnology (Figure 1). Current research in plants makes extensive use of metabolomics due to the benefit produced on human health of various products of plant origin, including food, pharmaceuticals, and industrial raw materials, as well as its use in plant breeding and nutrition assessment. In addition, the vast chemical diversity of plants compared with animals and microorganisms also contributes to the prominence of metabolomics in plant research [11].
Figure 1.
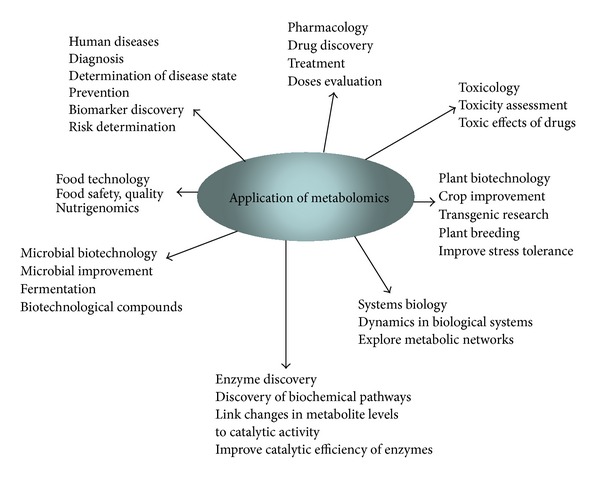
Applications of metabolomics.
Recent technological progress in NMR spectroscopy and MS, the two most accepted methods used in the measurement of metabolites, has improved the sensitivity and spectral resolution of analytic assays on metabolomic samples in attempts to achieve a comprehensive biochemical measurement. Although not all strategies used in metabolomics are universally accepted it is however possible to summarize the most popular ones [19–22] (see Table 1), including metabolite target analysis, metabolite profiling, metabolomics, metabolite flux analysis, and metabolic fingerprinting (and footprinting).
Table 1.
A comparison between the most used methods applied to metabolomic research.
| Technique | Use | Overview | References |
|---|---|---|---|
| HPLC-MS | a, b, c | High sensitivity, nonvolatile metabolites, expensive | [12, 13] |
| GC-MS | a, c | High sensitivity, only volatile compounds, nonexpensive | [14, 15] |
| NMR | b, d, e | For highly abundant metabolites, nondestructive, very reproducible, low sensitivity | [16] |
| MS | a, b, c, d, e | High sensitivity, usually coupled with other techniques | [17] |
| Raman spectroscopy | e | Low resolution, sensitivity, automated, high throughput | [18] |
Ref: a: metabolite target analysis; b: metabolite profiling; c: metabolomics; d: metabolite flux analysis; e: metabolic fingerprinting. MS: mass spectrometry; HPLC-MS: high-performance liquid chromatography-MS; GS-MS: gas chromatography-MS; NMR: nuclear magnetic resonance spectroscopy.
2. Metabolomics in Human Diseases
One of the most important applications of metabolomics in the study of human diseases is in the field of oncobiology. Because tumor cells are highly proliferative and have a high transcription and translation rates, as well as a higher energy demand, they have special metabolic requirements when compared to normal cells and frequently lose many regulatory functions [35]. Thus, one of the greatest challenges in medicine is the use of metabolomics in predicting the appearance of tumor cells. Initially, putative metabolic biomarkers for cancer detection and/or assessment of efficacy of anticancer treatment are discovered in preclinical analyses, followed by the validation of these biomarkers in biofluids (blood, urine, prostatic secretions, etc.) [36]. At present, different metabolites have been identified and proposed that would serve as markers for several tumor processes and other diseases (see Table 2). However, in most cases the combination of metabolomics with other genomic and/or proteomic techniques is extremely useful for both prevention and diagnosis of cancer.
Table 2.
Metabolites used as biomarkers of human diseases.
| Disease | Metabolite biomarker | References |
|---|---|---|
| Male infertility | Citrate, lactate, and glycerylphosphorylcholine | [23, 24] |
| Lung cancer | Metabolites that are shown to have a statistically significant difference between healthy individuals and lung cancer patients were hippurate, trigonelline, β-hydroxyisovalerate, α-hydroxyisobutyrate, N-acetylglutamine, and creatinine | [25] |
| Alzheimer's disease | Succinic anhydride, pyruvic acid, 2-aminopropanol, n,n-didemethylchlorpromazine, L-alanine n-butyl ester, L-glutamic acid dibutyl ester, L-dopa, taurine, creatine, creatinine, lactate, β-alanine, cysteine, fumaric acid, 2-octenedioic acid, and acetoacetic acid | [26, 27] |
| Respiratory diseases | Asthmatic children: acetate; chronic obstructive pulmonary disease (COPD): leucine, lactate, propionate, acetate, and pyruvate | [28, 29] |
| Huntington disease | 3-Nitropropionic acid | [30] |
| Multiple sclerosis | Elevated levels: 2-aminobutyrate, 1,3-dimethylurrate, glutamate, and acetate. Reduced levels: oxaloacetate, citrate, alanine, and 3-hydroxybutyrate. | [31] |
| Impaired glucose tolerance (IGT) | Significantly altered levels: glycine, lysophosphatidylcholine (LPC) (18:2), and acetylcarnitine | [32] |
| Renal cell carcinoma | Phospholipids, phenylalanine, tryptophan, acylcarnitines, cholesterol metabolites, and arachidonic acid metabolism | [33] |
| Colorectal cancer | Acteylcarnine, phenylacetylglutamin, leucylproline, and aspartyllysine | [34] |
| Kidney cancer | Quinolinate, 4-hydroxybenzoate, and gentisate | [34] |
One of the most frequent tumors with an increasing incidence in humans is the hepatocellular carcinoma (HCC). Liu et al. [37] reported the identification of cytokine biomarkers using antibody microarrays. They identified several protein markers such as a macrophage-derived chemokine and macrophage-stimulating protein differentially expressed in patients with liver carcinoma. In addition, different studies were performed in order to obtain a metabolite profile in patients with HCC. Using ultraperformance liquid chromatography-electrospray ionization and TOF mass spectrometry, Patterson et al. [38] have described an alteration of lipid metabolism in these patients, with an increase of glycodeoxycholate, deoxycholate-3-sulfate, and bilirubin. Moreover, the reduction in the levels of lysophosphocholine correlated with the appearance of HCC. Other metabolites found to be altered in liver cancer were polar compounds especially those from the amino acid group such as arginine, proline, alanine, lysine, and aspartate [39]. In addition, an increased level of some amino acids (including alanine) was found in other tumors such as brain tumors, gliomas, and neuroepithelial tumors [40]. Recently, it was described that compounds such as choline, phosphocholine, myoinositol, glycine, taurine and glycerophosphocholine are altered in some types of breast cancers. In this type of diseases, usually other molecules such as the estrogen receptor and the Human Epidermal Growth Factor Receptor 2 (Her2/neu) are widely used as biomarkers for prognostic or predictive purposes [10]. On the other hand, myo-inositol levels were also found to be increased in prostate cancer, colon adenocarcinoma, and ovarian carcinoma [41, 42].
