

Abstract
Vaccines that activate humoral and cell-mediated immune responses are urgently needed for many infectious agents, including the flaviviruses dengue and West Nile (WN) virus. Vaccine development would be greatly facilitated by a new approach, in which nanoscale modules (Ag, adjuvant, and carrier) are assembled into units that are optimized for stimulating immune responses to a specific pathogen. Toward that goal, we formulated biodegradable nanoparticles loaded with Ag and surface modified with the pathogen-associated molecular pattern CpG oligodeoxynucleotides. We chose to evaluate our construct using a recombinant envelope protein Ag from the WN virus and tested the efficiency of this system in eliciting humoral and cellular responses and providing protection against the live virus. Animals immunized with this system showed robust humoral responses polarized toward Th1 immune responses compared with predominately Th2-biased responses with the adjuvant aluminum hydroxide. Immunization with CpG oligodeoxynucleotide-modified nanoparticles resulted in a greater number of circulating effector T cells and greater activity of Ag-specific lymphocytes than unmodified nanoparticles or aluminum hydroxide. Ultimately, compared with alum, this system offered superior protection in a mouse model of WN virus encephalitis.
The flavivirus genus, which is a group of icosahedral, enveloped, positive, ssRNA viruses, includes many human pathogens of global epidemiological importance (1). Examples include agents involved in dengue fever and West Nile (WN) meningoencephalitis, for which there are no vaccines approved for human use (2). This unmet medical need is recognized by the National Institute of Allergy and Infectious Diseases (National Institutes of Health, Bethesda, MD), which includes WN virus and several other flaviviruses on its list of priority pathogens. Although human infections with WN virus are less prevalent than infections with some other flaviviruses, such as dengue virus, there are several well-established animal models for WN virus disease, allowing a clear path for the preclinical development of potential vaccines. In contrast, there are no good models for dengue fever in rodents, making vaccine development more challenging. However, given similarities seen among all flaviviruses, results obtained with WN virus may be broadly applicable to other flaviviruses.
Researchers have used multiple approaches to develop vaccines against WN and dengue viruses. These include killed virus preparations (3–5), attenuated virus strains (4–8), DNA vaccines (9, 10), recombinant canarypox virus or adenoviruses (11, 12), and recombinant subunit vaccines based on the viral E protein (4, 10, 13, 14). We recently reported the application of poly(D,L,-lactide-coglycolide) (PLGA) nanoparticles with surface-associated LPS as effective delivery vehicles for vaccination against WN virus (15).
It is instructive to note that viruses are nanoscale biological objects that are highly optimized for host infection. Therefore, we reasoned that the principles of nanoassembly can be used to design safe vectors that are highly optimized for protection against disease. Our aim in this work was to design and test a nanoparticulate system incorporating virus-inspired components for WN vaccine delivery. To achieve this, we sought to design a system that facilitates delivery of Ag and immune potentiator molecules to APCs. We chose CpG oligodeoxynucleotide (ODN) 1826 as the immune potentiator, a target for the intracellular receptor TLR9, and we chose the carrier to be a nanoparticulate formulation of a biodegradable polymer with an extensive history of use in humans, PLGA.
CpG ODN potentiation of a number of carriers was shown to be a promising methodology for induction of vaccine responses. For example, CpG ODNs have been incorporated into liposomes (16–19) and biodegradable particles (20–26) to create Ag-delivery vehicles with adjuvant properties. Unmethylated CpG sequences framed by two 3′ pyrimidines and two 5′ purines are recognized by TLR9 (27). Synthetic B-type CpG ODNs, such as ODN 1826, contain CpG-rich motifs on a nuclease-resistant phosphorothioate backbone and were shown to activate B cells, dendritic cells, and macrophages (reviewed in Ref. 28). Given that CpG ODN is quickly cleared by the body and degraded by nucleases (29, 30), particulates are able to protect and enhance the circulation time of encapsulated or adsorbed molecules. Also, particulates are able to colocalize Ag and CpG ODN and assure simultaneous transport of both agents to APCs. Conjugation or proximal delivery of Ag and CpG ODN was shown to be an effective strategy to maximize the benefits of both agents (31, 32). Furthermore, improved responses from Ag and CpG DNA coencapsulated in the same particle, compared with mixing particles encapsulating either agent (16, 17, 23, 33), suggests a necessity for both to reach the same APC.
We chose to create the vaccine-delivery system from PLGA, a biodegradable U.S. Food and Drug Administration-approved polymer. PLGA nano- and microparticles have been explored for use as vaccine-delivery vehicles for nearly 20 y (34). In our previous work, we developed a technology that facilitates enhanced surface presentation of targeting ligands on the surface of biodegradable PLGA (35). We demonstrated that biotinylated ligands bound to the surface of PLGA nanoparticles by an avidin-palmitic acid bridging element remain associated with the particles during encapsulant release (35). Although most research with biodegradable nanoparticles involved the coencapsulation of CpG ODN with Ag (22), our strategy tethers the molecules to the surface to keep the Ag and CpG ODN colocalized and to minimize release of CpG ODN. This is in contrast to CpG ODN adsorbed to cationic PLGA microparticles (20, 25), where up to 80% of all CpG ODN can be released from the surface after 24 h (20).
In this study, we developed CpG ODN-modified nanoparticles encapsulating a truncated version of the WN virus envelope protein, an Ag with proven vaccination efficacy in mice and horses (13). Particles were effectively endocytosed by dendritic cells, resulting in the secretion of proinflammatory cytokines and the upregulation of costimulatory molecules, enabling the activation of Ag-specific lymphocytes. CpG ODN-modified nanoparticles elicited robust Th1-biased humoral responses in immunized mice and a strong cellular response with a high number of circulating effector-like T cells, unlike vaccination with aluminum hydroxide, which favors a Th2 response. Ultimately, this system offered superior protection in a mouse model of WN encephalitis.
Materials and Methods
Materials
Expression of a truncated recombinant WN virus envelope protein (rWNVE) by Drosophila S2 cells was described previously (13). A type B CpG ODN 1826 (5′-TCC ATG ACG TTC CTG ACG TT-3′) with a phosphorotioate backbone was synthesized and conjugated to biotin on site by the Keck Facility (Yale University). Poly(vinyl alcohol) (PVA), sodium deoxycholate, palmitic acid N-hydroxy-succinimide ester, Nile red, Histopaque, OVA, and gelatin were all purchased from Sigma-Aldrich (St. Louis, MO). Methylene chloride and avidin were purchased from Thermo Fisher Scientific (Waltham, MA) and Invitrogen (Carlsbad, CA), respectively. Research-grade PLGA (50:50, ~50,000 g/mol), was purchased from Durect (Pelham, AL).
Nanoparticle fabrication and characterization
Avidin palmitate was prepared by a slightly modified method than described previously (35). Briefly, 5 mg/ml avidin in 2% sodium deoxycholate was reacted to a 15-fold molar excess of palmitic acid N-hydroxy-succinimide ester for 3 h at 37°C. Excess material was removed by dialysis in a 2% sodium deoxycholate bath over 24 h at 37°C.
