

Abstract
Localized Fgf10 expression in the distal mesenchyme adjacent to sites of lung bud formation has long been thought to drive stereotypic branching morphogenesis even though isolated lung epithelium branches in the presence of non-directional exogenous Fgf10 in Matrigel. Here, we show that lung agenesis in Fgf10 knockout mice can be rescued by ubiquitous overexpression of Fgf10, indicating that precisely localized Fgf10 expression is not required for lung branching morphogenesis in vivo. Fgf10 expression in the mesenchyme itself is regulated by Wnt signaling. Nevertheless, we found that during lung initiation simultaneous overexpression of Fgf10 is not sufficient to rescue the absence of primary lung field specification in embryos overexpressing Dkk1, a secreted inhibitor of Wnt signaling. However, after lung initiation, simultaneous overexpression of Fgf10 in lungs overexpressing Dkk1 is able to rescue defects in branching and proximal-distal differentiation. We also show that Fgf10 prevents the differentiation of distal epithelial progenitors into Sox2-expressing airway epithelial cells in part by activating epithelial β-catenin signaling, which negatively regulates Sox2 expression. As such, these findings support a model in which the main function of Fgf10 during lung development is to regulate proximal-distal differentiation. As the lung buds grow out, proximal epithelial cells become further and further displaced from the distal source of Fgf10 and differentiate into bronchial epithelial cells. Interestingly, our data presented here show that once epithelial cells are committed to the Sox2-positive airway epithelial cell fate, Fgf10 prevents ciliated cell differentiation and promotes basal cell differentiation.
Keywords: Basal cells, Branching, Dkk1, Fgf10, Lung development, Wnt signaling, Mouse
INTRODUCTION
Lung development in the mouse initiates at embryonic day (E)9-9.5 with the specification of the primary lung field and formation of lung buds, an outpocketing of endodermal respiratory progenitor cells from the ventral wall of the anterior foregut. Nkx2.1 is one of the earliest markers of the lung endoderm and is expressed in the ventral foregut at E9.5 (Lazzaro et al., 1991; Kimura et al., 1996; Minoo et al., 1999). Wnt2a/b signaling in the foregut endoderm is required for Nkx2.1 expression and maintenance of the respiratory fate (Monkley et al., 1996; Zakin et al., 1998; Goss et al., 2009; Harris-Johnson et al., 2009). Inactivation of Ctnnb1 (which encodes β-catenin) in endoderm, or inactivation of Wnt2a and Wnt2b, which are expressed in the mesoderm, results in complete absence of lung and trachea development (Goss et al., 2009; Harris-Johnson et al., 2009). An important role for Wnt2a/b is to regulate Fgf10 expression in the mesenchyme adjacent to the sites of lung bud formation (De Langhe et al., 2008; Yin et al., 2008; Goss et al., 2009; Goss et al., 2011). Interestingly, although knockout mice for Fgf10 (Fgf10-/-) or its receptor Fgfr2b (Fgfr2b-/-) do not develop a lung, these mice do not exhibit a defect in the initial specification of the primary lung field as the trachea does develop in the absence of Fgf10 signaling (Bellusci et al., 1997; Min et al., 1998; Sekine et al., 1999; De Moerlooze et al., 2000; Weaver et al., 2000; Abler et al., 2009). However, following formation of the primary lung buds, Fgf10 is absolutely required for their initial outgrowth and survival of lung epithelial progenitors.
Retinoic acid (RA) signaling is also essential for lung bud formation (Desai et al., 2004; Desai et al., 2006) by indirectly regulating Fgf10 expression through Wnt and Tgfβ signaling pathways. The Wnt antagonist Dkk1 is expressed throughout the foregut, but is excluded from the primary lung fields where RA signaling functions to suppress Dkk1 expression. This allows for increased Wnt2a/b signaling and upregulation of Fgf10 expression (Chen et al., 2007; Chen et al., 2010). RA signaling also induces Fgf10 expression by suppressing Tgfβ signaling in the lung mesenchyme (Chen et al., 2010).
Following primary lung bud outgrowth, lung development ensues by branching laterally and distally in a highly reproducible and repetitive pattern (Metzger et al., 2008). The specific molecules that pattern branching have not been identified. Fgf10 is dynamically localized in the mesenchyme surrounding prospective epithelial buds and has been shown in vitro to act as a chemoattractant on nearby epithelial cells (Bellusci et al., 1997; Park et al., 1998; Weaver et al., 2000). These data have led to the current prevailing theory that directional bud outgrowth is dependent on the precisely localized expression of Fgf10 in mesenchyme distal to the branch point, which regulates the formation of the initial domain branches and all subsequent branches (Bellusci et al., 1997; Warburton, 2008; Ornitz and Yin, 2012).
Fgf10 acts on the distal lung epithelium via its receptor Fgfr2b, which activates β-catenin signaling and prevents the distal epithelial progenitors from differentiating into airway epithelial cells by initially inhibiting Sox2 expression (Park et al., 1998; Que et al., 2007; Ramasamy et al., 2007; Nyeng et al., 2008; Hashimoto et al., 2012). β-Catenin is not only a downstream transcriptional target of epithelial Fgf10 signaling (Lü et al., 2005), but increasing data also indicate that Fgf10 is able to increase nuclear β-catenin directly, via phosphorylation of β-catenin on Ser552 and inhibition of Gsk3β, through the PI3K/AKT pathway (He et al., 2007; Ramasamy et al., 2007; Volckaert et al., 2011). In addition, FGF signaling via Erk/MAPK phosphorylates the Wnt co-receptor Lrp6 on Ser1490 and Thr1572 and phosphorylates β-catenin directly on Tyr142, thereby releasing it from cadherin complexes (Krejci et al., 2012). In turn, epithelial β-catenin activation participates in the induction of Fgfr2b expression to increase Fgf10 signaling further (Shu et al., 2005). Epithelial β-catenin signaling, mediated primarily through Fgf10 signaling, is a regulator of branching morphogenesis and functions to maintain the distal epithelial progenitor cells in an undifferentiated state by inhibiting Sox2 expression (Mucenski et al., 2003; De Langhe et al., 2005; Shu et al., 2005; Hashimoto et al., 2012).
Here, we show that lung agenesis in Fgf10-/- mice can be rescued by ubiquitous overexpression of Fgf10, suggesting that epithelial branching morphogenesis is not dependent on exact Fgf10 localization. We also report that overexpression of the canonical Wnt inhibitor Dkk1 from E8.0 onwards using Rosa26-rtTa;Tet-Dkk1 mice prevents the specification of the primary lung field in the ventral foregut, confirming previous reports on the role of Wnt signaling in primary lung field specification (Cohen et al., 2009; Goss et al., 2009; Harris-Johnson et al., 2009; Chen et al., 2010). However, we found that specification of the primary lung field could not be rescued by simultaneous overexpression of Fgf10 using Rosa26-rtTa;Tet-Dkk1;Tet-Fgf10 mice.
Instead, our data show that after lung initiation, from E10.5 onwards, downregulation of Fgf10 expression is the primary cause for decreased branching and proximalization of lungs overexpressing Dkk1, as simultaneous overexpression of Fgf10 and Dkk1 rescues these defects in branching and proximal-distal differentiation. We therefore propose a model in which localized expression of Fgf10 is not required for stereotypic branching morphogenesis and suggest that Fgf10 signaling functions primarily as a permissive factor.
Interestingly, we found that in the trachea, Fgf10 plays an additional role in directing the differentiation of Sox2-expressing cells into the basal cell lineage and that overexpression of Fgf10 at later stages of lung development directs the differentiation of Sox2-expressing proximal airway epithelium to a basal cell fate while blocking the ciliated cell fate.
MATERIALS AND METHODS
Transgenic embryos
Tet-Dkk1 and Rosa26-rtTa mice were described previously (Volckaert et al., 2011). Rosa26-rtTA mice were crossed with Tet-sFgfr2b (Hokuto et al., 2003), Tet-Fgf10 (Clark et al., 2001) and Tet-Dkk1 mice to generate double transgenic embryos allowing for inducible expression by simply feeding pregnant females with doxycycline (dox)-containing food (rodent diet with 625 mg/kg doxycycline, Harlan Teklad TD.09761). TOPGAL mice were a generous gift from Dr Elaine Fuchs (DasGupta and Fuchs, 1999) and Fgf10LacZ mice were a generous gift from Robert Kelly (Kelly et al., 2001).
β-Galactosidase staining
Embryos and embryonic lungs containing TOPGAL or Fgf10LacZ alleles were stained as described previously (Volckaert et al., 2011).
Immunohistochemistry
All immunostaining was performed on paraffin sections of formalin-fixed lungs and embryos with the following primary antibodies: Nkx2.1 (1:200; Neomarkers), keratin 5 (1:200; Neomarkers), FoxA2 (1:2000; Seven Hills Bioreagents), Scgb1A1 (1:500; Seven Hills Bioreagents), Pro-Sftpc (1:500; Seven Hills Bioreagents), β-tubulin (1:500; Seven Hills Bioreagents), Sox2 (1:1000; Seven Hills Bioreagents), CGRP (1:5000; Sigma-Aldrich), Cy3-conjugated mouse anti-α-smooth muscle actin (1:200; Sigma-Aldrich), phospho-Histone-H3 (Ser10) (1:200; Cell Signaling Technology), P-β-catenin Ser552 (1:200; Cell Signaling Technology), P-Akt Ser473 (1:100; Cell Signaling Technology), fibronectin (1:500; a kind gift of Dr Kenneth Yamada, National Institute of Dental and Craniofacial Research, NIH, Bethesda, MD, USA), PDPN (1:50; Developmental Studies Hybridoma Bank), Scgb1A1 (1:200; Santa Cruz), p63 (1:200; Santa Cruz), Fgfr2 (1:200; Santa Cruz), CD31 (PECAM) (1:50; BD Biosciences), E-cadherin (1:200; BD Biosciences), Sox9 (1:100; Millipore). All fluorescent staining (except the Cy3-conjugated α-SMA) was performed with Cy3- and FITC-conjugated secondary antibodies from Jackson ImmunoResearch and mounted using Vectashield with DAPI (Vector Laboratories). Photographs were taken with a Zeiss AxioImager and Axiovision software.
