

Abstract
Kingella kingae is a human pathogen that causes pediatric osteoarticular infections and infective endocarditis in children and adults. The bacterium is usually susceptible to β-lactam antibiotics, although β-lactam resistance has been reported in rare isolates. This study was conducted to identify β-lactam-resistant strains and to characterize the resistance mechanism. Screening of a set of 90 K. kingae clinical isolates obtained from different geographic locations revealed high-level resistance to penicillins among 25% of the strains isolated from Minnesota and Iceland. These strains produced TEM-1 β-lactamase and were shown to contain additional ≥50-kb plasmids. Ion Torrent sequencing of extrachromosomal DNA from a β-lactamase-producing strain confirmed the plasmid location of the blaTEM gene. An identical plasmid pattern was demonstrated by multiplex PCR in all β-lactamase producers. The porin gene's fragments were analyzed to investigate the relatedness of bacterial strains. Phylogenetic analysis revealed 27 single-nucleotide polymorphisms (SNPs) in the por gene fragment, resulting in two major clusters with 11 allele types forming bacterial-strain subclusters. β-Lactamase producers were grouped together based on por genotyping. Our results suggest that the β-lactamase-producing strains likely originate from a single plasmid-bearing K. kingae isolate that traveled from Europe to the United States, or vice versa. This study highlights the prevalence of penicillin resistance among K. kingae strains in some regions and emphasizes the importance of surveillance for antibiotic resistance of the pathogen.
INTRODUCTION
A frequent cause of septic arthritis and osteomyelitis in children younger than 2 years old was recently demonstrated to be infections with Kingella kingae (1–8). The bacterium is also a cardiovascular pathogen, causing severe infective endocarditis in children and adults (4, 9–13).
K. kingae, a Gram-negative coccobacillus of the family Neisseriaceae, is a normal human oropharynx inhabitant and is transmitted from child to child (14). K. kingae osteoarticular infections were underestimated prior to new isolation and PCR identification techniques were developed in the 1990s. Several recent reports describe epidemiological cases of invasive K. kingae infections in day care centers, showing the bacterium's ability to cause outbreaks in pediatric communities (15–17).
β-Lactam antibiotics (penicillin and its derivatives) inhibit the formation of bacterial cell wall peptidoglycan cross-linkages and are widely used in the treatment of bacterial infections, including osteoarticular infections (14). Penicillins are essentially first-choice drugs for infective endocarditis prevention in high-risk populations (18). Different mechanisms of β-lactam resistance have been described among Gram-negative bacteria. They include the production of different types of β-lactamases and are also associated with the natural low membrane permeability and with efflux systems (19, 20).
K. kingae is generally characterized as highly susceptible to β-lactams, although β-lactamase production has been reported in rare isolates (21–23). Currently, the mechanisms of β-lactam resistance in K. kingae are unknown. We screened isolates from different geographic regions for β-lactam susceptibility and characterized plasmid-encoded β-lactamase produced by the organism. We also developed single-gene sequence typing (SGST) of the isolates based on the major outer membrane protein (porin) gene structure to investigate genetic heterogeneity among the strains.
MATERIALS AND METHODS
Bacterial strains.
We acquired a collection of 90 K. kingae clinical isolates from different geographic regions. These strains were predominantly obtained from pediatric patients with infective endocarditis (n = 1), osteomyelitis (n = 8), septic arthritis (n = 27), and bacteremia (n = 28), as well as from healthy carriers (n = 26). Forty-four strains isolated between 1991 and 2010 in Israel were provided by Pablo Yagupsky from the Ben-Gurion University Medical School in southern Israel. Some of these strains were used in previous studies (24–27). Thirty-one strains of U.S. origin, obtained from 2003 to 2012, were contributed by the Minnesota State Health Department. They include strains isolated in an investigation of an outbreak of K. kingae infections in 2003 (28). One U.S. strain was donated by Paul Planet (American Museum of Natural History, New York, NY). Fourteen Icelandic isolates, including strains described previously (23), were collected in the period between 1995 and 2010 and were provided by Hjordis Harðardóttir from the Department of Clinical Microbiology, Landspitali University Hospital (Iceland). Strain 23330 (Norway) was obtained from the ATCC. The bacteria were grown on Columbia agar (CA) with 5% sheep blood at 37°C with 10% CO2 and stored at −80°C in the growth medium with 10% dimethyl sulfoxide (DMSO). The identification of all strains was confirmed by sequence analysis of the 16S rRNA gene. The clinical information on K. kingae isolates utilized in this work is presented in Table S1 in the supplemental material. Other strains used in the study were Kingella oralis 51147 (ATCC), Neisseria perflava 14799 (ATCC), Escherichia coli DH5α (Invitrogen), and Aggregatibacter actinomycetemcomitans 1704 (29).
Antibiotic sensitivity testing.
A Thermo Scientific Remel Nitrocefin Disk was used to identify β-lactamase production. MICs of antibiotics were determined by the agar dilution method according to Clinical and Laboratory Standards Institute (CLSI) guidelines (1, 30). Growth inhibition was identified after incubation of bacteria on Mueller-Hinton agar supplemented with 5% defibrinated sheep blood and containing various concentrations of antibiotics for 24 h at 37°C with 10% CO2.
DNA purification.
Genomic DNA isolation was accomplished using the Wizard Genomic DNA Purification kit (Promega). Plasmid DNA was purified using a Qiagen Plasmid Midi Kit. The DNA concentration was determined using NanoDrop (NanoDrop Technologies, Wilmington, DE). For sequencing, 200 mg of plasmid DNA stained with ethidium bromide was subjected to ultracentrifugation for 20 h at 45,000 rpm in a 1-g/ml cesium chloride solution. The plasmid bands were collected with the needle positioned just below the band of plasmid DNA.
