

Abstract
The relevance of claudin-6 and claudin-9 in hepatitis C virus (HCV) entry remains elusive. We produced claudin-6- or claudin-9-specific monoclonal antibodies that inhibit HCV entry into nonhepatic cells expressing exogenous claudin-6 or claudin-9. These antibodies had no effect on HCV infection of hepatoma cells or primary hepatocytes. Thus, although claudin-6 and claudin-9 can serve as entry factors in cell lines, HCV infection into human hepatocytes is not dependent on claudin-6 and claudin-9.
TEXT
Hepatitis C virus (HCV) enters cells via a multistep process that requires viral and host cell factors, including the viral envelope glycoproteins E1 and E2, tetraspanin CD81, scavenger receptor class B type I (SR-BI), tight junction proteins claudin-1 (CLDN1) and occludin (OCLN), as well as coentry factors such as epidermal growth factor receptor (EGFR), ephrin receptor A2 (EphA2), Niemann-Pick C1-Like1 (NPC1L1) and transferrin receptor 1 (1–3). Understanding the mechanisms of viral entry is a prerequisite to defining antiviral therapies targeting an early step(s) in the viral life cycle.
CLDNs are critical components of tight junctions (TJ) and regulate paracellular permeability and polarity. The CLDN superfamily comprises more than 20 members that are expressed in a tissue-specific manner. CLDN1 is an essential host factor defining HCV entry (4), and CLDN1-specific antibodies inhibit HCV infection of human hepatocytes in vitro (5, 6). CLDN1 associates with CD81 in a variety of cell types, and the resulting receptor complex is essential for HCV infection (5, 7–9). CLDN6 and CLDN9 have been reported to mediate HCV entry in CLDN1-deficient Bel7402 hepatoma cells (10) and CLDN-null 293T embryonic kidney-derived cells (11–13). State-of-the-art cell culture models that support HCV replication include human hepatoma Huh7-derived cell lines and primary human hepatocytes (PHHs). However, the role of CLDN6 and CLDN9 in mediating HCV infection of these cells and as potential antiviral targets is unknown.
To investigate the functional role of CLDN6 and CLDN9 in HCV entry, we generated monoclonal antibodies (MAbs) by genetic immunization using full-length human CLDN6 or CLDN9 cDNA expression vectors as previously described (6, 14). Following lymphocyte fusion, 10 96-well plates were screened for each target using transiently transfected cells expressing specific CLDNs on their cell surface, and 54 positive clones were subsequently amplified and subcloned. We selected five CLDN6 (WU-8F5-E7, WU-9E1-G2, WU-5H6-D6, WU-10A4-B9, WU-3C9-B11)- and three CLDN9 (YD-4E9-A2, YD-6F9-H2, YD-1C4-A4)-specific MAbs that bind specific CLDNs expressed on 293T cells without any cross-reactivity for further studies (6) (Fig. 1A). To characterize these antibodies, we first used well-characterized 293T cells that do not endogenously express CLDNs (12, 13, 15) (Fig. 1B) and thus allow us to express each target CLDN individually (Fig. 1C). A previously described CLDN1-specific MAb (OM-7D3-B3) was used as the control (6). We confirmed that, in contrast to naive 293T cells, HCV pseudoparticles (HCVpp) expressing a diverse panel of glycoproteins (strains H77 [genotype 1a], HCV-J [1b], JFH1 [2a], UKN3A1.28 [3a], and UKN4.21.16 [4], described in reference 16) infect 293T cells engineered to express CLDN1, CLDN6, or CLDN9 (Fig. 1D). In contrast to previous reports (10, 11), we noted that HCV-JFH1 only infected 293T cells expressing CLDN1, suggesting that this strain cannot utilize CLDN6 or CLDN9. Interestingly, a recent study also reported that Huh6 cells, expressing CLDN6 but devoid of CLDN1, are resistant to HCVpp expressing genotype 2a glycoproteins in contrast to HCVpp of genotype 1 (17). Next, we assessed the ability of CLDN-specific MAbs to inhibit HCVpp entry into these cells by using a CD81-specific antibody as a positive control (18). All of the CLDN6- and CLDN9-specific MAbs inhibited the entry of HCVpp expressing representative genotype 1a and 1b glycoproteins into CLDN-expressing 293T cells (Fig. 1E). The two most potent MAbs were further characterized for their dose-dependent inhibition of entry of HCVpp genotype 1a and 1b (Fig. 2A and B) and cross-reactivity to inhibit a panel of HCVpp expressing diverse glycoproteins. In contrast to mouse leukemia virus pseudoparticle (MLVpp) entry (19), CLDN6- and CLDN9-specific MAbs inhibited the entry of HCVpp expressing glycoproteins of genotypes 3a and 4 into 293T-derived cells (Fig. 2C). These data demonstrate that CLDN6- and CLDN9-specific MAbs inhibit HCV entry of 293T cells in a genotype-independent manner.
