

Abstract
The concept of “lifestyle” includes different factors such as nutrition, behavior, stress, physical activity, working habits, smoking and alcohol consumption. Increasing evidence shows that environmental and lifestyle factors may influence epigenetic mechanisms, such as DNA methylation, histone acetylation and microRNA expression.
Several lifestyle factors have been identified that might modify epigenetic patterns, such as diet, obesity, physical activity, tobacco smoking, alcohol consumption, environmental pollutants, psychological stress, and working on night shifts.
Most studies conducted so far have been centered on DNA methylation, whereas only a few investigations have studied lifestyle factors in relation to histone modifications and miRNAs.
Here, we review current evidence indicating that lifestyle factors might affect human health via epigenetic mechanisms.
Keywords: Epigenetics, DNA methylation, Histone modifications, Environmental exposures, Lifestyle
Introduction
The term lifestyle is broadly used to describe the “typical way of life or manner of living characteristic of an individual or group” [1]. This concept includes different factors such as diet, behavior, stress, physical activity, working habits, smoking and alcohol consumption. Individual genetic background and environmental factors are intertwined to lifestyle in determining the health status of individuals (Figure 1). Increasing evidence shows that environmental and lifestyle factors may influence epigenetic mechanisms, such as DNA methylation, histone modifications and microRNA expression. Epigenetic mechanisms are flexible genomic parameters that can change genome function under exogenous influence but also provide a mechanism that allows for the stable propagation of gene activity states from one generation of cells to the next [2]. Alterations in epigenetic marks have also been associated with a variety of human diseases, including cancer, cardiovascular, respiratory and neurodegenerative diseases [3]. In this review we will discuss examples of lifestyle factors that have been investigated in relation to possible epigenetic effects, and the implication of lifestyle-related epigenetic changes in disease etiology (Table 1).
Fig. 1.
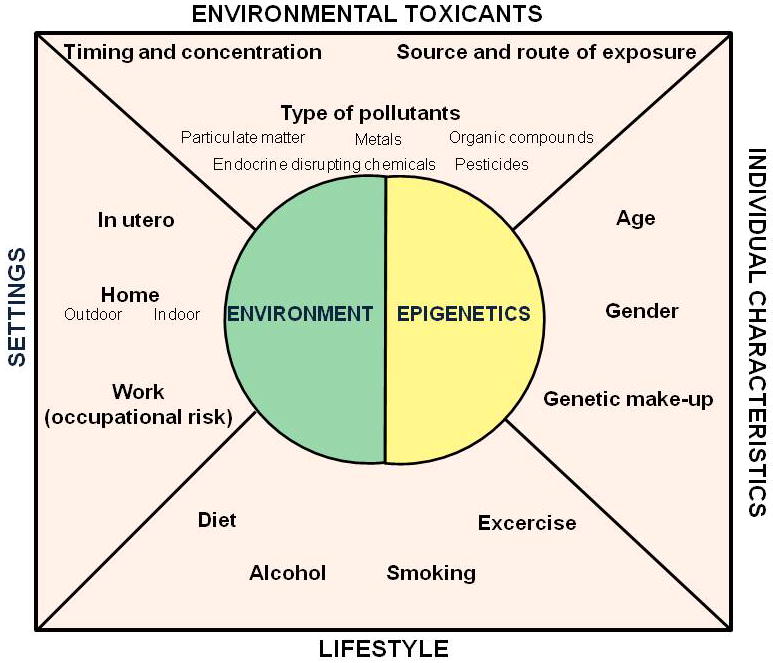
Environment-Epigenetics interactions
Table 1.
Lifestyle factors with epigenetic effects
| Factor | Example | Studies on: | Reference |
|---|---|---|---|
|
| |||
| Nutritional | Folate | humans | [16, 17] |
| Phytoestrogen | breast benign human cells | [36] | |
| human cancer cells | [35] | ||
| Polyphenols | human cancer cells | [27, 30–33] | |
| humans | 28, 29 | ||
| Selenium | human cancer cells | [38] | |
|
| |||
| Physical Activity | Exercise | human muscle biopsy tissues | [54] |
| humans | [51, 55] | ||
|
| |||
| Tobacco Smoke | Cigarette smoke | humans | [63] |
| lung cancer patients | [60] | ||
| Cigarette smoke condensate | placentas | [65] | |
| respiratory epithelia | [59] | ||
| rats and mice | [66] | ||
|
| |||
| Alcohol | High alcohol intake | humans | [73] |
|
| |||
| Pollutans | Arsenic | humans | [73–75] |
| PM10 | humans | [81, 87] | |
| Black carbon | humans | [86] | |
| Benzene | humans | [89] | |
| PAHs | humans | [93] | |
| human lymphoblastoid cells | [92] | ||
| human umbilical cord blood | [94] | ||
| POPs | humans | [95] | |
|
| |||
| Emotional | Stressful experiences | rats | [97] |
| mice | [100] | ||
| suicide victims | [101] | ||
|
| |||
| Shiftwork | Working at night | humans | [107] |
Foods
A possible role for nutrition in modifying epigenetic mechanisms has been examined in multiple investigations. For example, a diet rich in polyunsaturated fatty acids could generate mutagenic free radicals and oxidative stress [4], which has been directly linked to epigenetic alterations [5, 6]. Modulation of gene methylation has been observed in human endothelial cells incubated with arachidonic acid promoting up-regulation of a pro-angiogenic mechanisms [7]. Conversely, polyunsaturated fatty acids may have a suppressive function in tumorigenic processes through dampening of inflammation and NF-kappaB pathway [8]. Moreover, diets rich in fruits and vegetables, which contain many natural anti-oxidants, can yield anticancer protection [9]. Chen and Xu [10] have extensively reviewed the potential epigenetic effects of several nutritional components, mostly derived from vegetables. For instance, a study in healthy human subjects fed with a single serving of broccoli sprouts showed inhibition of histone deacetylase activity in circulating peripheral blood mononuclear cells 3–6 hours after consumption, with concurrent induction of histone H3 and H4 acetylation [11]. An in-vitro study on human tumor colon cell lines revealed that high doses of diallyl-disulfide from garlic increased histone H3 and H4 acetylation [12].
Folate and Vitamin B12 Intake
Folic acid and Vitamin B12 play an important role in DNA metabolism and are required for the synthesis of methionine and S-adenosylmethionine (SAM), the common methyl donor required for the maintenance of methylation patterns in DNA [13]. Methylation reactions could be influenced through the modification of the ratio between S-adenosylmethionine (SAM) and S-adenosylhomocysteine (SAH) ratio [14]. The SAM:SAH ratio is a primary determinant of the methylation capacity because SAM is converted to SAH by methionine adenosyltransferase. Taking into consideration that methionine is regenerated by methylation of homocysteine via the folate and B12 dependent reactions, a folate-deficient diet could interfere with this system [15]. The SAM/SAH ratio has been related with DNA methylation patterns. For example, a study conducted in rats examined the maternal folate status and DNA methylation in placenta. A significant positive correlation was found between placental DNA methylation, hepatic and plasma folate levels, and hepatic SAM/SAH ratio [16].
Low folate intakes have been associated with risk of colorectal cancer [17]. Moreover, folate depletion has been shown to cause lymphocyte DNA hypomethylation in healthy postmenopausal women, an alteration reverted by folate repletion [18, 19]. A recent study carried out among individuals susceptible to folate deficiency showed that methylation status can be corrected with choline supply at higher-than-recommended doses (500 mg/day) for 12-weeks [20]. In the context of the SAM cycle, choline can donate methyl groups to SAM since it is a trimethylated molecule. Folate has also been shown to reverse the dysregulation of miRNA expression associated with hepatocellular carcinogenesis, potentially by restoring dietary methyl donors [21]. The growing body of evidence showing that folate intake modulate epigenetic mechanisms has been actively investigated in relation to potential anticarcinogenic properties suggested by epidemiological studies [22–25]. Also, investigating epigenetic effects from folate might help understand paradoxical observations, such as those linking very high folate intakes with the development of colorectal carcinoma from adenomatous polyps [26].