The use of stable isotopes in cancer research has also been reported. Stable isotopes are nontoxic compounds such as 13C-labeled metabolites which can be used to investigate metabolic pathways in normal or highly proliferative cells [43]. Such isotopes can be distinguish easily by NMR or MS and provide information about changes in the regulation of biochemical pathways between cancer and noncancer cells, leading to the development of new diagnostic tools.
However, there are several gaps in the knowledge of the cancer metabolome. The metabolite profile could vary among the different tumor types making it difficult to generalize findings across tumor groups. There are also technical difficulties encountered while performing metabolomic assays that may hinder the characterization of a tumor metabolome, including sample-to-sample variation, the sensitivity and the physiological status of the tumor [10].
Another application of metabolomics in the field of human health includes nutrigenomics. The term nutrigenomics is associated with the interaction of the diet and the genes and reflects the change in gene expression that takes place after the exposure to different nutrients. The identification of certain compounds that would have the ability to act on the expression of target genes is extremely important in preventing diseases such as cancer. Moreover, this interaction could influence the absorption, digestion, and the elimination of metabolites [44]. Thus, the metabolomic approach may allow the discovery of bioprotective foods [20].
Finally, metabolomics became indispensable for the identification and prevention of coronary heart disease. The recognition of a myocardial ischemic episode is important for both the diagnosis and therapy of the disease [45]. At present, several protein and enzyme biomarkers are used for the identification of patients with risk of coronary diseases such as reactive C-protein, tumor necrosis factor α (TNF-α), receptors types 1 and 2 (sTNF-R1 and sTNF-R2), and interleukin-6 [46]. However, numerous efforts are underway to identify metabolites that may also serve as markers of disease using metabolomic techniques. Sabatine et al. (2005) reported an alteration in the levels of several metabolites that could serve as biomarkers of myocardial ischemia. Plasma levels of lactic acid and also metabolites involved in skeletal muscle AMP catabolism such as hypoxanthine and inosine as well as alanine were found to be increased after myocardial injury [47]. On the other hand, several metabolites such as Krebs cycle intermediates, uric acid, and GABA yielded significant but discordant changes in controls and cases. Mayr et al. (2008) used a combined metabolomic and proteomic approach to study patients presenting persistent atrial fibrillation. Those results showed a rise in beta-hydroxybutirate levels with an increase in different ketogenic amino acids and glycine [48]. The metabolomic data correlates with the increase in 3-oxoacid transferase, a key enzyme involved in ketolytic energy production. Recently, Shah et al. (2010) characterized the metabolic profile of patients with cardiovascular disease using 69 metabolites [49]. Different acylcarnitines as well as several amino acid levels including leucine, isoleucine, glutamate, glutamine, proline, and methionine were found to be altered in patients when compared to control groups [49]. In addition, it has been reported that the level of the metabolite dicarboxylacylcarnitine could be predictive of cardiovascular disease. More recently, using plasma metabolic fingerprint, it has been possible to classify patients with aortic abdominal aneurysm (AAA) and to predict the disease stage. In this assay, sphingolipids, lysophospholipids, cholesterol, acylcarnitines, and guanidinosuccinic acid were proposed as markers of AAA [50].
Although many efforts have been made in order to find new metabolic markers for the prevention of cardiac diseases, the metabolomic approach is still under development. To better understand metabolic pathways alterations, including changes in metabolite levels and gene transcription taking place in patients with risk of cardiac disease, it is necessary to use integrated approaches, combining biomarkers from genomics, proteomics, and metabolomics. This will result in a better picture and therefore a better selection of the potential biomarkers for prevention, diagnosis, and risk prediction of heart disease [51].
3. Metabolomics and Its Applications in Plant Biotechnology
Metabolomics originally developed from metabolic profiling. In the early 1970's GC-MS technologies were used to analyze steroids, acids, and neutral and acidic urinary drug metabolites [52–54]. Soon afterwards, the concept of using metabolic profiles to screen, diagnose, and assess health began to spread [55, 56]. However, it was not until the early 1990's that metabolite profiling was used to study plant organisms [14, 57, 58].
Plant metabolites are involved in many resistance and stress responses and also contribute to the color, taste, aroma, and scent of fruits and flowers [2]. As we mentioned previously, the biochemical phenotype of an organism is the final result of interactions between the genotype and the environmental stimuli; but it is also modulated by intracellular physiological fluctuations that are part of homeostasis [3]. Thus, the simultaneous identification and quantification of metabolites is necessary to understand the dynamics of the metabolome, analyze fluxes in metabolic pathways and decipher the role of each metabolite following various stimuli [59]. The challenge of metabolomics is to find changes in biochemical pathways, and metabolic networks that might correlate with the physiological and developmental phenotype of a cell, tissue, or organism [2, 3].
One of the greatest achievements of plant biology is the completion of the whole genome sequences of model plants such as Arabidopsis thaliana and rice. In Arabidopsis ~27000 genes were predicted based on nucleotide sequence information; however, only half of these genes have been functionally annotated based on sequence similarity to known genes, and among these the function of only ~11% has been confirmed with direct experimental evidence [60]. The elucidation of unknown genes function is therefore currently a major challenge in plant research. Because there is very little information on the number of genes in a particular gene family of a nonmodel plant, it becomes necessary to know the profile of expression of these genes under different conditions and stimuli. The integration of metabolomics with transcriptional profiles can provide clues for the identification of the functions of the unknown genes, regardless of whether they are from model or nonmodel plants [61].
Plants produce more than 200,000 metabolites, many of which play specific roles in allowing adaptation to specific ecological niches [5, 62]. Therefore, the main problems encountered when characterizing the plant metabolome have to do with the fact that in comparison to the proteome or transcriptome, the metabolome is highly complex in nature, due to the enormous chemical diversity of the compounds. In addition, there is a wide range of metabolite concentrations, which can vary over nine orders of magnitude (pM to mM). These large variations in the nature and the concentration of analytes to be studied provide challenges to all the analytical technologies employed in metabolomic strategies [63].
Using metabolomics, it is possible to identify pathways responsible for the production of important food metabolites that could be important in improving human health. There are several examples where the modification of certain metabolic pathways led to the production of plants with an increased nutritional value. This is the case of Golden Rice (GR), which is genetically modified rice that accumulates β-carotene in the endosperm [64]. The production of this variety of rice allowed alleviating vitamin A deficiency, a major nutritional problem worldwide. The nutritional value of GR was later improved by the overexpression of a phytoene synthase gene leading to the obtention of the GR2 variety, which accumulates higher amounts of carotenoids (84% of the total is β-carotene) [65]. Mehta et al. (2002) were able to express an S-adenosylmethionine decarboxylase gene under the inducible E8 promoter in tomato. The transgenic variety shows higher levels of different polyamines during fruit ripening, including spermidine and spermine, leading to an increase in the metabolite lycopene, which prolonged vine life and enhanced fruit juice and nutrient quality [66].
