

Abstract
Hac1 is the activator of the cellular response to the accumulation of unfolded proteins in the endoplasmic reticulum. Hac1 function requires the activity of Gcn4, which mainly acts as a regulator of the general amino acid control network providing Saccharomyces cerevisiae cells with amino acids. Here, we demonstrate novel functions of Hac1 and describe a mutual connection between Hac1 and Gcn4. Hac1 is required for induction of Gcn4-responsive promoter elements in haploid as well as diploid cells and therefore participates in the cellular amino acid supply. Furthermore, Hac1 and Gcn4 mutually influence their mRNA expression levels. Hac1 is also involved in FLO11 expression and adhesion upon amino acid starvation. Hac1 and Gcn4 act through the same promoter regions of the FLO11 flocculin. The results indicate an indirect effect of both transcription factors on FLO11 expression. Our data suggest a complex mutual cross talk between the Hac1- and Gcn4-controlled networks.
INTRODUCTION
The baker's yeast Saccharomyces cerevisiae executes two well-established pathways, the general amino acid control (GAAC) and the unfolded protein response (UPR). The GAAC regulatory network is induced not only by amino acid starvation or imbalances but also by other environmental stimuli, including supply of glucose (1), purines (2), and tRNA synthetases (3). A variety of physiological or environmental stress conditions such as calcium depletion, glucose deprivation, hypoxia, or misfolded proteins lead to an accumulation of misfolded or unfolded proteins in the endoplasmic reticulum (ER) lumen, which results in the induction of the UPR (4–7). These pathways are conserved in mammals, where they are essential.
The bZIP transcription factor Gcn4 represents the global key activator of the GAAC (8) and regulates transcription of numerous metabolic genes of amino acid or purine biosynthesis in response to amino acid starvation (9–12). In contrast to its mammalian homologues, yeast Gcn4 can bind only as a homodimer to a specific 9-bp palindromic nucleotide sequence (5′-ATGA[C/G]TCAT-3′) (termed Gcn4 protein recognition element [GCRE]) (13, 14). These GCREs are located upstream of many genes induced by amino acid starvation. Gcn4 can also bind to naturally occurring variants of this sequence (TGATTCA, TGACTCT, TGACTGA, TGACTAT, and ATGACTCT), and therefore, using a computer algorithm, this consensus site was generalized to RRRWGASTCA (R = purine, W = T or A, and S = G or C) (9). Gcn4 also binds to GCRE half-sites with high affinity in vitro (15, 16).
Gcn4 not only acts as a metabolic regulator but also has a developmental function. In response to nutrient starvation, Gcn4 is involved in the regulation of FLO11 expression (17, 18). The cell surface flocculin Flo11, also named Muc1, is required for diploid pseudohypha formation and for adhesion upon nutrient starvation (19–22).
Hac1 plays a central role in the yeast UPR system and represents a bZIP transcription factor, like Gcn4 (23, 24). Conserved from yeast to mammals is the sensing and response pathway that is transduced by Ire1, leading to an upregulation of transcription levels of approximately 400 genes, i.e., 7% to 8% of the yeast genome (25–30). In S. cerevisiae, Ire1 senses the stress and mediates a signaling cascade to upregulate responsive genes through unconventional splicing of HAC1 mRNA. Ire1 encodes a bifunctional transmembrane kinase/endoribonuclease consisting of an unfolded protein sensor domain in the ER lumen, a transmembrane domain, and a cytosolic effector domain, which contains an intrinsic serine/threonine kinase as well as an endoribonuclease in its C terminus (26, 27, 31–33). An accumulation of misfolded proteins in response to ER stress leads to oligomerization and trans-autophosphorylation of Ire1 (34, 35). This in turn results in an activated cytosolic endonuclease effector domain (33, 36). Ire1 recognizes two “loop” structures in the HAC1 mRNA. The transcript is constitutively synthesized as a precursor bearing a 252-nucleotide intron that blocks translation, and the endonuclease effector domain of Ire1 splices the HAC1 mRNA (37–39). Subsequently, the tRNA ligase Rlg1 religates, causing exons to produce the mature, efficiently translated HAC1 mRNA (33, 38). As the level of Hac1 rises in the cell, the genes that harbor unfolded protein response elements (UPREs) within their promoters are induced at the transcriptional level (40).
The synthesis of Hac1 in response to ER stress is regulated not only at its translational level but also by mechanisms that regulate the rate of turnover of Hac1. A similar mechanism had been described previously for the bZIP transcription factor Gcn4 (41–43). Like Gcn4, Hac1 is ubiquitinated by the SCFCdc4 E3 ligase complex, resulting in degradation by the 26S proteasome. Furthermore, phosphorylation by the cyclin-dependent kinase (CDK) Srb10 marks Hac1 for ubiquitination, similarly to Gcn4, whereas phosphorylation by the CDK Pho85 was not observed so far. Hac1 also contains a PEST region, which is typical for rapid turnover of transcription factors (44).
At least 381 UPR target genes were identified in yeast and encode functions ranging from protein folding, protein translocation, and protein transport to protein degradation within the secretory pathway. Whereas the predicted UPRE-1 consensus sequence (CAGNGTG) was absent in most of them (25), Patil and coworkers identified two further UPREs, which are recognized by Hac1 (UPRE-2, consensus sequence TACGTG; UPRE-3, consensus sequence AGGAACAAC) (45). Apart from its role as a transcriptional activator of the GAAC, Gcn4 and its activator Gcn2 are required for induction of a majority of UPR target genes upon ER stress. A direct binding of Gcn4 could be demonstrated for UPRE-1 and UPRE-2, while binding to UPRE-1 was Hac1 dependent. In contrast, Gcn4 is not necessary for the regulation of genes without a recognizable UPRE, which represent half of all UPR targets. Both Hac1 and Gcn4 are bZIP transcription factors. Heterodimer formation of Gcn4/Hac1 or its mammalian counterparts ATF4/XBP1 is an attractive hypothesis to explain the mechanism (45) but could not be verified yet. Recently, Fordyce and coworkers discovered that Hac1 binds only to UPRE-2 but not to the 7-bp UPRE-1 sequence, hereafter termed core UPRE-1 (cUPRE-1) (46). However, they could demonstrate that an extended core UPRE-1 (xcUPRE-1) containing flanking sequences is important for Hac1 binding. Therefore, the 7-bp cUPRE-1 consensus sequence can be extended to a 12-bp UPRE-1-like motif (GGACAGCGTGTC).
In this study, we identified novel functions of Hac1 in metabolic and developmental processes regulated by Gcn4. We demonstrate not only that Gcn4 is able to activate Hac1-specific target genes but also that Hac1 is involved in Gcn4-specific target gene regulation and FLO11 expression in response to amino acid starvation.
MATERIALS AND METHODS
Yeast strains and growth conditions.
All yeast strains used in this study are listed in Table 1. They are derivates of the S. cerevisiae strain background Σ1278b unless otherwise stated. Transformations were carried out using the lithium acetate method (47).
Table 1.
