

Abstract
Human pluripotent stem cell-derived cardiomyocytes (hPS-CM) may offer a number of advantages over previous cardiac models, however, questions of their immaturity complicate their adoption as a new in vitro model. hPS-CM differ from adult cardiomyocytes with respect to structure, proliferation, metabolism and electrophysiology, better approximating fetal cardiomyocytes. Time in culture appears to significantly impact phenotype, leading to what can be referred to as early and late hPS-CM. This work surveys the phenotype of hPS-CM, including structure, bioenergetics, sensitivity to damage, gene expression, and electrophysiology, including action potential, ion channels, and intracellular calcium stores, while contrasting fetal and adult CM with hPS-CM at early and late time points after onset of differentiation.
Keywords: Cardiomyocyte, Pluripotent stem cells, Phenotype, In vitro cell culture, Maturity
Introduction
There is an urgent need for novel cardiomyocyte models: ischemic heart disease remains the #1 killer in the western world [1], congenital cardiomyopathies affect 1%–2% of live births [2, 3], and drug-induced cardiotoxicity is a leading cause of market withdrawal [4]. Human pluripotent stem cell-derived cardiomyocytes (hPS-CM) may offer significant advances in the study of cardiac disease and treatments [5, 6]. Similar to currently available cardiomyocyte models, hPS-CM contract rhythmically [7] and respond appropriately to numerous cardioactive drugs [5, 8]. In addition, hPS-CM can also be manipulated genetically [9], maintained in in vitro culture long-term (1+ years) [10], and be created from adult patients with genetic conditions (in the case of cardiomyocytes sourced from induced pluripotent stem cells, hiPS) [11–14] and may engraft into damaged hearts in vivo [15–17].
Given the potential of these cells and the excitement surrounding them (>2,000 publications since the first report a decade ago [18]), it is timely to address the similarity of these cells to adult human cardiomyocytes, and how they might be used as models of such. Open questions surrounding these cells include: How do we best assess cardiomyocyte maturity? How well do hPS-CM model embryonic or adult CM in vitro? How does maturity change during in vitro culture? When can hPS-CM be used as models for adult CM?
It is frequently noted that hPS-CM resemble human fetal cardiomyocytes [7]; however, no previous review has systematically quantified the similarities. This is complicated by high variation in phenotype between hPS-CM studies, partially explained by differences in cell line of origin and culture conditions. Furthermore, evidence suggests that hPS-CM develop a more mature, adult-like phenotype with time in culture, yet differences between early and late phase hPS-CM have not yet been described. Therefore, this review will define “early” and “late” phase hPS-CM phenotype, and describe how hPS-CM resemble embryonic and adult cardiomyocytes with respect to key markers of maturity, including ultrastructure, metabolism, gene expression, and electrophysiology.
hPS-CM Structure and Function Resemble Embryonic Cardiomyocytes
Definition of Early and Late Phase hPS-CM
In this work, hPS-CM will be defined as spontaneously contractile cells derived from a human pluripotent cell line, to the exclusion of contractile cells derived from adult mesenchymal stem cells [19–22] or from mouse pluripotent stem cells, which have been described elsewhere [23–25]. Recent reviews have covered methods to create [17, 26–29] and purify hPS-CM [30], as well as their electrophysiology [31], drug response [5, 8], and function after transplant in vivo [16, 32]. CM derived from hiPS (hiPS-CM) and human embryonic stem (hES-CM) cells appear to be relatively similar but will be compared when data describing differences are available.
hPS-CM vary in maturity, thus, we will define hPS-CM as either early phase, defined as contractile cells, with some proliferative capacity and with embryonic like electrophysiology (i.e., small negative membrane potential and small action potential amplitude), or late phase, defined by loss of proliferative capacity and more adult-like electrophysiology. hPS-CM show early phase characteristics for generally the first month after initiation of contraction, with development of late phase characteristics arising afterward. Different elements of maturity appear to be affected by line [33–35], time in culture [35, 36], cocultured cells [37], and culture conditions [38, 39]; however, the factors affecting maturity remain largely unknown. This suggests that after initiation of contraction, genetic and environmental factors interact leading to a more mature phenotype; however, the process is incompletely understood.
Morphology
It has been widely reported that hPS-CM structurally resemble embryonic or fetal cardiomyocytes [40, 41]. However, potentially important differences are seen when these cells are compared to embryonic or adult CM. Adult CM are large and cylindrical (approximately 150 × 10 μm for ventricular cells) [42], while embryonic and fetal CM are smaller [43]. Similarly, early hPS-CM (initiation of contraction, 21 days) are small and round to slightly oblong, approximately 5–10 μm in diameter [7, 33, 44] (Fig. 1). Late hPS-CM (>35 days) develop a more oblong morphology (30 μm × 10 μm), similar to the dimensions of human embryonic CM but remain small compared to adult [7]. In addition, most adult CM are bi- or multinucleated, whereas hPS-CM are mono-nuclear, similar to early embryonic cardiomyocytes [43].
Figure 1.
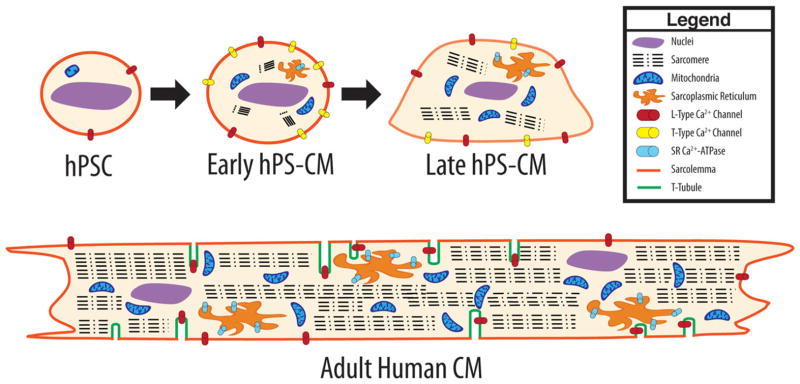
A visual comparison of early hPS-CM, late hPS-CM and adult CM morphology. Characteristics of hPS-CM depend strongly on time in culture since the initiation of contraction (early-proliferative, late-nonproliferative). Note that late hPS-CM differ from early hPS-CM with respect to shape, sarcomeric area and receptor expression. Adult CM are far larger, with multiple nuclei, large sarcomeric area, and large numbers of mitochondria. Abbreviations: hPSC, human pluripotent stem cell; hPS-CM, human pluripotent stem cell-derived cardiomyocytes.
The extensive t-tubule network present in adult ventricular CM is absent in both hPS-CM and embryonic CM [42]. As a result, excitation-contraction coupling is slower, and calcium primarily enters the cell through the sarcolemma instead of releasing from the sarcoplasmic reticulum (SR) [45–48]. Thus, early hPS-CM structurally resemble embryonic CM. With increasing time in culture, late hPS-CM develop a more adult-like morphology but do not appear to develop t-tubules or multinucleation (Fig. 1).
Function: Proliferation
Early hPS-CM proliferate [7, 39, 49], similar to embryonic or fetal mammalian cardiomyocytes [50–52]. In contrast, adult CM are among the most slowly dividing cell types [53]. Over time in culture, proliferative capacity of hPS-CM decreases from that of stem cells (24–48-hour doubling time [54]) to low levels: at 4 weeks, only 10% cells were BrdU+ after a 24-hour incorporation assay [39] and no Ki-67+ cells were observed [7], similar to changes seen in fetal cardiac development [50] (Fig. 2). Atrial natriuretic factor (involved in cardiomyocyte proliferation [55, 56]) is expressed in hPS-CM [57, 58]. In summary, early hPS-CM proliferate at a lower rate than their pluripotent progenitors whereas late hPS-CM can be considered nonproliferating cells.
Figure 2.

Visual comparison of early hPS-CM, late hPS-CM and adult CM phenotype. (A): An overview of major changes seen with increasing time in culture. Large changes in action potential characteristics (orange) are seen with time in culture, as well as expression of key ion channels (teal). (B): Calcium influx profiles for early and late hPS-CM compared with adult CM. Note that in early hPS-CM, almost no calcium is released from the sarcoplasmic reticulum, leading to slow, diffusion-limited calcium influx. Late hPS-CM perform better, but still show slow influx compared to adult. Abbreviation: hPS-CM, human pluripotent stem cell-derived cardiomyocytes.