Other examples include the engineering of plants to enhance anthocyanin content. Anthocyanins are flavonoids, a class of pigments that contribute to the colors and antioxidant qualities of plants. Moreover, these metabolites have been associated with the protection against several human diseases, but their natural levels in plants are inadequate to confer optimal benefits. Recently, it has been reported that the expression of two transcription factors in tomato led to the accumulation of higher quantities of anthocyanins at concentrations comparable to those founded in high antocyanin-containing plants such as blackberries and blueberries [67]. The new variety has an intense purple coloration and also a 3-fold enhanced antioxidant capacity. In addition, this study also reported an extension of the life-span of cancer susceptible mice which were fed with a diet supplemented with this tomato variety.
Plant metabolomics is being increasingly used for understanding other processes such as the cellular responses to stress conditions. The metabolic readjustment to sulfur deficiency is an example of this [74]. A close relationship was found between sulfur assimilation, nitrogen, lipid, and purine metabolism and enhanced photorespiration. Metabolomics has also been applied to the study of the cold stress response [75]. Other applications include metabolic engineering of biochemical pathways, gene function discovery, and engineering pathways for pharmaceuticals production (Table 3) [76].
Table 3.
Recent applications of metabolomics in plant biotechnology.
| Organism | Application | Technology used | Reference |
|---|---|---|---|
| Catharanthus roseus | Improvement of the production of anticancer indole alkaloid by overexpression of ORCA3 and G10H in C. roseus plants | NMR | [68] |
| Panicum virgatum (switchgrass) | Increased amounts of phenolic acids and a monolignol analog associated with more facile cell wall deconstruction | GC-MS | [69] |
| Solanum tuberosum (L) | Increased drought tolerance by expression of trehalose-6-phosphate synthase 1 | GC-MS | [70] |
| Oryza sativa | Modulation of salt tolerance by reduction of OsSUT1 (O. sativa sucrose transporter 1) expression | GC-TOF-MS | [71] |
| Arabidopsis thaliana | Distinguish transgenic and nontransgenic plants | NMR | [72] |
| Solanum Lycopersicum | Higher accumulation of flavonoids and thus nutritional value in tomato plants carrying a mutation in HP1/LeDDB1 gene | LC-ESI-MS/MS | [73] |
4. Methods Used for the Determination of Plant Metabolomes
The basic instrumentation useful for the evaluation of plant metabolome includes direct spectroscopy, chromatography (fingerprinting), or high-performance chromatography (GC or HPLC) combined with spectroscopy (IR, NIR, MS). The coupling of chromatographic methods with MS can substantially increase the depth of metabolome coverage, add an additional dimension for metabolite identification and enhance the biological context through the more rigorous identification of a greater number of metabolites [77].
GC-MS is one of the most widely used analytical techniques in plant metabolomics. It is utilized to analyze a qualitatively and quantitatively wide range of volatile and/or derivatized nonvolatile metabolites with high thermal stability. After separation, the eluted metabolites are identified by mass spectrophotometers. This technique has high analytical reproducibility and lower costs compared to other hyphenated techniques, such as LC-MS or LC-NMR [15, 77]. Alternatively, direct injection MS analysis may also be applied for the phenotyping of plants, that is, Fourier transformed-MS (FT-MS) provides ultimate limit of detection and mass measurement precision to enable metabolomic analyses [77, 78].
In the last years, the use of LC-MS in plant metabolomics has been increasing. LC-MS is more suitable than GC-MS for labile compounds as well as for those that are difficult to derivatize. The application of LC-MS in plant metabolomics includes the description of the tomato metabolome database (MoTo DB) [79], the identification of the accumulation of oxylipins after wound-induced stress in Arabidopsis [80], the identification of flavonoid and isoflavonoid compounds in Medicago truncatula [81], the untargeted large-scale analysis of plant metabolome [82], the characterization of metabolites that respond to stress [83, 84], and the identification of flavonoid glycoconjugates in roots form Medicago truncaluta [85].
Another technique used for the quantitative analysis of plant metabolites is capillary electrophoresis coupled to MS (CE-MS) [86]. CE-MS was used to characterize the amino acid profile in plant cell cultures [87], to characterize the metabolome of bacterial-infected orange leaves [88], and to identify the metabolite profile in Illicium anisatum [89]. CE coupled with MS has also the advantages of high resolution and high reliability [86].
Fourier transformed infrared spectroscopy (FT-IR), near infrared resonance (NIR) Raman spectroscopy, and, more recently, nuclear magnetic resonance (NMR) spectroscopy are constantly developing rapid, nondestructive and high-throughput techniques for a diverse range of sample types. FT-IR has also been introduced as a metabolic fingerprinting technique within the plant sciences [90, 91]. Raman spectroscopy coupled with microscopy has been recently used with great success, making possible the identification and quantification of phytochemicals and their distribution directly from plant tissues [92–94].
NMR techniques can uniquely identify and simultaneously quantify a wide range of organic compounds in the micromolar range [95, 96]. NMR-based methods can be broadly classified into solution NMR and insoluble or solid-state NMR, according to sample solubility. In onedimensional NMR, protons (1H) are usually observed (1H-NMR) due to the sensitivity and common occurrence of this magnetic nucleus. More detailed analyses, such as metabolite identification or flux assay, can be obtained with other nuclei, particularly 13C and 15N that are coupled with 1H nuclei in twodimensional or multidimensional NMR analysis [97, 98]. Recently, NMR has been used to identify secondary metabolites involved in host-plant resistance [99] and to characterize the metabolites produced after salt stress exposure in maize plants [100]. Other applications of NMR are in the investigation of food quality and in the standardization and control of different phytomedical preparations [101].
The resulting MS or NMR spectra are preprocessed to suppress noise and align the different peaks. Thereafter, the data are used to identify the different metabolites present in the sample using metabolite databases. In nontarget analyses, the spectra of unknown compounds can be subjected to statistical analyses [102]. In target analyses, the sets of spectral data that are associated with particular compounds are used as metabolic profiles for each compound in further analyses [102].
All the metabolomic technologies mentioned require data processing analysis due to, for example, some inaccuracies such as chromatogram shift and mass drift. Some of the most used methods to group metabolites in samples include multivariate statistical analyses such as principal component analysis (PCA), hierarchical clustering analysis (HCA), and self-organization mapping (SOM) [102]. In addition, the analysis and comprehension of the data obtained by metabolomics is often integrated with data from genomic and proteomic experiments [102].
5. Metabolomic Resources
Several metabolomic resources and metabolite databases are available (Table 4). The Human Metabolome Database is an electronic database which contains information on metabolites found in humans (http://www.hmdb.ca/). This database includes chemical, clinical, and biochemical data, linking known metabolites to several genes and proteins. The Golm Metabolome Database (http://gmd.mpimp-golm.mpg.de/) contains information about mass spectra from active metabolites quantified by GS-MS. The Madison Metabolomics Database is a resource for metabolomics research based on NMR and MS (http://mmcd.nmrfam.wisc.edu/). Another integrative database is Metabolights (http://www.ebi.ac.uk/metabolights/). This database is cross-species and contains information about metabolite structures, spectra and biological roles. Metabolomics at Rothamsted (MeT-RO) is an initiative that contains several resources that can be applied to plant and microbial metabolomics (http://www.metabolomics.bbsrc.ac.uk/MeT-RO.htm). The Metlin Metabolite Database contains information on about 55,000 metabolites and nearly 50,000 high resolution MS/MS spectra and tandem MS experiments (http://metlin.scripps.edu/). PRIMe, a platform for RIKEN metabolomics is a database that integrates genomic and metabolomic data. It contains information on metabolites obtained from NMR spectroscopy, GC-MS, LC-MS, and CE-MS (http://prime.psc.riken.jp/).