Polyphenols
Polyphenols are a large family of natural compounds widely distributed in plant foods, that have been shown to modify the activity of DNA methyltransferases, histone acetylases (HATs) and histone deacetylases (HDACs) [27, 28]. In particular, studies on cancer cells have shown that polyphenols can reverse in in-vitro models some of the epigenetic aberrations associated with malignant transformation [29]. Inhibitory effects on DNA methyltransferases have been observed both in-vitro and in-vivo using different dietary sources of polyphenols [28]. A retrospective analysis associated CDX2 and BMP-2 gene hypermethylation with past low intake of polyphenol sources such as cruciferous vegetables and green tea in patients with primary gastrocarcinoma [30, 31]. Green tea contains (−)-epigallocatechin-3-gallate (EGCG), which is an inhibitor of DNA methyltransferases activity. EGCG has been shown to reactivate methylation-silenced genes in cancer cell lines [32, 33]. In in-vitro experiments on esophageal, prostate, colon and breast cancer cells lines, several CpG islands in various loci were efficiently demethylated by EGCG, thus leading to the expression of previously silenced genes [32–35].
Soy beans are also extremely rich in polyphenols [28]. Soy polyphenols include phytoestrogens such as genistein, biochanin A, and daidzein [36]. These compounds have also been shown to inhibit DNA methyltransferases and histone deacetylases in cancer cell lines and to revert aberrant CpG island methylation [37]. Li et al. showed in benign (MCF-10AT) and cancer (MCF7) breast cells that all the three main DNA methyltransferases (DNMT1, DNMT3a, and DNMT3b) were down regulated by genistein [38]. These results might help explain epidemiology data indicating that soy consumption is associated with reduced risk of hormone-related cancers [39].
Selenium
Selenium can epigenetically modulate DNA and histones to activate methylation-silenced genes [40]. Increasing data suggest that selenium may have anticarcinogenic properties through modifications of epigenetic processes in the cell [41–43]. Selenium has been shown to directly inhibit DNMT expression and activity [44, 45]. Selenium can also restore the expression of hypermethylated genes, such as GSTP1, APC and CSR1, in human prostate cancer cells by downregulating DNMTs and inhibiting HDAC activity [40]. These genes are known to have anticancer activity by protection against oxidative damage, detoxification of carcinogenetic chemicals or tumor suppression [40]. Moreover, in animal models, a selenium-deficient diet has been shown to induce DNA hypomethylation [46, 47].
Obesity and Physical Activity
Overweight, obesity, and sedentary lifestyle are established and prevalent risk factors for several diseases, including cancer and cardiovascular disease [48–50]. Because body weight is regulated by genes controlling energy homeostasis, it has been hypothesized that dietary macronutrients that affect DNA methylation could contribute to develop obesity through epigenetic mechanisms [51]. Epigenetic biomarkers of obesity, including genes involved in adipogenesis, (SOCS1/SOCS3), methylation pattern of obesity-related genes (FGF2, PTEN, CDKN1A and ESR1), inflammation genes as well as intermediary metabolism and insulin signaling pathway genes, could help to predict susceptibility and prevent obesity [52].
Emerging evidence indicates that epigenetic mechanisms may be involved in mediating effects of physical activity. In a recent work, physical activity was associated with higher methylation in peripheral blood lymphocytes of LINE-1 elements, a class of repeated sequences highly repeated in the human genome [53]. Low methylation in LINE-1 repetitive elements has been associated with inflammatory responses, as well as with chromosomal instability [54]. Interestingly, elderly individuals with high LINE-1 methylation in peripheral blood lymphocytes have been recently shown to have lower incidence and mortality from ischemic heart disease and stroke [55]. Whether the decreased cardiovascular risks associated with LINE-1 methylation reflect beneficial effects from physical activity remains to be determined. In human muscle biopsies following exercise, a global increase in H3K36 acetylation has also been observed [56]. Moreover, a brief exercise has been shown to alter miRNA profiles in circulating neutrophils in humans, including 38 miRNAs involved in inflammatory pathways [57].
Tobacco smoke
Tabacco smoke contains a complex mixture of organic and inorganic chemicals, many of which have carcinogenic, pro-inflammatory and proaterogenic properties. Individual effects of these components have been examined through different epigenetic studies, but the results are still inconclusive. For example, an in-vitro chronic toxicity study of normal human fibroblast on Benzo[a]pyrene - a prominent carcinogenic polycyclic aromatic hydrocarbon (PAH) found in cigarette smoke - found no aberrant patterns of DNA methylation in genomic regions of relevance for lung cancer [58].
Conversely, cigarette smoke condensate has been shown in respiratory epithelial cells to decrease the nuclear levels of certain histone modifications such as H4K16 acetylation and H4K20 trimethylation [59]. These alterations were similar to changes in histone modifications that can be found in lung cancer tissues which commonly precede aberrant DNA methylation [60, 61]. For instance, demethylation in H19 and IGF2 occurred primarily to the DNA hypermethylation-mediated silencing of p16, MGMT, DAPK, E-cadherin, and cdh13 tumor suppressor genes as an early event in lung carcinonegesis induced by tobacco smoke [62].
P53 hypomethylation has been reported in peripheral blood lymphocytes of smoking lung cancer patients [63]. Despite the lack of consistent evidence for p53 gene aberrantly methylated in human cancer, p53 hypomethylation has been associated with early events in carcinogenesis such as DNA double-strand breaks and chromosomal instability [64, 65].
A study that evaluated global DNA methylation from buccal cells of children exposed to prenatal maternal smoking demonstrated hypomethylation of LINE-1 repetitive elements. In the same study, a microarray analysis of 1536 CpG sites identified differential methylation of CpG loci in eight genes. Two of them, AXL and PTPRO, were validated by pyrosequencing and showed significant increases in methylation [66]. Following findings indicating that miRNAs in human placentas are differentially expressed in association with adverse pregnancy outcomes [67], a recent study found that candidate miRNAs implicated in growth and developmental processes (i.e., miR-16, miR-21, and miR-146a) were significantly downregulated in cigarette smoke-exposed placentas compared to controls [68]. Moreover, downregulation of microRNA expression was also observed in animal experiments when lung of mice and rats were exposed to cigarette smoke. In this study, mir-34b, mir-345, mir-421, mir-450b, mir-466, and mir-469 were downregulated at high-dose of exposure; however, expression was restored one week after smoking cessation [69].
Alcohol consumption
In contrast to polycyclic aromatic hydrocarbons (PAHs) and other carcinogenic molecules found in tobacco smoke and tar, ethyl alcohol is not per se mutagenic, but rather acts mainly as a cocarcinogen [70]. A Netherlands cohort study on diet and cancer correlated the intake of folate and alcohol with changes in methylation of tumor suppressor and DNA repair genes (APC-1A, p14ARF, p16INK4A, hMLH1, O6-MGMT, and RASSF1A) in paraffin-embedded colorectal cancer tissues [71]. Also, this work suggested the association between the intake of other methyl donors such as methionine, vitamins B6, and B12 with an increased frequency of promoter hypermethylation of genes involved in colorectal carcinogenesis [71]. However, a second cohort study did not find any association of folate intake, methionine or alcohol with MLH1 hypermethylation, a frequent and well-characterized early event in the development of colorectal cancer [72]. A positive association between vitamin B6 intake and tumors showing MLH1 hypermethylation was found, suggesting B6 vitamin may enhance colorectal cancer risk [72]. Alcohol consumption has also been suggested to modify the association between blood markers of DNA methylation and disease. In a population-based case-control study on a Polish population, Hou et al. showed that repetitive elements hypomethylation in blood leukocyte DNA was associated with gastric cancer and that the association between LINE-1 hypomethylation and gastric cancer was stronger among individuals who were current alcohol drinkers [73].