Function: Gene Expression
The transcriptional profile of hPS-CM is starkly different from their originating pluripotent stem cells. Important differences include loss of pluripotency transcription factors and upregulation of mesodermal and cardiac markers [59–62]. Once differentiated, hPS-CM display a relatively homogeneous, cardiac-like gene expression program. Interestingly, gene expression of hiPS-CM and hES-CM is surprisingly similar, with only 1.9% of genes differentially expressed in these two cell types, despite dramatic differences between expression profiles in the undifferentiated hiPS and hES sources [63].
hPS-CM expression of contractile genes was not discernibly different from fetal heart tissue (20-week gestation) in one study of enriched early hPS-CM (age unknown) [59]. Global gene expression profile of purified early hPS-CM is more similar to fetal cardiac tissue (age unspecified) than to adult cardiac tissue; however, hPS-CM gene expression clustered more closely with either fetal or adult cardiac tissue than with pluripotent stem cells [64]. Bigger differences are seen when comparing hPS-CM and adult heart tissue, with important differences seen in a number of cardiac ion channel and calcium handling genes, once again highlighting the immature phenotype of hPS-CM [61, 65].
Function: Metabolism and Bioenergetics
Contractile machinery and mitochondria fill two-thirds of the cytoplasmic volume in adult CM (myofibril cell area, 40% [66] to 52% [67] and mitochondria, 15% [66] to 25% [67, 68]). In contrast, both hPS-CM [30] and embryonic CM [69] show smaller sarcomeric regions [58, 70] and have more moderate numbers of mitochondria [68] (Fig. 1). Similarly, expression of contractile and cytoskeletal genes is much lower in hPS-CM (unknown age) compared to fetal (20 weeks) or adult cardiomyocytes [59, 64].
Adult CM are highly metabolically active and depend on oxidative metabolism for synthesis of ATP (fatty acid oxidation accounts for 90% of acetyl-CoA production [71, 72]). In comparison, embryonic and fetal cardiomyocytes rely on glycolysis for production of ATP [69, 73] (fatty acid oxidation <15% of acetyl-CoA production [74]) resulting in a relatively hypoxia resistant phenotype and providing substrates for protein production [69]. hPS-CM showed primarily glycolytic metabolism in one study (in late, nonproliferative hPS-CM) which evaluated oxygen consumption rates [75] and mixed glycolytic and oxidative metabolism in another which assessed incorporation of radiolabeled carbon into metabolites (age unknown) [68]. hPS-CM are also able to metabolize lactate, unlike hPS [68]. Higher expression of oxidative phosphorylation genes and proteins are seen in hPS-CM compared to pluripotent stem cells, suggesting that these cells have the potential to use this metabolic pathway [68, 76, 77], though the expression level lags behind fetal tissue [59]. It remains unclear whether time in culture can alter hPS-CM preferred energy substrate.
Sensitivity to Damage and Apoptosis
It is unclear to what extent in vitro adult CM mimic the in vivo response to noxious stimuli. In vivo, adult CM may survive an entire lifetime (>80 years), while in vitro adult CM rarely survive more than a few days [78]. In stark contrast, hPS-CM are already culture-adapted, with reports of cells maintaining viability and contractility for a year [10, 79]. These observations clearly complicate comparisons of the sensitivity to damaging insults of hPS-CM with both in vivo and in vitro adult CM, and additional work is required to fully understand the differences in apoptotic cascades between these conditions.
Despite these limitations, some evidence suggests that hiPS-CM respond similarly to stimuli that cause damage to adult CM. For example, cardiotoxic tyrosine kinase inhibitors such as sunitinib and sorafenib demonstrate arrythmogenicity and increased apoptosis in hiPS-CM at clinically cardiotoxic doses of the drug [75, 80]. Likewise, doxorubicin, a cardiotoxic chemotherapeutic which is believed to act through oxidative stress [81], can induce apoptosis in hiPS-CM [82], as well as microtubule derangement [83]. Similarly, direct application of oxidizers such as hydrogen peroxide, induce apoptotic responses in hPS-CM [84, 85]. This process is mediated by opening of the mitochondrial permeability transition pore and could be prevented with anesthetic mediated preconditioning [84, 85], thus recapitulating the behavior seen in adult CM [86].
Embryonic human cardiomyocytes are resistant to hypoxia [87], whereas adult CM are highly dependent on an adequate oxygen supply [88]. As both hPS-CM and embryonic CM are predominantly glycolytic [75], it may be inferred that hPS-CM would likewise be resistant to hypoxia. However, the sensitivity of hPS-CM to ischemic stimuli has not been fully established. In summary, despite differences in metabolism, hPS-CM are sensitive to oxidative stress and cardiotoxic agents at levels expected from clinical use; however, their sensitivity to ischemia has not been characterized.
Cardic-Specific Inotropic and Chronotropic Receptors
Several key chronotropic responses are observed in hPS-CM and may be affected by time in culture. α, β1, and β2 adrenoceptor response have all been demonstrated in hPS-CM [89, 90]. A positive response to isoprenaline (β receptor agonist) challenge is almost universally performed in studies of hPS-CM [18, 34, 35, 58, 89–98], suggesting that all hPS-CM have some β-receptor expression, regardless of cell line of origin, differentiation method. As in vivo, isoprenaline increases contraction rate (positive chonotropy), increases the amplitude of the calcium transient, and decreases the relaxation time [90]. However, unlike adult CM, isoprenaline does not increase contraction force [98], once again demonstrating the immaturity of this cell type. β2 response accounts for 17%–37% [90] of the total response to isoprenaline, akin to fetal CM. With increased time in culture, hPS-CM demonstrated increased chronotropic β agonist response [90, 97]. In summary, β adrenoceptor response is present in hPS-CM and shares characteristics with fetal CM and may be amplified with time in in vitro culture.
Several studies have demonstrated a chronotropic response to carbacholine [89, 91, 92], thus showing muscarinic receptor activity. Finally, increased intraceullular cAMP increases contraction rate in hPS-CM via the phosphodiesterase inhibitor IBMX [18, 58] and the adenylyl cyclase activator forskolin [58, 89]. It is unclear whether in vitro maturation time affects the magnitude of these responses or whether these responses affect force of hPS-CM contraction.
Electrophysiology: Spontaneous Beating Rate
Spontaneous and synchronous contraction is seen as early as 5 days after the initiation of differentiation [99] and can be maintained for more than 1 year in culture [10] (in stark contrast to adult CM [78]). Different basal rhythms have been reported, ranging from 21 [93] to 52 beats per minute (BPM) [90], with most reporting ~40 BPM [35, 92, 100]. The rate of contraction may be affected by cell line, cultures conditions, time since differentiation, and time since the onset of contraction. hiPS-CM from iPS from patients with long QT syndrome show slower repolarization, thus recapitulating the in vivo phenotype [11, 12, 101–103].
Time in culture affects beating rate, though magnitude and direction of this change appears to vary with study. Several studies have reported moderate increases in contraction rate (30–75 BPM at 70 days [90] and 40–85 BPM at 60 days [35]) though a decrease has also been reported (45-5 BPM over the course of 63 days [92]). hES-CM show faster and stronger rhythms than hiPS-CM [35], which may be due to earlier initiation of contraction or the differences between hiPS and hES cells [63, 104]. In summary, spontaneous beating is the principal hallmark of differentiated hPS-CM, and beating rate is affected by line of origin and by time in culture.
Electrical Properties: Action Potential
hPS-CM contract spontaneously and synchronously, as noted previously, and are thus electrically active. Cells displaying atrial-, nodal-, and ventricular-like APs have been reported [105–107]. In addition, hPS-CM action potential characteristics vary between studies and within studies with different cell lines [35], differentiation methods [108], and time in culture [37]. Variation in a single population of hPS-CM has also been demonstrated, suggesting that even using the same cells, methods, and at the same time point, the electrophysiological characteristics of hiPS-CM are more heterogeneous than those in an adult heart [108]. hPS-CM from the same embryoid body (EB) (a more homogeneous environment) showed greater homogeneity in action potential duration than hPS-CM from same population but different EBs [35], highlighting the potential important role of a common extracellular environment in hPS-CM maturation.