Saccharomyces cerevisiae strains used in this study
| Strain | Genotype | Source |
|---|---|---|
| BY4741 | MATa his3::hisG leu2::hisG met15::hisG ura3::hisG (S288c background) | 48 |
| Y05650 | MATa his3::hisG leu2::hisG met15::hisG ura3::hisG Δhac1::kanMX4 (S288c background) | 48 |
| RH2398 | MATa/α ura3-52/ura3-52::GCRE6::lacZ::URA3 leu2::hisG/leu2::hisG his3::hisG/HIS3 trp1::hisG/trp1::hisG Δgcn4::LEU2/Δgcn4::LEU2 | 58 |
| RH2656 | MATa/α ura3-52/ura3-52 trp1::hisG/TRP1 | 18 |
| RH2658 | MATa/α ura3-52/ura3-52 leu2::hisG/leu2::hisG trp1::hisG/TRP1 Δgcn4::LEU2/Δgcn4::LEU2 | 18 |
| RH2661 | MATa/α ura3-52/ura3-52 trp1::hisG/TRP1 Δflo11::kanMX4/Δflo11::kanMX4 | 18 |
| RH2676 | MATa ura3-52 leu2::hisG trp1::hisG Δgcn4::LEU2 | Our collection |
| RH2681 | MATa ura3-52 trp1::hisG Δflo11::kanMX4 | Our collection |
| RH2694 | MATa/α ura3-52/ura3-52 leu2::hisG/leu2::hisG trp1::hisG/trp1::hisG Δgcn4::LEU2/Δgcn4::LEU2 | Our collection |
| RH2695 | MATa/α ura3-52/ura3-52::FLO11::lacZ::URA3 leu2::hisG/leu2::hisG trp1::hisG/trp1::hisG Δgcn4::LEU2/Δgcn4::LEU2 | Our collection |
| RH2816 | MATa ura3-52 trp1::hisG his3::hisG::HIS3 | 72 |
| RH2819 | MATα ura3-52 leu2-hisG his3::hisG::HIS3 | 72 |
| RH3349 | MATa/α ura3-52/ura3-52::FLO11::lacZ::URA3 leu2::hisG/leu2::hisG trp1::hisG/trp1::hisG Δgcn4::LEU2/Δgcn4::LEU2 Δhac1::kanMX4/Δhac1::kanMX4::natMX4 | This study |
| RH3350 | MATa/α ura3-52/ura3-52::GCRE6::lacZ::URA3 leu2::hisG/leu2::hisG trp1::hisG/trp1::hisG Δgcn4::LEU2/Δgcn4::LEU2 Δhac1::kanMX4/Δhac1::kanMX4::natMX4 | This study |
| RH3351 | MATa ura3-52 trp1::hisG Δhac1::kanMX4 | This study |
| RH3352 | MATα ura3-52 leu2::hisG Δhac1::kanMX4 | This study |
| RH3360 | MATa ura3-52::FLO11::lacZ::URA3 trp1::hisG Δhac1::kanMX4 | This study |
| RH3362 | MATa/α ura3-52/ura3-52::FLO11::lacZ::URA3 leu2::hisG/LEU2 trp1::hisG/TRP1 Δhac1::kanMX4/Δhac1::kanMX4 | This study |
| RH3363 | MATa ura3-52::GCRE6::lacZ::URA3 trp1::hisG Δhac1::kanMX4 | This study |
| RH3401 | MATα ura3-52 leu2::hisG trp1::hisG Δgcn4::LEU2 | Our collection |
| RH3402 | MATa ura3-52 leu2::hisG trp1::hisG Δgcn4::LEU2 Δhac1::kanMX4 | This study |
| RH3403 | MATα ura3-52 leu2::hisG trp1::hisG Δgcn4::LEU2 Δhac1::kanMX4 | This study |
| RH3404 | MATα ura3-52 leu2::hisG Δhac1::kanMX4::natMX4 | This study |
| RH3405 | MATα ura3-52 leu2::hisG trp1::hisG Δgcn4::LEU2 Δhac1::kanMX4::natMX4 | This study |
| RH3406 | MATa ura3-52::FLO11::lacZ::URA3 trp1::hisG | This study |
| RH3407 | MATa ura3-52::FLO11::lacZ::URA3 leu2::hisG trp1::hisG Δgcn4::LEU2 | This study |
| RH3408 | MATa ura3-52::FLO11::lacZ::URA3 leu2::hisG trp1::hisG Δgcn4::LEU2 Δhac1::kanMX4 | This study |
| RH3409 | MATa ura3-52::GCRE6::lacZ::URA3 trp1::hisG | This study |
| RH3410 | MATa ura3-52::GCRE6::lacZ::URA3 leu2::hisG trp1::hisG Δgcn4::LEU2 | This study |
| RH3411 | MATa ura3-52::GCRE6::lacZ::URA3 leu2::hisG trp1::hisG Δgcn4::LEU2 Δhac1::kanMX4 | This study |
| RH3412 | MATa/α ura3-52/ura3-52 leu2::hisG/LEU2 trp1::hisG/TRP1 Δhac1::kanMX4/Δhac1::kanMX4::natMX4 | This study |
| RH3413 | MATa/α ura3152/ura3-52 leu2::hisG/LEU2 trp1::hisG/TRP1 Δhac1::kanMX4/HAC1 | This study |
| RH3414 | MATa/α ura3-52/ura3-52 leu2::hisG/leu2::hisG trp1::hisG/TRP1 Δgcn4::LEU2/GCN4 | This study |
| RH3415 | MATa/α ura3-52/ura3-52 leu2::hisG/leu2::hisG trp1::hisG/trp1::hisG Δgcn4::LEU2/Δgcn4::LEU2 Δhac1::kanMX4/Δhac1::kanMX4::natMX4 | This study |
| RH3416 | MATa/α ura3-52/ura3-52 leu2::hisG/leu2::hisG trp1::hisG/TRP1 Δgcn4::LEU2/GCN4 Δhac1::kanMX4/HAC1 | This study |
| RH3417 | MATa/α ura3-52/ura3-52::FLO11::lacZ::URA3 leu2::hisG/LEU2 trp1::hisG/TRP1 | This study |
| RH3418 | MATa/α ura3-52/ura3-52::FLO11::lacZ::URA3 leu2::hisG/LEU2 trp1::hisG/TRP1 Δhac1::kanMX4/HAC1 | This study |
| RH3419 | MATa/α ura3-52/ura3-52::FLO11::lacZ::URA3 leu2::hisG/leu2::hisG trp1::hisG/TRP1 Δgcn4::LEU2/GCN4 | This study |
| RH3420 | MATa/α ura3-52/ura3-52::FLO11::lacZ::URA3 leu2::hisG/leu2::hisG trp1::hisG/TRP1 Δgcn4::LEU2/GCN4 Δhac1::kanMX4/HAC1 | This study |
| RH3421 | MATa/α ura3-52/ura3-52::GCRE6::lacZ::URA3 leu2::hisG/LEU2 trp1::hisG/TRP1 | This study |
| RH3422 | MATa/α ura3-52/ura3-52::GCRE6::lacZ::URA3 leu2::hisG/LEU2 trp1::hisG/TRP1 Δhac1::kanMX4/Δhac1::kanMX4::natMX4 | This study |
| RH3423 | MATa/α ura3-52/ura3-52::GCRE6::lacZ::URA3 leu2::hisG/LEU2 trp1::hisG/TRP1 Δhac1::kanMX4/HAC1 | This study |
| RH3424 | MATa/α ura3-52/ura3-52::GCRE6::lacZ::URA3 leu2::hisG/leu2::hisG trp1::hisG/TRP1 Δgcn4::LEU2/GCN4 | This study |
| RH3425 | MATa/α ura3-52/ura3-52::GCRE6::lacZ::URA3 leu2::hisG/leu2::hisG trp1::hisG/TRP1 Δgcn4::LEU2/GCN4 Δhac1::kanMX4/HAC1 | This study |
Yeast strains RH3351, RH3352, RH3402, and RH3403 containing a hac1 deletion were constructed by amplification of the kanamycin resistance cassette from the Euroscarf strain collection (48) containing sequence homologous to the up- and downstream regions of the relevant gene. Integration of deletion cassettes was obtained by homologous recombination in yeast strains RH2676, RH2816, RH2819, and RH3401. Positive transformants could be selected on yeast extract-peptone-dextrose (YEPD) medium supplemented with 200 μg/ml Geneticin G418 sulfate (Carl Roth GmbH, Karlsruhe, Germany). For genetic crosses, the kanamycin resistance cassette of RH3352 and RH3403 was replaced by a nourseothricin resistance cassette, which was amplified from plasmid pAG25. Transformants (RH3404 and RH3405) could be selected on YEPD medium containing 100 μg/ml nourseothricin (clonNAT; Werner BioAgents, Jena, Germany). Homo- and heterozygous diploid strains RH3412 to RH3416 were obtained by crossing strain RH2676, RH3351, or RH3402 with strain RH2819, RH3404, or RH3405. Haploid yeast strains RH3360, RH3406, RH3407, and RH3408 were obtained by introducing the FLO11::lacZ::URA3 cassette using ApaI-linearized plasmid pME2213 into the URA3 locus of yeast strains RH2676, RH2816, RH3351, and RH3402. The GCRE6::lacZ::URA3 reporter gene cassette was introduced in the same four haploid yeast strains by transformation with StuI-linearized GCRE6::lacZ reporter construct pME1112 to obtain yeast strains RH3363, RH3409, RH3410, and RH3411. The haploid FLO11- and GCRE6::lacZ-containing MATa strains were crossed with MATα strain RH2819, RH3352, RH3404, or RH3405 to produce diploid strains RH3349, RH3350, RH3362, and RH3417 to RH3425. All gene deletions, integrations, or replacements were confirmed by PCR and Southern blot analysis (49).
Strains were routinely cultivated in standard yeast extract-peptone-dextrose (YEPD; 1% yeast extract, 2% peptone, 2% dextrose) or minimal yeast nitrogen base medium (YNB; 1,5 g/liter yeast nitrogen base lacking amino acids and ammonium sulfate, 5 g/liter ammonium sulfate, 2% dextrose supplemented with the appropriate amino acids). Solid media were prepared using 2% agar.