Fig 1.

CLDN6- and CLDN9-specific MAbs inhibit HCVpp infection of 293T cells engineered to express CLDNs. (A) Reactivity of anti-CLDN MAbs for 293T cells transfected with different human CLDNs. CLDN-deficient 293T cells were transfected to express Aequorea coerulescens green fluorescent protein (AcGFP)-tagged human CLDN1, CLDN6, or CLDN9 as described previously (6) before detachment and staining with CLDN-specific MAbs or control isotype-matched irrelevant rat IgG at 20 μg/ml. Primary bound antibodies were detected with a goat anti-rat phycoerythrin (PE)-conjugated F(ab′)2 fragment (PN IM 1623; Beckman Coulter). After a washing step, the cells were fixed with 2% paraformaldehyde (PFA) and analyzed by flow cytometry (FACscan). AcGFP-tagged CLDN protein expression was confirmed by flow cytometric quantification of AcGFP expression (data not shown). Specific binding of CLDN1 (OM-7D3-B3) (6), CLDN6 (WU-8F5-E7, WU-9E1-G2, WU-5H6-D6, WU-10A4-B9, and WU-3C9-B11) and CLDN9 (YD-4E9-A2, YD-6F9-H2, and YD-1C4-A4) MAbs to AcGFP-tagged CLDNs is shown as the difference of the mean fluorescence intensities of cells stained with the CLDN-specific MAb and cells stained with the control rat MAb isotype. (B) Expression of HCV entry factors in 293T, HeLa, HepG2, Bcl-2, and Huh7.5 cells. RNA was purified from these cell lysates using a Qiagen RNA minikit by following the manufacturer's instructions. Samples were amplified using a Cells Direct reverse transcription-PCR (RT-PCR) kit (Invitrogen). A primer-limited glyceraldehyde-3-phosphate dehydrogenase (GAPDH) referent primer set was included in all PCRs and used to compare expression. (C) Entry factor expression in CLDN1-, CLDN6-, or CLDN9-transfected 293T cells. The relative expression of each entry factor was determined by flow cytometry and is indicated as n-fold expression compared to results with parental 293T cells. (D) HCVpp strains (H77 [genotype 1a], HCV-J [1b], JFH1 [2a], UKN3A1.28 [3a], and UKN4.21.16 [4]; produced as described in reference 16) were allowed to infect CLDN1-, CLDN6-, or CLDN9-293T cells, and infection was assessed by luciferase activity in cell lysates 72 h postinfection. Results are expressed in relative light units (RLU). The threshold for a detectable infection in this system is indicated by a dashed line, corresponding to the mean ± 3 standard deviations (SD) of background levels, i.e., luciferase activity of naive noninfected cells or cells infected with pseudotypes without HCV envelopes. Means ± SD from three independent experiments performed in triplicate are shown. (E) 293T cells engineered to express CDLN1 (left panel), CLDN6 (middle panel), or CLDN9 (right panel) were preincubated for 1 h at 37°C with a CD81-specific (10 μg/ml), CLDN-specific, or control (50 μg/ml) MAb before infection with HCVpp (strains H77 [genotype 1a] and HCV-J [1b]) for 4 h at 37°C. HCVpp entry was assessed as described for panel D. Means ± SD from three independent experiments performed in triplicate are shown. Statistical analysis relative to the control MAb was performed using the Student t test; *, P < 0.05.
Fig 2.