PCR amplification and typing.
We used the primers Bact8F and Bact1391R for the 16S rRNA gene to identify bacterial strains (31). To determine a β-lactamase type, primers for blaTEM, blaSHV (32), and blaCTX-M (33) were used for PCR amplification. The primers developed in this study are listed in Table 1. For plasmid typing, the PCR utilized the Expand High Fidelity PCR System (Roche) in 25 μl containing 1× buffer solution, 0.5 μM each primer, 0.2 mM deoxynucleoside triphosphate (dNTP), and 50 ng plasmid DNA. The amplification reaction was performed on the automated thermal cycler with an initial step of 10 min at 95°C, followed by 30 cycles of 1 min at 95°C, 1 min at 50°C, and 1 min at 72°C, and a final extension step of 10 min at 72°C. For porin gene typing, the reaction mixture contained bacterial genomic DNA as the template. The amplification reaction was performed with an initial step of 10 min at 95°C, followed by 30 cycles of 1 min at 95°C, 1 min at 55°C, and 1 min at 72°C, and a final extension step of 10 min at 72°C. The amplified products were visualized on a 0.8% agarose gel containing ethidium bromide.
Table 1.
Primers designed in the study
| Primer | Sequence | Analyzed product size (bp) |
|---|---|---|
| blaTEM 550 FW | 5′-GAGAATTATGCAGTGCTGCCATAACC-3′ | 550 |
| blaTEM 550 RV | 5′-GTTATCACTCATGGTTATGGCAG-3′ | 550 |
| por FW | 5′-GCACATAAGGAAATGAATAATG-3′ | 956 |
| por RV | 5′-CAAAATTAGAATTTATGGCTCATACCC-3′ | 956 |
| pAGA1 308-759 FW | 5′-GAAGATAAGGCATTAAGCAATTTG-3′ | 451 |
| pAGA1 308-759 RV | 5′-CTCGGCTCATAATCCTACCTG-3′ | 451 |
| pAGA1 4146-4789 FW | 5′-GGCTAAACGAACCACCGAAC-3′ | 643 |
| pAGA1 4146-4789 RV | 5′-CTGCTGCCTGTGTAAATCGTC-3′ | 643 |
| pAGA1 10146-11114 FW | 5′-GCCGAAATTGTCAAAAGTGTAG-3′ | 968 |
| pAGA1 10146-11114 RV | 5′-GCTATCACAAGAAGAAATAC-3′ | 968 |
| pAGA115085-15294 FW | 5′-CCATGAAGATGAAGCATTTC-3′ | 209 |
| pAGA115085-15294 RV | 5′-CTTTCAAGTGGGCTAGGTTG-3′ | 209 |
DNA sequencing.
The nucleotide sequences of the two strands of PCR products were determined by capillary sequencing using an ABI 3130xl Genetic Analyzer (Applied Biosystems, Foster City, CA). For plasmid sequencing, 10 mg of purified plasmid DNA was used for fragment library construction followed by sequencing on an Ion Torrent Platform 314 chip with 200-bp read lengths (Life Technologies). Sequence assembly was performed using Newbler v2.5 software without directed gap filling.
K. kingae mutagenesis.
The RTX toxin-deficient strain KKNB100 was obtained using the mariner transposon derivative Solo, which carries the aphA3 kanamycin resistance gene, using a previously described protocol (34) with modifications. Briefly, the rtxA gene was PCR amplified from genomic DNA of PYKK081 and cloned into the pUC19 vector (New England BioLabs). Then, the aphA3 gene, obtained from plasmid pFALCON2, was inserted in rtxA using the NsiI restriction site. The rtxA gene containing aphA3 was PCR amplified and used for the transformation of PYKK081. To accomplish this, the bacteria were grown overnight on CA with 5% sheep blood and resuspended to an A600 of 0.8 in Trypticase soy broth containing 2% yeast extract. After 30 min of incubation, the cells were resuspended in Trypticase soy broth containing 10 mM CaCl2, 12% horse plasma, and DNA. The transformation mixture was incubated for 1 h at 37°C and plated on CA-5% sheep blood plates containing kanamycin (40 μg/ml). The transconjugants were grown for 48 h at 37°C in a 10% CO2 incubator.
Mating experiments.
Donor and recipient strain cells (107) were mixed in a 1:1 ratio and incubated on CA plates with 5% blood for 18 h. The mixed cell culture was washed from the plates with 1 ml saline solution. Aliquots (0.2 ml) of each culture were plated on selective plates containing penicillin (2 μg/ml). Transconjugant colonies were recovered after 24 h and plated on new selective plates. Plasmid presence was confirmed by multiplex PCR.
Phylogenetic analysis.
Sequences were aligned with ClustalW (European Bioinformatics Institute), and a tree was constructed by applying the unweighted-pair group method using average linkages (UPGMA) and a Jukes and Cantor DNA distance matrix using Geneious 6.1.4 software. Bootstrap analysis was used to estimate the confidence of each group assignment. The tree was bootstrapped 100 times, and the proportion of data sets supporting each tree topology was recorded as a bootstrap probability value.
Nucleotide sequence accession numbers.
The sequence assembly of the K. kingae C2005004457 pAGA1 and pAGA2 plasmid DNA was deposited in GenBank (NCBI) under accession number AMPT00000000. The nucleotide sequence for the K. kingae TEM-1 gene has been submitted to GenBank under accession number JX855253. The nucleotide sequences for the K. kingae porin gene variants have been submitted to GenBank under accession numbers KC142159 to KC142169.
RESULTS
Distribution of β-lactam resistance among K. kingae clinical isolates.