Nanoparticles were prepared using an endotoxin-free, double-emulsion method. To remove pyrogens, all glassware was baked at 250°C for 2 h. All plastic instruments were decontaminated in Pyro-Clean (Thermo Fisher Scientific) for30min and subsequentlywashedinUltrapurewater (Invitrogen).As needed, purchased materials were screened for endotoxin activity. rWNVE wasmadeinaeukaryotic system, reducing concerns about contamination from endotoxin resulting from productioninabacterial system.Inthefirst emulsion, 1 mg soluble rWNVE (Nile red for optical studies in internalization experiments) in PBS was added dropwise to 100 mg PLGA (~50,000 g/mol) solubilized in 2 ml methylene chloride. In the second emulsion, the first emulsion was added dropwise to 10 mg avidin-palmitic acid in 1.25% PVA and 1% sodium deoxycholate. Both emulsion steps involved brief vortexing followed by 30 s of ultrasonication on ice using a Tekmar Sonic Distributor fitted with a CV26 sonicator (38% amplitude). Methylene chloride was evaporated by vigorously stirringasparticles hardenedin100 ml0.3% PVAinwater for3hat room temperature (RT). Particles were collected bycentrifugation at 18,500 × g and washed three times with deionized water. Next, the particles were flashfrozen, lyophilized, and stored at −20°C prior to use. Immediately before experiments, nanoparticles were suspended in PBS or media, and biotinylated CpG ODN was added at 0.5 μg/mg particles. Our preliminary experiments indicated that, at this concentration, all CpG ODN was attached to particles with no free CpG in solution.
To assay for rWNVE content, particles were dissolved in 0.2 N NaOH overnight, and protein content was assayed with the Micro BCA protein assay (Pierce, Rockford, IL). Optical densities obtained with rWNVE-free nanoparticles were used as negative control values. Controlled release of rWNVE was determined by suspending particles in a microcentrifuge tube with 1.1 ml PBS with 0.05% sodium azide to prevent bacterial growth. Particles were pelleted at various time points by centrifugation at 16,000 × g, and 1 ml supernatant was collected and replaced with 1 ml fresh buffer. Supernatant was frozen at −20°C until assayed for protein content by the Micro BCA assay. Particle size and distribution were measured by a liquidphase Stokes diffusion particle-tracking system (NanoSight; Wiltshire, U.K.) and visualized by scanning electron microscopy.
In vitro assaying of bone marrow-derived dendritic cell function with CpG ODN-modified nanoparticles
Bone marrow-derived dendritic cells (BMDCs) were pulsed with nanoparticles and subsequently analyzed for activation by cytokine secretion and surface marker upregulation. Bone marrow was isolated from the femurs of C57BL/6 mice at 6–8 wk of age and differentiated into dendritic cells in complete media (RPMI 1640 with 10% FBS, L-glutamine, MEM, nonessential amino acids, HEPES buffer, gentamicin, 2-ME) and supplemented with 10 ng/ml rGM-CSF in 24-well plates. Cells were given fresh media every other day and used on day 7.
Internalization of nanoparticles was confirmed by incubation of Nile redloaded, CpG ODN-modified nanoparticles with BMDCs for 1 h in complete mediaat37°Consterile polylysine-coatedcoverslips. Coverslipswerewashed, fixed in 4% paraformaldehyde for 15 min, and permeabilized with Triton-X twice for 5 min. Cells were then incubated with a 1:100 dilution of phalloidin-FITC in PBS for 30 min at RT in the dark. Coverslips were washed, mounted onto slides using hard set Vectashield, and visualized by fluorescent microscopy using a Zeiss Axiovert 200 Inverted Microscope with an MRm camera. As a control, BMDCs were incubated with free Nile red in media.
BMDCs were pulsed with rWNVE adsorbed to aluminum hydroxide gel (Alhydrogel; Accurate, Westbury, NY), unmodified rWNVE-loaded nanoparticles (−/rWNVE), or CpG ODN-modified rWNVE-loaded nanoparticles (CpG ODN/rWNVE) for 24 h at 37°C in complete media. For IL-1β assays, this was preceded by an overnight incubation with 50 ng/ml LPS (Sigma-Aldrich). Supernatant was analyzed by ELISA for the cytokines IL-6, IL-12 (BD Pharmingen, San Diego, CA), and IL-1β (R&D Systems, Minneapolis, MN), accordingtothe manufacturers’ protocols. Surface marker upregulation was detected by flow cytometry. Briefly, cells were washed of particles and then stained for mouse CD11c, CD80, and CD86 (eBioscience, San Diego, CA); fixed with Cytofix (BD Biosciences, San Jose, CA); and analyzed on the LSRII (BD Biosciences).
Animal immunization
Allmiceweremaintainedinapathogen-freefacility andtreatedincompliance with Yale University Institutional Animal Care&Use Committee regulations. Six- to eight-week-old female C3H/HeN mice (Charles River Laboratories, Wilmington, MA) were s.c. administered 10 μg rWNVE encapsulated in nanoparticles, adsorbed to Alhydrogel, or given as a solution with or without soluble CpG ODN in PBS. Vaccine doses containing Alhydrogel were prepared by mixing rWNVE, Alhydrogel (1 μl/μg protein), and 20 mM Tris–HCl 100 mM NaCl solution (pH 8). Mice were boosted with anidentical dose at 2 wk. Blood and spleens were collected at 4 and 6 wk for Ab and cellular-response analysis, respectively.
C57BL/6 mice and mice that were deficient in TLR9 (on a C57BL/6 background) were vaccinated with a single s.c. dose of CpG-modified LPS-modified (Escherichia coli 0111:B4; Sigma-Aldrich) or unmodified nanoparticles encapsulating rWNVE. LPS-modified nanoparticles were prepared as previously described (15). At 4 wk postvaccination, mice were analyzed for circulating Ag-specific Abs.
For survival studies, groups of 6-wk-old C3H/HeN mice were vaccinated s.c. once with 2.5 μg CpG ODN-modified nanoparticle-encapsulated or adsorbed rWNVE. At 2 wk, mice were bled and injected i.p. with 1000 PFU WN virus isolate 2741 in 1% gelatin. Mice were observed for morbidity and mortality for 21 d postchallenge.
Ab titer analysis
Ab isotype titers were measured from the serum of vaccinated mice. Blood samples collected retro-orbitally were allowed to clot at 4°C overnight and centrifuged at 1000 × g for 10 min. Serum was collected and stored at −80°C until assayed. Anti-rWNVE IgG isotypes were measured by ELISA. High-binding plates (Corning Glass, Corning, NY) were coated with 100 ng rWNVE per well overnight in PBS at 4°C. Plates were incubated with blocking buffer (PBS + 5% BSA [Sigma-Aldrich] + 0.1% Tween 20 [Sigma-Aldrich]) for 1 h at RT. Plates were washed three times in wash buffer (PBS + 0.05% Tween 20) and then incubated with serial dilutions of serum in blocking buffer for 1 h at RT. Plates were washed three times and then HRP-conjugated goat anti-mouse IgG1, IgG2a, IgG2b (Invitrogen), and total IgG (Jackson ImmunoResearch Laboratories, West Grove, PA) were added in blocking buffer at a 1:2000 dilution. After 1 h, plates were washed six times, followed by development with tetramethylbenzidine substrate (Kirkegaard & Perry Laboratories, Gaithersburg, MD). Reaction was stopped with 1 N HCl after 15 min. Absorbance was measured by a spectrophotometer at 450 nm. Titer was calculated as the inverse dilution at which the absorbance equaled that of the control (samples from mice receiving no Ag) plus 2 SD. Linear regression of dilution versus Ab curve was calculated after a log transformation of values.