Currently, there are demonstrations of alcohol effects on growth and neuronal development through epigenetic marks. Mouse fetal cortical neurons chronically exposed to ethanol in vitro, had NR2B gene demethylation which encodes an ionotropic glutamate receptor possibly involved in certain memory and learning processes [74, 75]. Instead, acute exposure to ethanol induced hypermethylation of specific cell cycle genes inhibiting the growth factor-regulated cell cycle progression in monolayer cultures of neural stem cells. Lengthening the time between G1 and S phase was observed when cells were exposed for 48 h [76]. In the mouse strain C57BL/6, alcohol exposure at early embryonic altered the DNA methylation in embryos with a neural tube defect phenotype changing the expression for genes involved in metabolism and development such as Nlgn3, Elavl2, Sox21, Sim1, Nlgn3, Elavl2, Sox21 and Sim1. These disturbances may contribute to malformations and abnormal fetal development [77]. Subsequently, Zhou et al. found a reduction in expression of neurogenin, Sox5, Bhlhe22, Igf1, Efemp1, Tieg and Edil3 in mice embryo cultures. In this case, the gene expression responsible for the neural tube development is modulated by changes in DNA methylation patterns [78].
Environmental pollutants
In environmental studies, the flexibility of epigenetic states has generated growing interest in evaluating whether environmental exposures can modify epigenetic states, including DNA methylation and histone modifications [79]. Studies of DNA methylation and histone modification in relation to environmental exposures to potentially toxic chemicals have been examined in detail in a recent review article [80]. Here, we briefly review the main classes of environmental exposures that are most frequently considered epigenetic toxicants.
Arsenic
In a human study from India, significant DNA hypermethylation of p53 and p16 promoter regions was observed in blood DNA of subjects exposed to toxic arsenic levels compared to controls [81]. In this study, p53 and p16 hypermethylation showed a dose-response relationship with arsenic measured in drinking water. A large body of in vitro and animal studies have shown that arsenic subtracts methyl donors from DNA methylation reactions and induces global DNA hypomethylation [82]. An unexpected finding was recently reported in vivo, as a global dose-dependent hypermethylation of blood DNA was observed in Bangladeshi adults with chronic arsenic exposure. This effect was modified by folate, suggesting that arsenic-induced increases in DNA methylation were dependent from methyl availability [82]. The same group, however, subsequently reported that lower blood DNA methylation was a strongly associated with arsenic-induced skin lesions in a related Bangladeshi population [83].
Air pollution
Exposure to air pollution, particularly to particulate matter (PM), has been associated with increased morbidity and mortality from cardiorespiratory disease, as well as with increased lung cancer risk [84–88]. In a human study, Tarantini et al recently demonstrated that iNOS (inducible Nitric Oxide Synthase) promoter methylation decreased in blood samples of foundry workers with well-characterized exposure to PM10 in samples taken at the end of a four-day work week compared to baseline samples [89]. iNOS demethylation is expected to increase expression and activity of the iNOS protein, an established key player in inflammation and oxidative stress generation, two primary mechanisms that have been suggested to link inhalation of air pollutants to their acute health effects [90–92]. In the same study, long-term exposure to PM10 was negatively associated with methylation in both Alu and LINE-1 [89]. Decreased LINE-1 methylation was also observed in association to exposure to black carbon (BC), a marker of traffic particles, on 1,097 blood DNA samples from the Normative Aging Study (NAS), a repeated measure investigation of elderly men in the Boston area. As blood LINE-1 hypomethylation has been found in patients with cancer [93] and cardiovascular disease [94], such changes may reproduce epigenetic processes related to disease development and represent mechanisms by which particulate air pollution affects human health [94]. A recent occupational study has recently examined the effects of exposure to PM and metal components on miRNAs expression in 63 workers at an electric-furnace steel plant. miR-222 and miR-21 – two candidate miRNAs related to oxidative stress and inflammation – were overexpressed and positively correlated with the levels of lead exposure and oxidative DNA damage, respectively [95].
Aromatic hydrocarbons and other organic pollutants
High-level exposure to benzene has been associated with increased risk of acute myelogenous leukemia (AML) [96], which is characterized by aberrant global hypomethylation and gene-specific hypermethylation/hypomethylation. In a study of gasoline station attendants and traffic police officers, airborne benzene exposure was shown to be associated with a significant reduction in LINE-1 and Alu methylation in peripheral blood DNA [97]. Airborne benzene was also associated with hypermethylation in p15 and hypomethylation of the MAGE-1 cancer-antigen gene [97]. These findings show that benzene exposure at relatively low levels may induce altered DNA methylation reproducing the aberrant epigenetic patterns found in malignant cells. Also, benzene-associated demethylation of repetitive elements may help explain the epidemiological data linking benzene exposure with increases risk of multiple myeloma [98, 99], which also exhibits reduced methylation in Alu e LINE-1 repetitive elements [97]. These human data were recently confirmed by the finding of global hypomethylation in human TK6 lymphoblastoid cells treated for 48 hours with hydroquinone, one of the active benzene metabolites [100]. In a study of Polish male nonsmoking coke-oven workers, chronic exposure to PAHs has been shown to modify the methylation status of specific gene promoters (p53, p16, HIC1 and IL-6), as well as of Alu and LINE-1 repetitive elements [101]. Perera et al. published an exploratory study that used methylation sensitive restriction fingerprinting to analyze umbilical cord white blood cell DNA of 20 children exposed to PAHs. Over 30 DNA sequences were identified whose methylation status was dependent on the level of maternal PAH exposure [102]. Rusiecki et al. evaluated the relationship between plasma concentrations of persistent organic pollutants and blood global DNA methylation, estimated in Alu repeated elements, in 70 Greenlandic Inuit, a population presenting some of the highest reported levels of POPs worldwide. In this work, a significant inverse linear relationship was found for DDT, DDE, β-BHC, oxychlordane, α-chlordane, mirex, several PCBs, and sum of all POPs [103].
Psychological stress
Earlier studies have indicated that DNA methylation is sensitive to environmental stressful exposures in early development and later in life [104–109]. The glucocorticoid receptor gene promoter was studied in the hippocampus of human suicide victims and controls [109]. Hypermethylation of the glucocorticoid receptor gene was found among suicide victims with a history of abuse in childhood, but not among controls or suicide victims with a negative history of childhood abuse [109]. On the contrary, positive early social experience might have a mitigating effect on stress responses later in life via epigenetic mechanisms, suggesting a protective role for positive early parental care [110, 111]. This is shown in animal studies that have demonstrated that higher maternal care, as reflected in higher licking and grooming of the pups, induces hypomethylation of the glucocorticoid receptor gene in the hippocampus and reduces responses to stress [110].
Shiftwork
Recent advances in the epigenetic field have revealed that chronobiological regulators may induce chromatin remodelling [see review 112]. CLOCK gene regulates circadian rhythm through a histone-acetyltransferase activity which promotes chromatin-remodelling events implicated in circadian control of gene expression [113, 114]. The circadian adjustment may be affected by different factors such as shift-work. According to several epidemiological studies shift-work that requires working at night can have a negative impact on the health and well-being of workers due to a mismatch between the endogenous circadian timing system and the environmental synchronizers (e.g. light/dark cycle) [115]. An epigenetic reprogramming of circadian genes has been proposed as a potential response altered circadian rhythms [116, 117]. A recent study on a population of night-shift workers has shown alterations in blood DNA methylation, including changes in Alu repetitive elements methylation and gene-specific methylation of inflammatory genes such as IFN-γ and TNF-α [118].
Conclusions and Future Perspectives
In the last few years, several investigations have examined the relation between epigenetic marks and lifestyle factors, including nutrition, behavior, stress, physical activity, working habits, smoking and alcohol consumption. Although epigenetic modifications are influenced by the environment, most of these changes tend to be re-established each generation; however, this does not happen at some loci in the human genome [119, 120]. The possibility that this phenomenon impacts successive generations is referred as transgenerational epigenetic inheritance [121–124]. Epigenetics is expected to help explaining how gene expression is modulated by lifestyle and environmental factors, and to bring a more complete understanding of individual responses to environmental cues and acquired risk factors (Figure 1). Because both epigenetic mechanisms and lifestyle are modifiable, epigeneticists have largely untapped opportunities to determine how tightly epigenetic markers are dependent on lifestyle factors and whether and how much epigenetic mechanisms can be modified after positive or negative lifestyle changes are acquired and sustained (Figure 2). Considering that epidemiological research are moving into new technologies such as epigenetics, many of the studies cited here should be taken as presumptive while there is no further evidence.