Table 4.
Metabolomic resources.
| Name | URL | Information/Species |
|---|---|---|
| Human metabolome database | http://www.hmdb.ca/ | Chemical and biological data of human metabolites |
| Golm metabolome database | http://gmd.mpimp-golm.mpg.de/ | GS-MS |
| Madison metabolomics database | http://mmcd.nmrfam.wisc.edu/ | NMR and MS |
| Metabolights | http://www.ebi.ac.uk/metabolights/ | Metabolite structures, spectra, function/cross-species |
| Metabolomics at Rothamsted (MeT-RO) | http://www.metabolomics.bbsrc.ac.uk/MeT-RO.htm | Plant and microbial metabolites |
| Metlin metabolite database | http://metlin.scripps.edu/ | High resolution MS/MS spectra and tandem MS experiments |
| PRIMe | http://prime.psc.riken.jp/ | Genomic and metabolomics data/NMR spectroscopy, GC-MS, LC-MS and CE-MS |
| Plantmetabolomics | http://plantmetabolomics.vrac.iastate.edu/ver2/ | Arabidopsis and other plant species |
| Metabolome tomato database | [79] | Metabolites identified by LC-MS |
| Terpmed | http://www.terpmed.eu/databases.html | Plant terpenoids, natural products, secondary metabolites/Therapeutic drugs |
| Armec repository project | http://www.armec.org/MetaboliteLibrary/ | FIE-MS/HPLC-ESI-MS data/human and plant metabolomes for nutrition |
| MetaCyc | http://metacyc.org/ | Integration of metabolite data with metabolic pathways/2000 organisms |
| RiceCyc | http://pathway.gramene.org/gramene/ricecyc.shtml | Metabolic pathways, enzymes, metabolites |
| AraCyc | http://www.arabidopsis.org/biocyc/ | Metabolic pathways, compounds/Arabidopsis |
| Solanacea Genomics network (SolCyc) | http://solgenomics.net/tools/solcyc/index.pl | Pathway genome databases/solanacea species |
| HumanCyc | http://humancyc.org/ | Metabolic pathways, genome/human |
| KEGG Pathway database | http://www.genome.jp/kegg/pathway.html | Pathways, metabolism, genetic information Cellular processes, human diseases |
| Mapman | http://mapman.gabipd.org/web/guest/mapman | Datasets (e.g., Gene expression data, metabolic pathways) |
Several databases for plant species have also been developed, such as Plantmetabolomics (http://plantmetabolomics.vrac.iastate.edu/ver2/). This initiative started as a metabolomic and functional genomic tool for elucidating the functions of Arabidopsis genes. In addition, other databases such as the Metabolome Tomato Database are available and contain information on metabolites identified by LC-MS [79]. A related database is Terpmed (http://www.terpmed.eu/databases.html) which contains information about plant terpenoids, natural products, and other secondary metabolites important for use as therapeutic drugs. The Armec Repository Project was created as a tool to annotate flow injection electrospray MS (FIE-MS) data, but information about HPLC-ESI-MS can also be found in this database. Moreover, at present the Armec database is in expansion, including information about additional species focused on food crops and human metabolome for use in nutrition research (http://www.armec.org/MetaboliteLibrary/).
On the other hand, there are several resources that integrate metabolite data with metabolic pathways such as MetaCyc (http://metacyc.org/) that contains information on about 1800 pathways from more than 2000 organisms. Other related databases are RiceCyc (http://pathway.gramene.org/gramene/ricecyc.shtml), AraCyc (http://www.arabidopsis.org/biocyc/), Solanacea Genomics Network (SolCyc, http://solgenomics.net/tools/solcyc/index.pl), HumanCyc (http://humancyc.org/), KEGG Pathway database (http://www.genome.jp/kegg/pathway.html), and Mapman (http://mapman.gabipd.org/web/guest/mapman).
6. Conclusions
Metabolomic studies require knowledge in many areas such as biochemistry, biology, physiology, and bioinformatics. Metabolomics has the potential to make a large impact on different areas of biology, including human health, plant biotechnology, toxicology, and pharmacology, among others. The analysis and comprehension of the data obtained require the use of bioinformatics, as well as the integration with data obtained from genomic and proteomic analyses.
With the development of more sensitive technologies and also computational tools for statistical analysis and data interpretation, metabolomics has the potential to help us better understand the molecular mechanisms of disease. In addition, the identification and characterization of new biomarkers will allow the diagnosis and prevention of many diseases, as well as the discovery of new drugs. However, in most diseases the analysis of other biomarkers, like protein markers as well as the evaluation of the physiological state for diagnosis and treatment is also required.
In the area of plant biotechnology, metabolomics, together with genomic and proteomic studies, has identified new genes or genes with new functions. In this area, metabolomics has gained importance in the evaluation of transgenic plants, food quality, the increase disease resistance, and herbicide or salinity tolerance. Moreover, the integration of the three omics and systems biology is an excellent strategy for the discovery enzymes involved in unknown metabolic pathways. In addition, metabolomics applied to the study of plants becomes extremely important in the efforts that are performed in order to improve human health. As there are several examples where the manipulation of different metabolic pathways led to an increase in the production of a particular metabolite, the modification of plant metabolic pathways could lead to the production of new drugs that could be used for the treatment of many diseases.
Acknowledgments
This work was supported by Grants from ANPCyT (PICT 0729, PICT 0543, and 0512) and CONICET (2009-236). Maria V. Busi, Maria I. Zanor and Diego F. Gomez-Casati are research members from CONICET.