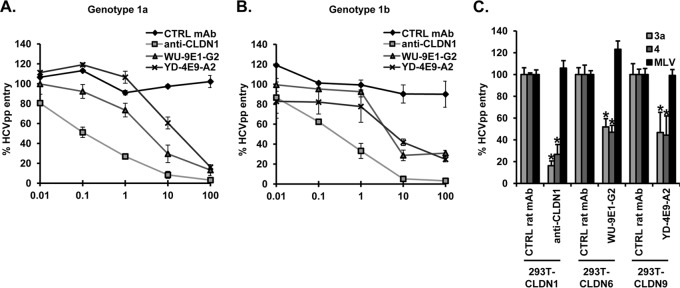
Anti-CLDN genotype-independent inhibition of HCVpp infection of 293T cells engineered to express CLDN receptors. CLDN1-, CLDN6-, and CLDN9-293T cells were preincubated for 1 h at 37°C with (A and B) serial dilutions of or (C) a fixed dose (100 μg/ml) of a CLDN1-, CLDN6 (WU-9E1-G2)-, CLDN9 (YD-4E9-A2)-specific, or control MAb before infection with HCVpp (strains H77 [genotype 1a], HCV-J [1b], UKN3A1.28 [3a], and UKN4.21.16 [4]) or MLVpp for 4 h at 37°C. Pseudoparticle entry was assessed as described in the legend to Fig. 1. Means ± SD from three independent experiments performed in triplicate are shown. Statistical analysis relative to the control MAb was performed using the Student t test, *P < 0.05.
CLDN6 and CLDN9 mRNA have been reported to be expressed in human liver, albeit at low levels (10). All of the CLDN6-specific MAbs bound Huh7.5.1 cells with values comparable to that of the CLDN1-specific MAb, whereas we failed to detect binding of the anti-CLDN9 MAbs (Fig. 3A). These data suggest that CLDN6 is expressed in this hepatoma cell line and that CLDN9 is either not or weakly expressed on Huh7.5.1 cells. This is consistent with CLDN6 and CLDN9 mRNA expression in these cells (Fig. 1B). Next, to investigate whether the antibodies inhibit HCV entry into human hepatoma cells, Huh7.5.1 cells were preincubated with CLDN-specific MAbs before infection with HCVpp from different genotypes (strains H77 [genotype 1a], HCV-J [1b], and JFH1 [2a]). Surprisingly, in contrast to CLDN1-specific MAb and to results obtained with CLDN-expressing 293T cells (Fig. 1 and 2), none of the CLDN6- or CLDN9-specific MAbs inhibited HCVpp entry into Huh7.5.1 cells, even at high concentrations (Fig. 3B). Furthermore, none of the CLDN6- or CLDN9-specific MAbs inhibited infection of cell culture-derived HCV (HCVcc; strains Luc-Con1 [genotype 1b/2a] and Luc-Jc1 [2a/2a], described in reference 20) in these cells (Fig. 3C).
Fig 3.

CLDN6- and CLDN9-specific MAbs do not inhibit HCV infection in Huh7.5.1 cells. (A) Binding of CLDN-specific MAbs to Huh7.5.1 cells. Huh7.5.1 cells were detached and incubated with CLDN-specific MAbs. MAb binding (4 μg/ml) was revealed by flow cytometry using a CLDN1-specific MAb (OM-7D3-B3) as a positive control. Inhibition of (B) HCVpp entry or (C) HCVcc infection by CLDN-specific MAbs. Huh7.5.1 cells were preincubated for 1 h at 37°C with CLDN-specific or control MAbs (100 μg/ml) before infection with HCVpp (strains H77 [genotype 1a], HCV-J [1b], and JFH1 [2a]) or HCVcc (strains Luc-Con1 [genotype 1b/2a] and Luc-Jc1 [2a/2a]) for 4 h at 37°C. HCV infection was assessed by luciferase activity in cell lysates 72 h postinfection. (D) Inhibition of HCVpp entry in CLDN1-silenced (siCLDN1) Huh7.5.1 cells. CLDN1-silenced Huh7.5.1 (4) were preincubated for 1 h at 37°C with CLDN-specific or control MAbs (100 μg/ml) before infection with HCVpp (strains H77 [1a], HCV-J [1b], JFH1 [2a], UKN3A1.28 [3a], and UKN4.21.16 [4]). Western blot demonstrating CLDN1 silencing is indicated below; siCTRL, silencing control. Means ± SD from three independent experiments performed in triplicate are shown. Statistical analysis relative to the control MAb was performed using the Student t test; *, P < 0.05. MW, molecular weight in thousands.