Quantification of P-Akt Ser473- and P-β-catenin Ser552-positive epithelial cells
Epithelial P-Akt Ser473 quantification was performed by measuring mean gray value over mean epithelial surface area using ImageJ software. P-β-catenin Ser552 quantification was performed by determining the ratio of P-β-catenin Ser552-labeled epithelial cells over the total number of epithelial cells per field. The percentage of P-β-catenin Ser552-labeled cells is reported. Statistical analysis was performed using Student’s t-test.
Quantitative real-time PCR
RNA was isolated from whole lungs or tracheas using RNALater (Ambion) and Total RNA Kit I (Omega Biotek) according to the manufacturer’s instructions. RNA concentration was determined by spectrophotometry. cDNA was generated using SuperScript III First-Strand Synthesis System (Invitrogen) according to the manufacturer’s instructions. Comparative real-time PCR was performed for β-glucuronidase (Mm00446953_m1), Dkk1 (Mm00438422_m1), Fgfr2b (Mm01269938_m1), Fgf10 (Mm01297079_m1), Nmyc (Mycn) (Mm00476449_m1), Sftpc (Mm00488144_m1), fibronectin (Mm01256744_m1), Scgb1a1 (Mm00442046_m1), Foxj1 (Mm00807215_m1), Cgrp (Calca) (Mm00801463_g1), Acta2 (SMA) (Mm00725412_s1), Spry2 (Mm00442344_m1), keratin 5 (Krt5) (Mm01305291_g1), p63 (Trp63) (Mm00495788_m1) Taqman Gene Expression Assays (Applied Biosystems) using a StepOne Plus system (Applied Biosystems). β-Glucuronidase was used as a reference control to normalize equal loading of template cDNA.
In situ hybridization
Whole-mount in situ hybridization was performed as previously described (De Langhe et al., 2005). Sections from paraffin-embedded embryonic lungs were hybridized as previously described (De Langhe et al., 2008; De Langhe et al., 2005). The following mouse cDNAs were used as templates for the synthesis of digoxigenin-labeled riboprobes: a 584-bp fragment of Fgf10 (Bellusci et al., 1997), a 948-bp full-length Spry2 cDNA and a 162-bp fragment of Fgfr2IIIb.
Quantification of branching
Peripheral lung buds were counted and statistical analysis was performed using Student’s t-test.
RESULTS
Fgf10 overexpression partially rescues lung and limb agenesis in Fgf10-/- mice
The importance of Fgf10 in lung morphogenesis has been illustrated by the drastic phenotype in mice lacking Fgf10 or its receptor Fgfr2b. The trachea still develops in these mice, but the lung fails to form, revealing an essential function for Fgf10 signaling in lung formation (Min et al., 1998; Arman et al., 1999; Sekine et al., 1999; De Moerlooze et al., 2000). Similarly, the lack of limbs in these mice illustrates the important role of Fgf10 signaling in limb formation.
The current prevailing theory is that directional lung bud outgrowth is dependent on the precisely localized, directional expression of Fgf10 in the lung mesenchyme distal to the branch point, which regulates the formation of the initial domain branches and all subsequent branches (Warburton, 2008; Ornitz and Yin, 2012). However, the observation that isolated lung epithelium branches when grown in Matrigel in the presence of recombinant Fgf10 (Bellusci et al., 1997; Park et al., 1998; Ohtsuka et al., 2001) contradicts this model for branching morphogenesis.
To investigate the importance of localized Fgf10 expression for branching morphogenesis in vivo, we generated Fgf10-/-;Rosa26-rtTa;Tet-Fgf10 mice in which we can induce Fgf10 expression ubiquitously. Although Fgf10-/- mice have no limbs (Fig. 1C) and show complete lung agenesis (Fig. 1F), ubiquitous overexpression of Fgf10 in a Fgf10-/- background from E9.5 onwards rescued lung development with a close to normal lobulation and epithelial branching pattern (Fig. 1E) compared with wild-type lungs (Fig. 1D). In addition, our data demonstrate that ubiquitous overexpression of Fgf10 partially rescues limb formation, with budding of both front and hind limbs at the correct position, similar to control embryos (Fig. 1A,B). This is interesting considering the observation that in chicken ectopic Fgf10 signaling results in the formation of ectopic limbs (Ohuchi et al., 1997).
Fig. 1.
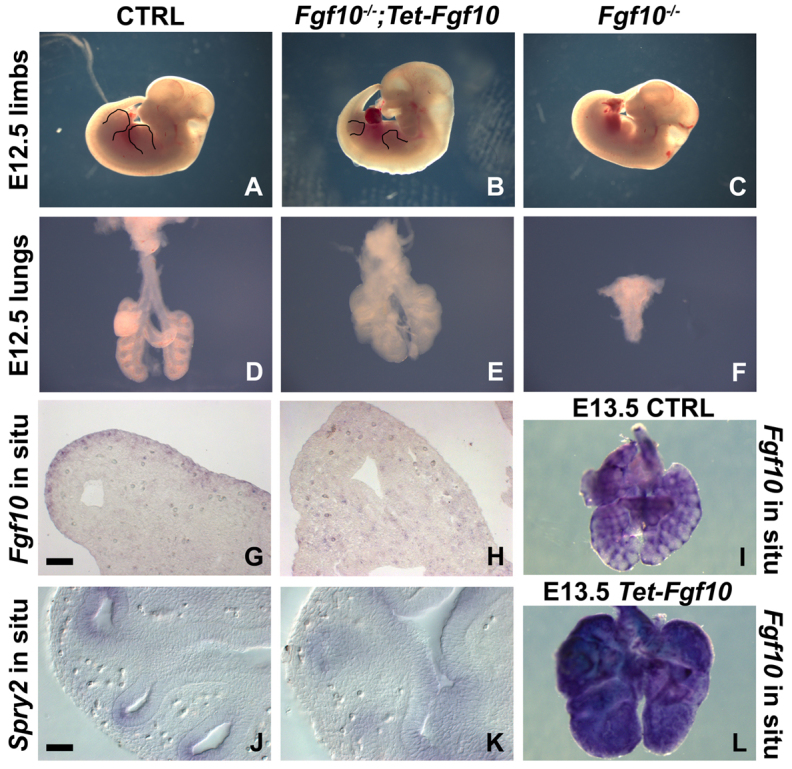
Fgf10 overexpression partially rescues lung and limb agenesis in Fgf10-/- mice. (A-C) E12.5 wild-type (A), Fgf10-/-;Rosa26-rtTA;Tet-Fgf10 dox-induced at E9.5 (B) and Fgf10-/- (C) embryos. Black outlines visualize front and hind limbs. (D-F) E12.5 lungs from wild-type (D), Fgf10-/-;Rosa26-rtTA;Tet-Fgf10 dox-induced at E9.5 (E) and Fgf10-/- (F) mice. (G,H) Fgf10 in situ hybridization on sections from E12.5 wild-type lung (G) and Fgf10-/-;Rosa26-rtTA;Tet-Fgf10 lung dox-induced at E9.5 (H). (J,K) Vibratome sections from whole-mount Spry2 in situ hybridization on E12.5 wild-type lung (J) and Fgf10-/-;Rosa26-rtTA;Tet-Fgf10 lung dox-induced at E9.5 (K). (I,L) Whole-mount Fgf10 in situ hybridization on E13.5 wild-type lungs (I) and Rosa26-rtTA;Tet-Fgf10 lungs dox-induced at E10.5 (L). n≥3. Scale bars: 50 μm (G,H,J,K).
To determine whether the ubiquitously overexpressed Fgf10 in our system is not post-transcriptionally regulated to allow localized expression and branching morphogenesis to occur, we performed in situ hybridization for Fgf10 on E12.5 wild-type and Fgf10-/-;Rosa26-rtTa;Tet-Fgf10-rescued lung sections. Fig. 1G illustrates that in E12.5 wild-type lungs, Fgf10 expression is limited to the distal mesenchyme whereas low ubiquitous levels of Fgf10 expression can be found throughout Fgf10-/-;Rosa26-rtTa;Tet-Fgf10 lungs in both the mesenchyme and epithelium (Fig. 1H). We found that low levels of Fgf10 were essential for proper lung development whereas higher levels perturbed the branching pattern. Thus, to demonstrate that Fgf10 is indeed ubiquitously expressed in our model, we also performed whole-mount in situ hybridization for Fgf10 on E13.5 wild-type (wt) (Fig. 1I) and Rosa26-rtTa;Tet-Fgf10 (Fig. 1L) lungs induced from E10.5. To further show that Fgf10 protein does not act locally in Fgf10-/-;Rosa26-rtTa;Tet-Fgf10-rescued lungs, we performed in situ hybridization for Spry2 expression. In the wt lung, Spry2 expression is regulated by Fgf10 and is restricted to the epithelium of the distal branching tips (Fig. 1J), whereas in Fgf10-/-;Rosa26-rtTa;Tet-Fgf10-rescued lungs Spry2 is expressed at low levels throughout the entire lung epithelium (Fig. 1K).