References
- 1.Hall R, Beale M, Fiehn O, Hardy N, Sumner L, Bino R. Plant metabolomics: the missing link in functional genomics strategies. Plant Cell. 2002;14(7):1437–1440. doi: 10.1105/tpc.140720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bino RJ, Hall RD, Fiehn O, et al. Potential of metabolomics as a functional genomics tool. Trends in Plant Science. 2004;9(9):418–425. doi: 10.1016/j.tplants.2004.07.004. [DOI] [PubMed] [Google Scholar]
- 3.Weckwerth W. Metabolomics in systems biology. Annual Review of Plant Biology. 2003;54:669–689. doi: 10.1146/annurev.arplant.54.031902.135014. [DOI] [PubMed] [Google Scholar]
- 4.Lei Z, Huhman DV, Sumner LW. Mass spectrometry strategies in metabolomics. Journal of Biological Chemistry. 2011;286(29):25435–25442. doi: 10.1074/jbc.R111.238691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Fiehn O. Metabolomics—the link between genotypes and phenotypes. Plant Molecular Biology. 2002;48(1-2):155–171. [PubMed] [Google Scholar]
- 6.Stitt M, Fernie AR. From measurements of metabolites to metabolomics: an “on the fly” perspective illustrated by recent studies of carbon-nitrogen interactions. Current Opinion in Biotechnology. 2003;14(2):136–144. doi: 10.1016/s0958-1669(03)00023-5. [DOI] [PubMed] [Google Scholar]
- 7.Griffin JL, Shockcor JP. Metabolic profiles of cancer cells. Nature Reviews Cancer. 2004;4(7):551–561. doi: 10.1038/nrc1390. [DOI] [PubMed] [Google Scholar]
- 8.Mendes P, Kell DB, Westerhoff HV. Channelling can decrease pool size. European Journal of Biochemistry. 1992;204(1):257–266. doi: 10.1111/j.1432-1033.1992.tb16632.x. [DOI] [PubMed] [Google Scholar]
- 9.Menedes P, Kell DB, Westerhoff HV. Why and when channelling can decrease pool size at constant net flux in a simple dynamic channel. Biochimica et Biophysica Acta. 1996;1289(2):175–186. doi: 10.1016/0304-4165(95)00152-2. [DOI] [PubMed] [Google Scholar]
- 10.Spratlin JL, Serkova NJ, Eckhardt SG. Clinical applications of metabolomics in oncology: a review. Clinical Cancer Research. 2009;15(2):431–440. doi: 10.1158/1078-0432.CCR-08-1059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Akiyama K, Chikayama E, Yuasa H, et al. PRIMe: a web site that assembles tools for metabolomics and transcriptomics. In Silico Biology. 2008;8(3-4):339–345. [PubMed] [Google Scholar]
- 12.Staack RF, Theobald DS, Maurer HH. Studies on the human metabolism and the toxicologic detection of the cough suppressant dropropizine in urine using gas chromatography-mass spectrometry. Therapeutic Drug Monitoring. 2004;26(4):441–449. doi: 10.1097/00007691-200408000-00015. [DOI] [PubMed] [Google Scholar]
- 13.Olah TV, McLoughlin DA, Gilbert JD. The simultaneous determination of mixtures of drug candidates by liquid chromatography/atmospheric pressure chemical ionization mass spectrometry as an in vivo drug screening procedure. Rapid Communications in Mass Spectrometry. 1997;11:17–12. doi: 10.1002/(SICI)1097-0231(19970115)11:1<17::AID-RCM812>3.0.CO;2-N. [DOI] [PubMed] [Google Scholar]
- 14.Roessner U, Luedemann A, Brust D, et al. Metabolic profiling allows comprehensive phenotyping of genetically or environmentally modified plant systems. Plant Cell. 2001;13(1):11–29. doi: 10.1105/tpc.13.1.11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Fiehn O, Kopka J, Dörmann P, Altmann T, Trethewey RN, Willmitzer L. Metabolite profiling for plant functional genomics. Nature Biotechnology. 2000;18:1157–1161. doi: 10.1038/81137. [DOI] [PubMed] [Google Scholar]
- 16.Lenz EM, Bright J, Wilson ID, Morgan SR, Nash AFP. A 1H NMR-based metabonomic study of urine and plasma samples obtained from healthy human subjects. Journal of Pharmaceutical and Biomedical Analysis. 2003;33(5):1103–1115. doi: 10.1016/s0731-7085(03)00410-2. [DOI] [PubMed] [Google Scholar]
- 17.Donato P, Cacciola F, Tranchida PQ, Dugo P, Mondello L. Mass spectrometry detection in comprehensive liquid chromatography: basic concepts, instrumental aspects, applications and trends. Mass Spectrometry Reviews. 2012;31(5):523–559. doi: 10.1002/mas.20353. [DOI] [PubMed] [Google Scholar]
- 18.Sikirzhytski V, Sikirzhytskaya A, Lednev IK. Multidimensional Raman spectroscopic signatures as a tool for forensic identification of body fluid traces: a review. Applied Spectroscopy. 2011;65(11):1223–1232. doi: 10.1366/11-06455. [DOI] [PubMed] [Google Scholar]
- 19.Fiehn O. Combining genomics, metabolome analysis, and biochemical modelling to understand metabolic networks. Comparative and Functional Genomics. 2001;2(3):155–168. doi: 10.1002/cfg.82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Goodacre R. Metabolomics of a superorganism. Journal of Nutrition. 2007;137(1):259–266. doi: 10.1093/jn/137.1.259S. [DOI] [PubMed] [Google Scholar]
- 21.Goodacre R, Vaidyanathan S, Dunn WB, Harrigan GG, Kell DB. Metabolomics by numbers: acquiring and understanding global metabolite data. Trends in Biotechnology. 2004;22(5):245–252. doi: 10.1016/j.tibtech.2004.03.007. [DOI] [PubMed] [Google Scholar]
- 22.Harrigan GG, Goodacre R. Metabolic Profiling: Its Role in Biomarker Discovery and Gene Function Analysis. Kluwer Academic Publishers; 2003. [Google Scholar]
- 23.Kovac JR, Pastuszak AW, Lamb DJ. The use of genomics, proteomics, and metabolomics in identifying biomarkers of male infertility. Fertility and Sterility. 2013;99:998–1007. doi: 10.1016/j.fertnstert.2013.01.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hamamah S, Seguin F, Barthelemy C, et al. 1H nuclear magnetic resonance studies of seminal plasma from fertile and infertile men. Journal of Reproduction and Fertility. 1993;97(1):51–55. doi: 10.1530/jrf.0.0970051. [DOI] [PubMed] [Google Scholar]
- 25.Carrola J, Rocha CM, Barros AS, et al. Metabolic signatures of lung cancer in biofluids: NMR-based metabonomics of urine. Journal of Proteome Research. 2011;10(1):221–230. doi: 10.1021/pr100899x. [DOI] [PubMed] [Google Scholar]