Fig. 2.
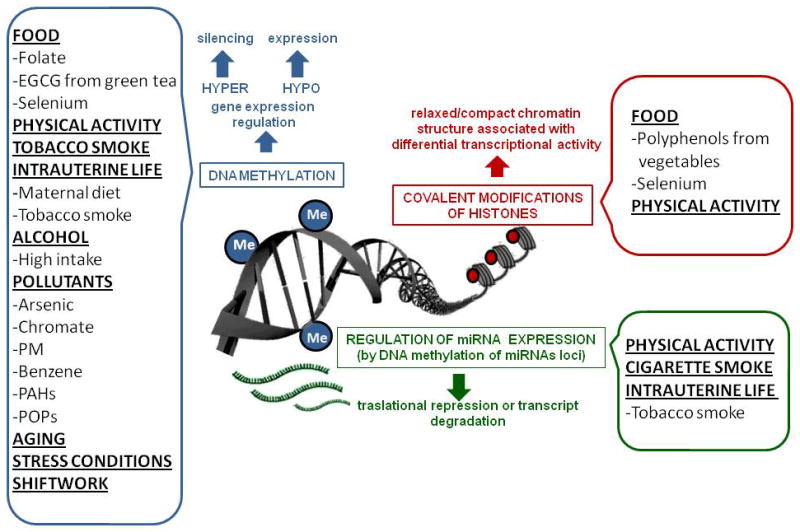
Lifestyle factors participating in environment-epigenetic interactions.
Executive summary.
Introduction
Lifestyle includes different factors such as nutrition, behavior, stress, physical activity, working habits, smoking and alcohol consumption.
Environmental and lifestyle factors may influence epigenetic mechanisms.
Nutrition
Folate and Vitamin B12 Intake
Epidemiological data support the anticarcinogenic property of folate.
A protective effect of low folate status against colorectal cancer was reported.
These contrasting results suggest that folic acid supplementation could exert a negative effect on already existing lesions.
Polyphenols
Polyphenols can impact DNA methyltransferases, Histone acetylases and Histone deacetylases inducing reversibility of epigenetic dysregulation.
Selenium
Selenium can impact the DNA methylation status interacting directly with DNA methyltransferases.
Obesity and Physical Activity
Macronutrient composition of the diet could help to develop obesity through epigenetic mechanisms.
Epigenetic mechanisms may be implicated in mediating the effects of physical activity.
Tobacco smoke
Tabacco smoke effects have been examined through different epigenetic studies, but the results are still under debate.
Smoking during pregnancy has been associated with increased risk for developing diseases in fetal or later life, through epigenetic mechanisms.
Alcohol consumption
Alcohol is an antagonist of folate metabolism and may have effects on DNA methylation.
Environmental pollutants
Arsenic
Hypo/hypermethylation was observed in blood DNA of subjects exposed to toxic level of arsenic.
Air pollution
Particulate air pollution may affect human health through DNA methylation alterations.
Aromatic hydrocarbons and other organic compounds
Repetitive element hypomethylation as well as either hyper- or hypomethylation of specific genes has been reported for benzene and PAH exposures.
Psychological stress
DNA methylation is sensitive to environmental stressful exposures early in development and later in life.
Shiftwork
An epigenetic reprogramming of circadian genes, changes in Alu repetitive elements methylation and gene-specific methylation of IFN-γ and TNF-α promoters have been observed.
Conclusion and Future Perspectives
Footnotes
Financial & competing interests disclosure
This work was funded by New Investigator funding from the HSPH-NIEHS Center for Environmental Health (ES000002), and Lombardy Region Founding (E.S.S.I.A.: Effetti sulla salute degli inquinanti aerodispersi in regione Lombardia).
The authors have no other relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the manuscript.
Contributor Information
Jorge Alejandro Alegría-Torres, Email: giorgio_alegretto@hotmail.com.
Andrea Baccarelli, Email: abaccare@hsph.harvard.edu.
Valentina Bollati, Email: valentina.bollati@unimi.it.
Bibliography
Papers of special note have been highlighted as:
*of interest
**of considerable interest
- 1.Apa: Thesaurus of Psychological Index Terms.
- 2.Probst Av, Dunleavy E, Almouzni G. Epigenetic inheritance during the cell cycle. Nat Rev Mol Cell Biol. 2009;10(3):192–206. doi: 10.1038/nrm2640. [DOI] [PubMed] [Google Scholar]
- 3.Santos-Reboucas Cb, Pimentel Mm. Implication of abnormal epigenetic patterns for human diseases. Eur J Hum Genet. 2007;15(1):10–17. doi: 10.1038/sj.ejhg.5201727. [DOI] [PubMed] [Google Scholar]
- 4.Bartsch H, Nair J. Oxidative stress and lipid peroxidation-derived DNA-lesions in inflammation driven carcinogenesis. Cancer Detect Prev. 2004;28(6):385–391. doi: 10.1016/j.cdp.2004.07.004. [DOI] [PubMed] [Google Scholar]
- 5.Lawless Mw, O’byrne Kj, Gray Sg. Oxidative stress induced lung cancer and COPD: opportunities for epigenetic therapy. J Cell Mol Med. 2009;13(9A):2800–2821. doi: 10.1111/j.1582-4934.2009.00845.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Arsova-Sarafinovska Z, Eken A, Matevska N, et al. Increased oxidative/nitrosative stress and decreased antioxidant enzyme activities in prostate cancer. Clin Biochem. 2009;42(12):1228–1235. doi: 10.1016/j.clinbiochem.2009.05.009. [DOI] [PubMed] [Google Scholar]
- 7.Kiec-Wilk B, Razny U, Mathers JC, Dembinska-Kiec A. DNA methylation, induced by beta-carotene and arachidonic acid, plays a regulatory role in the pro-angiogenic VEGF-receptor (KDR) gene expression in endothelial cells. J Physiol Pharmacol. 2009;60(4):49–53. [PubMed] [Google Scholar]
- 8.Nowak J, Weylandt KH, Habbel P, Wang J, Dignass A, Glickman JN, Kang JX. Colitis associated colon tumorigenesis is suppressed in transgenic mice rich in endogenous n-3 fatty acids. Carcinogenesis. 2007;28(9):1991–1995. doi: 10.1093/carcin/bgm166. [DOI] [PubMed] [Google Scholar]
- 9.Borek C. Dietary antioxidants and human cancer. Integr Cancer Ther. 2004;3(4):333–341. doi: 10.1177/1534735404270578. [DOI] [PubMed] [Google Scholar]
- 10**.Chen J, Xu X. Diet, epigenetic, and cancer prevention. Adv Genet. 2010;71:237–255. doi: 10.1016/B978-0-12-380864-6.00008-0. [DOI] [PubMed] [Google Scholar]
- 11.Dashwood Rh, Ho E. Dietary histone deacetylase inhibitors: from cells to mice to man. Semin Cancer Biol. 2007;17(5):363–369. doi: 10.1016/j.semcancer.2007.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Druesne N, Pagniez A, Mayeur C, et al. Diallyl disulfide (DADS) increases histone acetylation and p21(waf1/cip1) expression in human colon tumor cell lines. Carcinogenesis. 2004;25(7):1227–1236. doi: 10.1093/carcin/bgh123. [DOI] [PubMed] [Google Scholar]
- 13.Zingg Jm, Jones Pa. Genetic and epigenetic aspects of DNA methylation on genome expression, evolution, mutation and carcinogenesis. Carcinogenesis. 1997;18(5):869–882. doi: 10.1093/carcin/18.5.869. [DOI] [PubMed] [Google Scholar]
- 14.Gonzalez S, Huerta Jm, Alvarez-Uria J, Fernandez S, Patterson Am, Lasheras C. Serum selenium is associated with plasma homocysteine concentrations in elderly humans. J Nutr. 2004;134(7):1736–1740. doi: 10.1093/jn/134.7.1736. [DOI] [PubMed] [Google Scholar]
- 15.Johnson It, Belshaw Nj. Environment, diet and CpG island methylation: epigenetic signals in gastrointestinal neoplasia. Food Chem Toxicol. 2008;46(4):1346–1359. doi: 10.1016/j.fct.2007.09.101. [DOI] [PubMed] [Google Scholar]
- 16.Kim Jm, Hong K, Lee Jh, Lee S, Chang N. Effect of folate deficiency on placental DNA methylation in hyperhomocysteinemic rats. J Nutr Biochem. 2009;20(3):172–176. doi: 10.1016/j.jnutbio.2008.01.010. [DOI] [PubMed] [Google Scholar]
- 17.Giovannucci E. Epidemiologic studies of folate and colorectal neoplasia: a review. J Nutr. 2002;132(8 Suppl):2350S–2355S. doi: 10.1093/jn/132.8.2350S. [DOI] [PubMed] [Google Scholar]