Forty-four Israeli, 1 Norwegian, 14 Icelandic, and 31 U.S. K. kingae isolates were screened for susceptibility to penicillin G (4 μg/ml) and β-lactamase production using the nitrocefin method. Most of the strains, including all the Israeli strains, were negative for β-lactamase production and susceptible to penicillin. Eight U.S. isolates and 4 Icelandic strains were resistant to penicillin G and were β-lactamase producers (see Table S1 in the supplemental material). All penicillin-resistant strains from our collection (n = 12) and 12 random penicillin-susceptible strains were used to determine MICs. Using the agar dilution method, these strains were found to be resistant to penicillin V/G up to ≥250 μg/ml, ampicillin up to ≥200 μg/ml, carbenicillin up to 150 μg/ml, and amoxicillin up to 175 μg/ml, while nonresistant isolates were susceptible to these antibiotics at approximately 1,000 times lower concentrations (Table 2). All strains were susceptible to broad-spectrum cephalosporins (ceftazidime and ceftriaxone) and the carbapenem biapenem. MICs were strikingly similar between isolates from each group.
Table 2.
Antibiotic susceptibility testing
| Antibiotic | MIC (μg/ml)a |
|
|---|---|---|
| TEM+ (n = 12) | TEM− (n = 12) | |
| Penicillin G | ≥250 | 0.20 |
| Penicillin V | ≥250 | 0.20 |
| Ampicillin | ≥200 | 0.15 |
| Carbenicillin | ≥150 | 0.20 |
| Amoxicillin | ≥175 | 0.20 |
| Biapenem | 0.35 | 0.35 |
| Ceftazidime | 0.20 | 0.20 |
| Ceftriaxone | 0.20 | 0.20 |
MICs were identified by the agar dilution method in β-lactamase-producing (TEM+) and non-β-lactamase-producing (TEM−) strains.
Characterization of K. kingae β-lactamase.
The β-lactamase-producing strain C2005004457 of U.S. origin was selected for further analysis. To identify a β-lactamase type, primers for the three most common β-lactamase genes in Gram-negative bacteria, blaTEM, blaSHV, and blaCTX-M, were employed to amplify the product from C2005004457. PCR resulted in amplification of a product of about 1 kb using primers for blaTEM (Fig. 1A). The PCR product containing the gene for K. kingae TEM was sequenced. The flanking sequences of the blaTEM gene were sequenced using outward primers listed in Table 1. Using BLAST search analysis, the sequence was found to contain an 867-bp β-lactamase gene identical to one encoding Neisseria gonorrhoeae TEM-1 (NP_052173.1) (35). The TEM β-lactamase PCR product was obtained from other β-lactamase-positive strains. Comparative sequence analysis showed no variations in the blaTEM sequence between the K. kingae β-lactamase producers.
Fig 1.
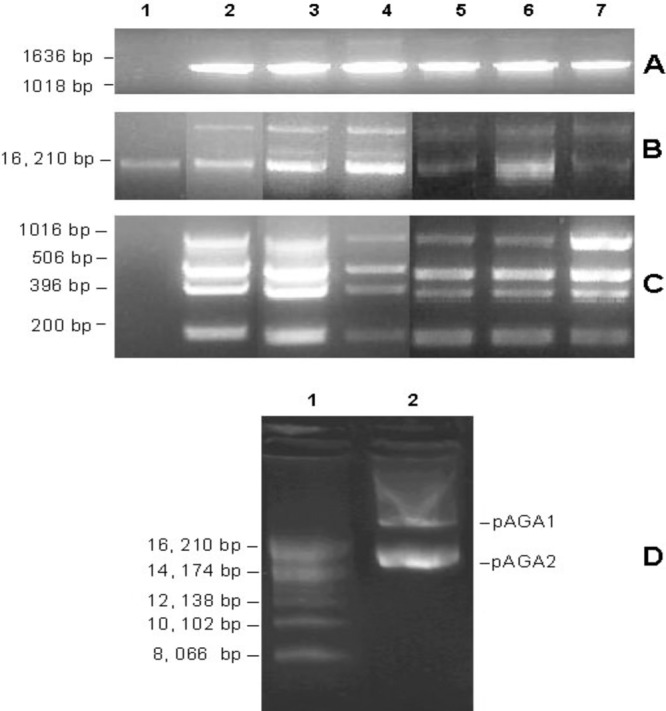
Plasmid DNA analysis in K. kingae strains. (A to C) The blaTEM gene amplified from K. kingae DNA (A), plasmid DNA identified in K. kingae strains (B), and multiplex PCR plasmid typing (C). Lanes: 1, PYKK081 (Israel; 1991); 2, C2005004457 (United States; 2005); 3, C2007000490 (United States; 2007); 4, M2004000037 (United States; 2004); 5, 0303 and 28260 (Iceland; 2003); 6, 0405 and 30002 (Iceland; 2004); 7, 9508 and 31135 (Iceland; 1995). (D) Plasmid DNA purified using a cesium chloride gradient. Lanes: 1, supercoiled DNA ladder (Invitrogen); 2, plasmid DNA purified from strain C2005004457.
K. kingae plasmid analysis.
To identify the locations of genes in the bacterial genome, plasmid DNA was isolated from all β-lactamase-producing strains and 3 random nonproducing strains (PYKK081, C2003003154, and M2003000170) (see Table S1 in the supplemental material). All tested K. kingae isolates contained a plasmid band of ∼15 kb. β-Lactamase-producing strains also contained a second, larger plasmid. Selected strains are shown in Fig. 1B. The small and large plasmids from strain C2005004457 were designated pAGA1 and pAGA2, respectively.
C2005004457 plasmid DNA sequencing.