Plaque-reduction neutralization tests
Invitroplaque-reduction neutralization tests(PRNTs)wereusedtoevaluatethe ability ofserum samplesto inhibit virus replication in cultured Vero cells. Sera from immunized mice were tested in a biosafety level 2 facility using an assay that used an attenuated chimeric virus WNV/DEN4Delta30 (kindly provided by Dr. Steven Whitehead, National Institute of Allergy and Infectious Diseases) (8, 36, 37). This method was described previously (38). In brief, Vero cells were maintained in Opti-MEM-Glutamax (Invitrogen) with 5% FBS and 100 μg/ml penicillin-streptomycin and seeded in 24-well plates at 105 cells/well 24 h before infection. Ten-fold serial Opti-MEM dilutions of heat-inactivated serum samples were mixed with virus (100 PFU/100 μl), maintained for ~10 min at RT, and added to the cells in culture for 1 h at 37°C, in 5% CO2. Overlay medium (1% methylcellulose in Opti-MEM–Glutamax with 2% FBS, 50 μg/ml gentamicin and 2.5 μg/ml amphotericin B) was added to the cells, and the plates were incubated at 37°C in 5% CO2 for 5 d. Viral plaque formation was detected by immunostaining using horse anti-rWNVE protein polyclonal Abs (39). Adherent cells were first fixed in 80% methanol, blocked in 5% skim milk in PBS for 10 min at RT, and incubated with 10 μg/ml 4G2 IgG at 37°C in 5% CO2 for 90 min. Cells were washed twicein5% skim milk in PBS and incubated with a 1:2000 dilution of peroxidase-labeled anti-horse Ab conjugate (Kirkegaard & Perry Laboratories) for 60 min at 37°C. Cells were washed twice in PBS and incubated in peroxidase substrate (True-Blue; Kirkegaard & Perry Laboratories) until plaque immunostaining was observed. PRNT 50 is the serum dilution value at which 50% inhibition of plaque formation is obtained.
Assaying for circulating T cell phenotype and ex vivo cellular activity
One week postvaccination, blood was collected retro-orbitally and transferred to sodium citrate buffer. Lymphocytes were isolated using Histopaque 1077, washed, and stained in FACS buffer with fluorescent Abs for CD8, CD44, CD127, and killer cell lectin-like receptor G1 (KLRG1; eBioscience). Cells were treated with Cytofix and analyzed by flow cytometry using the LSRII.
Splenocytes from vaccinated mice were pulsed ex vivo with Ag and assayed for IL-2 and IFN-γ release as an indicator of cell-mediated immunity. At 6 wk postinitial vaccination, splenocytes were isolated by mashing the spleen, lysing RBCs with ACK lysing buffer (Invitrogen), washing in PBS, and then transferring cells to a 96-well plate at 3 × 105 cells per well in complete media with 25 μg/ml rWNVE. After 48 h at 37°C, supernatant was collected and assayed for cytokines by ELISA (BD Pharmingen).
Statistical analysis
All experiments were performed multiple times and analyzed using Graph-Pad Prism Version5software. Data were analyzedby one-way ANOVA or the Student t test. Statistical significance in survival studies was determined by the log rank (Mantel–Cox) test.
Results
Characterization of the nanoparticle system
A schematic of the nanoparticle system is shown in Fig. 1A. rWNVE loading was found to be ~4.7 μg of protein per 1 mg of nanoparticles. A representative release curve of encapsulated rWNVE is shown in Fig. 1B. In an initial phase lasting 1–2 d, ~ 20% of the Ag is released from the nanoparticles. More than 60% of the encapsulant is then released over 5 wk. This biphasic nature and duration of release is typical for PLGA particles in this size range (40, 41).
FIGURE 1.

Nanoparticle characterization. A, PLGA nanoparticles were formulated to encapsulate rWNVE and were surface modified with an avidin-palmitate linker and biotinylated CpG ODN. Nanoparticles were characterized by measuring rWNVE release in triplicate over a 5-wk period (B), determining size distribution using the NanoSight system (C), and visualization by scanning electron microscopy (D). Scale bar, 500 nm. Original magnification × 51,200. E, NR was loaded into nanoparticles to confirm internalization of particlesbyBMDCs, visualized by fluorescent microscopy. Free NR was used as a control. Scale bar, 20 μm. Original magnification × 400. NR, Nile red.
Particles were sized by a novel modality (NanoSight) that tracks particle movement in liquid and derives size distribution and number of particles in a sample based on a conventional Stokes–Einstein equation that relates the Brownian Motion of nanoparticles to the hydrodynamic radius (42). Particles were found to have a mean diameter of 272 nm and a mode of 229 nm (Fig. 1C). Each milligram of material consisted of ~1 × 1010 individual particles as enumerated by this system. Particle morphology was determined using a scanning electron microscope (Fig. 1D). Rather than displaying a smooth surface characteristic of unmodified PLGA particles, modified nanoparticles displayed a surface texture indicative of modification with the avidin-palmitate conjugate.
Dendritic cells were observed to readily endocytose Nile redloaded, CpG ODN-modified nanoparticles 1 h after incubation (Fig. 1E). In comparison, free Nile red molecules alone did not associate with the cells during that time. We did not observe a preferential uptake of CpG ODN-modified nanoparticles compared with unmodified nanoparticles (data not shown), consistent with the fact that the receptor for CpG ODN (TLR9) is an intracellular one and, hence, does not impact surface receptor-mediated internalization kinetics.
Dendritic cells are activated after incubation with CpG ODN-modified nanoparticles
To determine the effect of particles on dendritic cell activation, BMDCs were pulsed with CpG ODN-modified and unmodified rWNVE-loaded nanoparticles, as well as rWNVE adsorbed to Alhydrogel,and were assessed for secreted cytokines and maturation surface markers. After a 24-h incubation, BMDCs were stained with Abs against the costimulatory molecules CD80 and CD86 and analyzed by flow cytometry. CD80 and CD86 expression on CD11c+ cells was elevated after treatment with CpG ODN-modified nanoparticles (Fig. 2). Unmodified nanoparticles and Alhydrogel were less successful at promoting costimulatory marker expression. Dendritic cells secreted higher amounts of the cytokines IL-6 and IL-12 after incubation with CpG-modified, rWNVE-loaded nanoparticles compared with blank nanoparticles; unmodified, rWNVE-loaded nanoparticles; and rWNVE adsorbed to Alhydrogel (Fig. 3A, 3B). IL-12 influences the differentiation of Th0 cells into a Th1 phenotype (43), whereas IL-6 regulates this action (44).
FIGURE 2.

BMDC surface marker expression. BMDCs in triplicate were pulsed with CpG ODN/rWNVE, −/rWNVE, or rWNVE adsorbed to Alhydrogel for 24 h and then stained with fluorescent Abs for CD80 (A) and CD86 (B) and analyzed by flow cytometry. CD11c+ live cells were gated, and the percentage of these cells positive for surface markers was enumerated. *p < 0.01 by ANOVA. Data shown are from a single experiment that was repeated three times with the same result.
FIGURE 3.

BMDC activation. BMDCs in triplicate were pulsed with CpG ODN/rWNVE, −/rWNVE, or rWNVE adsorbed to Alhydrogel for 24 h. Supernatant was analyzed for IL-6 (A) and IL-12 (B) by ELISA. C, Inflammasomeactivity.BMDCswere pretreated with 50 ng/ml of LPS overnight. Cells were then incubated with LPS-modified, CpG-modified, or unmodified rWNVE-loaded nanoparticles; rWNVE adsorbed to Alhydrogel; or rWNVE alone for 24 h. Supernatant was collected and analyzed for IL-1β by ELISA. *p < 0.001 by ANOVA. Data shown are from a single experiment that was repeated three times with the same result.