- 18.Jacob Ra, Gretz Dm, Taylor Pc, et al. Moderate folate depletion increases plasma homocysteine and decreases lymphocyte DNA methylation in postmenopausal women. J Nutr. 1998;128(7):1204–1212. doi: 10.1093/jn/128.7.1204. [DOI] [PubMed] [Google Scholar]
- 19.Rampersaud Gc, Kauwell Gp, Hutson Ad, Cerda Jj, Bailey Lb. Genomic DNA methylation decreases in response to moderate folate depletion in elderly women. Am J Clin Nutr. 2000;72(4):998–1003. doi: 10.1093/ajcn/72.4.998. [DOI] [PubMed] [Google Scholar]
- 20.Shin W, Yan J, Abratte Cm, Vermeylen F, Caudill Ma. Choline intake exceeding current dietary recommendations preserves markers of cellular methylation in a genetic subgroup of folate-compromised men. J Nutr. 2010;140(5):975–980. doi: 10.3945/jn.110.121186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21*.Ross Sa, Dwyer J, Umar A, et al. Introduction: diet, epigenetic events and cancer prevention. Nutr Rev. 2008;66(Suppl 1):S1–6. doi: 10.1111/j.1753-4887.2008.00055.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Bravi F, Polesel J, Bosetti C, et al. Dietary intake of selected micronutrients and the risk of pancreatic cancer: an Italian case-control study. Ann Oncol. 2011;22(1):202–206. doi: 10.1093/annonc/mdq302. [DOI] [PubMed] [Google Scholar]
- 23.Gonzalez Ca, Travier N, Lujan-Barroso L, et al. Dietary factors and in situ and invasive cervical cancer risk in the European prospective investigation into cancer and nutrition study. Int J Cancer. 2010 doi: 10.1002/ijc.25679. [DOI] [PubMed] [Google Scholar]
- 24.Shanmugham Jr, Zavras Ai, Rosner Ba, Giovannucci El. Alcohol-folate interactions in the risk of oral cancer in women: a prospective cohort study. Cancer Epidemiol Biomarkers Prev. 2010;19(10):2516–2524. doi: 10.1158/1055-9965.EPI-10-0499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Shitara K, Muro K, Ito S, et al. Folate intake along with genetic polymorphisms in methylenetetrahydrofolate reductase and thymidylate synthase in patients with advanced gastric cancer. Cancer Epidemiol Biomarkers Prev. 2010;19(5):1311–1319. doi: 10.1158/1055-9965.EPI-09-1257. [DOI] [PubMed] [Google Scholar]
- 26.Kim Yi. Folate: a magic bullet or a double edged sword for colorectal cancer prevention? Gut. 2006;55(10):1387–1389. doi: 10.1136/gut.2006.095463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Fini L, Selgrad M, Fogliano V, et al. Annurca apple polyphenols have potent demethylating activity and can reactivate silenced tumor suppressor genes in colorectal cancer cells. J Nutr. 2007;137(12):2622–2628. doi: 10.1093/jn/137.12.2622. [DOI] [PubMed] [Google Scholar]
- 28.Link A, Balaguer F, Goel A. Cancer chemoprevention by dietary polyphenols: promising role for epigenetics. Biochem Pharmacol. 2010;80(12):1771–1792. doi: 10.1016/j.bcp.2010.06.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Paluszczak J, Krajka-Kuzniak V, Malecka Z, et al. Frequent gene hypermethylation in laryngeal cancer cell lines and the resistance to demethylation induction by plant polyphenols. Toxicol In Vitro. 2011;25(1):213–221. doi: 10.1016/j.tiv.2010.11.003. [DOI] [PubMed] [Google Scholar]
- 30.Yuasa Y, Nagasaki H, Akiyama Y, et al. DNA methylation status is inversely correlated with green tea intake and physical activity in gastric cancer patients. Int J Cancer. 2009;124(11):2677–2682. doi: 10.1002/ijc.24231. [DOI] [PubMed] [Google Scholar]
- 31.Yuasa Y, Nagasaki H, Akiyama Y, et al. Relationship between CDX2 gene methylation and dietary factors in gastric cancer patients. Carcinogenesis. 2005;26(1):193–200. doi: 10.1093/carcin/bgh304. [DOI] [PubMed] [Google Scholar]
- 32.Fang Mz, Wang Y, Ai N, et al. Tea polyphenol (−)-epigallocatechin-3-gallate inhibits DNA methyltransferase and reactivates methylation-silenced genes in cancer cell lines. Cancer Res. 2003;63(22):7563–7570. [PubMed] [Google Scholar]
- 33.Lee Wj, Shim Jy, Zhu Bt. Mechanisms for the inhibition of DNA methyltransferases by tea catechins and bioflavonoids. Mol Pharmacol. 2005;68(4):1018–1030. doi: 10.1124/mol.104.008367. [DOI] [PubMed] [Google Scholar]
- 34.Fang M, Chen D, Yang Cs. Dietary polyphenols may affect DNA methylation. J Nutr. 2007;137(1 Suppl):223S–228S. doi: 10.1093/jn/137.1.223S. [DOI] [PubMed] [Google Scholar]
- 35.Lambert Jd, Lee Mj, Diamond L, et al. Dose-dependent levels of epigallocatechin-3-gallate in human colon cancer cells and mouse plasma and tissues. Drug Metab Dispos. 2006;34(1):8–11. doi: 10.1124/dmd.104.003434. [DOI] [PubMed] [Google Scholar]
- 36.Adlercreutz H, Mazur W. Phyto-oestrogens and Western diseases. Ann Med. 1997;29(2):95–120. doi: 10.3109/07853899709113696. [DOI] [PubMed] [Google Scholar]
- 37.Qin W, Zhu W, Shi H, et al. Soy isoflavones have an antiestrogenic effect and alter mammary promoter hypermethylation in healthy premenopausal women. Nutr Cancer. 2009;61(2):238–244. doi: 10.1080/01635580802404196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Li Y, Liu L, Andrews Lg, Tollefsbol To. Genistein depletes telomerase activity through cross-talk between genetic and epigenetic mechanisms. Int J Cancer. 2009;125(2):286–296. doi: 10.1002/ijc.24398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Cross Hs, Kallay E, Lechner D, Gerdenitsch W, Adlercreutz H, Armbrecht Hj. Phytoestrogens and vitamin D metabolism: a new concept for the prevention and therapy of colorectal, prostate, and mammary carcinomas. J Nutr. 2004;134(5):1207S–1212S. doi: 10.1093/jn/134.5.1207S. [DOI] [PubMed] [Google Scholar]
- 40.Xiang N, Zhao R, Song G, Zhong W. Selenite reactivates silenced genes by modifying DNA methylation and histones in prostate cancer cells. Carcinogenesis. 2008;29(11):2175–2181. doi: 10.1093/carcin/bgn179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Davis Cd, Milner J. Frontiers in nutrigenomics, proteomics, metabolomics and cancer prevention. Mutat Res. 2004;551(1–2):51–64. doi: 10.1016/j.mrfmmm.2004.01.012. [DOI] [PubMed] [Google Scholar]
- 42*.Huang S. Histone methyltransferases, diet nutrients and tumour suppressors. Nat Rev Cancer. 2002;2(6):469–476. doi: 10.1038/nrc819. [DOI] [PubMed] [Google Scholar]
- 43*.Davis Cd, Uthus Eo. DNA methylation, cancer susceptibility, and nutrient interactions. Exp Biol Med (Maywood) 2004;229(10):988–995. doi: 10.1177/153537020422901002. [DOI] [PubMed] [Google Scholar]
- 44.Cox R, Goorha S. A study of the mechanism of selenite-induced hypomethylated DNA and differentiation of Friend erythroleukemic cells. Carcinogenesis. 1986;7(12):2015–2018. doi: 10.1093/carcin/7.12.2015. [DOI] [PubMed] [Google Scholar]