To purify circular plasmid DNA from linear chromosomal DNA, the plasmid sample from strain C2005004457, obtained by a modified alkaline lysis procedure, was subjected to ultracentrifugal separation in a cesium chloride gradient (Fig. 1D). The pAGA1 and pAGA2 DNA bands were not clearly separated in the gradient, and therefore, total “plasmid bands” of DNA were isolated and used for fragment library preparation, followed by Torrent sequencing. The Torrent run yielded 136,258 individual reads with a mean read length of 209 bases. The total number of bases read was 28,593,075 (ca. 433-fold coverage). The completed extrachromosomal DNA sequence consisted of 65,896 bp with an average G+C content of 39%. The size of pAGA2 was previously identified as approximately 15 kb by using plasmid markers. Therefore, given the total size of the bacterial-plasmid sequence, the size of the other plasmid, pAGA1, was estimated to be approximately 50 kb. BLAST search analysis did not reveal any extensive regions of similarity between the newly generated C2005004457 plasmids' sequences and other known plasmid sequences. In the final assembly, 94.8% of the reads aligned into 127 unique contigs, but only 65 with gene calls and of sufficient length (≥200 bp) were selected for inclusion in GenBank. The contigs could not be assigned to either pAGA1 or pAGA2 in the present sequence state. The sequence was annotated using the NCBI Prokaryotic Genomes Automatic Annotation Pipeline server at NCBI (http://www.ncbi.nlm.nih.gov/genomes/static/Pipeline.html). There were 110 predicted protein-coding genes, and of these, 68 (62%) had functional predictions. Among those with functional predictions, 7 were associated with resistance to antibiotics (they are listed in Table 3) and 16 with phage-associated virulence. The blaTEM gene was localized in the sequence, suggesting that TEM-1 is plasmid encoded.
Table 3.
C2005004457 plasmid-encoded products associated with antibiotic resistance
| Gene product | Drug |
|---|---|
| Aminoglycoside/hydroxyurea antibiotic resistance kinase | Aminoglycosides/hydroxyurea |
| Aminoglycoside phosphotransferase | Aminoglycosides |
| Beta-lactamase class A; EC 3.5.2.6 | Penicillins |
| Streptomycin 6-kinase | Streptomycin |
| Streptomycin phosphotransferase | Streptomycin |
| Dihydropteroate synthase | Sulfonamides |
| Antitoxin of toxin-antitoxin stability | Antibiotic resistancea |
This gene product may provide resistance to many antibiotics.
Plasmid typing.
A multiplex PCR approach was developed to identify whether plasmids found in β-lactamase-producing strains are genetically variable. Using the C2005004457 plasmid DNA sequence, we designed a set of primers to amplify four 200- to 1,000-bp PCR products from random parts of the pAGA1 sequence (Table 1). Multiplex PCR resulted in a uniform PCR fragment pattern obtained from all β-lactamase-producing strains. Selected strains are shown in Fig. 1C. This result indicates that all β-lactam-resistant strains contain the same pAGA1 plasmid.
Plasmid transfer.
To investigate the possibility of R-plasmid horizontal transfer, mating experiments were carried out using different recipient strains sensitive to penicillin with C2005004457 as a donor (Table 4). We needed a K. kingae recipient strain to perform transformation and mating experiments on newly discovered plasmids. Strain KKNB100, carrying the aphA3 kanamycin resistance gene, was available in our laboratory for such studies. This strain lacks RtxA toxin activity and is not toxic to mammalian leukocytes. This feature was used as an additional phenotypic marker to distinguish between donor and recipient strains in mating experiments. When K. kingae KKNB100 was used as a recipient, the transconjugants formed on selective plates at high frequency and contained pAGA1, as demonstrated by multiplex PCR. However, further passages (less than four passages) on both nonselective and selective plates led to the loss of the strains' ability to produce β-lactamase and retain the plasmid. No spontaneous mutants were obtained on selective medium that contained two antibiotics, kanamycin (40 μg/ml) and penicillin (2 μg/ml), when a donor or recipient was plated alone. We conclude that pAGA1 is transmissible by conjugation to other K. kingae strains; however, there are special requirements for the cell to support the plasmid. In addition, we performed mating experiments using other bacterial species: E. coli DH5α, N. perflava, and A. actinomycetemcomitans 1704. Similar to KKNB100, transconjugants were formed in E. coli DH5α and N. perflava; however, the following passage led to the loss of the plasmid. We did not observe the formation of transconjugants in A. actinomycetemcomitans 1704.
Table 4.
Mating experiments between C2005004457 and Gram-negative bacteriaa
| Recipient strain | Selective mediumb | Transconjugantsc |
|---|---|---|
| K. kingae KKNB100 | CA-5% blood, KAN 40, PCN 2 | + |
| E. coli DH5α | LA, PCN 2 | + |
| N. perflava 14799 | MHA, PCN 2 | + |
| A. actinomycetemcomitans 1704 | CA-5% blood, KAN 40, PCN 2 | − |
C2005004457 and recipient strain cells (107) were mixed in a 1:1 ratio and incubated on CA with 5% blood for 18 h. Aliquots of the mixed cell culture were plated on selective plates containing penicillin G (2 μg/ml). Transconjugant colonies were recovered after 24 h.
KAN 40, kanamycin (40 μg/ml); PCN 2, penicillin (2 μg/ml). Luria-Bertani agar (LA) and Mueller-Hinton agar (MHA) media without blood did not support C2005004457 growth.
+, present; −, absent.
SGST sequence analysis.