A separate set ofexperiments was performed to elucidatewhether CpG ODN-modified nanoparticles trigger inflammasome activity. The inflammasome is an intracellular complex of proteins, including a member of the NOD-like receptor family and the apoptosis-associated speck-like adaptor protein, which mediates the activity of caspase-1 and results in the secretion of proinflammatory cytokines, such as IL-1β and IL-18. BMDCs were stimulated overnight with LPS to increase the expression of the proform of IL-1β and then were pulsed with particles for 24 h. To ascertain inflammasome activation, supernatant was analyzed for IL-1β. No IL-1β and, therefore, no inflammasome activity, was detected after pulsing with CpG ODN-modified nanoparticles (Fig. 3C). This is in contrast to aluminum adjuvants (45, 46) or LPS-modified nanoparticles (15), which were shown to activate the NLRP3 inflammasome (Fig. 3C).
Immunization with CpG ODN-modified nanoparticles generated high titers of Th1 Ag-specific Abs
C3H/HeN mice were immunized with encapsulated rWNVE in CpG ODN-modified or unmodified nanoparticles, free rWNVE with and without CpG ODN, or rWNVE adsorbed to Alhydrogel. The amount of nanoparticles administered was calculated such that each animal received 10 μg of rWNVE per dose. Mice were boosted 2 wk later, and serum was analyzed for circulating total anti-rWNVE IgG and IgG isotype titers. Alhydrogel and CpG ODN-modified nanoparticles successfully elicited robust humoral responses against the envelope protein, but the resulting isotype profiles were very different between the adjuvants. Alhydrogel and CpG ODN-modified nanoparticles induced roughly equivalent titers of total IgG (Fig. 4A). Unmodified particles produced a weaker response, whereas soluble rWNVE, with and without CpG ODN, was markedly less effective (Fig. 4A). As expected, Alhydrogel generated the highest levels of IgG1 (associated with a Th2-skewed response) (Fig. 4B). CpG ODN-modified particles resulted in a lower, but substantial, titer of IgG1. Interestingly, only CpG ODN-modified particles resulted in high IgG2a and IgG2b titers (Th1-biased response) (Fig. 4C, 4D). IgG1 and IgG2 titers were dose responsive after immunization with CpG ODN-modified nanoparticles (Supplemental Fig. 1). To assay for the neutralizing ability of circulating Abs, we performed a PRNTwith pooled serum from immunized animals. The neutralizing ability of Abs generated by CpG ODN-modified nanoparticles was slightly higher than that from Alhydrogel, although the difference was not statistically significant (Fig. 4E).
FIGURE 4.

Ag-specific Ab isotype titers after vaccination. Groups of four C3H/HeN mice were immunized twice with 10 μg of rWNVE in different formulations at 0 and 2 wk. Serum was isolated at 4 wk and analyzed for rWNVE-specific titers of total IgG (A), IgG1 (B), IgG2a (C), and IgG2b (D) by ELISA. E, Serum samples from mice given rWNVE encapsulated in nanoparticles or adsorbed to Alhydrogel were pooled into two groups and were analyzed for PRNTs that decreased plaque formation by 50% (PRNT 50). *p < 0.02 by ANOVA. Data shown are from a single experiment that was repeated twice with the same results.
To validate the role of TLR9 in the observed immunological activity with particles, we performed immunization experiments with TLR-deficient mice. Mice (C57BL/6 and TLR9−/−) were vaccinated with rWNVE encapsulated in CpG ODN-modified, LPS-modified, or unmodified nanoparticles. As expected, high titers of rWNVE-specific IgG1 and IgG2b were not present in mice lacking TLR9 after wild-type mice were vaccinated with particles containing CpG ODN (Fig. 5). However, mice deficient in TLR9 maintained a strong response to LPS-modified nanoparticles. Vaccination with CpG-modified and unmodified nanoparticles resulted in similar levels of Abs in TLR9-deficient mice. To validate this finding with a different Ag and, hence, establish generality, a similar experiment is shown in Supplemental Fig. 2 with the model Ag OVA (47).
FIGURE 5.

Ag-specific Ab titers in mice deficient for TLR 9. Groups of WT C57BL/6 or TLR9−/− mice (n = 3) were vaccinated s.c. with 10 μg of CpG/rWNVE, LPS/rWNVE, or −/rWNVE nanoparticles. After 4 wk, mice were bled retro-orbitally, and serum was analyzed for rWNVE-specific 1gG1 (A) and IgG2b (B) by ELISA. *p < 0.05. CpG/rWNVE, recombinant West Nile virus envelope protein encapsulated in CpG-modified nanoparticles; LPS/rWNVE, recombinant West Nile virus envelope protein encapsulated in LPS-modified nanoparticles. WT, wild-type.
CpG ODN-modified nanoparticles induce potent cellular immune responses
To measure levels of cellular activity, mice were vaccinated, and splenocytes were collected and pulsed with rWNVE to measure Ag-specific IL-2 and IFN-γ responses. Lymphocytes exhibited increased activity ex vivo after immunization with CpG ODN-modified nanoparticles compared with animals vaccinated with unmodified nanoparticles or Alhydrogel (Fig. 6A, 6B). High levels of Th1-biased humoral indicators and Ag-specific lymphocyte activity suggest a robust cell-mediated response from the TLR9-targeted, immunostimulatory nanoparticles. Alhydrogel induced a potent humoral response, but poor cytotoxic activity, which is generally associated with viral immunity.
FIGURE 6.

Indicators of cellular immunity. Ag-specific lymphocyte ex vivo activity. Splenocytes were harvested from immunized mice at week 6 and pulsed with 25 μg/ml of rWNVE for 48 h. Supernatant was collected and assayed for IFN-γ (A) and IL-2 (B) by ELISA. C, Effector cell phenotype. Mice were bled retro-orbitally 1 wk postimmunization, and lymphocytes were stained with fluorescent Abs for CD8, CD44, CD127, and KLRG1. Cells were gated for the CD8+CD44+ population, and the percentage of KLRG1+CD127– cells was determined.*p < 0.0001. Data shown are from single experiments that were repeated twice with the same results.
To test for the induction of effector T cell activity, C3H/HeN mice were immunized as before, and circulating lymphocytes were stained for CD8+ effector T cell subset markers at 1 wk postvaccination. High expression of KLRG1 has been correlated to downregulated IL-7R (CD127) (48) and represents a subset of terminally differentiated effector cells. CD8+CD44+ T cells from mice receiving CpG ODN/rWNVE nanoparticles had a larger KLRG1+ CD127− population than any of the mice receiving rWNVE with any other adjuvant (Fig. 6C). No difference in the KLRG1−CD127+ memory T cell subset was observed between any of the groups.
CpG ODN-modified nanoparticles provide strong protection against WN virus infection
To demonstrate protection against live WN virus, C3H/HeN mice were s.c. vaccinated with a single administration of rWNVE-loaded, CpG ODN-modified nanoparticles or rWNVE adsorbed to Alhydrogel and challenged with the live virus. At 2 wk postimmunization, mice were bled and transferred to a biosafety level 3 facility where an i.p. injection of 1000 PFU of WN isolate 2741 was administered. Animals vaccinated with CpG ODN-modified nanoparticles and Alhydrogel generated roughly equal titers of total IgG. CpG ODN elicited a humoral response favoring Th1 responses (high IgG2b) compared with Alhydrogel (predominantly IgG1). The ratios of IgG2b to IgG1 are shown in Fig. 7A. Remarkably, 94% of mice given CpG ODN-modified nanoparticles survived the challenge compared with only 44% of those immunized with Al-hydrogel (Fig. 7B).