In contrast to the current dogma, which ascribes a pivotal role to localized Fgf10 expression in directing lung epithelial bud and limb outgrowth, our data reveal that the spatial distribution of Fgf10 is less important for the process of epithelial bud outgrowth and stereotypic branching than was previously thought. Therefore, we speculate that localized Fgf10 expression might be important primarily for proximal-distal patterning by keeping the distal epithelium in an undifferentiated state.
Dkk1 overexpression effectively abrogates the initiation of the respiratory lineage, which cannot be rescued by simultaneous overexpression of Fgf10
Wnt/β-catenin signaling has proven to be essential in specifying lung progenitor cells in the developing foregut (Goss et al., 2009; Harris-Johnson et al., 2009). Recently, an elegantly performed series of foregut explant experiments revealed that RA is a major regulator of Wnt and Tgfβ pathways during the formation of the lung primordium (Chen et al., 2010). The authors proposed a mechanism through which RA activates canonical Wnt signaling by inhibiting its negative regulator Dkk1. Dkk1 binds to the Wnt co-receptor Lrp5/6, making it inaccessible to Wnt ligands (Bao et al., 2012). Previous experimental set-ups have shown that Dkk1 effectively inhibits Wnt/β-catenin signaling in the lung (De Langhe et al., 2005; Shu et al., 2005; Chen et al., 2010).
Based on this model, we hypothesized that ubiquitous overexpression of Dkk1 would override the effect of endogenous RA in vivo and result in failure of initiation of the respiratory lineage. Indeed, by analyzing expression of Nkx2.1, the earliest known marker for both lung and trachea (Kimura et al., 1996), and Foxa2, an endoderm marker (Ang et al., 1993), we found that overexpression of Dkk1 prior to lung initiation affects primary lung field specification in a dose-dependent manner (Fig. 2E-G,I-K). Mice homozygous for Dkk1 fail to specify the primary lung field illustrated by the lack of Nkx2.1 expression (Fig. 2G) and formation of lung branches (Fig. 2C,G,K), whereas heterozygous mice show proper specification of the primary lung field (Fig. 2F) but form smaller lung branches (Fig. 2B,F,J) compared with control lungs (Fig. 2A,E,I).
Fig. 2.

Dkk1 overexpression abrogates initial lung formation. (A-D) β-Gal staining on E10.5 control (ctrl) TOPGAL (A), Rosa26-rtTA;Tet-Dkk1+/-;TOPGAL (B), Rosa26-rtTA;Tet-Dkk1+/+;TOPGAL (C) and Rosa26-rtTA;Tet-Dkk1+/+;Tet-Fgf10;TOPGAL (D) lungs/foreguts dox-induced at E8.0. (E-H) Nkx2.1 immunostaining on E10.5 ctrl (E), Rosa26-rtTA;Tet-Dkk1+/- (F), Rosa26-rtTA;Tet-Dkk1+/+ (G) and Rosa26-rtTA;Tet-Dkk1+/+;Tet-Fgf10 (H) lungs/foreguts dox-induced at E8.0. (I-L) Foxa2 immunostaining on E10.5 ctrl (I), Rosa26-rtTA;Tet-Dkk1+/- (J), Rosa26-rtTA;Tet-Dkk1+/+ (K) and Rosa26-rtTA;Tet-Dkk1+/+;Tet-Fgf10 (L) lungs/foreguts dox-induced at E8.0. (M,N) qPCR analysis of relative Dkk1 (M) and Fgf10 (N) mRNA abundance on E10.5 Rosa26-rtTA;Tet-Dkk1+/+ and Rosa26-rtTA;Tet-Dkk1+/+;Tet-Fgf10 embryos dox-induced at E8.0. **P<0.01 (Student’s t-test); n≥3. Error bars represent standard error. (O,P) Whole-mount Fgf10 in situ hybridization on an E10.5 Rosa26-rtTA;Tet-Dkk1+/+ embryo (O) and a Rosa26-rtTA;Tet-Dkk1+/+;Tet-Fgf10 embryo dox-induced from E10.5 onwards (P). n≥3. Scale bars: 50 μm (E-L).
Through regulation of Wnt2a/b and Tgfβ signaling, RA also regulates mesenchymal Fgf10 expression around this stage of lung development (Chen et al., 2010). However, as specification of respiratory lineages in the foregut does occur in Fgf10-/- mice, Fgf10 is not thought to be essential for the initial specification of respiratory progenitors (Sekine et al., 1999). To investigate whether Fgf10 could be the main driver of β-catenin signaling in the epithelium at this early stage and as such could rescue lung agenesis, we simultaneously overexpressed Fgf10 and Dkk1 in vivo from E8.0 onwards using Rosa26-rtTa;Tet-Dkk1;Tet-Fgf10 mice (Fig. 2D,H,L-P). However, we found that lung formation was still abrogated in these embryos (Fig. 2D,H,L). This indicates that even though Fgf10 can activate/enhance epithelial β-catenin signaling at later stages of lung development (see below), Fgf10 alone is not sufficient to specify the lung progenitors in the developing foregut in the absence of Wnt ligand-mediated epithelial β-catenin signaling. Our findings, obtained using a novel model of Wnt signaling inhibition, emphasize the importance of canonical Wnt2a/b signaling in the initiation and specification of the respiratory cell lineage (Goss et al., 2009).
Dkk1 overexpression reduces Fgf10 expression and prevents amplification of a pool of Fgf10-expressing parabronchial smooth muscle progenitor cells
The distal lung comprises two pools of mesenchymal cell populations: sub-epithelial and sub-mesothelial (White et al., 2006). Whereas the former responds to epithelial-derived Shh, which inhibits Fgf10 expression (Pepicelli et al., 1998), we have previously shown that the sub-mesothelial cells are parabronchial smooth muscle cell (PSMC) progenitors (Mailleux et al., 2005) expressing high levels of Fgf10. Through mesenchymal-specific deletion of Ctnnb1 (the gene encoding β-catenin), we previously showed that Wnt/β-catenin signaling regulates Fgf10 expression and controls the amplification of these PSMC progenitors (De Langhe et al., 2008). Furthermore, we have previously shown that treatment of embryonic lung explants with Dkk1 prevents the differentiation of PSMCs (De Langhe et al., 2005). Since then, a specific role for Wnt ligands Wnt7b and Wnt2a in airway smooth muscle precursor development and Fgf10 expression has been described (Rajagopal et al., 2008; Cohen et al., 2009; Goss et al., 2011).
To broaden our understanding of the role of canonical Wnt signaling and its interplay with Fgf10 during early branching morphogenesis, we induced Rosa26-rtTa;Tet-Dkk1 mice from E10.5 onwards. At this stage, the lung is composed of the trachea and two growing lung buds. Using Fgf10LacZ+ reporter mice, which express lacZ under the transcriptional control of Fgf10 regulatory sequences, we confirmed that Fgf10 expression and the pool of PSMC progenitors in the distal mesenchyme is significantly reduced in E12.5 Rosa26-rtTa;Tet-Dkk1;Fgf10LacZ+ lungs (Fig. 3A,B; Fig. 4K) and that fewer β-galactosidase (β-gal)-positive PSMCs could be found around the bronchi (Fig. 3A-D). This also resulted in decreased epithelial Fgf10 signaling in Rosa26-rtTa;Tet-Dkk1 lungs, demonstrated by reduced and more ‘distalized’ epithelial expression of Spry2 (Fig. 3E-H; Fig. 4K) and Fgfr2b (Fig. 3I-L; Fig. 4K), two Fgf10 target genes (Mailleux et al., 2001; Shu et al., 2005). The reduction of PSMC progenitors in lungs overexpressing Dkk1 resulted in a discontinuous, thinner PSMC layer (Fig. 3N; Fig. 4K) compared with control lungs (Fig. 3M; Fig. 4K). Our findings recapitulate previously reported defects in PSMC formation after inhibition of mesenchymal β-catenin signaling (De Langhe et al., 2005; De Langhe et al., 2008; Cohen et al., 2009; Goss et al., 2011). Interestingly, endothelial cell differentiation visualized by Pecam immunostaining is normal in Rosa26-rtTa;Tet-Dkk1 lungs (Fig. 3O,P), in contrast to what we observed after mesenchymal-specific deletion of Ctnnb1 (De Langhe et al., 2008). This is possibly due to the fact that Dkk1 does not inhibit Fgf9-mediated activation of β-catenin signaling (Yin et al., 2008; Yin et al., 2011). Lastly, we found that fibronectin (FN) deposition was reduced in lungs overexpressing Dkk1 (Fig. 3N; Fig. 4K) compared with control lungs (Fig. 3M; Fig. 4K), confirming our previous observation in lung explants grown in the presence of Dkk1 (De Langhe et al., 2005).
Fig. 3.