To better understand the population structure of circulating K. kingae isolates and to identify the relatedness of β-lactamase producers, we classified the isolates based on porin (por) gene structure. Porins are essential pore-forming membrane proteins that create channels involved in hydrophilic-molecule transport (36). The K. kingae porin gene nucleotide sequence (1,053 bp) shares only 10% identity with Neisseria meningitidis and N. gonorrhoeae porin sequences. Comparative genome analysis of fully sequenced genomes of ATCC 23330 (GenBank accession number AFHS01000000) and PYKK081 (GenBank accession number AJGB01000000) revealed a number of polymorphic regions in the K. kingae porin gene, allowing distinction among strain groups. The genotyping was achieved by comparing 956-bp fragments of the por gene obtained from 90 isolates. Sequence analysis of the por gene fragment revealed 27 polymorphic nucleotides. Evaluation of sequence variations led to identification of two major K. kingae groups of strains with 11 por allele types. Seven por allele types were found in multiple patients, and 4 types were unique. The most predominant were por allele types 1, 9, and 2, which comprised 68% of all isolates. The genetic relatedness of por sequences is shown in a dendrogram (Fig. 2). Based on the por gene allele typing, β-lactamase production of U.S. and Icelandic isolates is limited to organisms exhibiting the por type 1 allele.
Fig 2.

Phylogeny of Kingella strains based on the 956-bp por gene sequence. Strains that produce β-lactamase are underlined. Strains isolated from outbreak patients in Minnesota are indicated with asterisks. The numbers on the right indicate the 11 allele types discussed in the text. The numbers within the dendrogram indicate the occurrence (percent) of the branching in 100 bootstrapped trees. K. oralis por was used as the outgroup.
DISCUSSION
K. kingae is generally known to be highly susceptible to β-lactams, and infections with the bacterium can be successfully treated by antibiotic therapy with penicillins and cephalosporins (14). The first β-lactamase-producing strain was reported in the United States in 1993 (21), followed by β-lactam-resistant isolates in Iceland in 1997 (23). In our collection, the β-lactamase-producing strains of U.S. origin were isolated in Minnesota from 2004 to 2007, and the Icelandic strains were isolated in different periods from 1995 to 2004. Interestingly, whereas the largest number of strains in our collection were obtained from Israel (n = 44), none of the Israeli strains were found to be resistant to β-lactams, which is in good agreement with previous studies (24, 37). These data may suggest the relative prevalence and persistence of such isolates in some regions.
Here, we report for the first time the production of plasmid-mediated TEM-1 in K. kingae. TEM-1 is a β-lactamase commonly found in Gram-negative bacteria. The enzyme has been reported in E. coli (38), Haemophilus influenzae (39), N. meningitidis (40), and N. gonorrhoeae (41). The native TEM-1 β-lactamase confers high-level resistance to ampicillin, penicillin, and some first-generation cephalosporins. However, mutations within the blaTEM structural gene may allow the enzyme to expand the hydrolysis capabilities to cephalosporins with an oxyimino side chain (42). Infections caused by the extended-spectrum β-lactamase (ESBL) have been associated with severe adverse clinical outcomes that have led to increased mortality, prolonged hospitalization, and additional medical costs (43–45).
Although K. kingae is being increasingly recognized as an important human pathogen, progress in understanding K. kingae genetics was made only recently, with the genomes of strains ATCC 23330, PYKK081 (46), and 11220434 (47) released. We have demonstrated that K. kingae represents a distinct taxonomic group in the family Neisseriaceae (46). Here, we report the unique R-plasmid pAGA1 in the organism. The identification and sequencing of new K. kingae plasmids will add to our understanding of the organism's genetic system. Plasmid typing, proposed here, can be applied to investigate Kingella plasmid diversity.
Pulsed-field gel electrophoresis (PFGE) (26, 27) and multilocus sequence typing (MLST) have been proposed for K. kingae genotyping (25). Fragments of the K. kingae RTX toxin gene, a putative virulence factor, have been analyzed (25, 26). These methods provide a good foundation to investigate evolutionary relationships among strains; however, each has some limitations. Specifically, PFGE techniques are known to be difficult to reproduce between laboratories. MLST is a laborious and expensive technique for bacterial typing and may not provide sufficient information about small clonal genetic variations. In most strains, the rtxA sequencing results were congruent with the allocation to sequence types by MLST and PFGE, but incongruence was observed for several alleles, and the gene was shown to have larger variations in some strains, such as 33-bp duplications (25, 26).
SGST is often used for finer discrimination than MLST, with more variable genes chosen for comparison. Often, these genes correspond to serotyping antigens, for example, porA in meningococci (48, 49) and spa in Staphylococcus aureus (50). Variations in the porin gene structure were used for genotyping other members of the family Neisseriaceae: N. meningitides, N. gonorrhoeae, and Neisseria lactamica (48, 51, 52). The porin (PorB) of N. gonorrhoeae has been implicated in the species' pathogenesis (53). The por gene genotyping proposed here can be used in addition to other typing methods as a molecular epidemiological tool for examination of genetic relationships among circulating K. kingae strains. In this study, the utility of this approach was demonstrated, revealing the relatedness of the β-lactam-resistant isolates. Notably, we have identified the K. kingae porin as a highly immunogenic protein (54). Future studies may find some correlation between the por allele type and the strain serotype. β-Lactamase production was exclusively associated with por allele 1 in K. kingae isolates from the United States and Iceland, suggesting similarity between the isolates from the two countries, whereas none of the 12 Israeli organisms exhibiting por allele 1 was an enzyme producer. Interestingly, 9 of these 12 isolates belonged to PFGE type K and two to the closely related PFGE types ua and E, indicating they belonged to the same subpopulation (26). Clone K is significantly overrepresented among invasive isolates in Israel, causing 28.2% of all invasive K. kingae infections (26) among the Jewish population (27), and members are also commonly isolated in French patients (25). Clone K, also characterized by MLST and rtxA gene sequencing, shows congruency between the different genotyping results. All clone K organisms belong to MLST sequence type 6 and to the closely related rtxA alleles 8, 9, and 10.