Most reported action potential characteristics are less mature than adult CM: maximum diastolic potential (MDP) for adult ventricular myocytes is −85 mV [109], whereas early hPS-CM MDP is approximately −30 mV [37] which improves to −60 to −75 mV in late hPS-CM [35, 102, 110–113]. The maximum rate of depolarization (dv/dtmax or Vmax) in adult CM is extremely fast, ranging from 300 V/second in healthy hearts [109] to about 100 V/second in heart failure [114]. In contrast, early hPS-CM show extremely slow depolarization depolarization speeds. Early hPS-CM depolarize at 2 V/second [37], improving in late hPS-CM to 10–40 V/second [35, 102, 112, 113] (with two studies reporting 130–150 V/second [108, 115] -Fig. 2). Similar parameters for embryonic or fetal cardiomyocytes are not available.
Electrical Properties: Ion Channels
The major ionic currents normally present in adult CM are expressed in hPS-CM, though frequently at abnormal levels (Fig. 2). The calcium channels are necessary for contractility, as is NCX [116, 117] and HCN [110]. In early hPS-CM, sodium channel inhibition does not prevent spontaneous contraction, but in late hPS-CM the same inhibition blocked spontaneous contraction [37].
The potassium currents considered to be responsible for arrhythmias are expressed in hPS-CM [12, 102, 118] (Fig. 2). As a result, considerable interest in using hPS-CM for antiar-rhythmic drug screening exists and has been reviewed [8, 34, 101, 57]. Some arrhythmias in hPS-CM are affected by time in culture, and thus may be a measure of in vitro maturity [108, 119].
Electrical Properties: Intracellular Calcium
The extent of the SR and its necessity for automaticity in hPS-CM is a matter of debate. In adult CM, calcium induced calcium release (CICR) from the SR contributes almost 70% of the total calcium release [120]. In contrast, hPS-CM, which have very little SR function in the early phase [46, 121–125], demonstrate calcium transients that are smaller and slower [126], with most cation influx is through the cell membrane [124, 127]. This results in abnormal diffusion of calcium into the cell [122] and reduces the synchrony in contraction necessary for large force generation [98] (Fig. 2B).
Reports vary as to the presence and function of the SR, possibly due to changes with maturity [45–47, 121, 124]. However, there is consensus that intracellular calcium stores are smaller than in adult CM [121, 128]. Calcium handling and response to compounds that modify calcium handling (e.g., nifedipine and ryanodine) appear to vary significantly between lines [122, 129], and between embryonic and induced pluripotent derived CM [122, 130], including larger intracellular calcium stores, though line to line differences dominate differences between hES and hiPS class [35]. Over time in culture, increased sarcoplasmic reticulum function is seen as assessed by caffeine-induced calcium release [121].
When paced, adult CM show a positive force-frequency relationship; that is, at faster pacing rates, greater calcium transients and force of contraction are seen [120]. This relationship requires both significant intracellular calcium stores and electrical coordination across the cell (the t-tubule network again ensures that the entire cell depolarizes rapidly and homogeneously [120]). In contrast, hPS-CM have consistently shown negative force-frequency relationships [45, 97, 128]. In these cells, calcium primarily enters the cell across the cell membrane and diffuses though the cytoplasm, a slower process [42]. Similarly, postrest potentiation (i.e., an increased uptake in calcium in resting cardiomyocytes after rapid pacing) is not seen [45] or seen only to a low extent [97] in hPS-CM. It has not been studied whether these properties improve with time in culture, but the increased sarcoplasmic reticulum function seen in late hPS-CM suggests they may be more adult-like.
Some evidence suggests that non-SR calcium stores play a key role in excitation-contraction coupling in hPS-CM [46]. IP3 receptor (IP3R) is expressed and colocalizes with sarcomeres and the cell nucleus [121, 124], suggesting it may play a role in release of non-SR calcium stores. In adult CM, IP3R appears to regulate noncontractile calcium signaling only [131–133], although abnormal IP3R expression can cause arrhythmia [131]. In hPS-CM, IP3R may be involved in contractility as contraction rate is sensitive to IP3 and IP3R antagonists [121, 124]; however, this observation may depend on inhibition of ryanodine receptors (RYRs) [37].
Structural and Functional Sarcoplasmic Reticulum Proteins
The structural and functional proteins in the SR show low and varied expression as would be expected from the evidence provided earlier on the underdeveloped SR in hPS-CM. Expression of the RYR is noted in a number of studies [37, 134], though at only a small fraction (0.1%) of the adult level [121]. Most reports state that application of ryanodine slows spontaneous contraction rate [14, 42, 97, 121, 124, 127, 135, 136], though two studies saw no such change [122, 125]. Similarly, one study reports close physical association between RYRs and L-type calcium channels [127], which would allow for efficient CICR [127], though other studies reported no such association [42, 134]. It should be noted that the colocalization of these two proteins in adult CM is debated [137]. SERCA, the sarcoplasmic reticulum Ca2+ ATPase pump, is also expressed in hPS-CM [122, 138] at levels similar to fetal cardiomyocytes [134], but a variable response to its inhibitor thaspargin has been reported [45, 124].
Not surprisingly, proteins known to regulate SR function are also abnormally expressed in hPS-CM. Calsequestrin, which binds calcium and allows for dense packing of the ion in the SR, is absent in a number of studies [45, 46, 123, 134] though present in one [97]. Interestingly, transgenic calse-questrin overexpression was enough to improve calcium handling and SR maturity in hiPS-CM [123]. Phospholamban, an endogenous inhibitor of SERCA, is absent in some studies [45, 46], though present in others [138], and its presence is inferred from a positive drug response [90]. Some of this variability may be due to variable (widely unreported) hPS-CM age or manual selection of spontaneously beating cells, as more rapidly beating cells may have less phospholamban expression (in vivo phospholamban is known to repress cardiac contractility) [139, 140]. Junctin and triadin, which potentiate RYR [141], were expressed at low levels in one study [122] and absent in another [134].
Conclusions
hPS-CM are a heterogeneous population of cells that recapitulate some features of embryonic and adult CM. hPS-CM contract spontaneously and synchronously, express numerous cardiac specific genes and proteins, and recapitulate several important electrophysiological features of adult CM. Recapitulation of fetal or adult CM phenotype may require novel culture methods better recapitulating the in vivo niche. Furthermore, time in culture, specifically time since the onset of differentiation or time since spontaneous contraction, is a major factor affecting proliferation, structure, intracellular calcium stores, and ion channel expression. It is unclear why time in culture should have such profound effects on hPS-CM phenotype, though several studies have emphasized the importance of paracrine signaling and cellular milieu in maturation, suggesting better recapitulation of the cardiac cellular niche will improve maturity. Nonetheless, it is convenient to define early and late phase hPS-CM based on phenotypic markers that include sarcomeric organization, sarcoplasmic reticulum, and membrane ion channels that impact such integrated behaviors as cell proliferation and the action potential. Despite limitations, hPS-CM demonstrate significant potential as a tool to enhance basic biological understanding, improve in vitro drug screening, and thus create new therapeutic options. Remaining challenges include improving the magnitude and consistency of intracellular calcium stores, improving sarcomeric volume and organization, creating consistent reproducible cell populations, and determining the mechanisms of increased maturity with time in culture.
Acknowledgments
This work was supported in part by NIH UH2 TR000481 (SCG), NSF Graduate Research Fellowship Program (CR), and the ARCS Foundation Scholarship (CR).
Footnotes
Disclosure of Potential Conflicts of Interest
The authors indicate no potential conflicts of interest.
Author contributions: Claire Robertson, Conception and Design, Manuscript Writing; David Tran, Manuscript Writing, Graphic Design; Steven George, Financial Support, Final Approval of Manuscript.