Dkk1 overexpression reduces Fgf10 expression and prevents amplification of distal parabronchial smooth muscle cell progenitors. (A-D) β-Gal-stained lungs (A,B) and corresponding vibratome sections through the medial lobes (C,D) of E12.5 control (ctrl) Fgf10LacZ (A,C) and Rosa26-rtTA;Tet-Dkk1;Fgf10LacZ lungs (B,D) dox-induced at E10.5. (E-L) Whole-mount in situ hybridization (E,F,I,J) and corresponding vibratome sections (G,H,K,L) for Spry2 (E-H) and Fgfr2b (I-L) on E13.5 ctrl (E,G,I,K) and Rosa26-rtTA;Tet-Dkk1 lungs (F,H,J,L) dox-induced at E10.5. (M,N) Immunostaining for fibronectin (FN) and α-SMA on E13.5 ctrl (M) and Rosa26-rtTA;Tet-Dkk1 (N) lungs dox-induced at E10.5. (O,P) Immunostaining for Pecam on E13.5 ctrl (O) and Rosa26-rtTA;Tet-Dkk1 (P) lungs dox-induced at E10.5. n≥3. Scale bars: 50 μm (C,D,G,H,K,L); 100 μm (M-P).
Fig. 4.

Fgf10 signaling prevents differentiation of distal epithelial progenitors by positively regulating β-catenin signaling. (A,B) β-Gal staining on E13.5 control (ctrl) TOPGAL (A) and Rosa26-rtTA;Tet-Fgf10;TOPGAL (B) lungs dox-induced at E10.5. (C-F) Immunostaining for E-cadherin and p-Akt-Ser473 (C,D) or β-catenin-Ser552 (E,F) on E13.5 ctrl (C,E) and Rosa26-rtTA;Tet-Fgf10 (D,F) lungs dox-induced at E10.5. (G,H) Immunostaining for Fgfr2b on E13.5 ctrl (G) and Rosa26-rtTA;Tet-Fgf10 (H) lungs dox-induced at E10.5. (I,J) Immunostaining for α-SMA and Sftpc on E13.5 ctrl (I) and Rosa26-rtTA;Tet-Fgf10 (J) lungs dox-induced at E10.5. (K) qPCR analysis of relative Fgf10, Spry2, Fgfr2b, α-Sma (Acta2) and Fn1 mRNA abundance on E11.5 (Fgf10, Spry2) or E12.5 (Fgfr2b, α-Sma, Fn1) ctrl and Rosa26-rtTA;Tet-Dkk1+/+ lungs dox-induced at E10.5. **P<0.01, *P<0.05 (Student’s t-test); n≥3. (To allow for better visualization, Fn1 expression levels presented in the graph are reduced by a factor of 2). (L) qPCR analysis of relative Nmyc, Fgfr2b and Sftpc mRNA abundance on E12.5 ctrl and Rosa26-rtTA;Tet-Fgf10 lungs dox-induced at E10.5. **P<0.01 (Student’s t-test); n≥3. Error bars represent standard error. Scale bars: 50 μm (C-F); 100 μm (G,H); 75 μm (I,J).
Fgf10 positively regulates β-catenin signaling through activation of Akt and prevents differentiation of the distal epithelial progenitors
Fgf10 has been shown to regulate epithelial β-catenin signaling during lung development as well as in the adult lung (Lü et al., 2005; Ramasamy et al., 2007; Volckaert et al., 2011). We therefore examined the status of epithelial Wnt/β-catenin signaling in lungs overexpressing Fgf10. To do this, we introduced the TOPGAL reporter allele into Rosa26-rtTa;Tet-Fgf10 mice and analyzed TOPGAL activity by β-gal staining. E13.5 Rosa26-rtTa;Tet-Fgf10;TOPGAL lungs induced from E10.5 showed a notable increase in epithelial TOPGAL activity compared with control TOPGAL lungs (Fig. 4A,B). In addition, we quantified activation of epithelial Wnt signaling by quantitative real-time PCR (qPCR) for Nmyc, an epithelial-specific Wnt target gene (Fig. 4L) (Shu et al., 2005), confirming our TOPGAL readout.
Fgf10 is thought to directly activate β-catenin signaling by activating the PI3K-AKT pathway (Volckaert et al., 2011). Phosphorylated Akt (P-Akt) acts to prevent the degradation of β-catenin by inhibiting its negative regulator Gsk3β and to help drive β-catenin to the nucleus by phosphorylating it directly on Ser552 (He et al., 2007). To elucidate whether Fgf10 directly activates β-catenin signaling through this mechanism, we performed immunostaining for P-Akt or P-β-cat-Ser552 together with E-cadherin (cadherin 1), to outline the epithelium on E13.5 control and Rosa26-rtTa;Tet-Fgf10 lungs induced from E10.5 (Fig. 4C-F). We found a 17±1.2% (mean±standard error; n≥4; P=0.017) increase in P-Akt phosphorylation and a 167±34% (n≥4; P=0.000021) increase in β-cat-Ser552 phosphorylation in the epithelium of lungs overexpressing Fgf10 versus wild-type lungs. Our results therefore indicate that Fgf10 signaling activates epithelial β-catenin signaling by activating Akt.
Proper lung morphogenesis is dependent on a tightly regulated balance between progenitor cell expansion and differentiation. This is reflected in the temporal and spatial expression of multiple genes involved in proximal-distal patterning in the developing lung, including Wnts, Shh, Nmyc, Fgf10 and Bmp4. Consequently, perturbations in these signaling systems can shift the balance either towards maintaining a distal population of undifferentiated, proliferating progenitor cells or towards differentiation into conducting airway epithelial cells (Rawlins et al., 2009a; Domyan and Sun, 2011).
Previous work has revealed an important function for Wnt/β-catenin signaling in the regulation of proximal-distal differentiation in the developing airway epithelium (Mucenski et al., 2003; Okubo and Hogan, 2004; De Langhe et al., 2005; Shu et al., 2005; Hashimoto et al., 2012). This led to a model in which Wnt/β-catenin signaling is crucial in maintaining the reservoir of epithelial and mesenchymal progenitor cells residing in the distal tips of the embryonic lung. Loss of Wnt signaling can therefore shift the balance from progenitor cell maintenance to differentiation. Our data show that ubiquitous overexpression of Fgf10 from E10.5 to E13.5, possibly by increasing epithelial β-catenin signaling, prevents the differentiation of distal epithelial progenitors, which is illustrated by the maintenance of Fgfr2b (Fig. 4G,H,L) and Sftpc (Fig. 4I,J,L) in more proximal epithelium.
Downregulation of Fgf10 expression is the primary cause for decreased branching and proximalization of lungs overexpressing Dkk1
To examine the status of epithelial Wnt/β-catenin signaling in lungs overexpressing Dkk1, we crossed the TOPGAL reporter allele with the Rosa26-rtTa;Tet-Dkk1 line and analyzed β-catenin signaling activity by β-gal staining. E11.5 Rosa26-rtTa;Tet-Dkk1;TOPGAL lungs induced from E10.5 showed a notable reduction in epithelial TOPGAL activity compared with control TOPGAL lungs (Fig. 5A,B). To analyze whether the reduction in Fgf10 signaling is the primary reason for reduced epithelial β-catenin signaling in lungs overexpressing Dkk1, we simultaneously overexpressed Fgf10 and found that epithelial Wnt/β-catenin signaling was rescued (Fig. 5A-C). Activation of epithelial Wnt signaling in these different lungs was further quantified by qPCR for Nmyc, an epithelial-specific Wnt target gene (Fig. 5D) (Shu et al., 2005), confirming our TOPGAL readout. Together, our data indicate that Fgf10 is able to activate epithelial Wnt signaling, probably independently of Wnt ligands or enhanced Wnt-mediated signaling, a mechanism that is also observed in the adult lung airway epithelium after injury (Volckaert et al., 2011).
Fig. 5.

Reduced epithelial Wnt signaling in lungs overexpressing Dkk1 results in proximal-distal differentiation defects that can be rescued by overexpressing Fgf10. (A-C) β-Gal staining on E11.5 control (ctrl) TOPGAL (A), Rosa26-rtTA;Tet-Dkk1;TOPGAL (B) Rosa26-rtTA;Tet-Dkk1;Tet-Fgf10;TOPGAL (C) lungs dox-induced at E10.5. (D) qPCR analysis of relative Nmyc mRNA abundance on E11.5 ctrl, Rosa26-rtTA;Tet-Dkk1+/+ and Rosa26-rtTA;Tet-Dkk1+/+;Tet-Fgf10 lungs dox-induced at E10.5. **P<0.01 (Student’s t-test); n≥3. Error bars represent standard error. (E-L) Immunostaining for Sox2 (E-H) and Sox9 (I-L) on E12.5 ctrl (E,I), Rosa26-rtTA;Tet-Dkk1 (F,J), Rosa26-rtTA;Tet-Dkk1;Tet-Fgf10 (G,K) and Fgf10-/-;Rosa26-rtTA;Tet-Fgf10 (H,L) lungs dox-induced at E10.5. n≥3. Scale bars: 200 μm (E-L).
We found that decreased Fgf10 expression is the primary cause for decreased branching and the premature differentiation of distal Sox9-expressing epithelial progenitors into Sox2-expressing conducting airway epithelial cells in lungs overexpressing Dkk1 as both branching and proximal-distal differentiation could be rescued by simultaneous overexpression of Fgf10 (Fig. 5E-G,I-K). As such, branching in E11.5 lungs overexpressing Dkk1 was reduced by 25±2.2% (n≥4; P=0.00004) compared with wild-type lungs, whereas simultaneous overexpression of Fgf10 with Dkk1 partially rescued branching morphogenesis by increasing branching by 12±0.8% (n≥4; P=0.008) compared with lungs overexpressing Dkk1 alone.