FIGURE 7.
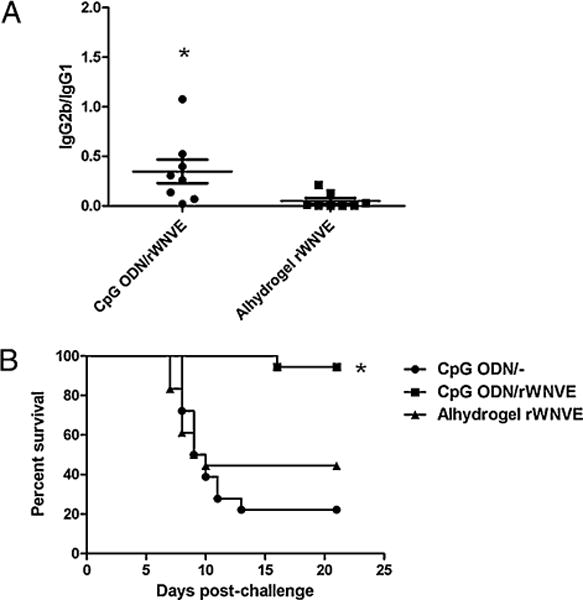
Survival analysis. A, Two weeks after a single immunization, serum samples were collected and analyzed for IgG2b and IgG1 titers by ELISA. Ratio of IgG2b to IgG1 is shown. *p < 0.05; unpaired t test. B, Mice were challenged i.p. with 1000 PFU of WN virus isolate 2741 and were monitored daily. At 21 d postchallenge, the percentage survival for each group was determined. Survival curves are the combined curves from two separate experiments (n = 8 and n = 10), with a combined N of 18 per group. *p < 0.001; log rank test.
Discussion
We believe that vaccine development would be greatly facilitated by a new approach, in which nanoscale modules (Ag, adjuvant, and carrier) are assembled into units that are optimized for stimulating immune responses to a specific pathogen. Toward this goal, our studies aimed to demonstrate this methodology, focusing on the WN virus for which no vaccine has yet been developed.
The components used in this work are well-established safe modules with clinical translational potential. Three clinical trials for CpG ODNs for use as vaccine adjuvants in humans have been completed (49), and PLGA is a U.S. Food and Drug Administration-approved biocompatible, biodegradable polymer with >30 y of use in the drug-delivery field. To keep the Ag and CpG ODN colocalized and to minimize its release, we attached biotinylated CpG ODN to the surface of the nanoparticles by modifying the surface of the nanoparticles with avidin-palmitate. This method minimizes the CpG ODN dose (0.5 μg/mouse), compared with much greater doses ≥100 μg (21) in similar studies. This is critical considering the concerns with autoimmunity and potential lymphoid architectural damage from exposure to high levels of CpG ODN (50).
Recovery from flavivirus infection is accompanied by the appearance of virus-specific, neutralizing Abs. It was shown that the viral envelope protein is the target of the majority of protective Abs (51). Passive immunization experiments showed that Abs may be sufficient to protect mice against a challenge with WN or dengue viruses (5, 37, 39, 52–54). Although control of viremia is generally assumed to be primarily mediated by neutralizing Abs, cellular immunity may play a role as well. For example, it was shown that CD8+ and CD4+ T cells participate in the immune response against a challenge with WN virus (55–57). It was proposed specifically that CD8+ T cells are essential to fully eliminate the infecting virus and prevent viral persistence. The importance of the cellular immune response was made clear by experiments showing that protection against various flaviviruses, such as Japanese encephalitis and tick-borne encephalitis viruses, could be achieved in the absence of neutralizing Abs (58–60). In the case of dengue, challenge of vaccinated rhesus macaques showed that control of viremia was not linearly correlated with neutralizing Ab titer (4). Thus, a vaccine that elicits a cellular immune response may be more effective in preventing infection with flaviviruses compared with one that does not.
Therefore, we hypothesized that the development of a Th1-biased response following immunization with CpG ODN-containing particles would result in a higher degree of protection. Consistent with previous work, which showed that CpG ODN, when given with domain III of the WN virus envelope protein, conferred protection to suckling mice upon transfer of sera from immunized mice (61), we demonstrate similar protective patterns of Th1-associated signals from CpG ODN-modified nanoparticles. Th1-biased responses have been documented elsewhere after administration of CpG ODN given with various delivery vehicles (16, 17, 21, 22, 62). Immunization with CpG ODN-modified nanoparticles resulted in a greater number of circulating effector-like T cells and a higher activity of Ag-specific lymphocytes ex vivo than did immunization with unmodified nanoparticles or Alhydrogel. Faster clearance of virus or infected cells may explain the increased survival in mice receiving the modified nanoparticles.
To generate cellular immunity, many efforts have focused on enhancing cross-presentation, in which exogenous Ag is presented on MHC class I, to promote a strong cytotoxic and Th1 inflammatory bias. Cross-presentation of exogenous Ag can be challenging, because most exogenous Ags that are internalized by a cell are more readily processed into the MHC class II pathway, which typically stimulates CD4 T cell humoral responses. For cross-presentation to occur, Ag must escape from the endosomal compartment into the cytosolic and endoplasmic reticular space where MHC class I processing occurs. Nanoparticles were reported to facilitate Ag cross-presentation, and this intrinsic ability to cross-present their encapsulated cargo was shown previously with PLGA nanoparticles (63). Work with mouse BMDCs showed that PLGA-encapsulated OVA enhanced and sustained Ag presentation in the MHC class I pathway to a much higher degree than did soluble Ag (63). We showed in this study that the addition of CpG motifs to PLGA nanoparticles further increases this Th1 polarization and the resultant cell-mediated immune response. This technology may be particularly beneficial against pathogens best controlled by a Th1-biased response.
The mechanistic aspects of how this system works in vivo have not been fully elucidated. Nanoparticles modified with CpG ODN do not create an IL-1β response in vitro. Thus, unlike alum, which elicits IL-1β secretion with a danger signal, CpG ODN-decorated nanoparticles are noninflammatory, which raises the possibility that the mechanism of action of CpG ODN systems in vivo may not involve inflammasome activation. Future work will delineate the intracellular mechanisms that render this particle system effective in vaccination.
Supplementary Material
Acknowledgments
We thank Dr. Sara Mootien, Jacob Tulipan, Atu Agawu, Michael Look, and Dr. Stephanie Eisenbarth for helpful discussions and assistance. TLR2/4-and TLR9-deficient breeding pairs were a generous gift from Drs. Ruslan Medzhitov and Akiko Iwasaki, respectively. We also thank Dr. Harald Foellmer and Deborah Beck for assistance with survival studies involving WN virus vaccination.
This work was supported by National Science Foundation Nanoscale Interdisciplinary Research Team Grant CTS-0609326 (to T.M.F.) and National Institutes of Health National Institute of Allergy and Infectious Diseases Grant R43 AI078649 (to M.L.).