References
- 1.Roger VL, Go AS, Lloyd-Jones D, et al. AHA statistical update heart disease and stroke statistics—2012 update. A report from the American Heart Association. Circulation. 2012;125:e2–e220. doi: 10.1161/CIR.0b013e31823ac046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bruneau BG. The developmental genetics of congenital heart disease. Nature. 2008;451:943–948. doi: 10.1038/nature06801. [DOI] [PubMed] [Google Scholar]
- 3.Lipshultz SE, Sleeper LA, Towbin JA, et al. The incidence of pediatric cardiomyopathy in two regions of the United States. N Engl J Med. 2003;348:1647–1655. doi: 10.1056/NEJMoa021715. [DOI] [PubMed] [Google Scholar]
- 4.Lexchin J. Drug withdrawals from the Canadian market for safety reasons, 1963–2004. CMAJ. 2005;172:765–767. doi: 10.1503/cmaj.045021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Denning C, Anderson D. Cardiomyocytes from human embryonic stem cells as predictors of cardiotoxicity. Drug Discov Today Ther Strateg. 2008;5:223–232. [Google Scholar]
- 6.Mandenius CF, Steel D, Noor F, et al. Cardiotoxicity testing using pluripotent stem cell-derived human cardiomyocytes and state-of-the-art bioanalytics: A review. J Appl Toxicol. 2011;31(3):191–205. doi: 10.1002/jat.1663. [DOI] [PubMed] [Google Scholar]
- 7.Snir M, Kehat I, Gepstein A, et al. Assessment of the ultrastructural and proliferative properties of human embryonic stem cell-derived cardiomyocytes. Am J Physiol Heart Circ Physiol. 2003;285:H2355–H2363. doi: 10.1152/ajpheart.00020.2003. [DOI] [PubMed] [Google Scholar]
- 8.Dick E, Rajamohan D, Ronksley J, et al. Evaluating the utility of cardiomyocytes from human pluripotent stem cells for drug screening. Biochem Soc Trans. 2010;38(4):1037–1045. doi: 10.1042/BST0381037. [DOI] [PubMed] [Google Scholar]
- 9.Faustino RS, Chiriac A, Niederlander NJ, et al. Decoded calreticulin-deficient embryonic stem cell transcriptome resolves latent cardio-phenotype. Stem Cells. 2010;28(7):1281–1291. doi: 10.1002/stem.447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Otsuji TG, Minami I, Kurose Y, et al. Progressive maturation in contracting cardiomyocytes derived from human embryonic stem cells: Qualitative effects on electrophysiological responses to drugs. Stem Cell Res. 2010;4(3):201–213. doi: 10.1016/j.scr.2010.01.002. [DOI] [PubMed] [Google Scholar]
- 11.Yazawa M, Hsueh B, Jia X, et al. Using induced pluripotent stem cells to investigate cardiac phenotypes in Timothy syndrome. Nature. 2011;471(7337):230–234. doi: 10.1038/nature09855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lahti AL, Kujala VJ, Chapman H, et al. Model for long QT syndrome type 2 using human iPS cells demonstrates arrhythmogenic characteristics in cell culture. Dis Model Mech. 2012;5(2):220–230. doi: 10.1242/dmm.008409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Itzhaki I, Maizels L, Huber I, et al. Modelling the long QT syndrome with induced pluripotent stem cells. Nature. 2011;471(7337):225–229. doi: 10.1038/nature09747. [DOI] [PubMed] [Google Scholar]
- 14.Jung CB, Moretti A, Mederos y Schnitzler M, et al. Dantrolene rescues arrhythmogenic RYR2 defect in a patient-specific stem cell model of catecholaminergic polymorphic ventricular tachycardia. EMBO Mol Med. 2012;4(3):180–191. doi: 10.1002/emmm.201100194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Caspi O, Gepstein L. Regenerating the heart using human embryonic stem cells—From cell to bedside. Isr Med Assoc J. 2006;8(3):208–214. [PubMed] [Google Scholar]
- 16.Liu J, Zhang Z, Liu Y, et al. Generation, characterization, and potential therapeutic applications of cardiomyocytes from various stem cells. Stem Cells Dev. 2012;21(12):2095–2110. doi: 10.1089/scd.2012.0031. [DOI] [PubMed] [Google Scholar]
- 17.Vidarsson H, Hyllner J, Sartipy P. Differentiation of human embryonic stem cells to cardiomyocytes for in vitro and in vivo applications. Stem Cell Rev. 2010;6(1):108–120. doi: 10.1007/s12015-010-9113-x. [DOI] [PubMed] [Google Scholar]
- 18.Xu C, Police S, Rao N, et al. Characterization and enrichment of cardiomyocytes derived from human embryonic stem cells. Circ Res. 2002;91(6):501–508. doi: 10.1161/01.res.0000035254.80718.91. [DOI] [PubMed] [Google Scholar]
- 19.Rose RA, Jiang H, Wang X, et al. Bone marrow-derived mesenchymal stromal cells express cardiac-specific markers, retain the stromal phenotype, and do not become functional cardiomyocytes in vitro. Stem Cells. 2008;26(11):2884–2892. doi: 10.1634/stemcells.2008-0329. [DOI] [PubMed] [Google Scholar]
- 20.Rangappa S, Fen C, Lee EH, et al. Transformation of adult mesenchymal stem cells isolated from the fatty tissue into cardiomyocytes. Ann Thorac Surg. 2003;75(3):775–779. doi: 10.1016/s0003-4975(02)04568-x. [DOI] [PubMed] [Google Scholar]
- 21.Heubach JF, Graf EM, Leutheuser J, et al. Electrophysiological properties of human mesenchymal stem cells. J Physiol. 2004;554(Pt 3):659–672. doi: 10.1113/jphysiol.2003.055806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Fukuda K. Reprogramming of bone marrow mesenchymal stem cells into cardiomyocytes. C R Biol. 2002;325(10):1027–1038. doi: 10.1016/s1631-0691(02)01524-x. [DOI] [PubMed] [Google Scholar]
- 23.Sachinidis A, Fleischmann BK, Kolossov E, et al. Cardiac specific differentiation of mouse embryonic stem cells. Cardiovasc Res. 2003;58(2):278–291. doi: 10.1016/s0008-6363(03)00248-7. [DOI] [PubMed] [Google Scholar]
- 24.Smith AG. Embryo-derived stem cells: of mice and men. Annu Rev Cell Dev Biol. 2001;17:435–462. doi: 10.1146/annurev.cellbio.17.1.435. [DOI] [PubMed] [Google Scholar]
- 25.Mummery C, Ward D, van den Brink CE, et al. Cardiomyocyte differentiation of mouse and human embryonic stem cells. J Anat. 2002;200(Pt 3):233–242. doi: 10.1046/j.1469-7580.2002.00031.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Boheler KR, Czyz J, Tweedie D, et al. Differentiation of pluripotent embryonic stem cells into cardiomyocytes. Circ Res. 2002;91(3):189–201. doi: 10.1161/01.res.0000027865.61704.32. [DOI] [PubMed] [Google Scholar]
- 27.Mummery CL, Zhang J, Ng ES, et al. Differentiation of human embryonic stem cells and induced pluripotent stem cells to cardiomyocytes: A methods overview. Circ Res. 2012;111(3):344–358. doi: 10.1161/CIRCRESAHA.110.227512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Jiang J, Han P, Zhang Q, et al. Cardiac differentiation of human pluripotent stem cells. J Cell Mol Med. 2012;16(8):1663–1668. doi: 10.1111/j.1582-4934.2012.01528.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.David R, Franz WM. From pluripotency to distinct cardiomyocyte subtypes. Physiology (Bethesda) 2012;27(3):119–129. doi: 10.1152/physiol.00044.2011. [DOI] [PubMed] [Google Scholar]
- 30.Habib M, Caspi O, Gepstein L. Human embryonic stem cells for cardiomyogenesis. J Mol Cell Cardiol. 2008;45(4):462–474. doi: 10.1016/j.yjmcc.2008.08.008. [DOI] [PubMed] [Google Scholar]