Exogenous expression of CLDN6 or CLDN9 in CLDN1-silenced Huh7.5 has been reported to confer a small but statistically significant level of HCVpp genotype 1b entry but to have no effect on HCVcc infection (11). To investigate whether CDLN6 or CLDN9 may be used as a substitute for CLDN1 in Huh7.5.1 cells, we assessed the ability of CLDN-specific MAbs to inhibit HCVpp entry in CLDN1-silenced Huh7.5.1 cells. In contrast to the CLDN1-specific MAb, neither CLDN6- nor CLDN9-specific MAbs were able to further decrease HCV entry into CLDN1-silenced Huh7.5.1 cells (Fig. 3D). To further investigate a potential interplay between CLDN1 and other CLDNs during HCV infection, we studied whether the combination of CLDN6- or CLDN9- and CLDN1-specific MAbs provided an additive or synergistic inhibitory effect(s) on HCVpp entry (6). Combining CLDN6- or CLDN9-specific MAbs with the CLDN1-specific MAb did not result in any additive effect on inhibition of HCVpp entry of different genotypes (Fig. 4). Taken together, these data suggest a limited role of CLDN6 and CLDN9 in HCV entry into Huh7-derived cell lines.
Fig 4.

Combining CLDN6- or CLDN9-specific MAbs with an anti-CLDN1 MAb does not result in an additive neutralizing effect. Huh7.5.1 cells were preincubated with serial concentrations of the CLDN1-specific or respective isotype control MAb for 1 h at 37°C and either a (A) CLDN6- (WU-9E1-G2, upper panel)- or (B) CLDN9 (YD-4E9-G2, lower panel)-specific MAb (10 or 100 μg/ml) before infection with HCVpp (strains H77 [genotype 1a], HCV-J [1b], JFH1 [2a], and UKN3A1.28 [3a]) in the presence of both compounds. HCVpp infection was analyzed as described in the legend to Fig. 3. Means ± SD from one representative experiment performed in triplicate are shown.
Finally, to further investigate the in vivo relevance of CLDN6 and CLDN9 as potential HCV entry factors and antiviral targets, we performed similar experiments on the natural and most relevant target cell of HCV, the human hepatocyte. In comparison to the CLDN1-specific MAb, we noted low to negligible binding of CLDN6- or CLDN9-specific MAbs to PHHs (Fig. 5A). These data suggest that CLDN6 and CLDN9 are either not or weakly expressed in PHHs. Most importantly, in contrast to the CLDN1-specific MAb, CLDN6- and CLDN9-specific MAbs had a minimal or an absent effect(s) on HCVpp infection of PHHs (Fig. 5B). These data are consistent with the limited detection of CLDN6 and CLDN9 mRNA in hepatocytes isolated from a range of donors, in contrast to the high-level expression of CD81, SR-BI, and CLDN1 (Fig. 5C). Another study recently reported high levels of the main HCV entry factors but highly variable CLDN6 mRNA levels in liver biopsy specimens from HCV patients (17). Immunostaining of liver sections also demonstrated that, in contrast to CLDN6-positive seminoma, CLDN6 was not detected in human liver sections (Fig. 5D).
Fig 5.

CLDN6- and CLDN9-specific MAbs do not inhibit HCVpp infection of PHHs. (A) Binding of CLDN-specific MAbs to PHHs. Following PHH isolation, cells were incubated with CLDN-specific MAbs. MAb binding (20 μg/ml) was assessed as described in the legend to Fig. 3. (B) Inhibition of HCVpp entry into PHHs by CLDN-specific MAbs (100 μg/ml) was performed as described in the legend to Fig. 3. Means ± SD from three independent experiments performed in triplicate are shown. Statistical analysis relative to the control MAb was performed using the Student t test; *, P < 0.05. (C) Expression of HCV entry factors in PHHs. Total RNA was extracted from hepatocyte samples (260 to 282) and HeLa, Huh7.5, and Caco-2 cells, and mRNA copies were determined by quantitative RT-PCR. Absolute quantities were normalized to GAPDH, and data are displayed as percent expression relative to CD81. Means ± SD from one experiment performed in quadruplicate are shown. Any signal of <0.1% of that of CD81 (1,000-fold lower) is negligible. (D) CLDN6 expression in normal liver. CLDN6 expression was analyzed by immunohistochemistry on formalin-fixed paraffin-embedded normal donor and normal liver (right panel), distal to tumor resection tissues (21). A seminoma case was used as a positive control (middle panel), and a control IgG was performed as a negative control (left panel). Positive staining is shown in red. Microscopic analysis of tissues showed strong membranous staining of the tumor cells for CLDN6 in the positive seminoma control section. Normal donor and resection liver tissue demonstrated no detectable CLND6 expression, and IgG controls on all sections were negative.