These results are in line with a model in which Fgf10 signaling localized distally, prevents the differentiation of the distal epithelial progenitors into airway epithelial cells by activating epithelial β-catenin signaling and inhibiting Sox2 expression (Hashimoto et al., 2012). Interestingly, a proximal-distal axis of differentiation is still present in Fgf10-/- lungs rescued by overexpression of Fgf10 ubiquitously (Fig. 5H,L), indicating that other factors besides Fgf10 play a role in distal epithelial progenitor maintenance.
Fgf10 inhibits Sox2 expression and proximal epithelial differentiation prior to E12.5 but promotes basal cell differentiation/amplification afterwards
We next determined the exact window of opportunity for Fgf10 to inhibit Sox2 expression and thus prevent the differentiation of distal epithelial progenitors into airway epithelial cells. To do this, we induced Rosa26-rtTa;Tet-Fgf10 lungs from either E11.5, E12.5 or E13.5 until E18.5 and monitored Sox2 (proximal epithelium), Sox9 (distal epithelial progenitors), p63 and K5 (basal cells), Scgb1a1 (Clara cells) and Sftpc [alveolar type II (ATII) or distal epithelial progenitor cells] markers of differentiation. We found that only overexpression of Fgf10 from E11.5 onwards was able to prevent the differentiation of almost all distal Sox9-expressing epithelial progenitors into Sox2-expressing airway epithelial cells (Fig. 6B,F,J,N). Overexpression of Fgf10 from E12.5 onwards did not prevent the differentiation of all distal epithelial progenitors as some had already differentiated into Sox2-expressing airway epithelial cells by this time (Fig. 6C,G,K,O). Fgf10 overexpression from E12.5 or E13.5 onwards did, however, prevent further differentiation of the remaining distal Sox9-expressing epithelial progenitors. This was evidenced by the persistent expression of Sox9 and Sftpc in distal epithelial cells in these lungs at E18.5 (Fig. 6G,H,O,P) as opposed to control lungs in which the distal epithelium had differentiated into ATII cells expressing Sftpc but not Sox9 (Fig. 6E,M). Interestingly, Fgf10 overexpression from E12.5 or E13.5 onwards also had an unexpected effect on Sox2-expressing airway epithelial cells, preventing neuroendocrine and ciliated cell differentiation (data not shown) and promoting differentiation of a subset of Sox2-positive cells into p63-positive basal cells (Fig. 6K,L). In addition, Fgf10 induced proliferation in Clara cells and ectopic basal cells (supplementary material Fig. S1B,C,E,F). Interestingly, we found that Fgf10 also induced expression of p63 in some Sox9-positive cells at the transition zone between Sox2- and Sox9-positive cells, suggesting that a subset of Sox9-positive cells can become basal cells or that at the transition zone some cells are triple positive, expressing Sox2, Sox9 and p63 simultaneously (supplementary material Fig. S1A,D).
Fig. 6.

Early Fgf10 overexpression prevents proximal differentiation whereas Fgf10 overexpression after E12.5 promotes basal cell differentiation. (A-D) Immunostaining for Sox2 on E18.5 control (ctrl) (A) and Rosa26-rtTA;Tet-Fgf10 lungs induced from E11.5 (B), E12.5 (C) and E13.5 (D). (E-H) Immunostaining for Sox9 on E18.5 ctrl (E) and Rosa26-rtTA;Tet-Fgf10 lungs induced from E11.5 (F), E12.5 (G) and E13.5 (H). (I-L) Immunostaining for p63 and K5 on E18.5 ctrl (I) and Rosa26-rtTA;Tet-Fgf10 lungs induced from E11.5 (J), E12.5 (K) and E13.5 (L). (M-P) Immunostaining for Sftpc and Scgb1a1 on E18.5 ctrl (M) and Rosa26-rtTA;Tet-Fgf10 lungs induced from E11.5 (N), E12.5 (O) and E13.5 (P). n≥3. Scale bars: 200 μm (A-H); 100 μm (I-P).
Opposing and similar effects of Dkk1 and Fgf10 on lung epithelial differentiation
Rosa26-rtTa;Tet-Dkk1 mice induced from E10.5 survive to E18.5 and demonstrate a more drastic phenotype than previously reported for Sftpc-rtTa;Tet-Dkk1 mice (Shu et al., 2005). This is likely to be due to higher and more ubiquitous levels of Dkk1 expression. E18.5 Rosa26-rtTa;Tet-Dkk1 lungs are dramatically ‘proximalized’ with the formation of very few primitive alveolar sacs, containing few alveolar type I (ATI) (Pdpn+) and II (ATII) (Sftpc+) cells (Fig. 7B,E) compared with control lungs (Fig. 7A,D). By contrast, induction of Fgf10 expression from E15.5 onwards prevented the conclusion of the pseudoglandular stage in Rosa26-rtTa;Tet-Fgf10 lungs (Fig. 7C,F). As such, Rosa26-rtTa;Tet-Fgf10 lungs induced from E15.5 failed to transition out of the branching or pseudoglandular stage into the canalicular and saccular stages. However, differentiation of ATI cells did occur based on podoplanin (Pdpn; also known as T1α) expression (Fig. 7F).
Fig. 7.

Opposing and similar effects of Dkk1 and Fgf10 on lung epithelial differentiation. (A-F) Immunostaining for α-SMA and Sftpc (A-C) or for Pdpn (D-F), on E18.5 control (ctrl) lungs (A,D), E18.5 Rosa26-rtTA;Tet-Dkk1 lungs dox-induced from E10.5 onwards (B,E) and E18.5 Rosa26-rtTA;Tet-Fgf10 (C,F) lungs dox-induced from E15.5 onwards. (G-I) Immunostaining for Scgb1a1 and Cgrp on E18.5 ctrl (G), Rosa26-rtTA;Tet-Dkk1 (H) and Rosa26-rtTA;Tet-Fgf10 (I) lungs induced from E10.5 and E15.5, respectively. (J-L) Immunostaining for Scgb1a1 and β-tubulin on E18.5 ctrl (J), Rosa26-rtTA;Tet-Dkk1 (K) and Rosa26-rtTA;Tet-Fgf10 (L) lungs induced from E10.5 and E15.5, respectively. (M-R) Immunostaining for keratin5 and p63 on E18.5 ctrl (M,P), Rosa26-rtTA;Tet-Dkk1 (N,Q) and Rosa26-rtTA;Tet-Fgf10 (O,R) lungs induced from E10.5 and E15.5, respectively. P-R are higher magnifications of M-O, respectively. n≥3. Scale bars: 100 μm (A-F,M-O); 50 μm (G-L); 25 μm (P-R).
We next investigated the effects of Dkk1 and late-stage Fgf10 overexpression on the differentiation of conducting airway epithelial cells. We found that overexpression of Dkk1 resulted in smaller neuroendocrine bodies (Fig. 7H) than were observed in control lungs (Fig. 7G), whereas overexpression of Fgf10 had the opposite effect (Fig. 7I). However, when quantified by qPCR, Rosa26-rtTA;Tet-Dkk1 lungs showed similar amounts of Cgrp expression as in control lungs whereas Rosa26-rtTA;Tet-Fgf10 lungs showed a reduction in overall Cgrp expression (Fig. 8E). This discrepancy between immunostaining and whole lung mRNA expression levels is likely to be due to overall changes in the ratio of cells present in these lungs. Alternatively, it is possible that Fgf10 overexpression promoted the amplification of neuroendocrine cells that had already differentiated by E15.5 but prevented further differentiation of new neuroendocrine cells. Interestingly, we found that overexpression of Fgf10 from E15.5 onwards was still able to completely block ciliated cell differentiation (Fig. 7L) compared with control lungs and lungs overexpressing Dkk1, showing a ‘salt and pepper’-like pattern of Clara (Scgb1a1) and ciliated (β-tubulin) cells (Fig. 7J,K). Changes in Clara versus ciliated cell distribution were further quantified by qPCR for Scgb1a1 and Foxj1 (ciliated cells) (Fig. 8E) and the absence of ciliated cells in Rosa26-rtTA;Tet-Fgf10 lungs was confirmed.
Fig. 8.

Fgf(10) signaling regulates basal cell differentiation in the trachea. (A-D) Immunostaining for p63 and K5 on E18.5 wild-type (A), Fgf10-/- (B), Rosa26-rtTA;Tet-sFgfr2b induced from E15.5 (C) and E15.5 wild-type (D) tracheas. Insets show high magnification images of boxed areas. (E) qPCR analysis of relative Scgb1a1, FoxJ1, Cgrp, keratin 5 and p63 mRNA abundance on E18.5 lungs from wild-type mice, Rosa26-rtTa;Tet-Dkk1 mice induced from E10.5 and Rosa26-rtTA-Tet-Fgf10 mice induced from E15.5. (F) qPCR analysis of relative keratin 5 and p63 mRNA abundance on E18.5 tracheas from wild-type mice, Fgf10-/- mice and Rosa26-rtTA-Tet-sFgfr2b mice induced from E15.5. **P<0.01, *P<0.05 (Student’s t-test). n≥3. (To allow for better visualization Scgb1a1, Foxj1 and Cgrp expression levels presented in the graph are reduced by a factor of 1500, 10 and 5, respectively.) Error bars represent standard error. Scale bars: 100 μm (A-D); 25 μm (insets in A-D).