- 31.Blazeski A, Zhu R, Hunter DW, et al. Electrophysiological and contractile function of cardiomyocytes derived from human embryonic stem cells. Prog Biophys Mol Biol. 2012;110(2):178–195. doi: 10.1016/j.pbiomolbio.2012.07.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Barile L, Chimenti I, Gaetani R, et al. Cardiac stem cells: Isolation, expansion and experimental use for myocardial regeneration. Nat Clin Pract Cardiovasc Med. 2007;4(Suppl 1):S9–S14. doi: 10.1038/ncpcardio0738. [DOI] [PubMed] [Google Scholar]
- 33.Gherghiceanu M, Barad L, Novak A, et al. Cardiomyocytes derived from human embryonic and induced pluripotent stem cells: Comparative ultrastructure. J Cell Mol Med. 2011;15(11):2539–2551. doi: 10.1111/j.1582-4934.2011.01417.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Yokoo N, Baba S, Kaichi S, et al. The effects of cardioactive drugs on cardiomyocytes derived from human induced pluripotent stem cells. Biochem Biophys Res Commun. 2009;387:482–488. doi: 10.1016/j.bbrc.2009.07.052. [DOI] [PubMed] [Google Scholar]
- 35.Zhang J, Wilson GF, Soerens AG, et al. Functional cardiomyocytes derived from human induced pluripotent stem cells. Circ Res. 2009;104(3):e30–e41. doi: 10.1161/CIRCRESAHA.108.192237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Sartiani L, Bochet P, Cerbai E, et al. Functional expression of the hyperpolarization-activated, non-selective cation current if in immortalized HL-1 cardiomyocytes. J Physiol. 2002;545(Pt 1):81–92. doi: 10.1113/jphysiol.2002.021535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Kim C, Majdi M, Xia P, et al. Non-cardiomyocytes influence the electrophysiological maturation of human embryonic stem cell-derived cardiomyocytes during differentiation. Stem Cells Dev. 2010;19(6):783–795. doi: 10.1089/scd.2009.0349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Bauwens CL, Peerani R, Niebruegge S, et al. Control of human embryonic stem cell colony and aggregate size heterogeneity influences differentiation trajectories. Stem Cells. 2008;26(9):2300–2310. doi: 10.1634/stemcells.2008-0183. [DOI] [PubMed] [Google Scholar]
- 39.McDevitt TC, Laflamme MA, Murry CE. Proliferation of cardiomyocytes derived from human embryonic stem cells is mediated via the IGF/PI 3-kinase/Akt signaling pathway. J Mol Cell Cardiol. 2005;39(6):865–873. doi: 10.1016/j.yjmcc.2005.09.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Mummery C, Wardvan Oostwaard D, Doevendans P, et al. Differentiation of human embryonic stem cells to cardiomyocytes: Role of coculture with visceral endoderm-like cells. Circulation. 2003;107(21):2733–2740. doi: 10.1161/01.CIR.0000068356.38592.68. [DOI] [PubMed] [Google Scholar]
- 41.Fijnvandraat AC, van Ginneken AC, de Boer PA, et al. Cardiomyocytes derived from embryonic stem cells resemble cardiomyocytes of the embryonic heart tube. Cardiovasc Res. 2003;58(2):399–409. doi: 10.1016/s0008-6363(03)00282-7. [DOI] [PubMed] [Google Scholar]
- 42.Lieu DK, Liu J, Siu CW, et al. Absence of transverse tubules contributes to non-uniform Ca(2+) wavefronts in mouse and human embryonic stem cell-derived cardiomyocytes. Stem Cells Dev. 2009;18(10):1493–1500. doi: 10.1089/scd.2009.0052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Smolich JJ. Ultrastructural and functional features of the developing mammalian heart: A brief overview. Reprod Fertil Dev. 1995;7(3):451–461. doi: 10.1071/rd9950451. [DOI] [PubMed] [Google Scholar]
- 44.Gepstein L, Snir M, Kehat I, et al. Assessment of the ultrastructural and proliferative properties of human embryonic stem cell-derived cardiomyocytes. Am J Physiol Heart Circ Physiol. 2003;285(6):H2355–H2363. doi: 10.1152/ajpheart.00020.2003. [DOI] [PubMed] [Google Scholar]
- 45.Binah O, Dolnikov K, Sadan O, et al. Functional and developmental properties of human embryonic stem cells-derived cardiomyocytes. J Electrocardiol. 2007;40(6 Suppl):S192–S196. doi: 10.1016/j.jelectrocard.2007.05.035. [DOI] [PubMed] [Google Scholar]
- 46.Dolnikov K, Shilkrut M, Zeevi Levin N, et al. Functional properties of human embryonic stem cell derived cardiomyocytes: Intracellular Ca2+ handling and the role of sarcoplasmic reticulum in the contraction. Stem Cells. 2006;24(2):236–245. doi: 10.1634/stemcells.2005-0036. [DOI] [PubMed] [Google Scholar]
- 47.Fu JD, Li J, Tweedie D, et al. Crucial role of the sarcoplasmic reticulum in the developmental regulation of Ca2+ transients and contraction in cardiomyocytes derived from embryonic stem cells. FASEB J. 2006;20(1):181–183. doi: 10.1096/fj.05-4501fje. [DOI] [PubMed] [Google Scholar]
- 48.Itzhaki I, Schiller J, Beyar R, et al. Calcium handling in embryonic stem cell-derived cardiac myocytes: Of mice and men. Ann N Y Acad Sci. 2006;1080:207–215. doi: 10.1196/annals.1380.017. [DOI] [PubMed] [Google Scholar]
- 49.Cui L, Johkura K, Takei S, et al. Structural differentiation, proliferation, and association of human embryonic stem cell-derived cardiomyocytes in vitro and in their extracardiac tissues. J Struct Biol. 2007;158:307–317. doi: 10.1016/j.jsb.2006.11.009. [DOI] [PubMed] [Google Scholar]
- 50.Erokhina IL, Semenova EG, Emel’ianova OI. Human fetal ventricular cardiomyocytes in vitro: Proliferation and differentiation. Tsitologiia. 2005;47(3):200–206. [PubMed] [Google Scholar]
- 51.Horigome H, Takahashi MI, Asaka M, et al. Magnetocardiographic determination of the developmental changes in PQ, Qrs and Qt intervals in the foetus. Acta Paediatr. 2000;89(1):64–67. doi: 10.1080/080352500750029086. [DOI] [PubMed] [Google Scholar]
- 52.Porrello ER, Mahmoud AI, Simpson E, et al. Transient regenerative potential of the neonatal mouse heart. Science. 2011;331(6020):1078–1080. doi: 10.1126/science.1200708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Bergmann O, Bhardwaj RD, Bernard S, et al. Evidence for cardiomyocyte renewal in humans. Science. 2009;324(5923):98–102. doi: 10.1126/science.1164680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Cowan CA, Klimanskaya I, McMahon J, et al. Derivation of embryonic stem-cell lines from human blastocysts. N Engl J Med. 2004;350:1353–1356. doi: 10.1056/NEJMsr040330. [DOI] [PubMed] [Google Scholar]
- 55.Horio T, Nishikimi T, Yoshihara F, et al. Inhibitory regulation of hypertrophy by endogenous atrial natriuretic peptide in cultured cardiac myocytes. Hypertension. 2000;35(1 Pt 1):19–24. doi: 10.1161/01.hyp.35.1.19. [DOI] [PubMed] [Google Scholar]
- 56.Holtwick R, van Eickels M, Skryabin BV, et al. Pressure-independent cardiac hypertrophy in mice with cardiomyocyte-restricted inactivation of the atrial natriuretic peptide receptor guanylyl cyclase-A. J Clin Invest. 2003;111(9):1399–1407. doi: 10.1172/JCI17061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Tanaka T, Tohyama S, Murata M, et al. In vitro pharmacologic testing using human induced pluripotent stem cell-derived cardiomyocytes. Biochem Biophys Res Commun. 2009;385(4):497–502. doi: 10.1016/j.bbrc.2009.05.073. [DOI] [PubMed] [Google Scholar]