For β-galactosidase assays, strains were cultivated in liquid synthetic minimal medium (YNB) overnight at 30°C, diluted into fresh medium, and cultivated for 6 h before assaying enzymatic activities. For amino acid starvation, 3-amino-1,2,4-triazole (3AT) (Sigma-Aldrich, Steinheim, Germany) was added to fresh diluted cultures to a final concentration of 10 mM, and cells were incubated for 8 h before further assays. For nitrogen starvation, cells grown to logarithmic phase were washed twice with 2% glucose and incubated for 24 h in liquid YNB medium containing only 50 μM ammonium sulfate (instead of 50 mM) as the sole nitrogen source. Tunicamycin (Tm) (Calbiochem/Merck KGaA, Darmstadt, Germany) was added to fresh diluted cultures to a final concentration of 1 μg/ml and incubated for 6 h to induce UPR stress. Additionally, cultures grown to log phase (4 h or instead 6 h in YNB) were treated with 1 μg/ml Tm for 15, 30, 60, 90, and 120 min.
To compare strains under different conditions in Western hybridization experiments, strains were cultivated in 250 ml liquid synthetic minimal medium (YNB) to an optical density at 600 nm (OD600) of 0.6 to 0.8 at 30°C, subsequently divided into 50-ml cultures, and cultivated for a further 90 min under different conditions each.
Plasmids.
All plasmids used in this study are listed in Table 2. Plasmid pME3498 expressing the HAC1 inclusive intron under the MET25 promoter was constructed by amplifying HAC1 with PfuUltra HF DNA polymerase (Promega, Mannheim, Germany) from genomic DNA and introducing it as a XbaI/ClaI fragment into SpeI/ClaI-restricted p426MET25.
Table 2.
Plasmids used in this study
| Plasmid | Description | Source |
|---|---|---|
| B3782 | 3-kbp FLO11::lacZ in YEp355 | 53 |
| p180 | GCN4::lacZ reporter construct (uORFs 1 to 4) on a centromere vector (URA3) | 59 |
| p227 | GCN4::lacZ reporter construct (without uORFs) on a centromere vector (URA3) | 73 |
| pAG25 | NatMX4 cassette in pFA6 | 74 |
| p426MET25 | pRS426 containing MET25 promoter, CYC1 terminator | 75 |
| pFLO11-2/1 to pFLO11-15/14 | 440-bp sequence elements cloned into pLG669Z from bp −1 to −420, bp −180 to −620, and bp −380 to −1020 until bp −2580 to −2980 | 53 |
| pLG669Z | lacZ shuttle vector | 76 |
| pMCZ-Y | UPRE-CYC1-lacZ reporter construct on multicopy vector (URA) | 23 |
| pME1092 | 2.8-kb GCN4 fragment in pRS314 | 57 |
| pME1112 | Integrative GCRE6::lacZ reporter construct | 57 |
| pME2212 | pLG669Z without UASa | 18 |
| pME2213 | Integrative FLO11::lacZ reporter construct | Our collection |
| pME2901 | GCN4prom-GCN4L267S-GCN4-term in pRS314 | 17 |
| pME3378 | GCN4prom-GCN4L253G-GCN4-term in pRS314 | 17 |
| pME3498 | MET25prom-HAC1 in p426MET25 | This study |
| pRS314 | TRP CEN Ampr ori | 77 |
| pRS426 | URA3 2μm Ampr ori | 77 |
UAS, upstream activation sequence.
Protein analysis.
Whole-cell extracts of S. cerevisiae were prepared from yeast cultures grown to exponential phase. Cells were washed in 2.5 ml ice-cold buffer B (100 mM Tris-HCl, pH 7.5, 200 mM NaCl, 5 mM EDTA, 20% glycerin), lysed with glass beads (diameter, 0.25 to 0.5 mm; Carl Roth GmbH, Karlsruhe, Germany) in 500 μl of B-buffer-containing protease inhibitors (Complete, EDTA-free; Roche Diagnostics GmbH, Mannheim, Germany) and 14.3 mM β-mercaptoethanol at 4°C, and centrifuged at 13,000 rpm for 12 min to remove glass beads and large pieces of cell debris. Extracts (10 μl) were removed to determine total protein concentration using the Bradford protein assay (50), and proteins were denatured in SDS loading dye by heating at 65°C for 15 min. Equal amounts of protein were subjected to SDS-PAGE and transferred to nitrocellulose membranes. Hac1, the α subunit of eukaryotic initiation factor 2 (eIF2α), and eIF2α-P were detected using ECL technology (Amersham, United Kingdom). For the first incubation, polyclonal rabbit anti-Hac1 (gift from Kazutoshi Mori, Kyoto University, Japan), polyclonal rabbit anti-eIF2α (pS52) (catalog no. 44728G; Invitrogen, Darmstadt, Germany), or rabbit anti-eIF2α (gift from Thomas Dever, NIH, Bethesda, MD, USA) antibodies were used. Peroxidase-coupled goat anti-rabbit IgG was used as a secondary antibody (catalog no. G21234; MoBiTec, Göttingen, Germany).
Adhesive growth tests.
Amino acid starvation-induced adhesive growth tests on solid YNB medium were performed as described previously (18, 51). For visualization of biofilms in wells of polystyrene plates, assays were performed as described in reference 19.
Growth tests.
Yeast strains were precultured to the same optical densities (OD600 = 0.6) and then diluted 10-fold, starting with 3 × 104 cells per 20 μl. For each dilution, 20 μl was spotted onto solid YNB medium with or without 0.5 μg/ml tunicamycin for ER stress survival assays and on selective YNB medium with or without 5 mM 3AT for resistance upon amino acid starvation. After incubation for 3 to 4 days at 30°C, plates were photographed under white light.
β-Galactosidase assay in S. cerevisiae.
Starting from one overnight culture, strains carrying either a UPRE-, a FLO11-, a GCRE6-, or a GCN4::lacZ reporter were diluted into fresh medium and further cultivated for 6 to 24 h before they were harvested. The incubation term was based on the medium (for details, see “Yeast strains and growth conditions”). Extracts were prepared and assayed for specific β-galactosidase activity as described previously (52) and normalized to the total protein (50), resulting in the specific enzyme activity (OD420 × 0.35)/(0.0045 × protein concentration × extract volume × time). Assays were performed for at least three independent cultures.
Analysis of FLO11 promoter elements.
Rupp et al. (53) constructed a set of 14 reporter constructs containing individual 400-bp FLO11 promoter fragments that overlap by 200 bp and were cloned in front of a CYC1::lacZ fusion gene. Thus, after transformation of these constructs in the diploid wild-type strain as well as in Δhac1/Δhac1 and Δgcn4/Δgcn4 mutant strains, the influence of the transcription factors on specific regions in the FLO11 promoter can be determined by β-galactosidase assays. A construct without an insert served for background measurements.
RNA isolation and quantitative real-time PCR (qRT-PCR).
Total RNA was isolated from yeast cells that were grown in YNB in the presence (8 h) or absence (6 h) of 10 mM 3AT using the High Pure RNA isolation kit (Roche Diagnostics GmbH, Mannheim, Germany) to determine FLO11 transcript levels. For analyzing GCN4 or HAC1 mRNA expression levels, yeast cells were grown to an OD600 of ∼0.6 at 30°C before division into independent cultures and further cultivation for 90 min under indicated conditions. cDNA synthesis was performed in duplicate for each sample using 0.8 μg RNA and the QuantiTect reverse transcription kit (Qiagen, Hilden, Germany) according to the manufacturer's instructions.
Twenty nanograms of cDNA was used as the template for quantitative real-time PCR (qRT-PCR) experiments, and amplification was performed in a LightCycler 2.0 (Roche Diagnostics GmbH, Mannheim, Germany) using the RealMaster SYBR Rox kit (5Prime GmbH, Hamburg, Germany). Independent PCRs were performed using the same cDNA for both genes of interest (FLO11, GCN4, and HAC1) and CDC28 or H2A as reference. The following temperature profile was applied after an initial denaturation at 95°C for 2 min 20 s: 20-s denaturation at 95°C, 22-s hybridization at 64°C, and 22-s elongation at 70°C. The 40 cycles were followed by construction of a melting curve to determine PCR specificity, contamination, and the absence of primer dimers. Gene expression was quantified using the threshold cycle (ΔCT) method with efficiency. qRT-PCR experiments were performed from three independent cultures for each strain and condition.
RESULTS
The high-copy-number suppressor Hac1 supports Gcn4 activity during amino acid starvation.