- 45.Fiala Es, Staretz Me, Pandya Ga, El-Bayoumy K, Hamilton Sr. Inhibition of DNA cytosine methyltransferase by chemopreventive selenium compounds, determined by an improved assay for DNA cytosine methyltransferase and DNA cytosine methylation. Carcinogenesis. 1998;19(4):597–604. doi: 10.1093/carcin/19.4.597. [DOI] [PubMed] [Google Scholar]
- 46.Davis Cd, Uthus Eo. Dietary selenite and azadeoxycytidine treatments affect di-methylhydrazine-induced aberrant crypt formation in rat colon and DNA methylation in HT-29 cells. J Nutr. 2002;132(2):292–297. doi: 10.1093/jn/132.2.292. [DOI] [PubMed] [Google Scholar]
- 47.Davis Cd, Uthus Eo, Finley Jw. Dietary selenium and arsenic affect DNA methylation in vitro in Caco-2 cells and in vivo in rat liver and colon. J Nutr. 2000;130(12):2903–2909. doi: 10.1093/jn/130.12.2903. [DOI] [PubMed] [Google Scholar]
- 48.Gastaldelli A, Basta G. Ectopic fat and cardiovascular disease: what is the link? Nutr Metab Cardiovasc Dis. 2010;20(7):481–490. doi: 10.1016/j.numecd.2010.05.005. [DOI] [PubMed] [Google Scholar]
- 49.Allender S, Rayner M. The burden of overweight and obesity-related ill health in the UK. Obes Rev. 2007;8(5):467–473. doi: 10.1111/j.1467-789X.2007.00394.x. [DOI] [PubMed] [Google Scholar]
- 50.Klein S, Allison Db, Heymsfield Sb, et al. Waist Circumference and Cardiometabolic Risk: a Consensus Statement from Shaping America’s Health: Association for Weight Management and Obesity Prevention; NAASO, the Obesity Society; the American Society for Nutrition; and the American Diabetes Association. Obesity (Silver Spring) 2007;15(5):1061–1067. doi: 10.1038/oby.2007.632. [DOI] [PubMed] [Google Scholar]
- 51.Lomba A, Milagro Fi, Garcia-Diaz Df, Marti A, Campion J, Martinez Ja. Obesity induced by a pair-fed high fat sucrose diet: methylation and expression pattern of genes related to energy homeostasis. Lipids Health Dis. 2010;9:60. doi: 10.1186/1476-511X-9-60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Campion J, Milagro Fi, Martinez Ja. Individuality and epigenetics in obesity. Obes Rev. 2009;10(4):383–392. doi: 10.1111/j.1467-789X.2009.00595.x. [DOI] [PubMed] [Google Scholar]
- 53.Zhang Ff, Cardarelli R, Carroll J, et al. Physical activity and global genomic DNA methylation in a cancer-free population. Epigenetics. 2011;6(3) doi: 10.4161/epi.6.3.14378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54**.Schulz Wa, Steinhoff C, Florl Ar. Methylation of endogenous human retroelements in health and disease. Curr Top Microbiol Immunol. 2006;310:211–250. doi: 10.1007/3-540-31181-5_11. [DOI] [PubMed] [Google Scholar]
- 55.Baccarelli A, Wright R, Bollati V, et al. Ischemic heart disease and stroke in relation to blood DNA methylation. Epidemiology. 2010;21(6):819–828. doi: 10.1097/EDE.0b013e3181f20457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Mcgee Sl, Fairlie E, Garnham Ap, Hargreaves M. Exercise-induced histone modifications in human skeletal muscle. J Physiol. 2009;587(Pt 24):5951–5958. doi: 10.1113/jphysiol.2009.181065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Radom-Aizik S, Zaldivar F, Jr, Oliver S, Galassetti P, Cooper Dm. Evidence for microRNA involvement in exercise-associated neutrophil gene expression changes. J Appl Physiol. 2010;109(1):252–261. doi: 10.1152/japplphysiol.01291.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Tommasi S, Kim Si, Zhong X, Wu X, Pfeifer Gp, Besaratinia A. Investigating the epigenetic effects of a prototype smoke-derived carcinogen in human cells. PLoS One. 2010;5(5):e10594. doi: 10.1371/journal.pone.0010594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Marwick Ja, Kirkham Pa, Stevenson Cs, et al. Cigarette smoke alters chromatin remodeling and induces proinflammatory genes in rat lungs. Am J Respir Cell Mol Biol. 2004;31(6):633–642. doi: 10.1165/rcmb.2004-0006OC. [DOI] [PubMed] [Google Scholar]
- 60.Van Den Broeck A, Brambilla E, Moro-Sibilot D, et al. Loss of histone H4K20 trimethylation occurs in preneoplasia and influences prognosis of non-small cell lung cancer. Clin Cancer Res. 2008;14(22):7237–7245. doi: 10.1158/1078-0432.CCR-08-0869. [DOI] [PubMed] [Google Scholar]
- 61.Toyooka S, Tokumo M, Shigematsu H, Matsuo K, Asano H, Tomii K, et al. Mutational and epigenetic evidence for independent pathways for lung adenocarcinomas arising in smokers and never smokers. Cancer Res. 2006;66 (3):1371–1375. doi: 10.1158/0008-5472.CAN-05-2625. [DOI] [PubMed] [Google Scholar]
- 62.Liu F, Killian Jk, Yang M, et al. Epigenomic alterations and gene expression profiles in respiratory epithelia exposed to cigarette smoke condensate. Oncogene. 2010;29(25):3650–3664. doi: 10.1038/onc.2010.129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Woodson K, Mason J, Choi Sw, et al. Hypomethylation of p53 in peripheral blood DNA is associated with the development of lung cancer. Cancer Epidemiol Bio-markers Prev. 2001;10(1):69–74. [PubMed] [Google Scholar]
- 64.Pogribny Ip, Basnakian Ag, Miller Bj, Lopatina Ng, Poirier La, James Sj. Breaks in genomic DNA and within the p53 gene are associated with hypomethylation in livers of folate/methyl-deficient rats. Cancer Res. 1995;55(9):1894–1901. [PubMed] [Google Scholar]
- 65.Kim Yi, Pogribny Ip, Basnakian Ag, et al. Folate deficiency in rats induces DNA strand breaks and hypomethylation within the p53 tumor suppressor gene. Am J Clin Nutr. 1997;65(1):46–52. doi: 10.1093/ajcn/65.1.46. [DOI] [PubMed] [Google Scholar]
- 66.Breton Cv, Byun Hm, Wenten M, Pan F, Yang A, Gilliland Fd. Prenatal tobacco smoke exposure affects global and gene-specific DNA methylation. Am J Respir Crit Care Med. 2009;180(5):462–467. doi: 10.1164/rccm.200901-0135OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Pineles Bl, Romero R, Montenegro D, et al. Distinct subsets of microRNAs are expressed differentially in the human placentas of patients with preeclampsia. Am J Obstet Gynecol. 2007;196(3):261, e261–266. doi: 10.1016/j.ajog.2007.01.008. [DOI] [PubMed] [Google Scholar]
- 68.Maccani Ma, Avissar-Whiting M, Banister Ce, Mcgonnigal B, Padbury Jf, Marsit Cj. Maternal cigarette smoking during pregnancy is associated with downregulation of miR-16, miR-21 and miR-146a in the placenta. Epigenetics. 2010;5(7) doi: 10.4161/epi.5.7.12762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Izzotti A, Larghero P, Longobardi M, et al. Dose-responsiveness and persistence of microRNA expression alterations induced by cigarette smoke in mouse lung. Mutat Res. 2010 doi: 10.1016/j.mrfmmm.2010.12.008. [DOI] [PubMed] [Google Scholar]