- 58.Kehat I, Kenyagin-Karsenti D, Snir M, et al. Human embryonic stem cells can differentiate into myocytes with structural and functional properties of cardiomyocytes. J Clin Invest. 2001;108(3):407–414. doi: 10.1172/JCI12131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Cao F, Wagner RA, Wilson KD, et al. Transcriptional and functional profiling of human embryonic stem cell-derived cardiomyocytes. PLoS One. 2008;3(10):e3474. doi: 10.1371/journal.pone.0003474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Synnergren J, Adak S, Englund MC, et al. Cardiomyogenic gene expression profiling of differentiating human embryonic stem cells. J Biotechnol. 2008;134:162–170. doi: 10.1016/j.jbiotec.2007.11.011. [DOI] [PubMed] [Google Scholar]
- 61.Synnergren J, Akesson K, Dahlenborg K, et al. Molecular signature of cardiomyocyte clusters derived from human embryonic stem cells. Stem Cells. 2008;26(7):1831–1840. doi: 10.1634/stemcells.2007-1033. [DOI] [PubMed] [Google Scholar]
- 62.Synnergren J, Giesler TL, Adak S, et al. Differentiating human embryonic stem cells express a unique housekeeping gene signature. Stem Cells. 2007;25(2):473–480. doi: 10.1634/stemcells.2006-0247. [DOI] [PubMed] [Google Scholar]
- 63.Gupta MK, Illich DJ, Gaarz A, et al. Global transcriptional profiles of beating clusters derived from human induced pluripotent stem cells and embryonic stem cells are highly similar. BMC Dev Biol. 2010;10:98. doi: 10.1186/1471-213X-10-98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Xu XQ, Soo SY, Sun W, et al. Global expression profile of highly enriched cardiomyocytes derived from human embryonic stem cells. Stem Cells. 2009;27:2163–2174. doi: 10.1002/stem.166. [DOI] [PubMed] [Google Scholar]
- 65.Synnergren J, Heins N, Brolen G, et al. Transcriptional profiling of human embryonic stem cells differentiating to definitive and primitive endoderm and further toward the hepatic lineage. Stem Cells Dev. 2010;19(7):961–978. doi: 10.1089/scd.2009.0220. [DOI] [PubMed] [Google Scholar]
- 66.Tashiro A, Masuda T, Segawa I. Morphometric comparison of mitochondria and myofibrils of cardiomyocytes between hypertrophic and dilated cardiomyopathies. Virchows Arch A: Pathol Anat Histopathol. 1990;416(6):473–478. doi: 10.1007/BF01600297. [DOI] [PubMed] [Google Scholar]
- 67.Barth E, Stammler G, Speiser B, et al. Ultrastructural quantitation of mitochondria and myofilaments in cardiac muscle from 10 different animal species including man. J Mol Cell Cardiol. 1992;24(7):669–681. doi: 10.1016/0022-2828(92)93381-s. [DOI] [PubMed] [Google Scholar]
- 68.Hattori F, Chen H, Yamashita H, et al. Nongenetic method for purifying stem cell-derived cardiomyocytes. Nat Methods. 2011;7(1):61–66. doi: 10.1038/nmeth.1403. [DOI] [PubMed] [Google Scholar]
- 69.Porter GA, Jr, Hom J, Hoffman D, et al. Bioenergetics, mitochondria, and cardiac myocyte differentiation. Prog Pediatr Cardiol. 2011;31(2):75–81. doi: 10.1016/j.ppedcard.2011.02.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Awasthi S, Matthews DL, Li RA, et al. Label-free identification and characterization of human pluripotent stem cell-derived cardiomyocytes using second harmonic generation (SHG) microscopy. J Biophotonics. 2012;5(1):57–66. doi: 10.1002/jbio.201100077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Giordano FJ. Oxygen, oxidative stress, hypoxia, and heart failure. J Clin Invest. 2005;115(3):500–508. doi: 10.1172/JCI200524408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Harris DA, Das AM. Control of mitochondrial ATP synthesis in the heart. Biochem J. 1991;280(Pt 3):561–573. doi: 10.1042/bj2800561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Lopaschuk GD, Collins-Nakai RL, Itoi T. Developmental changes in energy substrate use by the heart. Cardiovasc Res. 1992;26(12):1172–1180. doi: 10.1093/cvr/26.12.1172. [DOI] [PubMed] [Google Scholar]
- 74.Lopaschuk G, Jaswal J. Energy metabolic phenotype of the cardiomyocyte during development, differentiation and postnatal maturation. J Cardiovasc Pharmacol. 2010;56(2):130–140. doi: 10.1097/FJC.0b013e3181e74a14. [DOI] [PubMed] [Google Scholar]
- 75.Rana P, Anson B, Engle S, et al. Characterization of human induced pluripotent stem cell derived cardiomyocytes: Bioenergetics and utilization in safety screening. Toxicol Sci. 2012;130(1):117–131. doi: 10.1093/toxsci/kfs233. [DOI] [PubMed] [Google Scholar]
- 76.St John JC, Ramalho-Santos J, Gray HL, et al. The expression of mitochondrial DNA transcription factors during early cardiomyocyte in vitro differentiation from human embryonic stem cells. Cloning Stem Cells. 2005;7(3):141–153. doi: 10.1089/clo.2005.7.141. [DOI] [PubMed] [Google Scholar]
- 77.Chung S, Dzeja PP, Faustino RS, et al. Mitochondrial oxidative metabolism is required for the cardiac differentiation of stem cells. Nat Clin Pract Cardiovasc Med. 2007;4(Suppl 1):S60–S67. doi: 10.1038/ncpcardio0766. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Bistola V, Nikolopoulou M, Derventzi A, et al. Long-term primary cultures of human adult atrial cardiac myocytes: Cell viability, structural properties and BNP secretion in vitro. Int J Cardiol. 2008;131(1):113–122. doi: 10.1016/j.ijcard.2007.10.058. [DOI] [PubMed] [Google Scholar]
- 79.Mitcheson JS, Hancox JC, Levi AJ. Cultured adult cardiac myocytes: Future applications, culture methods, morphological and electrophysiological properties. Cardiovasc Res. 1998;39(2):280–300. doi: 10.1016/s0008-6363(98)00128-x. [DOI] [PubMed] [Google Scholar]
- 80.Cohen JD, Babiarz JE, Abrams RM, et al. Use of human stem cell derived cardiomyocytes to examine sunitinib mediated cardiotoxicity and electrophysiological alterations. Toxicol Appl Pharmacol. 2011;257(1):74–83. doi: 10.1016/j.taap.2011.08.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Simunek T, Sterba M, Popelova O, et al. Anthracycline-induced cardiotoxicity: Overview of studies examining the roles of oxidative stress and free cellular iron. Pharmacol Rep. 2009;61(1):154–171. doi: 10.1016/s1734-1140(09)70018-0. [DOI] [PubMed] [Google Scholar]
- 82.Andersson H, Steel D, Asp J, et al. Assaying cardiac biomarkers for toxicity testing using biosensing and cardiomyocytes derived from human embryonic stem cells. J Biotechnol. 2010;150(1):175–181. doi: 10.1016/j.jbiotec.2010.06.023. [DOI] [PubMed] [Google Scholar]
- 83.Shinozawa T, Furukawa H, Sato E, et al. A novel purification method of murine embryonic stem cell- and human-induced pluripotent stem cell-derived cardiomyocytes by simple manual dissociation. J Biomol Screen. 2012;17(5):683–691. doi: 10.1177/1087057111434145. [DOI] [PubMed] [Google Scholar]
- 84.Sepac A, Sedlic F, Si-Tayeb K, et al. Isoflurane preconditioning elicits competent endogenous mechanisms of protection from oxidative stress in cardiomyocytes derived from human embryonic stem cells. Anesthesiology. 2010;113(4):906–916. doi: 10.1097/ALN.0b013e3181eff6b7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Kim JH, Oh AY, Choi YM, et al. Isoflurane decreases death of human embryonic stem cell-derived, transcriptional marker Nkx2. 5(+) cardiac progenitor cells. Acta Anaesthesiol Scand. 2011;55(9):1124–1131. doi: 10.1111/j.1399-6576.2011.02509.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Ferdinandy P, Schulz R, Baxter GF. Interaction of cardiovascular risk factors with myocardial ischemia/reperfusion injury, preconditioning, and postconditioning. Pharmacol Rev. 2007;59(4):418–458. doi: 10.1124/pr.107.06002. [DOI] [PubMed] [Google Scholar]