In S. cerevisiae, amino acid starvation triggers the activation of the general amino acid control (GAAC) network and the cell surface flocculin gene FLO11 by the transcriptional activator Gcn4 (9, 18, 54). The Gcn4L267S variant carrying an amino acid substitution in the third of the four conserved leucines of the zipper dimerization domain results in a transcription factor which is less active than wild-type Gcn4. Indeed, Gcn4L267S has a reduced activation of metabolic genes in comparison to that of wild-type Gcn4, which is, however, sufficient to fulfill metabolic processes and to permit growth under amino acid starvation. In contrast, the developmental function of Gcn4L267S is diminished. Gcn4L253G carrying a helix breaker substitution at Leu253 results in a highly stable but transcriptionally inactive protein (17). We searched for a high-copy-number suppressor of Gcn4L267S that improves the transcriptional activity and growth during amino acid starvation. Patil et al. (45) described interplay between Gcn4 and the UPR bZIP transcription factor Hac1. We analyzed whether an overexpression of the native unspliced HAC1 influences growth of the gcn4 deletion strain expressing different GCN4 variants. Amino acid starvation was induced by addition of the histidine analogue 3-amino-1,2,4-triazole (3AT) to the growth medium, which acts as a false feedback inhibitor and inhibits the histidine biosynthetic enzyme His3 (55). Due to the fact that most natural yeasts are dimorphic and diploid and that the reduction in transcriptional activation capacity of Gcn4L267S is less severe in haploids, we used diploid yeast cells (17, 56). Growth of the diploid gcn4 deletion strain expressing GCN4L267S was enhanced during amino acid starvation when the bZIP transcription factor Hac1 was overexpressed (Fig. 1). Furthermore, growth of yeast cells expressing wild-type GCN4 in the presence of 3AT could be strengthened while overexpressing HAC1. In contrast, overexpression of HAC1 could not complement the growth defect of Δgcn4 cells expressing the stable but inactive GCN4L253G upon amino acid starvation.
Fig 1.
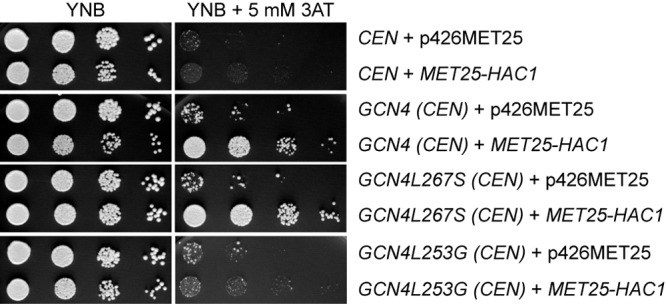
Hac1 enhances Gcn4 activity during amino acid starvation. The diploid gcn4 deletion strain (RH2694) expressing wild-type GCN4 (pME1092), the substituted GCN4L267S (pME2901) or GCN4L253G (pME3378) from CEN plasmids with GCN4 promoter and terminator, or, as a control, the empty vector (CEN) (pRS314) was additionally transformed with either independently overexpressed HAC1 (MET25-HAC1) (pME3498) or an empty vector (p426MET25). Strains were spotted in 10-fold dilutions either on YNB as a control or on YNB medium containing 5 mM 3AT to induce amino acid starvation. After incubation for 3 to 4 days at 30°C, plates were photographed.
This suppressor analysis supports a cross talk where the transcription factor Hac1 can support Gcn4 activity in amino acid biosynthesis. Hac1 controls the unfolded protein response (UPR), which is activated due to accumulation of misfolded proteins in the endoplasmic reticulum (23). The involvement of Hac1 in the Gcn4-mediated supply of amino acids (general amino acid control [GAAC]) has not been described yet, whereas it is known vice versa that Gcn4 is involved in the unfolded protein response (UPR) (45).
Hac1 is required for inducing GCREs in haploid and diploid yeast cells.
Subsequently, we examined whether Hac1 is required for activating the canonical Gcn4 promoter elements (GCREs). The influence of the hac1 deletion in comparison to the wild type was analyzed with a Gcn4-specific reporter which contains six GCRE binding sites upstream of the CYC1::lacZ minimal promoter. This construct was chromosomally integrated into the URA3 locus (57). Basal expression of the GCRE6::lacZ reporter was almost identical in wild-type and Δhac1 cells under nonstarvation conditions. However, Δhac1 cells were unable to induce the GCREs in response to amino acid starvation (3AT) and remained at the basal level as in sated wild-type cells without amino acid limitation (Fig. 2A). As expected, strains containing a gcn4 deletion were unable to activate GCRE6::lacZ expression in response to amino acid starvation. We compared the effect of Hac1 on the Gcn4 element GCRE with the effect of Gcn4 on the Hac1-dependent unfolded protein response element (UPRE). The reported UPRE::CYC1::lacZ, which is induced by tunicamycin-mediated ER stress, was tested (23). The strong induction of the UPRE::CYC1::lacZ reporter in the wild type is abolished when the transcription factor Hac1 is absent (Δhac1). A defective GCN4 gene (Δgcn4) results in partial induction. The double mutation Δgcn4 Δhac1 provides a phenotype similar to that of the Δhac1 mutation, further supporting the conclusion that UPRE transcription is primarily mediated by Hac1 (Fig. 2A).
Fig 2.

Hac1 is required to induce GCREs. (A) Expression of GCRE6::lacZ was determined in the haploid Σ1278b wild-type strain (WT) (RH3409) as well as in Δhac1 (RH3363), Δgcn4 (RH3410), and Δgcn4 Δhac1 (RH3411) mutant strains each carrying a chromosomally integrated GCRE6::lacZ reporter. Expression was measured under different nutritional conditions. Starting from one overnight culture, strains were diluted into fresh medium and further cultivated for 6 to 24 h in the respective media before specific β-galactosidase activities were assayed. Cultures were grown to log phase in YNB under nonstarvation conditions [light gray bars, YNB (6 h)]. Amino acid starvation and ER stress conditions were induced by addition of either 10 mM 3AT [dark gray bars, +3AT (8 h)] or 1 μg/ml tunicamycin [black bars, +Tm (6 h)]. For nitrogen starvation (N-starv), yeast cells were washed twice with 2% glucose before incubation for 24 h in minimal medium containing only 50 μM ammonium sulfate as the sole nitrogen source [white bars, N-starv (24 h)]. Units of specific β-galactosidase activities are shown in nanomoles per minutes per milligram. The bars represent the mean values of at least three independent measurements. As additional control, the haploid Σ1278b wild-type strain (WT) (RH2816), as well as Δhac1 (RH3351), Δgcn4 (RH2676), and Δgcn4 Δhac1 (RH3402) mutant strains, was transformed with the UPRE-CYC1-lacZ reporter gene carried on a multicopy vector (pMCZ-Y). Starting from one overnight culture, strains were diluted into fresh medium and further cultivated for 6 h in the respective media before specific β-galactosidase activities were assayed. Either cultures were grown to log phase in YNB medium [light gray bars, YNB (6 h)], or ER stress conditions were induced by the addition of 1 μg/ml tunicamycin for 2 h [black bars, YNB (4 h) + Tm (2 h)]. Units of specific β-galactosidase activities are shown in nanomoles per minutes per milligram. The bars represent the mean values based on quadruplicate determinations of at least three independent transformants. (B) The diploid wild-type yeast strain (WT) (RH3421), as well as homo- and heterozygous Δhac1/Δhac1 (RH3422), Δhac1/HAC1 (RH3423), Δgcn4/Δgcn4 (RH2398), Δgcn4/GCN4 (RH3424), Δgcn4/Δgcn4 Δhac1/Δhac1 (RH3350), and Δgcn4/GCN4 Δhac1/HAC1 (RH3425) mutant strains each carrying a chromosomally integrated GCRE6::lacZ reporter, was grown to log phase in YNB in the absence (light gray bars, YNB) or presence (dark gray bars, +3AT) of 10 mM 3AT before specific β-galactosidase activities were assayed. As a further control, the diploid yeast strains indicated on the abscissa were transformed with the UPRE-CYC1-lacZ reporter gene (pMCZ-Y) and transformants were grown in the presence or absence of tunicamycin before being used for β-galactosidase assays.