In 2003, 2 confirmed cases of K. kingae osteomyelitis/septic arthritis occurred among children in the same toddler classroom of a Minnesota child care center (28). According to por genotyping, both outbreak strains belong to por allele 9 and are not β-lactamase producers. The two strains had indistinguishable PFGE patterns (23), suggesting correlation between different typing techniques. The por allele 9 type was the predominant group among mucosal and invasive K. kingae isolates from Minnesota. This group was associated with septic arthritis and bacteremia in different geographic regions, indicating that it is truly an international invasive clone.
Since no vaccines for K. kingae are available, antimicrobial therapy is the primary measure for infection control. Our results should alert clinicians to the need for β-lactam resistance testing in suspected K. kingae infections. In addition, a recent study reported reduced susceptibility to clindamycin and oxacillin in a substantial number of organisms (14, 24). Taken together, these data emphasize the importance of surveillance for antibiotic resistance of the pathogen.
Supplementary Material
ACKNOWLEDGMENTS
We thank Pablo Yagupsky, Paul Planet, Hjördís Harðardóttir, and the Minnesota Department of Health Infectious Disease Laboratory for contributing bacterial strains, as well as Joseph St. Geme III for providing the mariner transposon mutagenesis system. We thank Ashlee N. Geesey, Anthony Leonetti, and Rachel Thomas for performing genotyping. The plasmid-sequencing service was provided by SeqWright Inc.
This work was supported in part by NIH grants AI82392 (to J.B.K.) and AI80844 (to N.V.B.) and by American Heart Association grant 9SDG2310194 (to N.V.B.).
Footnotes
Published ahead of print 24 June 2013
Supplemental material for this article may be found at http://dx.doi.org/10.1128/AAC.00318-13.
REFERENCES
- 1.Ceroni D, Cherkaoui A, Kaelin A, Schrenzel J. 2010. Kingella kingae spondylodiscitis in young children: toward a new approach for bacteriological investigations? A preliminary report. J. Child Orthop. 4:173–175 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Chometon S, Benito Y, Chaker M, Boisset S, Ploton C, Berard J, Vandenesch F, Freydiere AM. 2007. Specific real-time polymerase chain reaction places Kingella kingae as the most common cause of osteoarticular infections in young children. Pediatr. Infect. Dis. J. 26:377–381 [DOI] [PubMed] [Google Scholar]
- 3.Ilharreborde B, Bidet P, Lorrot M, Even J, Mariani-Kurkdjian P, Liguori S, Vitoux C, Lefevre Y, Doit C, Fitoussi F, Pennecot G, Bingen E, Mazda K, Bonacorsi S. 2009. New real-time PCR-based method for Kingella kingae DNA detection: application to samples collected from 89 children with acute arthritis. J. Clin. Microbiol. 47:1837–1841 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Korach A, Olshtain-Pops K, Schwartz D, Moses A. 2009. Kingella kingae prosthetic valve endocarditis complicated by a paravalvular abscess. Isr. Med. Assoc. J. 11:251–253 [PubMed] [Google Scholar]
- 5.Moumile K, Merckx J, Glorion C, Berche P, Ferroni A. 2003. Osteoarticular infections caused by Kingella kingae in children: contribution of polymerase chain reaction to the microbiologic diagnosis. Pediatr. Infect. Dis. J. 22:837–839 [DOI] [PubMed] [Google Scholar]
- 6.Nguyen S, Fayad G, Modine T, Leroy O. 2009. Mitral acute bacterial endocarditis caused by HACEK microorganisms. J. Heart Valve Dis. 18:353–354 [PubMed] [Google Scholar]
- 7.Verdier I, Gayet-Ageron A, Ploton C, Taylor P, Benito Y, Freydiere AM, Chotel F, Berard J, Vanhems P, Vandenesch F. 2005. Contribution of a broad range polymerase chain reaction to the diagnosis of osteoarticular infections caused by Kingella kingae: description of twenty-four recent pediatric diagnoses. Pediatr. Infect. Dis. J. 24:692–696 [DOI] [PubMed] [Google Scholar]
- 8.Yagupsky P, Dagan R, Prajgrod F, Merires M. 1995. Respiratory carriage of Kingella kingae among healthy children. Pediatr. Infect. Dis. J. 14:673–678 [DOI] [PubMed] [Google Scholar]
- 9.Verbruggen AM, Hauglustaine D, Schildermans F, van der Hauwaert L, Rombouts JJ, Wauters G, Vandepitte J. 1986. Infections caused by Kingella kingae: report of four cases and review. J. Infect. 13:133–142 [DOI] [PubMed] [Google Scholar]
- 10.Lewis MB, Bamford JM. 2000. Global aphasia without hemiparesis secondary to Kingella kingae endocarditis. Arch. Neurol. 57:1774–1775 [DOI] [PubMed] [Google Scholar]
- 11.Goutzmanis JJ, Gonis G, Gilbert GL. 1991. Kingella kingae infection in children: ten cases and a review of the literature. Pediatr. Infect. Dis. J. 10:677–683 [PubMed] [Google Scholar]
- 12.Elyes B, Mehdi G, Kamel BH, Hela Z, Imen BS. 2006. Kingella kingae septic arthritis with endocarditis in an adult. Joint Bone Spine 73:472–473 [DOI] [PubMed] [Google Scholar]
- 13.Brachlow A, Chatterjee A, Stamato T. 2004. Endocarditis due to Kingella kingae: a patient report. Clin. Pediatr. 43:283–286 [DOI] [PubMed] [Google Scholar]
- 14.Yagupsky P, Porsch E, St Geme JW., III 2011. Kingella kingae: an emerging pathogen in young children. Pediatrics 127:557–565 [DOI] [PubMed] [Google Scholar]