The aim of this study was to explore the functional role of CLDN6 and CLDN9 in the HCV entry process in PHHs and hepatoma cells. Understanding the mechanism of HCV entry in the most relevant cell culture model will allow the development of more efficient antivirals. We generated novel CLDN6- and CLDN9-specific MAbs that specifically interact with CLDN6 or CLDN9 (Fig. 1A) and inhibit HCVpp entry into human embryonic 293T kidney cells expressing defined CLDN receptors that have been classically used to assess the role of CLDNs in the HCV entry process (Fig. 1 and 2). These data demonstrate that the antibodies are functional with respect to binding cognate receptors and inhibiting HCV internalization. In line with mRNA expression data (Fig. 1B), antibody binding studies demonstrate expression of CLDN6 on Huh7.5.1 cells (Fig. 3A), albeit with lower binding values than those observed with 293T cells. However, CLDN6- or CLDN9-specific MAbs showed low or absent binding to PHHs (Fig. 5A), suggesting minimal expression, a conclusion further supported by the low mRNA levels of these CLDNs in PHHs isolated from multiple donors and absent detection of CLDN6 in liver sections (Fig. 5C and D). It is noteworthy that functional studies demonstrate that CLDN6- and CLDN9-specific MAbs had no effect on HCV infection of Huh7.5.1 cells or PHHs (Fig. 3 to 5). Interestingly, silencing of CLDN6 in Huh7.5 cells also did not affect HCV infection (17). Furthermore, these MAbs were unable to further decrease HCVpp entry into CLDN1-silenced Huh7.5.1 cells or when added simultaneously with the CLDN1-specific MAb to naive Huh7.5.1 cells, suggesting that CLDN6 and CLDN9 are not able to complement the absence of CLDN1 expression or blockage by the anti-CLDN1 MAb (Fig. 4). Given their low levels of expression (Fig. 5C and D), other CLDN members may not be able to compensate CLDN1-specific inhibition of HCV entry into PHHs. In contrast, a recent study showed that silencing CLDN6 expression in Huh7.5 cells further increased CLDN1-specific antibody-mediated inhibition of HCV infection of genotype 1b and 3a (17). Differences of CLDN6 expression in Huh7-based cell lines or experimental design may account for the different findings. Given the very low levels of CLDN6 protein expression in the human liver in vivo (Fig. 5) and its absent or limited functional role in human hepatocytes (Fig. 5), the relevance of CLDN6 escape observed with Huh7.5 cells (17) remains to be determined with human hepatocytes or in vivo. If relevant, the CLDN6-specific MAbs (Fig. 1 and 2) could address this issue.
In summary, this study demonstrates that although CLDN6 and CLDN9 can serve as entry factors upon exogenous expression in certain cell culture models, such as CLDN-deficient 293T cells, they do not play a major role for HCV entry into human hepatocytes. Yet given their functional activity, the novel antibodies described in this study may represent interesting tools to investigate the role of CLDN6 and CLDN9 in physiology and disease.
ACKNOWLEDGMENTS
This work was supported by Inserm, University of Strasbourg, the Zentrales Innovationsprogramm Mittelstand, the European Union (ERC-2008-AdG-233130-HEPCENT, INTERREG-IV-Rhin Supérieur-FEDER-Hepato-Regio-Net 2009 and 2012, EU FP7 HEPAMAB), ANRS (2011/132, 2012/239, 2012/318, 2013/108), and Laboratoire d'excellence LabEx HepSYS (Investissement d'Avenir; ANR-10-LAB-28). Research in the McKeating lab is funded by the Medical Research Council and NIHR Liver Biomedical Research Unit.