- 87.Sedmera D, Kucera P, Raddatz E. Developmental changes in cardiac recovery from anoxia-reoxygenation. Am J Physiol Regul Integr Comp Physiol. 2002;283(2):R379–R388. doi: 10.1152/ajpregu.00534.2001. [DOI] [PubMed] [Google Scholar]
- 88.Kang PM, Haunstetter A, Aoki H, et al. Morphological and molecular characterization of adult cardiomyocyte apoptosis during hypoxia and reoxygenation. Circ Res. 2000;87(2):118–125. doi: 10.1161/01.res.87.2.118. [DOI] [PubMed] [Google Scholar]
- 89.Norstrom A, Akesson K, Hardarson T, et al. Molecular and pharmacological properties of human embryonic stem cell-derived cardiomyocytes. Exp Biol Med. 2006;231(11):1753–1762. doi: 10.1177/153537020623101113. [DOI] [PubMed] [Google Scholar]
- 90.Brito-Martins M, Harding SE, Ali NN. beta(1)- and beta(2)-adrenoceptor responses in cardiomyocytes derived from human embryonic stem cells: Comparison with failing and non-failing adult human heart. Br J Pharmacol. 2008;153(4):751–759. doi: 10.1038/sj.bjp.0707619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Mehta A, Chung YY, Ng A, et al. Pharmacological response of human cardiomyocytes derived from virus-free induced pluripotent stem cells. Cardiovasc Res. 2011;91(4):577–586. doi: 10.1093/cvr/cvr132. [DOI] [PubMed] [Google Scholar]
- 92.Reppel M, Boettinger C, Hescheler J. Beta-adrenergic and muscarinic modulation of human embryonic stem cell-derived cardiomyocytes. Cell Physiol Biochem. 2004;14(4–6):187–196. doi: 10.1159/000080326. [DOI] [PubMed] [Google Scholar]
- 93.Zwi L, Caspi O, Arbel G, et al. Cardiomyocyte differentiation of human induced pluripotent stem cells. Circulation. 2009;120(15):1513–1523. doi: 10.1161/CIRCULATIONAHA.109.868885. [DOI] [PubMed] [Google Scholar]
- 94.Zwi L, Caspi O, Arbel G, et al. Cardiomyocyte differentiation of human induced pluripotent stem cells. Hum Gene Ther. 2010;21(15):658–658. doi: 10.1161/CIRCULATIONAHA.109.868885. [DOI] [PubMed] [Google Scholar]
- 95.Xue T, Cho HC, Akar FG, et al. Functional integration of electrically active cardiac derivatives from genetically engineered human embryonic stem cells with quiescent recipient ventricular cardiomyocytes: Insights into the development of cell-based pacemakers. Circulation. 2005;111(1):11–20. doi: 10.1161/01.CIR.0000151313.18547.A2. [DOI] [PubMed] [Google Scholar]
- 96.Satin J, Kehat I, Caspi O, et al. Mechanism of spontaneous excitability in human embryonic stem cell derived cardiomyocytes. J Physiol. 2004;559(Pt 2):479–496. doi: 10.1113/jphysiol.2004.068213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Germanguz I, Sedan O, Zeevi-Levin N, et al. Molecular characterization and functional properties of cardiomyocytes derived from human inducible pluripotent stem cells. J Cell Mol Med. 2011;15(1):38–51. doi: 10.1111/j.1582-4934.2009.00996.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Pillekamp F, Haustein M, Khalil M, et al. Contractile properties of early human embryonic stem cell-derived cardiomyocytes: Beta-adrenergic stimulation induces positive chronotropy and lusitropy but not inotropy. Stem Cells Dev. 2012;10(21):2111–2121. doi: 10.1089/scd.2011.0312. [DOI] [PubMed] [Google Scholar]
- 99.Burridge PW, Thompson S, Millrod MA, et al. A universal system for highly efficient cardiac differentiation of human induced pluripotent stem cells that eliminates interline variability. PLoS One. 2011;6(4):e18293. doi: 10.1371/journal.pone.0018293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Xu RH, Chen X, Li DS, et al. BMP4 initiates human embryonic stem cell differentiation to trophoblast. Nat Biotechnol. 2002;20(12):1261–1264. doi: 10.1038/nbt761. [DOI] [PubMed] [Google Scholar]
- 101.Braam SR, Tertoolen L, van de Stolpe A, et al. Prediction of drug-induced cardiotoxicity using human embryonic stem cell-derived cardiomyocytes. Stem Cell Res. 2010;4(2):107–116. doi: 10.1016/j.scr.2009.11.004. [DOI] [PubMed] [Google Scholar]
- 102.Wang K, Terrenoire C, Sampson KJ, et al. Biophysical properties of slow potassium channels in human embryonic stem cell derived cardiomyocytes implicate subunit stoichiometry. J Physiol. 2011;589(24):6093–6104. doi: 10.1113/jphysiol.2011.220863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Moretti A, Bellin M, Welling A, et al. Patient-specific induced pluripotent stem-cell models for long-QT syndrome. N Engl J Med. 2010;363:1397–1409. doi: 10.1056/NEJMoa0908679. [DOI] [PubMed] [Google Scholar]
- 104.Bilic J, Izpisua Belmonte JC. Concise review: Induced pluripotent stem cells versus embryonic stem cells: Close enough or yet too far apart? Stem Cells. 2012;30(1):33–41. doi: 10.1002/stem.700. [DOI] [PubMed] [Google Scholar]
- 105.He JQ, Ma Y, Lee Y, et al. Human embryonic stem cells develop into multiple types of cardiac myocytes: Action potential characterization. Circ Res. 2003;93(1):32–39. doi: 10.1161/01.RES.0000080317.92718.99. [DOI] [PubMed] [Google Scholar]
- 106.Zhu WZ, Xie Y, Moyes KW, et al. Neuregulin/ErbB signaling regulates cardiac subtype specification in differentiating human embryonic stem cells. Circ Res. 2010;107(6):776–786. doi: 10.1161/CIRCRESAHA.110.223917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Moore JC, Fu J, Chan YC, et al. Distinct cardiogenic preferences of two human embryonic stem cell (hESC) lines are imprinted in their proteomes in the pluripotent state. Biochem Biophys Res Commun. 2008;372(4):553–558. doi: 10.1016/j.bbrc.2008.05.076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Pekkanen-Mattila M, Chapman H, Kerkela E, et al. Human embryonic stem cell-derived cardiomyocytes: demonstration of a portion of cardiac cells with fairly mature electrical phenotype. Exp Biol Med. 2010;235(4):522–530. doi: 10.1258/ebm.2010.009345. [DOI] [PubMed] [Google Scholar]
- 109.Drouin E, Lande G, Charpentier F. Amiodarone reduces transmural heterogeneity of repolarization in the human heart. J Am Coll Cardiol. 1998;32(4):1063–1067. doi: 10.1016/s0735-1097(98)00330-1. [DOI] [PubMed] [Google Scholar]
- 110.Yanagi K, Takano M, Narazaki G, et al. Hyperpolarization-activated cyclic nucleotide-gated channels and T-type calcium channels confer automaticity of embryonic stem cell-derived cardiomyocytes. Stem Cells. 2007;25(11):2712–2719. doi: 10.1634/stemcells.2006-0388. [DOI] [PubMed] [Google Scholar]
- 111.Pekkanen-Mattila M, Ojala M, Kerkela E, et al. The effect of human and mouse fibroblast feeder cells on cardiac differentiation of human pluripotent stem cells. Stem Cells Int. 2012 doi: 10.1155/2012/875059. [Epub ahead of print] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Zhang Q, Jiang J, Han P, et al. Direct differentiation of atrial and ventricular myocytes from human embryonic stem cells by alternating retinoid signals. Cell Res. 2011;21:579–587. doi: 10.1038/cr.2010.163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Ma J, Guo L, Fiene SJ, et al. High purity human-induced pluripotent stem cell-derived cardiomyocytes: Electrophysiological properties of action potentials and ionic currents. Am J Physiol Heart Circ Physiol. 2011;301(4):H2006–H2017. doi: 10.1152/ajpheart.00694.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Veldkamp MW, Verkerk AO, van Ginneken AC, et al. Norepinephrine induces action potential prolongation and early afterdepolarizations in ventricular myocytes isolated from human end-stage failing hearts. Eur Heart J. 2001;22(11):955–963. doi: 10.1053/euhj.2000.2499. [DOI] [PubMed] [Google Scholar]