Abbreviations
- BMDC
bone marrow-derived dendritic cell
- CpG ODN/rWNVE
CpG oligodeoxynucleotide-modified recombinant West Nile virus envelope protein-loaded nanoparticles
- CpG/rWNVE
recombinant West Nile virus envelope protein encapsulated in CpG-modified nanoparticles
- KLRG1
killer cell lectin-like receptor G1
- LPS/rWNVE
recombinant West Nile virus envelope protein encapsulated in LPS-modified nanoparticles
- NR
Nile red
- ODN
oligodeoxynucleo-tide
- PLGA
poly(D,L,-lactide-coglycolide)
- PRNT
plaque-reduction neutralization test
- PVA
poly(vinyl alcohol)
- RT
room temperature
- —/rWNVE
unmodified recombinant West Nile virus envelope protein-loaded nanoparticles
- rWNVE
recombinant West Nile virus envelope protein
- WN
West Nile
- WT
wild-type
Footnotes
The online version of this article contains supplemental information.
Disclosures
T.M.F. is a consultant with L2 Diagnostics. The work described in this article is not a commercial product.
References
- 1.Tsai TF. Flaviviruses. In: Mandell GL, Bennett JE, Dolin R, editors. Principles and Practice of Infectious Diseases. Churchill Livingstone; Philadelphia, PA: 2000. pp. 1196–1206. [Google Scholar]
- 2.Granwehr BP, Lillibridge KM, Higgs S, Mason PW, Aronson JF, Campbell GA, Barrett AD. West Nile virus: where are we now? Lancet Infect Dis. 2004;4:547–556. doi: 10.1016/S1473-3099(04)01128-4. [DOI] [PubMed] [Google Scholar]
- 3.Ng T, Hathaway D, Jennings N, Champ D, Chiang YW, Chu HJ. Equine vaccine for West Nile virus. Dev Biol (Basel) 2003;114:221–227. [PubMed] [Google Scholar]
- 4.Robert Putnak JBA, Coller G, Voss DW, Vaughn D, Clements I, Peters G, Bignami HS, Houng RC, Chen DA, Barvir, et al. An evaluation of dengue type-2 inactivated, recombinant subunit, and live-attenuated vaccine candidates in the rhesus macaque model. Vaccine. 2005;23:4442–4452. doi: 10.1016/j.vaccine.2005.03.042. [DOI] [PubMed] [Google Scholar]
- 5.Tesh RB, Arroyo J, Travassos Da Rosa AP, Guzman H, Xiao SY, Monath TP. Efficacy of killed virus vaccine, live attenuated chimeric virus vaccine, and passive immunization for prevention of West Nile virus encephalitis in hamster model. Emerg Infect Dis. 2002;8:1392–1397. doi: 10.3201/eid0812.020229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Arroyo J, Miller C, Catalan J, Myers GA, Ratterree MS, Trent DW, Monath TP. ChimeriVax-West Nile virus live-attenuated vaccine: preclinical evaluation of safety, immunogenicity, and efficacy. J Virol. 2004;78:12497–12507. doi: 10.1128/JVI.78.22.12497-12507.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Monath TP, Liu J, Kanesa-Thasan N, Myers GA, Nichols R, Deary A, McCarthy K, Johnson C, Ermak T, Shin S, et al. A live, attenuated recombinant West Nile virus vaccine. Proc Natl Acad Sci USA. 2006;103:6694–6699. doi: 10.1073/pnas.0601932103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Pletnev AG, Putnak R, Speicher J, Wagar EJ, Vaughn DW. West Nile virus/dengue type 4 virus chimeras that are reduced in neurovirulence and peripheral virulence without loss of immunogenicity or protective efficacy. Proc Natl Acad Sci USA. 2002;99:3036–3041. doi: 10.1073/pnas.022652799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Davis BS, Chang GJ, Cropp B, Roehrig JT, Martin DA, Mitchell CJ, Bowen R, Bunning ML. West Nile virus recombinant DNA vaccine protects mouse and horse from virus challenge and expresses in vitro a noninfectious recombinant antigen that can be used in enzyme-linked immunosorbent assays. J Virol. 2001;75:4040–4047. doi: 10.1128/JVI.75.9.4040-4047.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Simmons M, Murphy GS, Kochel T, Raviprakash K, Hayes CG. Characterization of antibody responses to combinations of a dengue-2 DNA and dengue-2 recombinant subunit vaccine. Am J Trop Med Hyg. 2001;65:420–426. doi: 10.4269/ajtmh.2001.65.420. [DOI] [PubMed] [Google Scholar]
- 11.Holman DH, Wang D, Raviprakash K, Raja NU, Luo M, Zhang J, Porter KR, Dong JY. Two complex, adenovirus-based vaccines that together induce immune responses to all four dengue virus serotypes. Clin Vaccine Immunol. 2007;14:182–189. doi: 10.1128/CVI.00330-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Minke JM, Siger L, Karaca K, Austgen L, Gordy P, Bowen R, Renshaw RW, Loosmore S, Audonnet JC, Nordgren B. Recombinant canarypoxvirus vaccine carrying the prM/E genes of West Nile virus protects horses against a West Nile virus-mosquito challenge. Arch Virol Suppl. 2004:221–230. doi: 10.1007/978-3-7091-0572-6_20. [DOI] [PubMed] [Google Scholar]
- 13.Ledizet M, Kar K, Foellmer HG, Wang T, Bushmich SL, Anderson JF, Fikrig E, Koski RA. A recombinant envelope protein vaccine against West Nile virus. Vaccine. 2005;23:3915–3924. doi: 10.1016/j.vaccine.2005.03.006. [DOI] [PubMed] [Google Scholar]
- 14.Lieberman MM, Clements DE, Ogata S, Wang G, Corpuz G, Wong T, Martyak T, Gilson L, Coller BA, Leung J, et al. Preparation and immunogenic properties of a recombinant West Nile subunit vaccine. Vaccine. 2007;25:414–423. doi: 10.1016/j.vaccine.2006.08.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Demento SL, Eisenbarth SC, Foellmer HG, Platt C, Caplan MJ, Mark Saltzman W, Mellman I, Ledizet M, Fikrig E, Flavell RA, Fahmy TM. Inflammasome-activating nanoparticles as modular systems for optimizing vaccine efficacy. Vaccine. 2009;27:3013–3021. doi: 10.1016/j.vaccine.2009.03.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Gürsel M, Tunca S, Ozkan M, Ozcengiz G, Alaeddinoglu G. Immunoadjuvant action of plasmid DNA in liposomes. Vaccine. 1999;17:1376–1383. doi: 10.1016/s0264-410x(98)00383-1. [DOI] [PubMed] [Google Scholar]
- 17.Gursel I, Gursel M, Ishii KJ, Klinman DM. Sterically stabilized cationic liposomes improve the uptake and immunostimulatory activity of CpG oligonucleotides. J Immunol. 2001;167:3324–3328. doi: 10.4049/jimmunol.167.6.3324. [DOI] [PubMed] [Google Scholar]
- 18.Li WM, Bally MB, Schutze-Redelmeier MP. Enhanced immune response to T-independent antigen by using CpG oligodeoxynucleotides encapsulated in liposomes. Vaccine. 2001;20:148–157. doi: 10.1016/s0264-410x(01)00277-8. [DOI] [PubMed] [Google Scholar]
- 19.Joseph A, Louria-Hayon I, Plis-Finarov A, Zeira E, Zakay-Rones Z, Raz E, Hayashi T, Takabayashi K, Barenholz Y, Kedar E. Liposomal immunostimulatory DNA sequence (ISS-ODN): an efficient parenteral and mucosal adjuvant for influenza and hepatitis B vaccines. Vaccine. 2002;20:3342–3354. doi: 10.1016/s0264-410x(02)00295-5. [DOI] [PubMed] [Google Scholar]
- 20.Singh M, Ott G, Kazzaz J, Ugozzoli M, Briones M, Donnelly J, O’Hagan DT. Cationic microparticles are an effective delivery system for immune stimulatory cpG DNA. Pharm Res. 2001;18:1476–1479. doi: 10.1023/a:1012269226066. [DOI] [PubMed] [Google Scholar]