We thank R. Bartenschlager (University of Heidelberg, Germany), F.-L. Cosset (Inserm U1111, ENS Lyon, France), and J. Ball (University of Nottingham, United Kingdom) for providing plasmids for production of HCVcc and HCVpp, C. M. Rice (The Rockefeller University, New York, NY) and F. V. Chisari (The Scripps Research Institute, La Jolla, CA) for Huh7.5 and Huh7.5.1 cells, respectively, P. Bachellier (Service de Chirurgie Générale, Hépatique, Endocrinienne et Transplantation, Hôpitaux Universitaires de Strasbourg, France) for providing primary human hepatocytes, D. Trono (Ecole Polytechnique Fédérale de Lausanne, Switzerland) for lentiviral expression constructs, and T. Pietschmann (TWINCORE, Centre for Experimental and Clinical Infection Research, Germany) for helpful discussions. The authors acknowledge P. Balfe (University of Birmingham, United Kingdom) for RT-PCR data as well as the excellent technical assistance of S. Glauben (Aldevron GmbH, Germany) and C. Bach (Inserm U1110, France).
T. F. B. designed research. I.F., L.Z., C.T., L.H., S.C.D., J. L., H.E.B., P.P., G.M.R., J.A.M., F.G., J.T., M.B.Z, and T.F.B. performed research. I.F., L.Z., C.T., L.H., S.C.D., J.L., P.P., H.E.B., G.M.R., J.A.M., F.G., J.T., M.B.Z, and T.F.B. analyzed data. C.G. contributed essential reagents. I.F., M.B.Z., and T.F.B. wrote the paper.
The authors declare no conflict of interest. Inserm and Aldevron have previously filed a patent application on the use of anti-Claudin-1 antibodies for inhibition of HCV infection (WO2010034812).
Footnotes
Published ahead of print 17 July 2013
REFERENCES
- 1. Meredith LW, Wilson GK, Fletcher NF, McKeating JA. 2012. Hepatitis C virus entry: beyond receptors. Rev. Med. Virol. 22:182–193 [DOI] [PubMed] [Google Scholar]
- 2. Zeisel MB, Lupberger J, Fofana I, Baumert TF. 2013. Host-targeting agents for prevention and treatment of chronic hepatitis C: perspectives and challenges. J. Hepatol. 58:375–384 [DOI] [PubMed] [Google Scholar]
- 3. Martin DN, Uprichard SL. 2013. Identification of transferrin receptor 1 as a hepatitis C virus entry factor. Proc. Natl. Acad. Sci. U. S. A. 110:10777–10782 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Evans MJ, von Hahn T, Tscherne DM, Syder AJ, Panis M, Wolk B, Hatziioannou T, McKeating JA, Bieniasz PD, Rice CM. 2007. Claudin-1 is a hepatitis C virus co-receptor required for a late step in entry. Nature 446:801–805 [DOI] [PubMed] [Google Scholar]
- 5. Krieger SE, Zeisel MB, Davis C, Thumann C, Harris HJ, Schnober EK, Mee C, Soulier E, Royer C, Lambotin M, Grunert F, Dao Thi VL, Dreux M, Cosset FL, McKeating JA, Schuster C, Baumert TF. 2010. Inhibition of hepatitis C virus infection by anti-claudin-1 antibodies is mediated by neutralization of E2-CD81-claudin-1 associations. Hepatology 51:1144–1157 [DOI] [PubMed] [Google Scholar]
- 6. Fofana I, Krieger SE, Grunert F, Glauben S, Xiao F, Fafi-Kremer S, Soulier E, Royer C, Thumann C, Mee CJ, McKeating JA, Dragic T, Schuster C, Thompson J, Baumert TF. 2010. Monoclonal anti-claudin 1 antibodies for prevention of hepatitis C virus infection. Gastroenterology 139:953–964, 964.e951-954 [DOI] [PubMed] [Google Scholar]
- 7. Harris HJ, Davis C, Mullins JG, Hu K, Goodall M, Farquhar MJ, Mee CJ, McCaffrey K, Young S, Drummer H, Balfe P, McKeating JA. 2010. Claudin association with CD81 defines hepatitis C virus entry. J. Biol. Chem. 285:21092–21102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Harris HJ, Farquhar MJ, Mee CJ, Davis C, Reynolds GM, Jennings A, Hu K, Yuan F, Deng H, Hubscher SG, Han JH, Balfe P, McKeating JA. 2008. CD81 and claudin 1 coreceptor association: role in hepatitis C virus entry. J. Virol. 82:5007–5020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Zona L, Lupberger J, Sidahmed-Adrar N, Thumann C, Harris HJ, Barnes A, Florentin J, Tawar RG, Xiao F, Turek M, Durand SC, Duong FH, Heim MH, Cosset FL, Hirsch I, Samuel D, Brino L, Zeisel MB, Le Naour F, McKeating JA, Baumert TF. 2013. HRas signal transduction promotes hepatitis C virus cell entry by triggering assembly of the host tetraspanin receptor complex. Cell Host Microbe 13:302–313 [DOI] [PubMed] [Google Scholar]