- 70**.Irigaray P, Newby Ja, Clapp R, et al. Lifestyle-related factors and environmental agents causing cancer: an overview. Biomed Pharmacother. 2007;61(10):640–658. doi: 10.1016/j.biopha.2007.10.006. [DOI] [PubMed] [Google Scholar]
- 71.Van Engeland M, Weijenberg Mp, Roemen Gm, et al. Effects of dietary folate and alcohol intake on promoter methylation in sporadic colorectal cancer: the Netherlands cohort study on diet and cancer. Cancer Res. 2003;63(12):3133–3137. [PubMed] [Google Scholar]
- 72.De Vogel S, Bongaerts Bw, Wouters Ka, et al. Associations of dietary methyl donor intake with MLH1 promoter hypermethylation and related molecular phenotypes in sporadic colorectal cancer. Carcinogenesis. 2008;29(9):1765–1773. doi: 10.1093/carcin/bgn074. [DOI] [PubMed] [Google Scholar]
- 73.Hou L, Wang H, Sartori S, et al. Blood leukocyte DNA hypomethylation and gastric cancer risk in a high-risk Polish population. Int J Cancer. 2010;127(8):1866–1874. doi: 10.1002/ijc.25190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Marutha Ravindran CR, Ticku MK. Changes in methylation pattern of NMDA receptor NR2B gene in cortical neurons after chronic ethanol treatment in mice. Brain Res Mol Brain Res. 2004;121(1–2):19–27. doi: 10.1016/j.molbrainres.2003.10.025. [DOI] [PubMed] [Google Scholar]
- 75.Marutha Ravindran CR, Ticku MK. Role of CpG islands in the up-regulation of NMDA receptor NR2B gene expression following chronic ethanol treatment of cultured cortical neurons of mice. Neurochem Int. 2005;46(4):313–327. doi: 10.1016/j.neuint.2004.10.004. [DOI] [PubMed] [Google Scholar]
- 76.Hicks SD, Middleton FA, Miller MW. Ethanol-induced methylation of cell cycle genes in neural stem cells. J Neurochem. 2010;114(6):1767–1780. doi: 10.1111/j.1471-4159.2010.06886.x. [DOI] [PubMed] [Google Scholar]
- 77.Liu Y, Balaraman Y, Wang G, Nephew KP, Zhou FC. Alcohol exposure alters DNA methylation profiles in mouse embryos at early neurulation. Epigenetics. 2009;4(7):500–511. doi: 10.4161/epi.4.7.9925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Zhou FC, Zhao Q, Liu Y, Goodlett CR, Liang T, McClintick JN, Edenberg HJ, Li L. Alteration of gene expression by alcohol exposure at early neurulation. BMC Genomics. 2001;12:124–140. doi: 10.1186/1471-2164-12-124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Bollati V, Baccarelli A. Environmental epigenetics. Heredity. 2010;105(1):105–112. doi: 10.1038/hdy.2010.2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Baccarelli A, Bollati V. Epigenetics and environmental chemicals. Curr Opin Pediatr. 2009;21(2):243–251. doi: 10.1097/mop.0b013e32832925cc. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Chanda S, Dasgupta Ub, Guhamazumder D, et al. DNA hypermethylation of promoter of gene p53 and p16 in arsenic-exposed people with and without malignancy. Toxicol Sci. 2006;89(2):431–437. doi: 10.1093/toxsci/kfj030. [DOI] [PubMed] [Google Scholar]
- 82.Pilsner Jr, Liu X, Ahsan H, et al. Genomic methylation of peripheral blood leukocyte DNA: influences of arsenic and folate in Bangladeshi adults. Am J Clin Nutr. 2007;86(4):1179–1186. doi: 10.1093/ajcn/86.4.1179. [DOI] [PubMed] [Google Scholar]
- 83.Pilsner Jr, Liu X, Ahsan H, et al. Folate deficiency, hyperhomocysteinemia, low urinary creatinine, and hypomethylation of leukocyte DNA are risk factors for arsenic-induced skin lesions. Environ Health Perspect. 2009;117(2):254–260. doi: 10.1289/ehp.11872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Baccarelli A, Cassano Pa, Litonjua A, et al. Cardiac autonomic dysfunction: effects from particulate air pollution and protection by dietary methyl nutrients and metabolic polymorphisms. Circulation. 2008;117(14):1802–1809. doi: 10.1161/CIRCULATIONAHA.107.726067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Brook Rd, Franklin B, Cascio W, et al. Air pollution and cardiovascular disease: a statement for healthcare professionals from the Expert Panel on Population and Prevention Science of the American Heart Association. Circulation. 2004;109(21):2655–2671. doi: 10.1161/01.CIR.0000128587.30041.C8. [DOI] [PubMed] [Google Scholar]
- 86.Peters A. Particulate matter and heart disease: evidence from epidemiological studies. Toxicol Appl Pharmacol. 2005;207(2 Suppl):477–482. doi: 10.1016/j.taap.2005.04.030. [DOI] [PubMed] [Google Scholar]
- 87.Samet Jm, Dominici F, Curriero Fc, Coursac I, Zeger Sl. Fine particulate air pollution and mortality in 20 U.S. cities, 1987–1994. N Engl J Med. 2000;343(24):1742–1749. doi: 10.1056/NEJM200012143432401. [DOI] [PubMed] [Google Scholar]
- 88.Vineis P, Husgafvel-Pursiainen K. Air pollution and cancer: biomarker studies in human populations. Carcinogenesis. 2005;26(11):1846–1855. doi: 10.1093/carcin/bgi216. [DOI] [PubMed] [Google Scholar]
- 89.Tarantini L, Bonzini M, Apostoli P, et al. Effects of particulate matter on genomic DNA methylation content and iNOS promoter methylation. Environ Health Perspect. 2009;117(2):217–222. doi: 10.1289/ehp.11898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Alexeeff Se, Litonjua Aa, Wright Ro, et al. Ozone exposure, antioxidant genes, and lung function in an elderly cohort: VA normative aging study. Occup Environ Med. 2008;65(11):736–742. doi: 10.1136/oem.2007.035253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Baccarelli A, Zanobetti A, Martinelli I, et al. Air pollution, smoking, and plasma homocysteine. Environ Health Perspect. 2007;115(2):176–181. doi: 10.1289/ehp.9517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Chahine T, Baccarelli A, Litonjua A, et al. Particulate air pollution, oxidative stress genes, and heart rate variability in an elderly cohort. Environ Health Perspect. 2007;115(11):1617–1622. doi: 10.1289/ehp.10318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93*.Ehrlich M. DNA hypomethylation in cancer cells. Epigenomics. 2009;1(2):239–259. doi: 10.2217/epi.09.33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94*.Castro R, Rivera I, Struys Ea, et al. Increased homocysteine and S-adenosylhomocysteine concentrations and DNA hypomethylation in vascular disease. Clin Chem. 2003;49(8):1292–1296. doi: 10.1373/49.8.1292. [DOI] [PubMed] [Google Scholar]
- 95.Bollati V, Marinelli B, Apostoli P, et al. Exposure to metal-rich particulate matter modifies the expression of candidate microRNAs in peripheral blood leukocytes. Environ Health Perspect. 2010;118(6):763–768. doi: 10.1289/ehp.0901300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Snyder R. Benzene and leukemia. Crit Rev Toxicol. 2002;32(3):155–210. doi: 10.1080/20024091064219. [DOI] [PubMed] [Google Scholar]