For analyzing the impact of Hac1 on Gcn4 target gene expression in diploids, haploid MATa strains containing the GCRE6::lacZ reporter gene were crossed with respective MATα strains to obtain homo- or heterozygous diploid strains deleted for HAC1, GCN4, or both. Heterozygous yeast strains revealed that a single intact copy of either GCN4 or HAC1 is sufficient to provide induction of the GCRE6::lacZ reporter upon amino acid starvation. A strain carrying only one intact GCN4 and one intact HAC1 copy was hardly able to induce the GCRE reporter. Induction of the GCRE reporter by 3AT was abolished when diploid homozygous strains deleted for HAC1 or GCN4 were analyzed (Fig. 2B). Therefore, neither haploid nor diploid Δhac1 cells showed intrinsic activation of GCRE upon amino acid starvation. In accordance with the data with haploids, the analysis of the corresponding activation of the UPRE-mediated unfolded protein response in diploids revealed that the homozygous Δgcn4 cells showed a reduced response to ER stress, whereas homozygous Δhac1 cells failed to activate UPRE-lacZ expression after tunicamycin treatment (Fig. 2B).
These data suggest that there is a cross talk in both directions between Hac1 and Gcn4. Besides the known requirement of Gcn4 for UPRE activation, there is also a Hac1 influence on the activation of Gcn4-specific target genes in response to amino acid starvation in haploids, as well as in diploids. One copy of either GCN4 or HAC1 is sufficient to fulfill the activation of Gcn4-specific target gene expression in diploids. The noncomplementation of the double heterozygous mutants could be a consequence of direct Gcn4 and Hac1 interactions.
ER stress inhibits GCN4 mRNA by a Hac1-independent mechanism.
Since the ER stress regulator Hac1 is required for the GCRE response during amino acid starvation, we analyzed the effect of tunicamycin-mediated ER stress on GCRE activity. ER stress represses GCRE activity independently of the presence or absence of Gcn4 or Hac1, suggesting an additional molecular mechanism (Fig. 2A). Nitrogen starvation was used as a control and was achieved by decreasing the ammonium sulfate concentration in the culture medium (from 50 mM to 50 μM). GCRE-driven gene expression was drastically reduced by ER stress, and the effect was similar to, and even more pronounced than, the described repressive effect of nitrogen starvation on GCREs (58). The kinetics of the observed repression of GCREs by ER stress suggests that it starts after about 60 min. A transient upregulation of Gcn4 protein levels 15 and 30 min after tunicamycin treatment had been described previously (45). However, it did not result in increased GCRE activation in the corresponding period (see Fig. S1 in the supplemental material).
We analyzed how the repressive effect of ER stress on GCREs correlates with the translational control of the GCN4 mRNA, which was monitored by a GCN4::lacZ reporter containing the wild-type leader with all four intact upstream open reading frames (uORFs) (59). Translational efficiency of GCN4 mRNA decreased in response to tunicamycin-mediated ER stress in haploid wild-type and Δhac1 and Δgcn4 single and double mutant strains (Fig. 3A). The increased activity in the Δgcn4 strain corresponds to permanent limitation caused by the lack of the Gcn4 regulator, resulting in an induced translation of the GCN4 reporter. This further suggests that ER stress activates an additional mechanism, which controls gene expression independently of Hac1. The repression of GCN4::lacZ by tunicamycin, which was measured as the control, is similar to the described effect of nitrogen repression (58) (Fig. 3A). There is no hint of a translational control of GCN4 by Hac1 in haploids since Hac1 influenced neither GCN4::lacZ expression of a reporter containing all four uORFs nor the expression of a reporter with mutated uORFs (Fig. 3A; see also Fig. S2 in the supplemental material).
Fig 3.

Tunicamycin-mediated ER stress represses GCN4 mRNA translation by a Hac1-independent mechanism. (A) Expression of a GCN4::lacZ fusion gene (p180) was measured in a haploid Σ1278b wild-type strain (WT) (RH2816) as well as in Δhac1 (RH3351), Δgcn4 (RH2676), and Δgcn4 Δhac1 (RH3402) mutant strains under different nutritional conditions. Bars depict means of at least three independent measurements of β-galactosidase activities. (B) Crude protein extracts were prepared from haploid Σ1278b wild-type cells (WT) (RH2816) as well as from Δgcn4 (RH2676) and Δhac1 (RH3351) mutant strains, respectively. Cells were grown either under normal conditions (untreated) or in the presence of ER stress conditions induced by 1 μg/ml tunicamycin (Tm). Additional amino acid starvation was obtained by adding 10 mM 3AT. Starting from one main culture with an OD600 of ∼0.8 at 30°C, cultures were quartered and cultivated for a further 90 min under the indicated conditions. Protein levels of Hac1 and eIF2α-P were analyzed by immunoblotting using specific antibodies. eIF2α was used as a loading control.
Amino acid starvation activates the general control system primarily on the level of translation. The eukaryotic initiation factor of translation eIF2α is phosphorylated (eIF2α-P) resulting in two effects: (i) the overall translation rate is reduced to save precursors of translation during amino acid limitation and (ii) GCN4 mRNA translation is increased to produce larger amounts of this transcription factor, which then activates numerous genes for amino acid biosynthesis (11). We monitored phosphorylation of eIF2α to analyze how ER stress (tunicamycin) in addition to simultaneous amino acid limitation (3AT) affects the levels of eIF2α-P (Fig. 3B). Phosphorylation of eIF2α upon amino acid starvation is unaffected by additional ER stress. Accordingly, ER stress without amino acid starvation does not result in phosphorylation of eIF2α. We also analyzed whether Hac1 is required for phosphorylation of eIF2α. Hac1 protein levels are significantly increased as a result of ER stress. Additional amino acid limitation still shows elevated Hac1 protein levels, but due to the generally reduced translation rate as a result of the increased levels of eIF2α-P, the amounts of Hac1 are decreased (Fig. 3B). In the absence of Hac1, eIF2α phosphorylation is still functional during simultaneous amino acid starvation and ER stress. In contrast, upon additional amino acid starvation Hac1 protein levels are reduced in Δgcn4 cells. In conclusion, these data suggest that ER stress inhibits translation of GCN4 mRNA in a molecular mechanism, which does not affect eIF2α phosphorylation caused by amino acid starvation. This novel ER stress-induced process on GCN4 and subsequently GCRE expression is independent of Hac1. In contrast, Gcn4 appears to influence Hac1 upon amino acid starvation, which further supports a cross talk between the two transcription factors.
Hac1 and Gcn4 mutually influence their mRNA expression levels.
We further analyzed the impact of Hac1 on the mRNA expression level of GCN4 and vice versa. Therefore, we performed quantitative real-time PCR (qRT-PCR) experiments under amino acid starvation as well as under ER stress conditions. The levels of GCN4 and HAC1 mRNA were upregulated by induction of amino acid starvation (3AT) in both haploid and diploid wild-type strains (Fig. 4). Deletion of HAC1 slightly reduced GCN4 mRNA expression levels upon 3AT treatment in haploids, whereas this effect was very prominent in diploid deletion strains. In contrast to amino acid starvation, 90-min tunicamycin-mediated ER stress did not result in enhanced GCN4 mRNA expression levels (Fig. 4A). The data are consistent with our results obtained from the GCN4::lacZ experiments (Fig. 3A). As expected, HAC1 mRNA expression levels increased under ER stress conditions in both haploid and diploid wild-type cells. Deletion of GCN4 impaired HAC1 mRNA expression levels in response to amino acid starvation as well as under ER stress conditions. The data are consistent with our results where Hac1 protein levels were reduced in Δgcn4 cells when they were treated with 3AT (Fig. 3B). In contrast to GCN4 mRNA levels, HAC1 mRNA expression levels were similar in haploid and diploid Δgcn4 cells (Fig. 4B).
Fig 4.

Hac1 and Gcn4 mutually influence their mRNA expression levels. (A) Relative GCN4 mRNA levels of either a haploid or diploid wild-type strain (RH2816 and RH2656) and a haploid or diploid HAC1 deletion strain (RH3351 and RH3412) were determined by qRT-PCR and normalized against H2A. Starting from one main culture with an OD600 of ∼0.6 at 30°C, cultures were divided and cultivated for a further 90 min under the indicated conditions. Amino acid starvation was induced by 10 mM 3AT (dark gray bars, +3AT), ER stress conditions were achieved by 1 μg/ml tunicamycin (black bars, +Tm). Experiments were performed from three independent cultures for each strain and condition. The mRNA levels were normalized to the respective untreated (YNB) wild type. Standard deviations are indicated as error bars. (B) HAC1 mRNA levels were analyzed similarly in either a haploid or a diploid wild-type strain (RH2816 and RH2656) and a haploid or a diploid GCN4 deletion strain (RH2676 and RH2658).