- 15.Yagupsky P, Erlich Y, Ariela S, Trefler R, Porat N. 2006. Outbreak of Kingella kingae skeletal system infections in children in daycare. Pediatr. Infect. Dis. J. 25:526–532 [DOI] [PubMed] [Google Scholar]
- 16.Sena AC, Seed P, Nicholson B, Joyce M, Cunningham CK. 2010. Kingella kingae endocarditis and a cluster investigation among daycare attendees. Pediatr. Infect. Dis. J. 29:86–88 [DOI] [PubMed] [Google Scholar]
- 17.Kraig E, Dailey T, Kolodrubetz D. 1990. Nucleotide sequence of the leukotoxin gene from Actinobacillus actinomycetemcomitans: homology to the alpha-hemolysin/leukotoxin gene family. Infect. Immun. 58:920–929 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Wilson W, Taubert KA, Gewitz M, Lockhart PB, Baddour LM, Levison M, Bolger A, Cabell CH, Takahashi M, Baltimore RS, Newburger JW, Strom BL, Tani LY, Gerber M, Bonow RO, Pallasch T, Shulman ST, Rowley AH, Burns JC, Ferrieri P, Gardner T, Goff D, Durack DT. 2007. Prevention of infective endocarditis: guidelines from the American Heart Association: a guideline from the American Heart Association Rheumatic Fever, Endocarditis, and Kawasaki Disease Committee, Council on Cardiovascular Disease in the Young, and the Council on Clinical Cardiology, Council on Cardiovascular Surgery and Anesthesia, and the Quality of Care and Outcomes Research Interdisciplinary Working Group. Circulation 116:1736–1754 [DOI] [PubMed] [Google Scholar]
- 19.Poole K. 2004. Resistance to beta-lactam antibiotics. Cell. Mol. Life Sci. 61:2200–2223 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Poirel L, Bonnin RA, Nordmann P. 2011. Genetic basis of antibiotic resistance in pathogenic Acinetobacter species. IUBMB Life 63:1061–1067 [DOI] [PubMed] [Google Scholar]
- 21.Sordillo EM, Rendel M, Sood R, Belinfanti J, Murray O, Brook D. 1993. Septicemia due to beta-lactamase-positive Kingella kingae. Clin. Infect. Dis. 17:818–819 [DOI] [PubMed] [Google Scholar]
- 22.Jensen KT, Schonheyder H, Thomsen VF. 1994. In-vitro activity of beta-lactam and other antimicrobial agents against Kingella kingae. J. Antimicrob. Chemother. 33:635–640 [DOI] [PubMed] [Google Scholar]
- 23.Birgisson H, Steingrimsson O, Gudnason T. 1997. Kingella kingae infections in paediatric patients: 5 cases of septic arthritis, osteomyelitis and bacteraemia. Scand. J. Infect. Dis. 29:495–498 [DOI] [PubMed] [Google Scholar]
- 24.Yagupsky P. 2012. Antibiotic susceptibility of Kingella kingae isolates from children with skeletal system infections. Pediatr. Infect. Dis. J. 31:212. [DOI] [PubMed] [Google Scholar]
- 25.Basmaci R, Yagupsky P, Ilharreborde B, Guyot K, Porat N, Chomton M, Thiberge JM, Mazda K, Bingen E, Bonacorsi S, Bidet P. 2012. Multilocus sequence typing and rtxA toxin gene sequencing analysis of Kingella kingae isolates demonstrates genetic diversity and international clones. PLoS One 7:e38078. 10.1371/journal.pone.0038078 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Amit U, Porat N, Basmaci R, Bidet P, Bonacorsi S, Dagan R, Yagupsky P. 2012. Genotyping of invasive Kingella kingae isolates reveals predominant clones and association with specific clinical syndromes. Clin. Infect. Dis. 55:1074–1079 [DOI] [PubMed] [Google Scholar]
- 27.Amit U, Dagan R, Porat N, Trefler R, Yagupsky P. 2012. Epidemiology of invasive Kingella kingae infections in 2 distinct pediatric populations cohabiting in one geographic area. Pediatr. Infect. Dis. J. 31:415–417 [DOI] [PubMed] [Google Scholar]
- 28.Kiang KM, Ogunmodede F, Juni BA, Boxrud DJ, Glennen A, Bartkus JM, Cebelinski EA, Harriman K, Koop S, Faville R, Danila R, Lynfield R. 2005. Outbreak of osteomyelitis/septic arthritis caused by Kingella kingae among child care center attendees. Pediatrics 116:e206–213. 10.1542/peds.2004-2051 [DOI] [PubMed] [Google Scholar]
- 29.Balashova NV, Crosby JA, Al Ghofaily L, Kachlany SC. 2006. Leukotoxin confers beta-hemolytic activity to Actinobacillus actinomycetemcomitans. Infect. Immun. 74:2015–2021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Clinical and Laboratory Standards Institute Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically; approved standard. Document M07-A9, 9th ed, vol 32 Clinical and Laboratory Standards Institute, Wayne, PA [Google Scholar]
- 31.Palmer C, Bik EM, Eisen MB, Eckburg PB, Sana TR, Wolber PK, Relman DA, Brown PO. 2006. Rapid quantitative profiling of complex microbial populations. Nucleic Acids Res. 34:e5. 10.1093/nar/gnj007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Nuesch-Inderbinen MT, Hachler H, Kayser FH. 1996. Detection of genes coding for extended-spectrum SHV beta-lactamases in clinical isolates by a molecular genetic method, and comparison with the E test. Eur. J. Clin. Microbiol. Infect. Dis. 15:398–402 [DOI] [PubMed] [Google Scholar]