- 26.Xu XH, Huang Y, Wang G, Chen SD. Metabolomics: a novel approach to identify potential diagnostic biomarkers and pathogenesis in Alzheimer's disease. Neuroscience Bulletin. 2012;28:641–648. doi: 10.1007/s12264-012-1272-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Trushina E, Dutta T, Persson XM, Mielke MM, Petersen RC. Identification of altered metabolic pathways in plasma and CSF in mild cognitive impairment and Alzheimer's disease using metabolomics. PLoS ONE. 2013;8 doi: 10.1371/journal.pone.0063644.e63644 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Carraro S, Rezzi S, Reniero F, et al. Metabolomics applied to exhaled breath condensate in childhood asthma. American Journal of Respiratory and Critical Care Medicine. 2007;175(10):986–990. doi: 10.1164/rccm.200606-769OC. [DOI] [PubMed] [Google Scholar]
- 29.De Laurentiis G, Paris D, Melck D, et al. Metabonomic analysis of exhaled breath condensate in adults by nuclear magnetic resonance spectroscopy. European Respiratory Journal. 2008;32(5):1175–1183. doi: 10.1183/09031936.00072408. [DOI] [PubMed] [Google Scholar]
- 30.Henry P-G, Mochel F. The search for sensitive biomarkers in presymptomatic Huntington disease. Journal of Cerebral Blood Flow and Metabolism. 2012;32(5):769–770. doi: 10.1038/jcbfm.2012.17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Smolinska A, Blanchet L, Buydens LM, Wijmenga SS. NMR and pattern recognition methods in metabolomics: from data acquisition to biomarker discovery: a review. Analytica Chimica Acta. 2012;750:82–97. doi: 10.1016/j.aca.2012.05.049. [DOI] [PubMed] [Google Scholar]
- 32.Wang-Sattler R, Yu Z, Herder C, et al. Novel biomarkers for pre-diabetes identified by metabolomics. Molecular Systems Biology. 2012;8:p. 615. doi: 10.1038/msb.2012.43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Liu G, Snapp HM, Ji QC, Arnold ME. Strategy of accelerated method development for high-throughput bioanalytical assays using ultra high-performance liquid chromatography coupled with mass spectrometry. Analytical Chemistry. 2009;81(22):9225–9232. doi: 10.1021/ac901316w. [DOI] [PubMed] [Google Scholar]
- 34.Kim K, Taylor SL, Ganti S, Guo L, Osier MV, Weiss RH. Urine metabolomic analysis identifies potential biomarkers and pathogenic pathways in kidney cancer. OMICS. 2011;15(5):293–303. doi: 10.1089/omi.2010.0094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Hanahan D, Weinberg RA. The hallmarks of cancer. Cell. 2000;100(1):57–70. doi: 10.1016/s0092-8674(00)81683-9. [DOI] [PubMed] [Google Scholar]
- 36.Serkova NJ, Glunde K. Metabolomics of cancer. Methods in Molecular Biology. 2009;520:273–295. doi: 10.1007/978-1-60327-811-9_20. [DOI] [PubMed] [Google Scholar]
- 37.Liu T, Xue R, Dong L, Wu H, Zhang D, Shen X. Rapid determination of serological cytokine biomarkers for hepatitis B virus-related hepatocellular carcinoma using antibody microarrays. Acta Biochimica et Biophysica Sinica. 2011;43(1):45–51. doi: 10.1093/abbs/gmq111. [DOI] [PubMed] [Google Scholar]
- 38.Patterson AD, Maurhofer O, Beyoğlu D, et al. Aberrant lipid metabolism in hepatocellular carcinoma revealed by plasma metabolomics and lipid profiling. Cancer Research. 2011;71(21):6590–6600. doi: 10.1158/0008-5472.CAN-11-0885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Chen J, Wang W, Lv S, et al. Metabonomics study of liver cancer based on ultra performance liquid chromatography coupled to mass spectrometry with HILIC and RPLC separations. Analytica Chimica Acta. 2009;650(1):3–9. doi: 10.1016/j.aca.2009.03.039. [DOI] [PubMed] [Google Scholar]
- 40.Masuo Y, Imai T, Shibato J, et al. Omic analyses unravels global molecular changes in the brain and liver of a rat model for chronic Sake (Japanese alcoholic beverage) intake. Electrophoresis. 2009;30(8):1259–1275. doi: 10.1002/elps.200900045. [DOI] [PubMed] [Google Scholar]
- 41.Lee DG, Lee J-H, Choi BK, et al. H+-myo-inositol transporter SLC2A13 as a potential marker for cancer stem cells in an oral squamous cell carcinoma. Current Cancer Drug Targets. 2011;11(8):966–975. doi: 10.2174/156800911797264752. [DOI] [PubMed] [Google Scholar]
- 42.Serkova NJ, Gamito EJ, Jones RH, et al. The metabolites citrate, myo-inositol, and spermine are potential age-independent markers of prostate cancer in human expressed prostatic secretions. Prostate. 2008;68(6):620–628. doi: 10.1002/pros.20727. [DOI] [PubMed] [Google Scholar]
- 43.Lane AN, Fan TW-M, Higashi RM. Stable isotope-assisted metabolomics in cancer research. IUBMB Life. 2008;60(2):124–129. doi: 10.1002/iub.17. [DOI] [PubMed] [Google Scholar]
- 44.Riscuta G, Dumitrescu RG. Nutrigenomics: implications for breast and colon cancer prevention. Methods in Molecular Biology. 2012;863:343–358. doi: 10.1007/978-1-61779-612-8_22. [DOI] [PubMed] [Google Scholar]
- 45.Griffin JL, Atherton H, Shockcor J, Atzori L. Metabolomics as a tool for cardiac research. Nature Reviews Cardiology. 2011;8(11):630–643. doi: 10.1038/nrcardio.2011.138. [DOI] [PubMed] [Google Scholar]
- 46.Pai JK, Pischon T, Ma J, et al. Inflammatory markers and the risk of coronary heart disease in men and women. New England Journal of Medicine. 2004;351:2599–2610. doi: 10.1056/NEJMoa040967. [DOI] [PubMed] [Google Scholar]
- 47.Sabatine MS, Liu E, Morrow DA, et al. Metabolomic identification of novel biomarkers of myocardial ischemia. Circulation. 2005;112(25):3868–3875. doi: 10.1161/CIRCULATIONAHA.105.569137. [DOI] [PubMed] [Google Scholar]
- 48.Mayr M, Yusuf S, Weir G, et al. Combined metabolomic and proteomic analysis of human atrial fibrillation. Journal of the American College of Cardiology. 2008;51:585–594. doi: 10.1016/j.jacc.2007.09.055. [DOI] [PubMed] [Google Scholar]
- 49.Shah SH, Bain JR, Muehlbauer MJ, et al. Association of a peripheral blood metabolic profile with coronary artery disease and risk of subsequent cardiovascular events. Circulation. 2010;3(2):207–214. doi: 10.1161/CIRCGENETICS.109.852814. [DOI] [PubMed] [Google Scholar]
- 50.Ciborowski M, Teul J, Martin-Ventura JL, Egido J, Barbas C. Metabolomics with LC-QTOF-MS permits the prediction of disease stage in aortic abdominal aneurysm based on plasma metabolic fingerprint. PLoS ONE. 2012;7(2) doi: 10.1371/journal.pone.0031982.e31982 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Herder C, Karakas M, Koenig W. Biomarkers for the prediction of type 2 diabetes and cardiovascular disease. Clinical Pharmacology and Therapeutics. 2011;90(1):52–66. doi: 10.1038/clpt.2011.93. [DOI] [PubMed] [Google Scholar]