These findings further support our assumption that there is a cross talk between Hac1 and Gcn4. Gcn4 appears to be involved in HAC1 transcription in both haploid and diploid cells, whereas Hac1 appears to be absolutely required for GCN4 transcription in diploids.
Hac1 supports FLO11 expression and adhesive growth in haploid or diploid cells during amino acid starvation.
Besides the activation of target genes by binding to specific Gcn4 recognition elements in their promoter regions, Gcn4 evokes a strong adhesion of yeast cells on surfaces or on each other upon amino acid starvation. This adhesion is mediated by the flocculin Flo11. The FLO11 promoter lacks a typical GCRE and therefore is not a typical Gcn4-dependent promoter (18). Since Hac1 is required for activation of Gcn4 promoter elements (GCREs), we analyzed Hac1-dependent FLO11 expression in more detail in haploid and diploid cells. We investigated how Hac1 influences expression of a FLO11::lacZ reporter construct that contains 3,500 bp of the FLO11 promoter in front of a CYC1::lacZ minimal promoter integrated into the URA3 locus. Amino acid starvation (3AT) led to only a partial induction of β-galactosidase activity in Δhac1 cells in comparison to haploid wild-type cells. Haploid cells containing a gcn4 deletion showed no induction upon amino acid starvation. These findings were confirmed by quantitative real-time PCR (qRT-PCR) (Fig. 5A).
Fig 5.

Hac1 supports FLO11 expression during amino acid starvation. (A) Expression of FLO11::lacZ was assayed in the haploid Σ1278b wild-type yeast strain (WT) (RH3406) as well as in Δhac1 (RH3360), Δgcn4 (RH3407), and Δgcn4 Δhac1 (RH3408) mutant strains each carrying a chromosomally integrated FLO11::lacZ reporter. Cultures were grown to log phase in YNB in the absence (light gray bars, YNB) or presence of 10 mM 3AT (dark gray bars, +3AT) before specific β-galactosidase activities were measured. Relative FLO11 mRNA abundances determined by qRT-PCR were measured in haploid Σ1278b wild-type (WT) (RH2816), Δhac1 (RH3351), Δgcn4 (RH2676), and Δgcn4 Δhac1 (RH3402) yeast strains and normalized against CDC28. Experiments were performed from three independent cultures for each strain and condition, and cultivation was accomplished as described for FLO11::lacZ expression. The ΔCT method including efficiencies was used for quantification. Standard deviations are indicated as error bars. (B) The untransformed yeast strains described in panel A as well as a Δflo11 (RH2681) mutant strain were streaked out on solid YNB medium (nonstarved cells) and with 10 mM 3AT (histidine-starved cells), respectively. After incubation for 3 days at 30°C, adhesive growth was determined. Plates were photographed before (before wash) and after washing under a stream of water (after wash) to document remaining cells on the agar surface. The same yeast strains were grown in liquid YNB medium until reaching an optical density of 0.6 before 300 μl of each culture was transferred in a microtiter well. Cells were grown in the absence or presence of 5 or 10 mM 3AT to induce starvation-dependent adhesive growth. After incubation for 24 h at 30°C, sedimented cells were dyed with crystal violet and carefully washed. Finally, plates were photographed to document adhesive growth. (C) FLO11::lacZ expression was also determined in the diploid Σ1278b wild-type yeast strain (WT) (RH3417) as well as in diploid homo- and heterozygous Δhac1/Δhac1 (RH3362), Δhac1/HAC1 (RH3418), Δgcn4/Δgcn4 (RH2695), Δgcn4/GCN4 (RH3419), Δgcn4/Δgcn4 Δhac1/Δhac1 (RH3349), and Δgcn4/GCN4 Δhac1/HAC1 (RH3420) mutant yeast strains each carrying a chromosomally integrated FLO11::lacZ reporter. For testing amino acid starvation-induced adhesive growth, the diploid wild-type yeast strain (WT) (RH2656) as well as Δflo11/Δflo11 (RH2661), Δhac1/Δhac1 (RH3412), Δhac1/HAC1 (RH3413), Δgcn4/Δgcn4 (RH2658), Δgcn4/GCN4 (RH3414), Δgcn4/Δgcn4 Δhac1/Δhac1 (RH3415), and Δgcn4/GCN4 Δhac1/HAC1 (RH3416) mutant strains was used, and the assay was performed as described for panel B.
Subsequently, the consequences of the reduced FLO11 expression of Δhac1 cells on the adherence phenotype were analyzed. We investigated to what extent Δhac1 cells can still grow adhesively when starved for amino acids on either agar or plastic surfaces. Cells deleted for GCN4 were not able to grow adhesively and were comparable to the Δflo11 strain deficient in the structural gene for the adhesin. Haploid wild-type and Δhac1 cells became adhesive on agar when starved for amino acids (3AT), but Δhac1 cells showed only a constricted adhesive growth on plastic, which reflected the halved FLO11 expression (Fig. 5B).
We also explored Hac1-dependent FLO11 expression in diploids and found a much more pronounced phenotype. Haploid MATa strains containing the FLO11::lacZ reporter gene were crossed with respective MATα strains to obtain homo- and heterozygous diploid strains deleted for HAC1, GCN4, or both. Starvation-dependent FLO11::lacZ expression was strongly decreased in diploid homozygous Δhac1 and Δgcn4 strains, and the cells were not able to grow adhesively (Fig. 5C). A single copy of HAC1 was sufficient in heterozygous Δhac1/HAC1 cells to restore FLO11::lacZ expression and adhesive growth, whereas heterozygous Δgcn4/GCN4 resulted only in a partial FLO11 induction and reduced adherence.
These data support an auxiliary role of Hac1 for FLO11 expression during amino acid starvation in haploid and even more in diploid yeasts.
Hac1 and Gcn4 act on similar promoter elements in the FLO11 promoter.
Both Hac1 and Gcn4 affect FLO11 expression during amino acid starvation. Gcn4 action on the presumably longest promoter of S. cerevisiae, spanning approximately 3.5 kb, might be indirect, because no promoter binding had been detected. Gcn4 action is also complex because it includes several promoter regions, including also basal promoter elements (18). We compared the effects of the presence or absence of Hac1 and Gcn4 on a set of 14 described reporter constructs containing individual 400-bp FLO11 promoter fragments that overlap by 200 bp in front of a CYC1::lacZ fusion gene (53). Homozygous diploid yeast cells were analyzed, because they show the most pronounced effect of Hac1 on FLO11 expression (Fig. 5). The comparison between diploid wild-type, Δgcn4, and Δhac1 cells shows two promoter regions, which are significantly affected by both Gcn4 and Hac1. These regions are located approximately 2 or 1 kb upstream of the AUG of the FLO11 open reading frame, respectively (Fig. 6; see also Table S1 in the supplemental material). Already in the absence of amino acid limitation, the 1-kb upstream region requires not only Gcn4 but also Hac1 for basal expression. Activation by amino acid limitation (3AT) is reduced when Hac1 is missing and is abolished in the absence of Gcn4. This regulatory region corresponds to the major regulatory region of the FLO11 promoter (18, 21, 60). The second upstream region at −2 kb is also affected by Hac1 and Gcn4. There are differences between the two transcription factors, because only Hac1 seems to participate in repression of this promoter region in the absence of amino acid limitation. The effect of Hac1 and Gcn4 on the same FLO11 promoter elements further corroborates the interplay between the two transcription factors.
Fig 6.

Effect of Hac1 and Gcn4 on FLO11::lacZ promoter elements. The β-galactosidase activity of 14 isolated 400-bp elements of the FLO11 promoter region was assayed in diploid wild-type cells (RH2656) or in corresponding cells deleted for the transcription factor HAC1 (RH3412) or GCN4 (RH2658) (see Table S1 in the supplemental material). The activity was determined under nonstarvation conditions (YNB, upper graphs) or in the presence of amino acid starvation (3AT, lower graphs) induced by the addition of 10 mM 3AT. Each breakpoint represents the measured reporter construct expression less the measured expression of the construct without insert (pME2212) in the indicated genetic background. β-Galactosidase activities measured in the wild type are shown by diamonds, Δhac1/Δhac1 breakpoints are shown by squares, and Δgcn4/Δgcn4 breakpoints are shown by triangles.