- 115.Pekkanen-Mattila M, Kerkela E, Tanskanen JM, et al. Substantial variation in the cardiac differentiation of human embryonic stem cell lines derived and propagated under the same conditions—A comparison of multiple cell lines. Ann Med. 2009;41(5):360–370. doi: 10.1080/07853890802609542. [DOI] [PubMed] [Google Scholar]
- 116.Fu JD, Jiang P, Rushing S, et al. Na+/Ca2+ exchanger is a determinant of excitation-contraction coupling in human embryonic stem cell-derived ventricular cardiomyocytes. Stem Cells Dev. 2010;19(6):773–782. doi: 10.1089/scd.2009.0184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Zahanich I, Sirenko SG, Maltseva LA, et al. Rhythmic beating of stem cell-derived cardiac cells requires dynamic coupling of electro-physiology and Ca cycling. J Mol Cell Cardiol. 2011;50(1):66–76. doi: 10.1016/j.yjmcc.2010.09.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Caspi O, Itzhaki I, Kehat I, et al. In vitro electrophysiological drug testing using human embryonic stem cell derived cardiomyocytes. Stem Cells Dev. 2009;18(1):161–172. doi: 10.1089/scd.2007.0280. [DOI] [PubMed] [Google Scholar]
- 119.Gorelik J, Kadir SHSA, Ali NN, et al. Embryonic stem cell-derived cardiomyocytes as a model to study fetal arrhythmia related to maternal disease. J Cell Mol Med. 2009;13(9B):3730–3741. doi: 10.1111/j.1582-4934.2009.00741.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Bers DM. Cardiac excitation-contraction coupling. Nature. 2002;415(6868):198–205. doi: 10.1038/415198a. [DOI] [PubMed] [Google Scholar]
- 121.Satin J, Itzhaki I, Rapoport S, et al. Calcium handling in human embryonic stem cell-derived cardiomyocytes. Stem Cells. 2008;26:1961–1972. doi: 10.1634/stemcells.2007-0591. [DOI] [PubMed] [Google Scholar]
- 122.Lee YK, Ng KM, Lai WH, et al. Calcium homeostasis in human induced pluripotent stem cell-derived cardiomyocytes. Stem Cell Rev. 2011;7(4):976–986. doi: 10.1007/s12015-011-9273-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Liu J, Lieu DK, Siu CW, et al. Facilitated maturation of Ca2+ handling properties of human embryonic stem cell-derived cardiomyocytes by calsequestrin expression. Am J Physiol Cell Physiol. 2009;297(1):C152–C159. doi: 10.1152/ajpcell.00060.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Itzhaki I, Rapoport S, Huber I, et al. Calcium handling in human induced pluripotent stem cell derived cardiomyocytes. PLoS One. 2011;7(4):976–986. doi: 10.1371/journal.pone.0018037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Dolnikov K, Shilkrut M, Zeevi-Levin N, et al. Functional properties of human embryonic stem cell-derived cardiomyocytes. Ann N Y Acad Sci. 2005;1047:66–75. doi: 10.1196/annals.1341.006. [DOI] [PubMed] [Google Scholar]
- 126.Kang J, Chen X, Ji J, et al. Ca2+ channel activators reveal differential L-type Ca2+ channel pharmacology between native and stem cell-derived cardiomyocytes. J Pharmacol Exp Ther. 2012;341(2):510–517. doi: 10.1124/jpet.112.192609. [DOI] [PubMed] [Google Scholar]
- 127.Zhu WZ, Santana LF, Laflamme MA. Local control of excitation-contraction coupling in human embryonic stem cell-derived cardiomyocytes. PLoS One. 2009;4(4):e5407. doi: 10.1371/journal.pone.0005407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Xi J, Khalil M, Shishechian N, et al. Comparison of contractile behavior of native murine ventricular tissue and cardiomyocytes derived from embryonic or induced pluripotent stem cells. FASEB J. 2010;24(8):2739–2751. doi: 10.1096/fj.09-145177. [DOI] [PubMed] [Google Scholar]
- 129.Mandel Y, Weissman A, Schick R, et al. Human embryonic and induced pluripotent stem cell-derived cardiomyocytes exhibit beat rate variability and power-law behavior. Circulation. 2012;125(7):883–893. doi: 10.1161/CIRCULATIONAHA.111.045146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Matsa E, Denning C. In vitro uses of human pluripotent stem cell-derived cardiomyocytes. J Cardiovasc Transl Res. 2012;5(5):581–592. doi: 10.1007/s12265-012-9376-5. [DOI] [PubMed] [Google Scholar]
- 131.Roderick HL, Bootman MD. Pacemaking, arrhythmias, inotropy and hypertrophy: The many possible facets of IP3 signalling in cardiac myocytes. J Physiol. 2007;581(3):883–884. doi: 10.1113/jphysiol.2007.133819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Nakayama H, Bodi I, Maillet M, et al. The IP3 receptor regulates cardiac hypertrophy in response to select stimuli. Circ Res. 2010;107(5):659–666. doi: 10.1161/CIRCRESAHA.110.220038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Hund TJ, Ziman AP, Lederer WJ, et al. The cardiac IP3 receptor: Uncovering the role of “the other” calcium-release channel. J Mol Cell Cardiol. 2008;45(2):159–161. doi: 10.1016/j.yjmcc.2008.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Liu J, Fu JD, Siu CW, et al. Functional sarcoplasmic reticulum for calcium handling of human embryonic stem cell-derived cardiomyocytes: Insights for driven maturation. Stem Cells. 2007;25(12):3038–3044. doi: 10.1634/stemcells.2007-0549. [DOI] [PubMed] [Google Scholar]
- 135.Lee YK, Ng KM, Chan YC, et al. Triiodothyronine promotes cardiac differentiation and maturation of embryonic stem cells via the classical genomic pathway. Mol Endocrinol. 2010;24(9):1728–1736. doi: 10.1210/me.2010-0032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Ng KM, Lee YK, Chan YC, et al. Exogenous expression of HIF-1 alpha promotes cardiac differentiation of embryonic stem cells. J Mol Cell Cardiol. 2010;48(6):1129–1137. doi: 10.1016/j.yjmcc.2010.01.015. [DOI] [PubMed] [Google Scholar]
- 137.Bers DM. Excitation-Contraction Coupling and Cardiac Contractile Force. Dordrecht: Kluwer Academic Publishers; 2001. [Google Scholar]
- 138.Chaudhary KW, Barrezueta NX, Bauchmann MB, et al. Embryonic stem cells in predictive cardiotoxicity: Laser capture microscopy enables assay development. Toxicol Sci. 2006;90(1):149–158. doi: 10.1093/toxsci/kfj078. [DOI] [PubMed] [Google Scholar]
- 139.Luo W, Grupp IL, Harrer J, et al. Targeted ablation of the phospho-lamban gene is associated with markedly enhanced myocardial contractility and loss of beta-agonist stimulation. Circ Res. 1994;75(3):401–409. doi: 10.1161/01.res.75.3.401. [DOI] [PubMed] [Google Scholar]
- 140.MacLennan DH, Kranias EG. Phospholamban: A crucial regulator of cardiac contractility. Nat Rev Mol Cell Biol. 2003;4(7):566–577. doi: 10.1038/nrm1151. [DOI] [PubMed] [Google Scholar]
- 141.Gyorke I, Hester N, Jones LR, et al. The role of calsequestrin, triadin, and junctin in conferring cardiac ryanodine receptor responsiveness to luminal calcium. Biophys J. 2004;86(4):2121–2128. doi: 10.1016/S0006-3495(04)74271-X. [DOI] [PMC free article] [PubMed] [Google Scholar]