- 52.Devaux PG, Homing MG, Homing BC. Benyzloxime derivative of steroids, a new metabolic profile procedure for human urinary steroids. Analytical Letters. 1971;4:p. 151. [Google Scholar]
- 53.Horning BC, Horning MG. Metabolic Profiles: Chromatographic Methods for Isolation and Characterization of a Variety of Metabolites in Man. Chicago, Ill, USA: Yearbook Medical Publishers; 1970. [PubMed] [Google Scholar]
- 54.Horning BC, Horning MG. Human metabolic profiles obtained by GC and GC/MS. Journal of Chromatographic Science. 1971;9:129–140. [Google Scholar]
- 55.Cunnick WR, Cromie JB, Cortell R, et al. Value of biochemical profiling in a periodic health examination program: analysis of 1,000 cases. Bulletin of the New York Academy of Medicine. 1972;18(1):5–22. [PMC free article] [PubMed] [Google Scholar]
- 56.Mroczek WJ. Biochemical profiling and the natural history of hypertensive diseases. Circulation. 1972;45(6):1332–1333. doi: 10.1161/01.cir.45.6.1332. [DOI] [PubMed] [Google Scholar]
- 57.Roessner U, Wagner C, Kopka J, Trethewey RN, Willmitzer L. Simultaneous analysis of metabolites in potato tuber by gas chromatography-mass spectrometry. Plant Journal. 2000;23(1):131–142. doi: 10.1046/j.1365-313x.2000.00774.x. [DOI] [PubMed] [Google Scholar]
- 58.Maloney V. Plant metabolomics. Biotechnology Journal. 2004;2:92–99. [Google Scholar]
- 59.Fiehn O, Weckwerth W. Deciphering metabolic networks. European Journal of Biochemistry. 2003;270(4):579–588. doi: 10.1046/j.1432-1033.2003.03427.x. [DOI] [PubMed] [Google Scholar]
- 60.MASC. The Multinational Coordinated Arabidopsis thaliana Functional Genomics Project—Annual Report 2007. 2007 [Google Scholar]
- 61.Saito K, Hirai MY, Yonekura-Sakakibara K. Decoding genes with coexpression networks and metabolomics—“majority report by precogs”. Trends in Plant Science. 2008;13(1):36–43. doi: 10.1016/j.tplants.2007.10.006. [DOI] [PubMed] [Google Scholar]
- 62.Oksman-Caldentey K-M, Inzé D, Orešič M. Connecting genes to metabolites by a systems biology approach. Proceedings of the National Academy of Sciences of the United States of America. 2004;101(27):9949–9950. doi: 10.1073/pnas.0403636101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Dunn WB, Bailey NJC, Johnson HE. Measuring the metabolome: current analytical technologies. Analyst. 2005;130(5):606–625. doi: 10.1039/b418288j. [DOI] [PubMed] [Google Scholar]
- 64.Ye X, Al-Babili S, Kloti A, et al. Engineering the provitamin A (β-carotene) biosynthetic pathway into (carotenoid-free) rice endosperm. Science. 2000;287(5451):303–305. doi: 10.1126/science.287.5451.303. [DOI] [PubMed] [Google Scholar]
- 65.Paine JA, Shipton CA, Chaggar S, et al. Improving the nutritional value of Golden Rice through increased pro-vitamin A content. Nature Biotechnology. 2005;23(4):482–487. doi: 10.1038/nbt1082. [DOI] [PubMed] [Google Scholar]
- 66.Mehta RA, Cassol T, Li N, Ali N, Handa AK, Mattoo AK. Engineered polyamine accumulation in tomato enhances phytonutrient content, juice quality, and vine life. Nature Biotechnology. 2002;20(6):613–618. doi: 10.1038/nbt0602-613. [DOI] [PubMed] [Google Scholar]
- 67.Butelli E, Titta L, Giorgio M, et al. Enrichment of tomato fruit with health-promoting anthocyanins by expression of select transcription factors. Nature Biotechnology. 2008;26(11):1301–1308. doi: 10.1038/nbt.1506. [DOI] [PubMed] [Google Scholar]
- 68.Pan Q, Wang Q, Yuan F, et al. Overexpression of ORCA3 and G10H in Catharanthus roseus plants regulated alkaloid biosynthesis and metabolism revealed by NMR-metabolomics. PLoS ONE. 2012;7 doi: 10.1371/journal.pone.0043038.e43038 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Tschaplinski TJ, Standaert RF, Engle NL, et al. Down-regulation of the caffeic acid O-methyltransferase gene in switchgrass reveals a novel monolignol analog. Biotechnology for Biofuels. 2012;5:p. 71. doi: 10.1186/1754-6834-5-71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Kondrak M, Marincs F, Antal F, Juhasz Z, Banfalvi Z. Effects of yeast trehalose-6-phosphate synthase 1 on gene expression and carbohydrate contents of potato leaves under drought stress conditions. BMC Plant Biology. 2012;12, article 74 doi: 10.1186/1471-2229-12-74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Siahpoosh MR, Sanchez DH, Schlereth A, et al. Modification of OsSUT1 gene expression modulates the salt response of rice Oryza sativa cv. Taipei 309. Plant Science. 2012;182(1):101–111. doi: 10.1016/j.plantsci.2011.01.001. [DOI] [PubMed] [Google Scholar]
- 72.Ren Y, Wang T, Peng Y, Xia B, Qu L-J. Distinguishing transgenic from non-transgenic Arabidopsis plants by 1H NMR-based metabolic fingerprinting. Journal of Genetics and Genomics. 2009;36(10):621–628. doi: 10.1016/S1673-8527(08)60154-X. [DOI] [PubMed] [Google Scholar]
- 73.Calvenzani V, Martinelli M, Lazzeri V, et al. Response of wild-type and high pigment-1 tomato fruit to UV-B depletion: flavonoid profiling and gene expression. Planta. 2010;231(3):755–765. doi: 10.1007/s00425-009-1082-4. [DOI] [PubMed] [Google Scholar]
- 74.Nikiforova VJ, Kopka J, Tolstikov V, et al. Systems rebalancing of metabolism in response to sulfur deprivation, as revealed by metabolome analysis of Arabidopsis plants. Plant Physiology. 2005;138(1):304–318. doi: 10.1104/pp.104.053793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Cook D, Fowler S, Fiehn O, Thomashow MF. A prominent role for the CBF cold response pathway in configuring the low-temperature metabolome of Arabidopsis . Proceedings of the National Academy of Sciences of the United States of America. 2004;101(42):15243–15248. doi: 10.1073/pnas.0406069101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Bhalla R, Narasimhan K, Swarup S. Metabolomics and its role in understanding cellular responses in plants. Plant Cell Reports. 2005;24(10):562–571. doi: 10.1007/s00299-005-0054-9. [DOI] [PubMed] [Google Scholar]
- 77.Bedair M, Sumner LW. Current and emerging mass-spectrometry technologies for metabolomics. Trends in Analytical Chemistry. 2008;27(3):238–250. [Google Scholar]
- 78.Aharoni A, De Vos CHR, Verhoeven HA, et al. Nontargeted metabolome analysis by use of Fourier Transform Ion Cyclotron Mass Spectrometry. OMICS. 2002;6(3):217–234. doi: 10.1089/15362310260256882. [DOI] [PubMed] [Google Scholar]