- 21.Diwan M, Tafaghodi M, Samuel J. Enhancement of immune responses by co-delivery of a CpG oligodeoxynucleotide and tetanus toxoid in biodegradable nanospheres. J Control Release. 2002;85:247–262. doi: 10.1016/s0168-3659(02)00275-4. [DOI] [PubMed] [Google Scholar]
- 22.Zhang XQ, Dahle CE, Baman NK, Rich N, Weiner GJ, Salem AK. Potent antigen-specific immune responses stimulated by codelivery of CpG ODN and antigens in degradable microparticles. J Immunother. 2007;30:469–478. doi: 10.1097/CJI.0b013e31802fd8c6. [DOI] [PubMed] [Google Scholar]
- 23.Kaiser-Schulz G, Heit A, Quintanilla-Martinez L, Hammerschmidt F, Hess S, Jennen L, Rezaei H, Wagner H, Schätzl HM. Polylactide-coglycolide microspheres co-encapsulating recombinant tandem prion protein with CpG-oligonucleotide break self-tolerance to prion protein in wild-type mice and induce CD4 and CD8 T cell responses. J Immunol. 2007;179:2797–2807. doi: 10.4049/jimmunol.179.5.2797. [DOI] [PubMed] [Google Scholar]
- 24.Borges O, Cordeiro-da-Silva A, Tavares J, Santarém N, de Sousa A, Borchard G, Junginger HE. Immune response by nasal delivery of hepatitis B surface antigen and codelivery of a CpG ODN in alginate coated chitosan nanoparticles. Eur J Pharm Biopharm. 2008;69:405–416. doi: 10.1016/j.ejpb.2008.01.019. [DOI] [PubMed] [Google Scholar]
- 25.Xie H, Gursel I, Ivins BE, Singh M, O’Hagan DT, Ulmer JB, Klinman DM. CpG oligodeoxynucleotides adsorbed onto polylactide-co-glycolide microparticles improve the immunogenicity and protective activity of the licensed anthrax vaccine. Infect Immun. 2005;73:828–833. doi: 10.1128/IAI.73.2.828-833.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Heit A, Schmitz F, Haas T, Busch DH, Wagner H. Antigen co-encapsulated with adjuvants efficiently drive protective T cell immunity. Eur J Immunol. 2007;37:2063–2074. doi: 10.1002/eji.200737169. [DOI] [PubMed] [Google Scholar]
- 27.Hemmi H, Takeuchi O, Kawai T, Kaisho T, Sato S, Sanjo H, Matsumoto M, Hoshino K, Wagner H, Takeda K, Akira S. A Toll-like receptor recognizes bacterial DNA. Nature. 2000;408:740–745. doi: 10.1038/35047123. [DOI] [PubMed] [Google Scholar]
- 28.Krieg AM. CpG motifs in bacterial DNA and their immune effects. Annu Rev Immunol. 2002;20:709–760. doi: 10.1146/annurev.immunol.20.100301.064842. [DOI] [PubMed] [Google Scholar]
- 29.Barry ME, Pinto-González D, Orson FM, McKenzie GJ, Petry GR, Barry MA. Role of endogenous endonucleases and tissue site in transfection and CpG-mediated immune activation after naked DNA injection. Hum Gene Ther. 1999;10:2461–2480. doi: 10.1089/10430349950016816. [DOI] [PubMed] [Google Scholar]
- 30.Soni PN, Brown D, Saffie R, Savage K, Moore D, Gregoriadis G, Dusheiko GM. Biodistribution, stability, and antiviral efficacy of liposome-entrapped phosphorothioate antisense oligodeoxynucleotides in ducks for the treatment of chronic duck hepatitis B virus infection. Hepatology. 1998;28:1402–1410. doi: 10.1002/hep.510280532. [DOI] [PubMed] [Google Scholar]
- 31.Tighe H, Takabayashi K, Schwartz D, Marsden R, Beck L, Corbeil J, Richman DD, Eiden JJ, Jr, Spiegelberg HL, Raz E. Conjugation of protein to immunostimulatory DNA results in a rapid, long-lasting and potent induction of cell-mediated and humoral immunity. Eur J Immunol. 2000;30:1939–1947. doi: 10.1002/1521-4141(200007)30:7<1939::AID-IMMU1939>3.0.CO;2-#. [DOI] [PubMed] [Google Scholar]
- 32.Mutwiri GK, Nichani AK, Babiuk S, Babiuk LA. Strategies for enhancing the immunostimulatory effects of CpG oligodeoxynucleotides. J Control Release. 2004;97:1–17. doi: 10.1016/j.jconrel.2004.02.022. [DOI] [PubMed] [Google Scholar]
- 33.Nierkens S, den Brok MH, Sutmuller RP, Grauer OM, Bennink E, Morgan ME, Figdor CG, Ruers TJ, Adema GJ. In vivo colocalization of antigen and CpG [corrected] within dendritic cells is associated with the efficacy of cancer immunotherapy. Cancer Res. 2008;68:5390–5396. doi: 10.1158/0008-5472.CAN-07-6023. [DOI] [PubMed] [Google Scholar]
- 34.O’Hagan DT, Rahman D, McGee JP, Jeffery H, Davies MC, Williams P, Davis SS, Challacombe SJ. Biodegradable microparticles as controlled release antigen delivery systems. Immunology. 1991;73:239–242. [PMC free article] [PubMed] [Google Scholar]
- 35.Fahmy TM, Samstein RM, Harness CC, Mark Saltzman W. Surface modification of biodegradable polyesters with fatty acid conjugates for improved drug targeting. Biomaterials. 2005;26:5727–5736. doi: 10.1016/j.biomaterials.2005.02.025. [DOI] [PubMed] [Google Scholar]
- 36.Durbin AP, Karron RA, Sun W, Vaughn DW, Reynolds MJ, Perreault JR, Thumar B, Men R, Lai CJ, Elkins WR, et al. Attenuation and immunogenicity in humans of a live dengue virus type-4 vaccine candidate with a 30 nucleotide deletion in its 3′-untranslated region. Am J Trop Med Hyg. 2001;65:405–413. doi: 10.4269/ajtmh.2001.65.405. [DOI] [PubMed] [Google Scholar]
- 37.Goncalvez AP, Men R, Wernly C, Purcell RH, Lai CJ. Chimpanzee Fab fragments and a derived humanized immunoglobulin G1 antibody that efficiently cross-neutralize dengue type 1 and type 2 viruses. J Virol. 2004;78:12910–12918. doi: 10.1128/JVI.78.23.12910-12918.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Bonafé N, Rininger JA, Chubet RG, Foellmer HG, Fader S, Anderson JF, Bushmich SL, Anthony K, Ledizet M, Fikrig E, et al. A recombinant West Nile virus envelope protein vaccine candidate produced in Spodoptera frugiperda expresSF+ cells. Vaccine. 2009;27:213–222. doi: 10.1016/j.vaccine.2008.10.046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Ledizet M, Kar K, Foellmer HG, Bonafé N, Anthony KG, Gould LH, Bushmich SL, Fikrig E, Koski RA. Antibodies targeting linear determinants of the envelope protein protect mice against West Nile virus. J Infect Dis. 2007;196:1741–1748. doi: 10.1086/523654. [DOI] [PubMed] [Google Scholar]
- 40.Mundargi RC, Babu VR, Rangaswamy V, Patel P, Aminabhavi TM. Nano/micro technologies for delivering macromolecular therapeutics using poly(D,L-lactide-co-glycolide) and its derivatives. J Control Release. 2008;125:193–209. doi: 10.1016/j.jconrel.2007.09.013. [DOI] [PubMed] [Google Scholar]
- 41.Panyam J, Dali MM, Sahoo SK, Ma W, Chakravarthi SS, Amidon GL, Levy RJ, Labhasetwar V. Polymer degradation and in vitro release of a model protein from poly(D,L-lactide-co-glycolide) nano- and. micropar-ticles. J Control Release. 2003;92:173–187. doi: 10.1016/s0168-3659(03)00328-6. [DOI] [PubMed] [Google Scholar]
- 42.Carr R, Warren J, Knowles J. 2009 Available at: http://www.nanosight.com. Accessed May.