In wt lungs, K5+/p63+ basal cells can be found only in the main stem bronchi and trachea (Fig. 7M,P). Surprisingly, analysis of E18.5 Rosa26-rtTa;Tet-Dkk1 lungs induced from E10.5 revealed K5+/p63+ basal cells lining the entire airway epithelium (Fig. 7N,Q). This indicates that inhibition of Wnt signaling promotes the differentiation of basal cells probably through premature airway epithelial cell differentiation and increased Sox2 expression. This would be consistent with a recent report showing that β-catenin negatively regulates Sox2, whereas Sox2 regulates p63, a transcription factor essential for basal cell differentiation (Koster et al., 2004; Hashimoto et al., 2012). Interestingly, we found that overexpression of Dkk1 after E12.5 had no effect on basal cell amplification (data not shown). By contrast, we found that overexpression of Fgf10 from E15.5 onwards still promotes basal cell differentiation of a subset of Sox2-positive airway epithelial cells (Fig. 7O,R). qPCR analysis of K5 and p63 confirms that E18.5 Rosa26-rtTa;Tet-Dkk1 lungs induced from E10.5 show an approximately sevenfold increase in K5 and an approximately fivefold increase in p63 expression (Fig. 8E) compared with E18.5 wild-type lungs, whereas E18.5 Rosa26-rtTa;Tet-Fgf10 lungs induced from E15.5 show an approximately fourfold increase in K5 and ∼15-fold increase in p63 expression (Fig. 8E).
It is tempting to speculate that the reason why basal cells can only be found in the main stem bronchi and trachea in wild-type mice is the presence of Fgf10 expression in the mesenchyme between the cartilaginous rings (Tiozzo et al., 2009) but not in the airway smooth muscle cells that line the bronchi (Mailleux et al., 2005).
Fgf(10) signaling regulates basal cell differentiation in the trachea
To investigate whether FGF signaling is important for basal cell differentiation or their maintenance in the trachea, we analyzed E18.5 Fgf10-/- tracheas and found that basal cells were still present in the upper trachea (data not shown) but significantly reduced in the lower trachea (Fig. 8B) compared with wild-type tracheas (Fig. 8A). This was confirmed by qPCR analysis in which we found a 33% reduction in K5 and p63 expression compared with wild-type tracheas (Fig. 8F). It is possible that other Fgfs such as Fgf7, which is highly expressed in the trachea (Finch et al., 1995) and acts via the same Fgfr2b receptor, might also play a role in basal cell differentiation in the trachea. To test this, we also analyzed E18.5 Rosa26-rtTa;Tet-sFgfr2b tracheas induced from E15.5, which overexpress a secreted dominant-negative receptor for Fgf10 and Fgf7, and found them to be almost devoid of basal cells (Fig. 8C). This is in line with a 54% reduction in K5 and p63 expression compared with wild-type tracheas as shown by qPCR analysis (Fig. 8F). Interestingly, we found that at E15.5, the trachea is already lined with immature p63/K5-positive basal cells (Fig. 8D), therefore attributing a potentially crucial role for FGF signaling in basal cell maintenance.
DISCUSSION
We have shown here that localized expression of Fgf10 is not required to direct stereotypic branching and that the role of Fgf10 signaling during early lung development lies primarily in preventing distal epithelial cell differentiation, in part by activating epithelial β-catenin signaling. Our data demonstrating a crucial role for Fgf10/Fgfr2b signaling in regulating proximal-distal patterning of the developing airway epithelium is in accordance with several previous reports. Hypomorphic Fgf10 mutant lungs show a reduction in distal epithelial markers and decreased epithelial β-catenin signaling (Ramasamy et al., 2007). Conditional inactivation of Fgf10 in the mesenchyme or its receptor Fgfr2b in the epithelium leads to downregulation of the distal epithelial marker Sox9, and the expansion of the proximal marker Sox2 (Abler et al., 2009). Finally, Fgf10 overexpression has been shown to inhibit terminal differentiation of the lung epithelium causing these cells to adopt a distal fate (Nyeng et al., 2008) or induction of adenomatous malformations (Clark et al., 2001). Our current data confirm and expand on some of these earlier findings.
The lung has a complex three-dimensional structure that features major differences along its proximodistal axis in terms of the composition of the endoderm-derived epithelium. The trachea and primary lung buds arise by different morphogenetic processes from contiguous regions of the embryonic foregut (Cardoso and Lü, 2006). A distinguishing feature of the adult mouse cartilaginous airways (i.e. trachea and primary bronchi) is that Fgf10 is expressed in the mesenchyme between the cartilage rings (Tiozzo et al., 2009; Sala et al., 2011) and that they contain a discontinuous population of basal stem cells that express p63 and specific keratins (K14 and K5) (Rock et al., 2010). The more distal airways (small bronchi and bronchioles) lack basal cells and are surrounded by airway smooth muscle, which does not express Fgf10 during normal homeostasis (Mailleux et al., 2005). In the cartilaginous airways, basal cells are considered to be on top of the stem cell hierarchy and are able to self renew and give rise to Clara cells, goblet cells and ciliated cells (Rock et al., 2009). Clara cells themselves are also considered to be stem cells and during normal homeostasis they can give rise to new Clara cells and terminally differentiated ciliated cells (Evans et al., 1976; Rawlins et al., 2009b). Interestingly, p63 is a master regulator required for the development of basal cells (Daniely et al., 2004) and it induces a basal cell phenotype and squamous metaplasia when ectopically expressed in Clara cells (Koster et al., 2004). This form of Clara cell reprogramming may occur to some extent after airway epithelial injury, as Clara cells under such conditions have been shown to be able to give rise to basal cells (Rawlins et al., 2009b).
Very little is known about how basal cells arise during lung development. Our data indicate that blocking Wnt signaling during the very early stages of lung development results in a profound ‘proximalization’ of the lung epithelium and increased basal cell differentiation. Blocking Wnt signaling is likely to result in increased Sox2 expression, which is a positive regulator of p63 expression (Hashimoto et al., 2012).
In addition, we have discovered a new important role for Fgf10 signaling in the differentiation of Sox2-expressing airway epithelial cells, by preventing ciliated cell differentiation and promoting basal cell differentiation. Fgf10 probably prevents ciliated cell differentiation through activation of the Notch pathway in Clara cells (Guseh et al., 2009; Tsao et al., 2009; Morimoto et al., 2010; Morimoto et al., 2012), which prevents ciliated cell differentiation, similar to our observations in the adult lung (Volckaert et al., 2011). The mechanism through which Fgf10 promotes basal cell differentiation is still unclear.
These findings have potentially important consequences with regard to the role of basal cells and Fgf10 signaling in adult lung injury and repair. Basal cells are not present during normal homeostasis in the adult mouse distal conducting airway epithelium, but can be found only in the trachea and main stem bronchi (Rock et al., 2010). However, after H1N1 infection, ‘distal airway stem cells’ (DASCs) with basal cell characteristics can be found in the distal mouse airway and have been implicated in the regeneration of the alveolar epithelium (Kumar et al., 2011). Interestingly, a recent study using direct lineage tracing of ATII cells after bleomycin-induced lung injury provides evidence that the majority of the newly generated ATII cells are not derived from the pre-existing ATII cells (Chapman et al., 2011). In support of this, it was shown by lineage tracing that distal airway Clara stem cells can regenerate alveolar epithelium after catastrophic bleomycin- and H1N1-mediated injury, giving rise to (most of the) newly generated ATII cells (Rock et al., 2011; Chen et al., 2012; Zheng et al., 2012). We have previously demonstrated an important role for Fgf10 secreted by airway smooth muscle cells in the regeneration of the airway epithelium after naphthalene-mediated epithelial injury (Volckaert et al., 2011), suggesting that Fgf10 secreted from airway smooth muscle after injury might promote basal cell differentiation/amplification after injury and thus generate the stem cells required to regenerate the injured lungs. Future experiments will be required to test this hypothesis.
Supplementary Material
Footnotes
Funding
These studies were supported by funding from the National Institutes of Health [HL092967 to S.D.L.]. P.M. is Hastings Professor of Pediatrics. Deposited in PMC for release after 12 months.
Competing interests statement
The authors declare no competing financial interests.
Author contributions
This project was conceived and executed by S.D.L. Experiments were designed by T.V. and S.D.L. T.V. and S.D.L. wrote the manuscript. T.V., C.L., P.M. and S.D.L. edited the manuscript. T.V., A.C., E.D. and S.D.L. performed all experiments and characterizations of the phenotypes. C.L., P.M. and S.D.L. generated Tet-Dkk1 mice.