- 10. Zheng A, Yuan F, Li Y, Zhu F, Hou P, Li J, Song X, Ding M, Deng H. 2007. Claudin-6 and claudin-9 function as additional coreceptors for hepatitis C virus. J. Virol. 81:12465–12471 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Meertens L, Bertaux C, Cukierman L, Cormier E, Lavillette D, Cosset FL, Dragic T. 2008. The tight junction proteins claudin-1, -6, and -9 are entry cofactors for hepatitis C virus. J. Virol. 82:3555–3560 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Graham FL, Smiley J, Russell WC, Nairn R. 1977. Characteristics of a human cell line transformed by DNA from human adenovirus type 5. J. Gen. Virol. 36:59–74 [DOI] [PubMed] [Google Scholar]
- 13. Shaw G, Morse S, Ararat M, Graham FL. 2002. Preferential transformation of human neuronal cells by human adenoviruses and the origin of HEK 293 cells. FASEB J. 16:869–871 [DOI] [PubMed] [Google Scholar]
- 14. Zahid MN, Turek M, Xiao F, Dao Thi VL, Guerin M, Fofana I, Bachellier P, Thompson J, Delang L, Neyts J, Bankwitz D, Pietschmann T, Dreux M, Cosset FL, Grunert F, Baumert TF, Zeisel MB. 2013. The postbinding activity of scavenger receptor class B type I mediates initiation of hepatitis C virus infection and viral dissemination. Hepatology 57:492–504 [DOI] [PubMed] [Google Scholar]
- 15. Da Costa D, Turek M, Felmlee DJ, Girardi E, Pfeffer S, Long G, Bartenschlager R, Zeisel MB, Baumert TF. 2012. Reconstitution of the entire hepatitis C virus life cycle in nonhepatic cells. J. Virol. 86:11919–11925 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Fafi-Kremer S, Fofana I, Soulier E, Carolla P, Meuleman P, Leroux-Roels G, Patel AH, Cosset FL, Pessaux P, Doffoël M, Wolf P, Stoll-Keller F, Baumert TF. 2010. Enhanced viral entry and escape from antibody-mediated neutralization are key determinants for hepatitis C virus re-infection in liver transplantation. J. Exp. Med. 207:2019–2031 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Haid S, Grethe C, Dill MT, Heim M, Kaderali L, Pietschmann T. 14 June 2013, posting date Isolate-dependent use of Claudins for cell entry by hepatitis C virus. Hepatology. 10.1002/hep.26567 [DOI] [PubMed] [Google Scholar]
- 18. Fofana I, Xiao F, Thumann C, Turek M, Zona L, Tawar RG, Grunert F, Thompson J, Zeisel MB, Baumert TF. 2013. A novel monoclonal anti-CD81 antibody produced by genetic immunization efficiently inhibits hepatitis C virus cell-cell transmission. PLoS One 8:e64221. 10.1371/journal.pone.0064221 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Bartosch B, Dubuisson J, Cosset FL. 2003. Infectious hepatitis C virus pseudo-particles containing functional E1-E2 envelope protein complexes. J. Exp. Med. 197:633–642 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Koutsoudakis G, Kaul A, Steinmann E, Kallis S, Lohmann V, Pietschmann T, Bartenschlager R. 2006. Characterization of the early steps of hepatitis C virus infection by using luciferase reporter viruses. J. Virol. 80:5308–5320 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Reynolds GM, Harris HJ, Jennings A, Hu K, Grove J, Lalor PF, Adams DH, Balfe P, Hubscher SG, McKeating JA. 2008. Hepatitis C virus receptor expression in normal and diseased liver tissue. Hepatology 47:418–427 [DOI] [PubMed] [Google Scholar]