- 33.Edelstein M, Pimkin M, Palagin I, Edelstein I, Stratchounski L. 2003. Prevalence and molecular epidemiology of CTX-M extended-spectrum beta-lactamase-producing Escherichia coli and Klebsiella pneumoniae in Russian hospitals. Antimicrob. Agents Chemother. 47:3724–3732 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Kehl-Fie TE, St Geme JW., III 2007. Identification and characterization of an RTX toxin in the emerging pathogen Kingella kingae. J. Bacteriol. 189:430–436 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Pagotto F, Aman AT, Ng LK, Yeung KH, Brett M, Dillon JA. 2000. Sequence analysis of the family of penicillinase-producing plasmids of Neisseria gonorrhoeae. Plasmid 43:24–34 [DOI] [PubMed] [Google Scholar]
- 36.Olesky M, Zhao S, Rosenberg RL, Nicholas RA. 2006. Porin-mediated antibiotic resistance in Neisseria gonorrhoeae: ion, solute, and antibiotic permeation through PIB proteins with penB mutations. J. Bacteriol. 188:2300–2308 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Yagupsky P, Katz O, Peled N. 2001. Antibiotic susceptibility of Kingella kingae isolates from respiratory carriers and patients with invasive infections. J. Antimicrob. Chemother. 47:191–193 [DOI] [PubMed] [Google Scholar]
- 38.Livermore DM. 1995. β-Lactamases in laboratory and clinical resistance. Clin. Microbiol. Rev. 8:557–584 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Farrell DJ, Morrissey I, Bakker S, Buckridge S, Felmingham D. 2005. Global distribution of TEM-1 and ROB-1 beta-lactamases in Haemophilus influenzae. J. Antimicrob. Chemother. 56:773–776 [DOI] [PubMed] [Google Scholar]
- 40.Backman A, Orvelid P, Vazquez JA, Skold O, Olcen P. 2000. Complete sequence of a beta-lactamase-encoding plasmid in Neisseria meningitidis. Antimicrob. Agents Chemother. 44:210–212 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Hermida M, Roy C, Baro MT, Reig R, Tirado M. 1993. Characterization of penicillinase-producing strains of Neisseria gonorrhoeae. Eur. J. Clin. Microbiol. Infect. Dis. 12:45–48 [DOI] [PubMed] [Google Scholar]
- 42.Bush K, Jacoby GA. 2010. Updated functional classification of beta-lactamases. Antimicrob. Agents Chemother. 54:969–976 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Tumbarello M, Sanguinetti M, Montuori E, Trecarichi EM, Posteraro B, Fiori B, Citton R, D'Inzeo T, Fadda G, Cauda R, Spanu T. 2007. Predictors of mortality in patients with bloodstream infections caused by extended-spectrum-beta-lactamase-producing Enterobacteriaceae: importance of inadequate initial antimicrobial treatment. Antimicrob. Agents Chemother. 51:1987–1994 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Rodriguez-Bano J, Picon E, Gijon P, Hernandez JR, Cisneros JM, Pena C, Almela M, Almirante B, Grill F, Colomina J, Molinos S, Oliver A, Fernandez-Mazarrasa C, Navarro G, Coloma A, Lopez-Cerero L, Pascual A. 2010. Risk factors and prognosis of nosocomial bloodstream infections caused by extended-spectrum-beta-lactamase-producing Escherichia coli. J. Clin. Microbiol. 48:1726–1731 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Ikeda Y, Mamiya T, Nishiyama H, Koseki T, Mouri A, Nabeshima T. 2012. Risk factors for extended-spectrum beta-lactamase-producing Escherichia coli infection in hospitalized patients. Nagoya J. Med. Sci. 74:105–114 [PMC free article] [PubMed] [Google Scholar]
- 46.Kaplan JB, Lo C, Xie G, Johnson SL, Chain PS, Donnelly R, Kachlany SC, Balashova NV. 2012. Genome sequence of Kingella kingae septic arthritis isolate PYKK081. J. Bacteriol. 194:3017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Fournier PE, Rouli L, El Karkouri K, Nguyen TT, Yagupsky P, Raoult D. 2012. Genomic comparison of Kingella kingae strains. J. Bacteriol. 194:5972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Urwin R, Kaczmarski EB, Guiver M, Fox AJ, Maiden MC. 1998. Amplification of the meningococcal porB gene for non-culture serotype characterization. Epidemiol. Infect. 120:257–262 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Urwin R, Fox AJ, Musilek M, Kriz P, Maiden MC. 1998. Heterogeneity of the PorB protein in serotype 22 Neisseria meningitidis. J. Clin. Microbiol. 36:3680–3682 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Cai Y, Kong F, Wang Q, Tong Z, Sintchenko V, Zeng X, Gilbert GL. 2007. Comparison of single- and multilocus sequence typing and toxin gene profiling for characterization of methicillin-resistant Staphylococcus aureus. J. Clin. Microbiol. 45:3302–3308 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.McKnew DL, Lynn F, Zenilman JM, Bash MC. 2003. Porin variation among clinical isolates of Neisseria gonorrhoeae over a 10-year period, as determined by Por variable region typing. J. Infect. Dis. 187:1213–1222 [DOI] [PubMed] [Google Scholar]
- 52.Bennett JS, Callaghan MJ, Derrick JP, Maiden MC. 2008. Variation in the Neisseria lactamica porin, and its relationship to meningococcal PorB. Microbiology 154:1525–1534 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Mosleh IM, Huber LA, Steinlein P, Pasquali C, Gunther D, Meyer TF. 1998. Neisseria gonorrhoeae porin modulates phagosome maturation. J. Biol. Chem. 273:35332–35338 [DOI] [PubMed] [Google Scholar]
- 54.Maldonado R, Wei R, Kachlany SC, Kazi M, Balashova NV. 2011. Cytotoxic effects of Kingella kingae outer membrane vesicles on human cells. Microb. Pathog. 51:22–30 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.