Supplementary material
Supplementary material available online at http://dev.biologists.org/lookup/suppl/doi:10.1242/dev.096560/-/DC1
References
- Abler L. L., Mansour S. L., Sun X. (2009). Conditional gene inactivation reveals roles for Fgf10 and Fgfr2 in establishing a normal pattern of epithelial branching in the mouse lung. Dev. Dyn. 238, 1999–2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ang S. L., Wierda A., Wong D., Stevens K. A., Cascio S., Rossant J., Zaret K. S. (1993). The formation and maintenance of the definitive endoderm lineage in the mouse: involvement of HNF3/forkhead proteins. Development 119, 1301–1315 [DOI] [PubMed] [Google Scholar]
- Arman E., Haffner-Krausz R., Gorivodsky M., Lonai P. (1999). Fgfr2 is required for limb outgrowth and lung-branching morphogenesis. Proc. Natl. Acad. Sci. USA 96, 11895–11899 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bao J., Zheng J. J., Wu D. (2012). The structural basis of DKK-mediated inhibition of Wnt/LRP signaling. Sci. Signal. 5, pe22 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bellusci S., Grindley J., Emoto H., Itoh N., Hogan B. L. (1997). Fibroblast growth factor 10 (FGF10) and branching morphogenesis in the embryonic mouse lung. Development 124, 4867–4878 [DOI] [PubMed] [Google Scholar]
- Cardoso W. V., Lü J. (2006). Regulation of early lung morphogenesis: questions, facts and controversies. Development 133, 1611–1624 [DOI] [PubMed] [Google Scholar]
- Chapman H. A., Li X., Alexander J. P., Brumwell A., Lorizio W., Tan K., Sonnenberg A., Wei Y., Vu T. H. (2011). Integrin α6β4 identifies an adult distal lung epithelial population with regenerative potential in mice. J. Clin. Invest. 121, 2855–2862 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen F., Desai T. J., Qian J., Niederreither K., Lü J., Cardoso W. V. (2007). Inhibition of Tgf beta signaling by endogenous retinoic acid is essential for primary lung bud induction. Development 134, 2969–2979 [DOI] [PubMed] [Google Scholar]
- Chen F., Cao Y., Qian J., Shao F., Niederreither K., Cardoso W. V. (2010). A retinoic acid-dependent network in the foregut controls formation of the mouse lung primordium. J. Clin. Invest. 120, 2040–2048 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen H., Matsumoto K., Brockway B. L., Rackley C. R., Liang J., Lee J. H., Jiang D., Noble P. W., Randell S. H., Kim C. F., et al. (2012). Airway epithelial progenitors are region specific and show differential responses to bleomycin-induced lung injury. Stem Cells 30, 1948–1960 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clark J. C., Tichelaar J. W., Wert S. E., Itoh N., Perl A. K., Stahlman M. T., Whitsett J. A. (2001). FGF-10 disrupts lung morphogenesis and causes pulmonary adenomas in vivo. Am. J. Physiol. 280, L705–L715 [DOI] [PubMed] [Google Scholar]
- Cohen E. D., Ihida-Stansbury K., Lu M. M., Panettieri R. A., Jones P. L., Morrisey E. E. (2009). Wnt signaling regulates smooth muscle precursor development in the mouse lung via a tenascin C/PDGFR pathway. J. Clin. Invest. 119, 2538–2549 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daniely Y., Liao G., Dixon D., Linnoila R. I., Lori A., Randell S. H., Oren M., Jetten A. M. (2004). Critical role of p63 in the development of a normal esophageal and tracheobronchial epithelium. Am. J. Physiol. 287, C171–C181 [DOI] [PubMed] [Google Scholar]
- DasGupta R., Fuchs E. (1999). Multiple roles for activated LEF/TCF transcription complexes during hair follicle development and differentiation. Development 126, 4557–4568 [DOI] [PubMed] [Google Scholar]
- De Langhe S. P., Sala F. G., Del Moral P. M., Fairbanks T. J., Yamada K. M., Warburton D., Burns R. C., Bellusci S. (2005). Dickkopf-1 (DKK1) reveals that fibronectin is a major target of Wnt signaling in branching morphogenesis of the mouse embryonic lung. Dev. Biol. 277, 316–331 [DOI] [PubMed] [Google Scholar]
- De Langhe S. P., Carraro G., Tefft D., Li C., Xu X., Chai Y., Minoo P., Hajihosseini M. K., Drouin J., Kaartinen V., et al. (2008). Formation and differentiation of multiple mesenchymal lineages during lung development is regulated by beta-catenin signaling. PLoS One 3, e1516 [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Moerlooze L., Spencer-Dene B., Revest J. M., Hajihosseini M., Rosewell I., Dickson C. (2000). An important role for the IIIb isoform of fibroblast growth factor receptor 2 (FGFR2) in mesenchymal-epithelial signalling during mouse organogenesis. Development 127, 483–492 [DOI] [PubMed] [Google Scholar]
- Desai T. J., Malpel S., Flentke G. R., Smith S. M., Cardoso W. V. (2004). Retinoic acid selectively regulates Fgf10 expression and maintains cell identity in the prospective lung field of the developing foregut. Dev. Biol. 273, 402–415 [DOI] [PubMed] [Google Scholar]
- Desai T. J., Chen F., Lü J., Qian J., Niederreither K., Dollé P., Chambon P., Cardoso W. V. (2006). Distinct roles for retinoic acid receptors alpha and beta in early lung morphogenesis. Dev. Biol. 291, 12–24 [DOI] [PubMed] [Google Scholar]
- Domyan E. T., Sun X. (2011). Patterning and plasticity in development of the respiratory lineage. Dev. Dyn. 240, 477–485 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evans M. J., Johnson L. V., Stephens R. J., Freeman G. (1976). Renewal of the terminal bronchiolar epithelium in the rat following exposure to NO2 or O3. Lab. Invest. 35, 246–257 [PubMed] [Google Scholar]
- Finch P. W., Cunha G. R., Rubin J. S., Wong J., Ron D. (1995). Pattern of keratinocyte growth factor and keratinocyte growth factor receptor expression during mouse fetal development suggests a role in mediating morphogenetic mesenchymal-epithelial interactions. Dev. Dyn. 203, 223–240 [DOI] [PubMed] [Google Scholar]
- Goss A. M., Tian Y., Tsukiyama T., Cohen E. D., Zhou D., Lu M. M., Yamaguchi T. P., Morrisey E. E. (2009). Wnt2/2b and beta-catenin signaling are necessary and sufficient to specify lung progenitors in the foregut. Dev. Cell 17, 290–298 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goss A. M., Tian Y., Cheng L., Yang J., Zhou D., Cohen E. D., Morrisey E. E. (2011). Wnt2 signaling is necessary and sufficient to activate the airway smooth muscle program in the lung by regulating myocardin/Mrtf-B and Fgf10 expression. Dev. Biol. 356, 541–552 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guseh J. S., Bores S. A., Stanger B. Z., Zhou Q., Anderson W. J., Melton D. A., Rajagopal J. (2009). Notch signaling promotes airway mucous metaplasia and inhibits alveolar development. Development 136, 1751–1759 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harris-Johnson K. S., Domyan E. T., Vezina C. M., Sun X. (2009). beta-Catenin promotes respiratory progenitor identity in mouse foregut. Proc. Natl. Acad. Sci. USA 106, 16287–16292 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hashimoto S., Chen H., Que J., Brockway B. L., Drake J. A., Snyder J. C., Randell S. H., Stripp B. R. (2012). β-Catenin-SOX2 signaling regulates the fate of developing airway epithelium. J. Cell Sci. 125, 932–942 [DOI] [PMC free article] [PubMed] [Google Scholar]
- He X. C., Yin T., Grindley J. C., Tian Q., Sato T., Tao W. A., Dirisina R., Porter-Westpfahl K. S., Hembree M., Johnson T., et al. (2007). PTEN-deficient intestinal stem cells initiate intestinal polyposis. Nat. Genet. 39, 189–198 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hokuto I., Perl A. K., Whitsett J. A. (2003). Prenatal, but not postnatal, inhibition of fibroblast growth factor receptor signaling causes emphysema. J. Biol. Chem. 278, 415–421 [DOI] [PubMed] [Google Scholar]
- Kelly R. G., Brown N. A., Buckingham M. E. (2001). The arterial pole of the mouse heart forms from Fgf10-expressing cells in pharyngeal mesoderm. Dev. Cell 1, 435–440 [DOI] [PubMed] [Google Scholar]
- Kimura S., Hara Y., Pineau T., Fernandez-Salguero P., Fox C. H., Ward J. M., Gonzalez F. J. (1996). The T/ebp null mouse: thyroid-specific enhancer-binding protein is essential for the organogenesis of the thyroid, lung, ventral forebrain, and pituitary. Genes Dev. 10, 60–69 [DOI] [PubMed] [Google Scholar]
- Koster M. I., Kim S., Mills A. A., DeMayo F. J., Roop D. R. (2004). p63 is the molecular switch for initiation of an epithelial stratification program. Genes Dev. 18, 126–131 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krejci P., Aklian A., Kaucka M., Sevcikova E., Prochazkova J., Masek J. K., Mikolka P., Pospisilova T., Spoustova T., Weis M., et al. (2012). Receptor tyrosine kinases activate canonical WNT/β-catenin signaling via MAP kinase/LRP6 pathway and direct β-catenin phosphorylation. PLoS ONE 7, e35826 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar P. A., Hu Y., Yamamoto Y., Hoe N. B., Wei T. S., Mu D., Sun Y., Joo L. S., Dagher R., Zielonka E. M., et al. (2011). Distal airway stem cells yield alveoli in vitro and during lung regeneration following H1N1 influenza infection. Cell 147, 525–538 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lazzaro D., Price M., de Felice M., Di Lauro R. (1991). The transcription factor TTF-1 is expressed at the onset of thyroid and lung morphogenesis and in restricted regions of the foetal brain. Development 113, 1093–1104 [DOI] [PubMed] [Google Scholar]
- Lü J., Izvolsky K. I., Qian J., Cardoso W. V. (2005). Identification of FGF10 targets in the embryonic lung epithelium during bud morphogenesis. J. Biol. Chem. 280, 4834–4841 [DOI] [PubMed] [Google Scholar]