- 79.Moco S, Bino RJ, Vorst O, et al. A liquid chromatography-mass spectrometry-based metabolome database for tomato. Plant Physiology. 2006;141(4):1205–1218. doi: 10.1104/pp.106.078428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Thiocone A, Farmer EE, Wolfender J-L. Screening for wound-induced oxylipins in Arabidopsis thaliana by differential HPLC-APCI/MS profiling of crude leaf extracts and subsequent characterisation by capillary-scale NMR. Phytochemical Analysis. 2008;19(3):198–205. doi: 10.1002/pca.1051. [DOI] [PubMed] [Google Scholar]
- 81.Farag MA, Huhman DV, Lei Z, Sumner LW. Metabolic profiling and systematic identification of flavonoids and isoflavonoids in roots and cell suspension cultures of Medicago truncatula using HPLC-UV-ESI-MS and GC-MS. Phytochemistry. 2007;68(3):342–354. doi: 10.1016/j.phytochem.2006.10.023. [DOI] [PubMed] [Google Scholar]
- 82.De Vos RCH, Moco S, Lommen A, Keurentjes JJB, Bino RJ, Hall RD. Untargeted large-scale plant metabolomics using liquid chromatography coupled to mass spectrometry. Nature Protocols. 2007;2(4):778–791. doi: 10.1038/nprot.2007.95. [DOI] [PubMed] [Google Scholar]
- 83.Kim JK, Bamba T, Harada K, Fukusaki E, Kobayashi A. Time-course metabolic profiling in Arabidopsis thaliana cell cultures after salt stress treatment. Journal of Experimental Botany. 2007;58(3):415–424. doi: 10.1093/jxb/erl216. [DOI] [PubMed] [Google Scholar]
- 84.Sanchez DH, Siahpoosh MR, Roessner U, Udvardi M, Kopka J. Plant metabolomics reveals conserved and divergent metabolic responses to salinity. Physiologia Plantarum. 2008;132(2):209–219. doi: 10.1111/j.1399-3054.2007.00993.x. [DOI] [PubMed] [Google Scholar]
- 85.Staszków A, Swarcewicz B, Banasiak J, Muth D, Jasiński M, Stobiecki M. LC/MS profiling of flavonoid glycoconjugates isolated from hairy roots, suspension root cell cultures and seedling roots of Medicago truncatula. Metabolomics. 2011;7(4):604–613. doi: 10.1007/s11306-011-0287-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Soga T. Capillary electrophoresis-mass spectrometry for metabolomics. Methods in Molecular Biology. 2007;358:129–137. doi: 10.1007/978-1-59745-244-1_8. [DOI] [PubMed] [Google Scholar]
- 87.Williams BJ, Cameron CJ, Workman R, Broeckling CD, Sumner LW, Smith JT. Amino acid profiling in plant cell cultures: an inter-laboratory comparison of CE-MS and GC-MS. Electrophoresis. 2007;28(9):1371–1379. doi: 10.1002/elps.200600609. [DOI] [PubMed] [Google Scholar]
- 88.Cevallos-Cevallos JM, Rouseff R, Reyes-De-Corcuera JI. Untargeted metabolite analysis of healthy and Huanglongbing-infected orange leaves by CE-DAD. Electrophoresis. 2009;30(7):1240–1247. doi: 10.1002/elps.200800594. [DOI] [PubMed] [Google Scholar]
- 89.Urakami K, Zangiacomi V, Yamaguchi K, Kusuhara M. Quantitative metabolome profiling of Illicium anisatum by capillary electro-phoresis time-of-flight mass spectrometry. Biomedical Research. 2010;31(2):161–163. doi: 10.2220/biomedres.31.161. [DOI] [PubMed] [Google Scholar]
- 90.Baranska M, Schulz H, Rösch P, Strehle MA, Popp J. Identification of secondary metabolites in medicinal and spice plants by NIR-FT-Raman microspectroscopic mapping. Analyst. 2004;129(10):926–930. doi: 10.1039/b408933m. [DOI] [PubMed] [Google Scholar]
- 91.Kaderbhai NN, Broadhurst DI, Ellis DI, Goodacre R, Kell DB. Functional genomics via metabolic footprinting: Monitoring metabolite secretion by Escherichia coli tryptophan metabolism mutants using FT-IR and direct injection electrospray mass spectrometry. Comparative and Functional Genomics. 2003;4(4):376–391. doi: 10.1002/cfg.302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Baranska M, Schulz H, Krüger H, Quilitzsch R. Chemotaxonomy of aromatic plants of the genus Origanum via vibrational spectroscopy. Analytical and Bioanalytical Chemistry. 2005;381(6):1241–1247. doi: 10.1007/s00216-004-3018-y. [DOI] [PubMed] [Google Scholar]
- 93.Schulz H, Baranska M, Quilitzsch R, Schütze W, Lösing G. Characterization of peppercorn, pepper oil, and pepper oleoresin by vibrational spectroscopy methods. Journal of Agricultural and Food Chemistry. 2005;53(9):3358–3363. doi: 10.1021/jf048137m. [DOI] [PubMed] [Google Scholar]
- 94.Schulz H, Schrader B, Quilitzsch R, Pfeffer S, Krüger H. Rapid classification of basil chemotypes by various vibrational spectroscopy methods. Journal of Agricultural and Food Chemistry. 2003;51(9):2475–2481. doi: 10.1021/jf021139r. [DOI] [PubMed] [Google Scholar]
- 95.Kim HK, Choi YH, Verpoorte R. NMR-based plant metabolomics: where do we stand, where do we go? Trends in Biotechnology. 2011;29(6):267–275. doi: 10.1016/j.tibtech.2011.02.001. [DOI] [PubMed] [Google Scholar]
- 96.Schripsema J. Application of NMR in plant metabolomics: techniques, problems and prospects. Phytochemical Analysis. 2010;21(1):14–21. doi: 10.1002/pca.1185. [DOI] [PubMed] [Google Scholar]
- 97.Kikuchi J, Shinozaki K, Hirayama T. Stable isotope labeling of Arabidopsis thaliana for an NMR-based metabolomics approach. Plant and Cell Physiology. 2004;45(8):1099–1104. doi: 10.1093/pcp/pch117. [DOI] [PubMed] [Google Scholar]
- 98.Sekiyama Y, Kikuchi J. Towards dynamic metabolic network measurements by multi-dimensional NMR-based fluxomics. Phytochemistry. 2007;68(16–18):2320–2329. doi: 10.1016/j.phytochem.2007.04.011. [DOI] [PubMed] [Google Scholar]
- 99.Leiss KA, Choi YH, Verpoorte R, Klinkhamer PGL. An overview of NMR-based metabolomics to identify secondary plant compounds involved in host plant resistance. Phytochemistry Reviews. 2011;10(2):205–216. doi: 10.1007/s11101-010-9175-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Gavaghan CL, Li JV, Hadfield ST, et al. Application of NMR-based metabolomics to the investigation of salt stress in maize (Zea mays) Phytochemical Analysis. 2011;22(3):214–224. doi: 10.1002/pca.1268. [DOI] [PubMed] [Google Scholar]
- 101.Ward JL, Baker JM, Beale MH. Recent applications of NMR spectroscopy in plant metabolomics. FEBS Journal. 2007;274(5):1126–1131. doi: 10.1111/j.1742-4658.2007.05675.x. [DOI] [PubMed] [Google Scholar]
- 102.Mochida K, Shinozaki K. Genomics and bioinformatics resources for crop improvement. Plant and Cell Physiology. 2010;51(4):497–523. doi: 10.1093/pcp/pcq027. [DOI] [PMC free article] [PubMed] [Google Scholar]