DISCUSSION
XBP1 and ATF4, the mammalian homologues of Hac1 and Gcn4, represent essential genes, which are involved in a multiplicity of metabolic and developmental processes ensuring the survival of the organism. The general amino acid control (GAAC) and the unfolded protein response (UPR) of baker's yeast are, in contrast to those of mammals, not essential. Yeast has been used here to study the interplay between the two networks. Patil et al. (45) discovered a linkage between the two pathways where Gcn4 is required for induction of a majority of UPR target genes during ER stress. We demonstrate here that this is a mutual interplay, which also applies vice versa. The initial finding was that the survival of diploid cells expressing either wild-type GCN4 or the partially active GCN4L267S (17) could be increased upon amino acid starvation when native HAC1 was overexpressed. Hac1 can only improve Gcn4 function but is unable to complement a gcn4 deletion strain or a strain with an inactive Gcn4 variant.
Hac1 and the general amino acid control.
Gcn4 target gene expression and GCN4 mRNA expression itself are influenced by Hac1 in both haploid and diploid cells, but effects are stronger and more prominent in diploids. In haploids, Hac1 is not involved in basal expression of Gcn4 target genes but is required for the Gcn4-mediated response to amino acid starvation. Diploid cells show the same lack of induction during starvation, but in addition, Hac1 is required for the basal expression levels. This could be a direct effect where Hac1 itself or in combination with Gcn4 activates Gcn4 targets, or an indirect effect like Hac1-dependent stabilization of Gcn4 as earlier proposed (45). Several findings indicate that the effects of Hac1 on Gcn4 are indirect: (i) Hac1 itself does not influence eIF2α phosphorylation, (ii) Hac1 does not affect GCN4 mRNA translation in haploids, (iii) ER stress inhibits GCN4 mRNA translation in a Hac1-independent mechanism, (iv) ER stress represses Gcn4 target gene expression, and (v) Hac1 is hardly detectable in Δgcn4 cells upon ER stress conditions and additional amino acid starvation. A regulation of the HAC1 gene by Gcn4 is supported by the fact that in the absence of GCN4 and ER stress constitutive expression of HAC1 does not activate transcription of Hac1 target genes (45). Furthermore, Hac1 contains two Gcn4-specific consensus sequences in its promoter, which is not the case vice versa. Furthermore, Gcn4 has a weak but distinct RNase activity, and therefore, it might be possible that Gcn4 regulates HAC1 mRNA stability (61).
The situation is more complicated because Hac1 is responsible for full activation of Gcn4 target gene expression upon amino acid starvation. Vice versa, it is described that Gcn4 directly interacts with two of the three Hac1-specific promoter elements, which share half-site similarity: three bases (GTG) are identical. Gcn4 is able to bind to half-sites (15), and the shared half-site is present in the consensus sequence which is found in the promoters of Gcn4 target genes (RRRWGASTCA, with R = purine, W = T or A, and S = G or C) (9). An attractive explanation is that both Gcn4 and Hac1 regulate the activation of the other target genes by binding to half-sites in their promoter regions. Hac1 and Gcn4 could also act together at target promoters, but we could not copurify a Gcn4/Hac1 heterodimer in our experiments. Nevertheless, a collaboration between the two factors which might depend on a stable physical interaction is supported by the findings that (i) overexpression of HAC1 increased survival of diploid cells expressing either wild type or the partially active GCN4L267S during amino acid starvation and (ii) Hac1- and Gcn4-specific target gene expression is strongly decreased when the responsible main transcription factor is deleted.
Hac1 function in dimorphism.
CaHac1 and HacA, the Hac1 homologues of Candida albicans and Aspergillus fumigatus, respectively, play important roles in regulating morphology, which in turn is important for virulence of these pathogenic fungi (62, 63). Furthermore, both CaGcn4 and CpcA, the Gcn4 homologue proteins, are also involved in pathogenicity (64–66). The mammalian Hac1-like XBP1 and the unfolded protein response (UPR) play important roles in tumorigenesis, and UPR suppressors are proposed as therapeutic agents (67–69). In S. cerevisiae, Gcn4 is required for adhesion and pseudohyphal development upon nutrient starvation (18). In this study, we identified Hac1 as an auxiliary regulator of FLO11 expression and therefore dimorphism of S. cerevisiae. Gcn4 and Hac1 influence an identical FLO11 promoter element and presumably act indirectly in combination with other transcriptional regulators because there is neither a Gcn4 nor a Hac1 predicted recognition element in the FLO11 promoter. It is yet unclear why both Hac1 and Gcn4 are specifically required in diploids to induce FLO11 expression. The overlapping elements, which are influenced by the transcription factors Tec1, Ste12, and Flo8, support this hypothesis (53, 70). STE12 and TEC1 represent potential target genes of Gcn4, since both carry at least one Gcn4 recognition element in their promoters. Furthermore, the promoter of TEC1 actually contains five independent UPREs, whereas three of them are arranged on the complementary strand. These findings enforce our hypothesis that both Gcn4 and Hac1 are involved in Flo11 regulation per se, however, presumably by binding to another transcription factor, e.g., the TEA protein Tec1 as one of the major regulators of the FLO11 promoter. However, Tec1 protein levels did not depend on the absence or presence of Gcn4 or Hac1 (data not shown). Our data suggest that the impact of Gcn4 and Hac1 on FLO11 is complex. It includes the same regions of the FLO11 promoter and is at least partially mediated by changes of the amount of another transcription factor, which in turn is able to bind to the FLO11 promoter.
In summary, Hac1 not only affects the UPR to reduce misfolded or unfolded proteins but also is involved in metabolic and developmental processes generally regulated by Gcn4 in response to amino acid starvation. At least 13 bZIP transcription factors exist in S. cerevisiae, whereas the increasing complexity during evolution is reflected by a minimum of 51 bZIP factors in humans (71). Understanding the complex regulation of dimorphism, stability control, nuclear trafficking, and cell death pathways in fungal models might be relevant for tumor therapy in humans. The mutual cross talk between the UPR and the GAAC in yeast is much broader than supposed, and it will be interesting to analyze if such a cross talk also exists in higher eukaryotes.
Supplementary Material
ACKNOWLEDGMENTS
We thank Kai Heimel and Susanna Braus-Stromeyer for critically reading the manuscript and helpful comments. We are grateful to Agnieszka Kolka and Maria Meyer for their technical assistance during the experiments. Special thanks to Olaf Grundmann, Thomas Dever, Alan Hinnebusch, and Steffen Rupp for strains, plasmids, and antibodies.
This work was supported by grants from the Deutsche Forschungsgemeinschaft, the Cluster of Excellence and DFG Research Center Nanoscale Microscopy and Molecular Physiology of the Brain, the Volkswagenstiftung, and the Fonds der Chemischen Industrie.
Footnotes
Published ahead of print 21 June 2013
Supplemental material for this article may be found at http://dx.doi.org/10.1128/EC.00123-13.