- 43.Macatonia SE, Hosken NA, Litton M, Vieira P, Hsieh CS, Culpepper JA, Wysocka M, Trinchieri G, Murphy KM, O’Garra A. Dendritic cells produce IL-12 and direct the development of Th1 cells from naive CD4+ T cells. J Immunol. 1995;154:5071–5079. [PubMed] [Google Scholar]
- 44.Diehl S, Rincón M. The two faces of IL-6 on Th1/Th2 differentiation. Mol Immunol. 2002;39:531–536. doi: 10.1016/s0161-5890(02)00210-9. [DOI] [PubMed] [Google Scholar]
- 45.Eisenbarth SC, Colegio OR, O’Connor W, Sutterwala FS, Flavell RA. Crucial role for the Nalp3 inflammasome in the immunostimulatory properties of aluminium adjuvants. Nature. 2008;453:1122–1126. doi: 10.1038/nature06939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Sharp FA, Ruane D, Claass B, Creagh E, Harris J, Malyala P, Singh M, O’Hagan DT, Pétrilli V, Tschopp J, et al. Uptake of particulate vaccine adjuvants by dendritic cells activates the NALP3 inflammasome. Proc Natl Acad Sci USA. 2009;106:870–875. doi: 10.1073/pnas.0804897106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Felnerova D, Viret JF, Glück R, Moser C. Liposomes and virosomes as delivery systems for antigens, nucleic acids and drugs. Curr Opin Biotechnol. 2004;15:518–529. doi: 10.1016/j.copbio.2004.10.005. [DOI] [PubMed] [Google Scholar]
- 48.Joshi NS, Cui W, Chandele A, Lee HK, Urso DR, Hagman J, Gapin L, Kaech SM. Inflammation directs memory precursor and short-lived effector CD8(+) T cell fates via the graded expression of T-bet transcription factor. Immunity. 2007;27:281–295. doi: 10.1016/j.immuni.2007.07.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Klinman DM, Klaschik S, Sato T, Tross D. CpG oligonucleotides as adjuvants for vaccines targeting infectious diseases. Adv Drug Deliv Rev. 2009;61:248–255. doi: 10.1016/j.addr.2008.12.012. [DOI] [PubMed] [Google Scholar]
- 50.Heikenwalder M, Polymenidou M, Junt T, Sigurdson C, Wagner H, Akira S, Zinkernagel R, Aguzzi A. Lymphoid follicle destruction and immunosuppression after repeated CpG oligodeoxynucleotide administration. Nat Med. 2004;10:187–192. doi: 10.1038/nm987. [DOI] [PubMed] [Google Scholar]
- 51.Brinton MA. The molecular biology of West Nile Virus: a new invader of the western hemisphere. Annu Rev Microbiol. 2002;56:371–402. doi: 10.1146/annurev.micro.56.012302.160654. [DOI] [PubMed] [Google Scholar]
- 52.Engle MJ, Diamond MS. Antibody prophylaxis and therapy against West Nile virus infection in wild-type and immunodeficient mice. J Virol. 2003;77:12941–12949. doi: 10.1128/JVI.77.24.12941-12949.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Gould LH, Sui J, Foellmer H, Oliphant T, Wang T, Ledizet M, Murakami A, Noonan K, Lambeth C, Kar K, et al. Protective and therapeutic capacity of human single-chain Fv-Fc fusion proteins against West Nile virus. J Virol. 2005;79:14606–14613. doi: 10.1128/JVI.79.23.14606-14613.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Oliphant T, Engle M, Nybakken GE, Doane C, Johnson S, Huang L, Gorlatov S, Mehlhop E, Marri A, Chung KM, et al. Development of a humanized monoclonal antibody with therapeutic potential against West Nile virus. Nat Med. 2005;11:522–530. doi: 10.1038/nm1240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Purtha WE, Myers N, Mitaksov V, Sitati E, Connolly J, Fremont DH, Hansen TH, Diamond MS. Antigen-specific cytotoxic T lymphocytes protect against lethal West Nile virus encephalitis. Eur. J Immunol. 2007;37:1845–1854. doi: 10.1002/eji.200737192. [DOI] [PubMed] [Google Scholar]
- 56.Shrestha B, Diamond MS. Role of CD8+ T cells in control of West Nile virus infection. J Virol. 2004;78:8312–8321. doi: 10.1128/JVI.78.15.8312-8321.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Sitati EM, Diamond MS. CD4+ T-cell responses are required for clearance of West Nile virus from the central nervous system. J Virol. 2006;80:12060–12069. doi: 10.1128/JVI.01650-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Ashok MS, Rangarajan PN. Immunization with plasmid DNA encoding the envelope glycoprotein of Japanese Encephalitis virus confers significant protection against intracerebral viral challenge without inducing detectable antiviral antibodies. Vaccine. 1999;18:68–75. doi: 10.1016/s0264-410x(99)00180-2. [DOI] [PubMed] [Google Scholar]
- 59.Chen HW, Pan CH, Liau MY, Jou R, Tsai CJ, Wu HJ, Lin YL, Tao MH. Screening of protective antigens of Japanese encephalitis virus by DNA immunization: a comparative study with conventional viral vaccines. J Virol. 1999;73:10137–10145. doi: 10.1128/jvi.73.12.10137-10145.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Kofler RM, Aberle JH, Aberle SW, Allison SL, Heinz FX, Mandl CW. Mimicking live flavivirus immunization with a noninfectious RNA vaccine. Proc Natl Acad Sci USA. 2004;101:1951–1956. doi: 10.1073/pnas.0307145101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Chu JH, Chiang CC, Ng ML. Immunization of flavivirus West Nile recombinant envelope domain III protein induced specific immune response and protection against West Nile virus infection. J Immunol. 2007;178:2699–2705. doi: 10.4049/jimmunol.178.5.2699. [DOI] [PubMed] [Google Scholar]
- 62.Mui B, Raney SG, Semple SC, Hope MJ. Immune stimulation by a CpG-containing oligodeoxynucleotide is enhanced when encapsulated and delivered in lipid particles. J Pharmacol Exp Ther. 2001;298:1185–1192. [PubMed] [Google Scholar]
- 63.Shen H, Ackerman AL, Cody V, Giodini A, Hinson ER, Cresswell P, Edelson RL, Saltzman WM, Hanlon DJ. Enhanced and prolonged cross-presentation following endosomal escape of exogenous antigens encapsulated in biodegradable nanoparticles. Immunology. 2006;117:78–88. doi: 10.1111/j.1365-2567.2005.02268.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.