- Mailleux A. A., Tefft D., Ndiaye D., Itoh N., Thiery J. P., Warburton D., Bellusci S. (2001). Evidence that SPROUTY2 functions as an inhibitor of mouse embryonic lung growth and morphogenesis. Mech. Dev. 102, 81–94 [DOI] [PubMed] [Google Scholar]
- Mailleux A. A., Kelly R., Veltmaat J. M., De Langhe S. P., Zaffran S., Thiery J. P., Bellusci S. (2005). Fgf10 expression identifies parabronchial smooth muscle cell progenitors and is required for their entry into the smooth muscle cell lineage. Development 132, 2157–2166 [DOI] [PubMed] [Google Scholar]
- Metzger R. J., Klein O. D., Martin G. R., Krasnow M. A. (2008). The branching programme of mouse lung development. Nature 453, 745–750 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Min H., Danilenko D. M., Scully S. A., Bolon B., Ring B. D., Tarpley J. E., DeRose M., Simonet W. S. (1998). Fgf-10 is required for both limb and lung development and exhibits striking functional similarity to Drosophila branchless. Genes Dev. 12, 3156–3161 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Minoo P., Su G., Drum H., Bringas P., Kimura S. (1999). Defects in tracheoesophageal and lung morphogenesis in Nkx2.1(-/-) mouse embryos. Dev. Biol. 209, 60–71 [DOI] [PubMed] [Google Scholar]
- Monkley S. J., Delaney S. J., Pennisi D. J., Christiansen J. H., Wainwright B. J. (1996). Targeted disruption of the Wnt2 gene results in placentation defects. Development 122, 3343–3353 [DOI] [PubMed] [Google Scholar]
- Morimoto M., Liu Z., Cheng H. T., Winters N., Bader D., Kopan R. (2010). Canonical Notch signaling in the developing lung is required for determination of arterial smooth muscle cells and selection of Clara versus ciliated cell fate. J. Cell Sci. 123, 213–224 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morimoto M., Nishinakamura R., Saga Y., Kopan R. (2012). Different assemblies of Notch receptors coordinate the distribution of the major bronchial Clara, ciliated and neuroendocrine cells. Development 139, 4365–4373 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mucenski M. L., Wert S. E., Nation J. M., Loudy D. E., Huelsken J., Birchmeier W., Morrisey E. E., Whitsett J. A. (2003). beta-Catenin is required for specification of proximal/distal cell fate during lung morphogenesis. J. Biol. Chem. 278, 40231–40238 [DOI] [PubMed] [Google Scholar]
- Nyeng P., Norgaard G. A., Kobberup S., Jensen J. (2008). FGF10 maintains distal lung bud epithelium and excessive signaling leads to progenitor state arrest, distalization, and goblet cell metaplasia. BMC Dev. Biol. 8, 2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohtsuka N., Urase K., Momoi T., Nogawa H. (2001). Induction of bud formation of embryonic mouse tracheal epithelium by fibroblast growth factor plus transferrin in mesenchyme-free culture. Dev. Dyn. 222, 263–272 [DOI] [PubMed] [Google Scholar]
- Ohuchi H., Nakagawa T., Yamamoto A., Araga A., Ohata T., Ishimaru Y., Yoshioka H., Kuwana T., Nohno T., Yamasaki M., et al. (1997). The mesenchymal factor, FGF10, initiates and maintains the outgrowth of the chick limb bud through interaction with FGF8, an apical ectodermal factor. Development 124, 2235–2244 [DOI] [PubMed] [Google Scholar]
- Okubo T., Hogan B. L. (2004). Hyperactive Wnt signaling changes the developmental potential of embryonic lung endoderm. J. Biol. 3, 11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ornitz D. M., Yin Y. (2012). Signaling networks regulating development of the lower respiratory tract. Cold Spring Harb. Perspect. Biol. 4, a008318 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park W. Y., Miranda B., Lebeche D., Hashimoto G., Cardoso W. V. (1998). FGF-10 is a chemotactic factor for distal epithelial buds during lung development. Dev. Biol. 201, 125–134 [DOI] [PubMed] [Google Scholar]
- Pepicelli C. V., Lewis P. M., McMahon A. P. (1998). Sonic hedgehog regulates branching morphogenesis in the mammalian lung. Curr. Biol. 8, 1083–1086 [DOI] [PubMed] [Google Scholar]
- Que J., Okubo T., Goldenring J. R., Nam K. T., Kurotani R., Morrisey E. E., Taranova O., Pevny L. H., Hogan B. L. (2007). Multiple dose-dependent roles for Sox2 in the patterning and differentiation of anterior foregut endoderm. Development 134, 2521–2531 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rajagopal J., Carroll T. J., Guseh J. S., Bores S. A., Blank L. J., Anderson W. J., Yu J., Zhou Q., McMahon A. P., Melton D. A. (2008). Wnt7b stimulates embryonic lung growth by coordinately increasing the replication of epithelium and mesenchyme. Development 135, 1625–1634 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramasamy S. K., Mailleux A. A., Gupte V. V., Mata F., Sala F. G., Veltmaat J. M., Del Moral P. M., De Langhe S., Parsa S., Kelly L. K., et al. (2007). Fgf10 dosage is critical for the amplification of epithelial cell progenitors and for the formation of multiple mesenchymal lineages during lung development. Dev. Biol. 307, 237–247 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rawlins E. L., Clark C. P., Xue Y., Hogan B. L. (2009a). The Id2+ distal tip lung epithelium contains individual multipotent embryonic progenitor cells. Development 136, 3741–3745 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rawlins E. L., Okubo T., Xue Y., Brass D. M., Auten R. L., Hasegawa H., Wang F., Hogan B. L. (2009b). The role of Scgb1a1+ Clara cells in the long-term maintenance and repair of lung airway, but not alveolar, epithelium. Cell Stem Cell 4, 525–534 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rock J. R., Onaitis M. W., Rawlins E. L., Lu Y., Clark C. P., Xue Y., Randell S. H., Hogan B. L. (2009). Basal cells as stem cells of the mouse trachea and human airway epithelium. Proc. Natl. Acad. Sci. USA 106, 12771–12775 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rock J. R., Randell S. H., Hogan B. L. (2010). Airway basal stem cells: a perspective on their roles in epithelial homeostasis and remodeling. Dis. Model. Mech. 3, 545–556 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rock J. R., Barkauskas C. E., Cronce M. J., Xue Y., Harris J. R., Liang J., Noble P. W., Hogan B. L. (2011). Multiple stromal populations contribute to pulmonary fibrosis without evidence for epithelial to mesenchymal transition. Proc. Natl. Acad. Sci. USA 108, E1475–E1483 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sala F. G., Del Moral P. M., Tiozzo C., Alam D. A., Warburton D., Grikscheit T., Veltmaat J. M., Bellusci S. (2011). FGF10 controls the patterning of the tracheal cartilage rings via Shh. Development 138, 273–282 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sekine K., Ohuchi H., Fujiwara M., Yamasaki M., Yoshizawa T., Sato T., Yagishita N., Matsui D., Koga Y., Itoh N., et al. (1999). Fgf10 is essential for limb and lung formation. Nat. Genet. 21, 138–141 [DOI] [PubMed] [Google Scholar]
- Shu W., Guttentag S., Wang Z., Andl T., Ballard P., Lu M. M., Piccolo S., Birchmeier W., Whitsett J. A., Millar S. E., et al. (2005). Wnt/beta-catenin signaling acts upstream of N-myc, BMP4, and FGF signaling to regulate proximal-distal patterning in the lung. Dev. Biol. 283, 226–239 [DOI] [PubMed] [Google Scholar]
- Tiozzo C., De Langhe S., Carraro G., Alam D. A., Nagy A., Wigfall C., Hajihosseini M. K., Warburton D., Minoo P., Bellusci S. (2009). Fibroblast growth factor 10 plays a causative role in the tracheal cartilage defects in a mouse model of Apert syndrome. Pediatr. Res. 66, 386–390 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsao P. N., Vasconcelos M., Izvolsky K. I., Qian J., Lu J., Cardoso W. V. (2009). Notch signaling controls the balance of ciliated and secretory cell fates in developing airways. Development 136, 2297–2307 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Volckaert T., Dill E., Campbell A., Tiozzo C., Majka S., Bellusci S., De Langhe S. P. (2011). Parabronchial smooth muscle constitutes an airway epithelial stem cell niche in the mouse lung after injury. J. Clin. Invest. 121, 4409–4419 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Warburton D. (2008). Developmental biology: order in the lung. Nature 453, 733–735 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weaver M., Dunn N. R., Hogan B. L. (2000). Bmp4 and Fgf10 play opposing roles during lung bud morphogenesis. Development 127, 2695–2704 [DOI] [PubMed] [Google Scholar]
- White A. C., Xu J., Yin Y., Smith C., Schmid G., Ornitz D. M. (2006). FGF9 and SHH signaling coordinate lung growth and development through regulation of distinct mesenchymal domains. Development 133, 1507–1517 [DOI] [PubMed] [Google Scholar]
- Yin Y., White A. C., Huh S. H., Hilton M. J., Kanazawa H., Long F., Ornitz D. M. (2008). An FGF-WNT gene regulatory network controls lung mesenchyme development. Dev. Biol. 319, 426–436 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yin Y., Wang F., Ornitz D. M. (2011). Mesothelial- and epithelial-derived FGF9 have distinct functions in the regulation of lung development. Development 138, 3169–3177 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zakin L. D., Mazan S., Maury M., Martin N., Guénet J. L., Brûlet P. (1998). Structure and expression of Wnt13, a novel mouse Wnt2 related gene. Mech. Dev. 73, 107–116 [DOI] [PubMed] [Google Scholar]
- Zheng D., Limmon G. V., Yin L., Leung N. H., Yu H., Chow V. T., Chen J. (2012). Regeneration of alveolar type I and II cells from Scgb1a1-expressing cells following severe pulmonary damage induced by bleomycin and influenza. PLoS ONE 7, e48451 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.