- 97.Bollati V, Baccarelli A, Hou L, et al. Changes in DNA methylation patterns in subjects exposed to low-dose benzene. Cancer Res. 2007;67(3):876–880. doi: 10.1158/0008-5472.CAN-06-2995. [DOI] [PubMed] [Google Scholar]
- 98.Costantini As, Benvenuti A, Vineis P, et al. Risk of leukemia and multiple myeloma associated with exposure to benzene and other organic solvents: evidence from the Italian Multicenter Case-control study. Am J Ind Med. 2008;51(11):803–811. doi: 10.1002/ajim.20592. [DOI] [PubMed] [Google Scholar]
- 99.Kirkeleit J, Riise T, Bratveit M, Moen Be. Increased risk of acute myelogenous leukemia and multiple myeloma in a historical cohort of upstream petroleum workers exposed to crude oil. Cancer Causes Control. 2008;19(1):13–23. doi: 10.1007/s10552-007-9065-x. [DOI] [PubMed] [Google Scholar]
- 100.Ji Z, Zhang L, Peng V, Ren X, Mchale Cm, Smith Mt. A comparison of the cytogenetic alterations and global DNA hypomethylation induced by the benzene metabolite, hydroquinone, with those induced by melphalan and etoposide. Leukemia. 2010;24(5):986–991. doi: 10.1038/leu.2010.43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Pavanello S, Bollati V, Pesatori Ac, et al. Global and gene-specific promoter methylation changes are related to anti-B[a]PDE-DNA adduct levels and influence micronuclei levels in polycyclic aromatic hydrocarbon-exposed individuals. Int J Cancer. 2009;125(7):1692–1697. doi: 10.1002/ijc.24492. [DOI] [PubMed] [Google Scholar]
- 102.Perera F, Tang Wy, Herbstman J, et al. Relation of DNA methylation of 5′-CpG island of ACSL3 to transplacental exposure to airborne polycyclic aromatic hydrocarbons and childhood asthma. PLoS One. 2009;4(2):e4488. doi: 10.1371/journal.pone.0004488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Rusiecki Ja, Baccarelli A, Bollati V, Tarantini L, Moore Le, Bonefeld-Jorgensen Ec. Global DNA hypomethylation is associated with high serum-persistent organic pollutants in Greenlandic Inuit. Environ Health Perspect. 2008;116(11):1547–1552. doi: 10.1289/ehp.11338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104**.Jirtle Rl, Skinner Mk. Environmental epigenomics and disease susceptibility. Nat Rev Genet. 2007;8(4):253–262. doi: 10.1038/nrg2045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Miller Ca, Sweatt Jd. Covalent modification of DNA regulates memory formation. Neuron. 2007;53(6):857–869. doi: 10.1016/j.neuron.2007.02.022. [DOI] [PubMed] [Google Scholar]
- 106**.Szyf M, Mcgowan P, Meaney Mj. The social environment and the epigenome. Environ Mol Mutagen. 2008;49(1):46–60. doi: 10.1002/em.20357. [DOI] [PubMed] [Google Scholar]
- 107.Miller Ca, Gavin Cf, White Ja, et al. Cortical DNA methylation maintains remote memory. Nat Neurosci. 2010;13(6):664–666. doi: 10.1038/nn.2560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Murgatroyd C, Patchev Av, Wu Y, et al. Dynamic DNA methylation programs persistent adverse effects of early-life stress. Nat Neurosci. 2009;12(12):1559–1566. doi: 10.1038/nn.2436. [DOI] [PubMed] [Google Scholar]
- 109.Mcgowan Po, Sasaki A, D’alessio Ac, et al. Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse. Nat Neurosci. 2009;12(3):342–348. doi: 10.1038/nn.2270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110*.Weaver Ic. Epigenetic programming by maternal behavior and pharmacological intervention. Nature versus nurture: let’s call the whole thing off. Epigenetics. 2007;2(1):22–28. doi: 10.4161/epi.2.1.3881. [DOI] [PubMed] [Google Scholar]
- 111.Weaver Ic, Cervoni N, Champagne Fa, et al. Epigenetic programming by maternal behavior. Nat Neurosci. 2004;7(8):847–854. doi: 10.1038/nn1276. [DOI] [PubMed] [Google Scholar]
- 112.Nakahata Y, Grimaldi B, Sahar S, Hirayama J, Sassone-Corsi P. Signaling to the circadian clock: plasticity by chromatin remodeling. Curr Opin Cell Biol. 2007;19 (2):230–237. doi: 10.1016/j.ceb.2007.02.016. [DOI] [PubMed] [Google Scholar]
- 113.Hirayama J, Sahar S, Grimaldi B, Tamaru T, Takamatsu K, Nakahata Y, Sasson Corsi P. CLOCK-mediated acetylation of BMAL1 controls circadian function. Nature. 2007;450 (7172):1086–1090. doi: 10.1038/nature06394. [DOI] [PubMed] [Google Scholar]
- 114.Grimaldi B, Nakahata Y, Kaluzova M, Masubuchi S, Sassone-Corsi P. Chromatin remodeling, metabolism and circadian clocks: the interplay of CLOCK and SIRT1. Int J Biochem Cell Biol. 2009;41(1):81–86. doi: 10.1016/j.biocel.2008.08.035. [DOI] [PubMed] [Google Scholar]
- 115.Costa G. The problem: shiftwork. Chronobiol Int. 1997;14(2):89–98. doi: 10.3109/07420529709001147. [DOI] [PubMed] [Google Scholar]
- 116.Zhu Y, Zheng T, Stevens Rg, Zhang Y, Boyle P. Does “clock” matter in prostate cancer? Cancer Epidemiol Biomarkers Prev. 2006;15(1):3–5. doi: 10.1158/1055-9965.EPI-05-0631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Sahar S, Sassone-Corsi P. Circadian clock and breast cancer: a molecular link. Cell Cycle. 2007;6(11):1329–1331. doi: 10.4161/cc.6.11.4295. [DOI] [PubMed] [Google Scholar]
- 118.Bollati V, Baccarelli A, Sartori S, et al. Epigenetic effects of shiftwork on blood DNA methylation. Chronobiol Int. 2010;27(5):1093–1104. doi: 10.3109/07420528.2010.490065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Morgan DK, Whitelaw E. The case for transgenerational epigenetic inheritance in humans. Mamm Genome. 2008;19(6):394–397. doi: 10.1007/s00335-008-9124-y. [DOI] [PubMed] [Google Scholar]
- 120.Chao MJ, Ramagopalan SV, Herrera BM, Lincoln MR, Dyment DA, Sadovnick AD, Ebers GC. Epigenetics in multiple sclerosis susceptibility: difference in transgenerational risk localizes to the major histocompatibility complex. Hum Mol Genet. 2009;18(2):261–266. doi: 10.1093/hmg/ddn353. [DOI] [PubMed] [Google Scholar]
- 121.Daxinger L, Whitelaw E. Transgenerational epigenetic inheritance: more questions than answers. Genome Res. 2010;20(12):1623–1628. doi: 10.1101/gr.106138.110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Nelson VR, Nadeau JH. Transgenerational genetic effects. Epigenomics. 2010;2 (6):797–806. doi: 10.2217/epi.10.57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123*.Skinner MK, Guerrero-Bosagna C. Environmental signals and transgenerational epigenetics. Epigenomics. 2009;1(1):111–117. doi: 10.2217/epi.09.11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Skinner MK, Manikkam M, Guerrero-Bosagna C. Epigenetic transgenerational actions of environmental factors in disease etiology. Trends Endocrinol Metab. 2010;21(4):214–222. doi: 10.1016/j.tem.2009.12.007. [DOI] [PMC free article] [PubMed] [Google Scholar]