REFERENCES
- 1. Yang R, Wek SA, Wek RC. 2000. Glucose limitation induces GCN4 translation by activation of Gcn2 protein kinase. Mol. Cell. Biol. 20:2706–2717 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Mösch HU. 2000. Pseudohyphal development of Saccharomyces cerevisiae. Contrib. Microbiol. 5:185–200 [DOI] [PubMed] [Google Scholar]
- 3. Meussdoerffer F, Fink GR. 1983. Structure and expression of two aminoacyl-tRNA synthetase genes from Saccharomyces cerevisiae. J. Biol. Chem. 258:6293–6299 [PubMed] [Google Scholar]
- 4. Ron D. 2002. Translational control in the endoplasmic reticulum stress response. J. Clin. Invest. 110:1383–1388 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Kozutsumi Y, Segal M, Normington K, Gething MJ, Sambrook J. 1988. The presence of malfolded proteins in the endoplasmic reticulum signals the induction of glucose-regulated proteins. Nature 332:462–464 [DOI] [PubMed] [Google Scholar]
- 6. Schröder M, Kaufman RJ. 2005. ER stress and the unfolded protein response. Mutat. Res. 569:29–63 [DOI] [PubMed] [Google Scholar]
- 7. Patil C, Walter P. 2001. Intracellular signaling from the endoplasmic reticulum to the nucleus: the unfolded protein response in yeast and mammals. Curr. Opin. Cell Biol. 13:349–355 [DOI] [PubMed] [Google Scholar]
- 8. Drysdale CM, Duenas E, Jackson BM, Reusser U, Braus GH, Hinnebusch AG. 1995. The transcriptional activator GCN4 contains multiple activation domains that are critically dependent on hydrophobic amino acids. Mol. Cell. Biol. 15:1220–1233 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Natarajan K, Meyer MR, Jackson BM, Slade D, Roberts C, Hinnebusch AG, Marton MJ. 2001. Transcriptional profiling shows that Gcn4p is a master regulator of gene expression during amino acid starvation in yeast. Mol. Cell. Biol. 21:4347–4368 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Hinnebusch AG. 1984. Evidence for translational regulation of the activator of general amino acid control in yeast. Proc. Natl. Acad. Sci. U. S. A. 81:6442–6446 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Hinnebusch AG. 2005. Translational regulation of GCN4 and the general amino acid control of yeast. Annu. Rev. Microbiol. 59:407–450 [DOI] [PubMed] [Google Scholar]
- 12. Mösch HU, Scheier B, Lahti R, Mantsala P, Braus GH. 1991. Transcriptional activation of yeast nucleotide biosynthetic gene ADE4 by GCN4. J. Biol. Chem. 266:20453–20456 [PubMed] [Google Scholar]
- 13. Hope IA, Struhl K. 1986. Functional dissection of a eukaryotic transcriptional activator protein, GCN4 of yeast. Cell 46:885–894 [DOI] [PubMed] [Google Scholar]
- 14. Oliphant AR, Brandl CJ, Struhl K. 1989. Defining the sequence specificity of DNA-binding proteins by selecting binding sites from random-sequence oligonucleotides: analysis of yeast GCN4 protein. Mol. Cell. Biol. 9:2944–2949 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Hollenbeck JJ, Oakley MG. 2000. GCN4 binds with high affinity to DNA sequences containing a single consensus half-site. Biochemistry 39:6380–6389 [DOI] [PubMed] [Google Scholar]
- 16. Chan IS, Fedorova AV, Shin JA. 2007. The GCN4 bZIP targets noncognate gene regulatory sequences: quantitative investigation of binding at full and half sites. Biochemistry 46:1663–1671 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Herzog B, Streckfuss-Bömeke K, Braus GH. 2011. A feedback circuit between transcriptional activation and self-destruction of Gcn4 separates its metabolic and morphogenic response in diploid yeasts. J. Mol. Biol. 405:909–925 [DOI] [PubMed] [Google Scholar]
- 18. Braus GH, Grundmann O, Brückner S, Mösch HU. 2003. Amino acid starvation and Gcn4p regulate adhesive growth and FLO11 gene expression in Saccharomyces cerevisiae. Mol. Biol. Cell 14:4272–4284 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Reynolds TB, Fink GR. 2001. Bakers' yeast, a model for fungal biofilm formation. Science 291:878–881 [DOI] [PubMed] [Google Scholar]
- 20. Gimeno CJ, Ljungdahl PO, Styles CA, Fink GR. 1992. Unipolar cell divisions in the yeast S. cerevisiae lead to filamentous growth: regulation by starvation and RAS. Cell 68:1077–1090 [DOI] [PubMed] [Google Scholar]
- 21. Brückner S, Mösch HU. 2012. Choosing the right lifestyle: adhesion and development in Saccharomyces cerevisiae. FEMS Microbiol. Rev. 36:25–58 [DOI] [PubMed] [Google Scholar]
- 22. Lambrechts MG, Bauer FF, Marmur J, Pretorius IS. 1996. Muc1, a mucin-like protein that is regulated by Mss10, is critical for pseudohyphal differentiation in yeast. Proc. Natl. Acad. Sci. U. S. A. 93:8419–8424 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Mori K, Kawahara T, Yoshida H, Yanagi H, Yura T. 1996. Signalling from endoplasmic reticulum to nucleus: transcription factor with a basic-leucine zipper motif is required for the unfolded protein-response pathway. Genes Cells 1:803–817 [DOI] [PubMed] [Google Scholar]
- 24. Weiss MA, Ellenberger T, Wobbe CR, Lee JP, Harrison SC, Struhl K. 1990. Folding transition in the DNA-binding domain of GCN4 on specific binding to DNA. Nature 347:575–578 [DOI] [PubMed] [Google Scholar]
- 25. Travers KJ, Patil CK, Wodicka L, Lockhart DJ, Weissman JS, Walter P. 2000. Functional and genomic analyses reveal an essential coordination between the unfolded protein response and ER-associated degradation. Cell 101:249–258 [DOI] [PubMed] [Google Scholar]
- 26. Cox JS, Shamu CE, Walter P. 1993. Transcriptional induction of genes encoding endoplasmic reticulum resident proteins requires a transmembrane protein kinase. Cell 73:1197–1206 [DOI] [PubMed] [Google Scholar]
- 27. Mori K, Ma W, Gething MJ, Sambrook J. 1993. A transmembrane protein with a cdc2+/CDC28-related kinase activity is required for signaling from the ER to the nucleus. Cell 74:743–756 [DOI] [PubMed] [Google Scholar]
- 28. van Anken E, Braakman I. 2005. Endoplasmic reticulum stress and the making of a professional secretory cell. Crit. Rev. Biochem. Mol. Biol. 40:269–283 [DOI] [PubMed] [Google Scholar]
- 29. Kawaguchi S, Ng DT. 2011. Sensing ER stress. Science 333:1830–1831 [DOI] [PubMed] [Google Scholar]
- 30. Walter P, Ron D. 2011. The unfolded protein response: from stress pathway to homeostatic regulation. Science 334:1081–1086 [DOI] [PubMed] [Google Scholar]
- 31. Liu CY, Wong HN, Schauerte JA, Kaufman RJ. 2002. The protein kinase/endoribonuclease IRE1α that signals the unfolded protein response has a luminal N-terminal ligand-independent dimerization domain. J. Biol. Chem. 277:18346–18356 [DOI] [PubMed] [Google Scholar]
- 32. Nikawa J, Yamashita S. 1992. IRE1 encodes a putative protein kinase containing a membrane-spanning domain and is required for inositol phototrophy in Saccharomyces cerevisiae. Mol. Microbiol. 6:1441–1446 [DOI] [PubMed] [Google Scholar]
- 33. Sidrauski C, Walter P. 1997. The transmembrane kinase Ire1p is a site-specific endonuclease that initiates mRNA splicing in the unfolded protein response. Cell 90:1031–1039 [DOI] [PubMed] [Google Scholar]
- 34. Shamu CE, Walter P. 1996. Oligomerization and phosphorylation of the Ire1p kinase during intracellular signaling from the endoplasmic reticulum to the nucleus. EMBO J. 15:3028–3039 [PMC free article] [PubMed] [Google Scholar]
- 35. Tirasophon W, Welihinda AA, Kaufman RJ. 1998. A stress response pathway from the endoplasmic reticulum to the nucleus requires a novel bifunctional protein kinase/endoribonuclease (Ire1p) in mammalian cells. Genes Dev. 12:1812–1824 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Kohno K. 2007. How transmembrane proteins sense endoplasmic reticulum stress. Antioxid. Redox Signal. 9:2295–2303 [DOI] [PubMed] [Google Scholar]
- 37. Ruegsegger U, Leber JH, Walter P. 2001. Block of HAC1 mRNA translation by long-range base pairing is released by cytoplasmic splicing upon induction of the unfolded protein response. Cell 107:103–114 [DOI] [PubMed] [Google Scholar]
- 38. Kawahara T, Yanagi H, Yura T, Mori K. 1997. Endoplasmic reticulum stress-induced mRNA splicing permits synthesis of transcription factor Hac1p/Ern4p that activates the unfolded protein response. Mol. Biol. Cell 8:1845–1862 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Chapman RE, Walter P. 1997. Translational attenuation mediated by an mRNA intron. Curr. Biol. 7:850–859 [DOI] [PubMed] [Google Scholar]
- 40. Cox JS, Walter P. 1996. A novel mechanism for regulating activity of a transcription factor that controls the unfolded protein response. Cell 87:391–404 [DOI] [PubMed] [Google Scholar]
- 41. Irniger S, Braus GH. 2003. Controlling transcription by destruction: the regulation of yeast Gcn4p stability. Curr. Genet. 44:8–18 [DOI] [PubMed] [Google Scholar]
- 42. Kornitzer D, Raboy B, Kulka RG, Fink GR. 1994. Regulated degradation of the transcription factor Gcn4. EMBO J. 13:6021–6030 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Shemer R, Meimoun A, Holtzman T, Kornitzer D. 2002. Regulation of the transcription factor Gcn4 by Pho85 cyclin PCL5. Mol. Cell. Biol. 22:5395–5404 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Pal B, Chan NC, Helfenbaum L, Tan K, Tansey WP, Gething MJ. 2007. SCFCdc4-mediated degradation of the Hac1p transcription factor regulates the unfolded protein response in Saccharomyces cerevisiae. Mol. Biol. Cell 18:426–440 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Patil CK, Li H, Walter P. 2004. Gcn4p and novel upstream activating sequences regulate targets of the unfolded protein response. PLoS Biol. 2:E246. 10.1371/journal.pbio.0020246 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Fordyce PM, Pincus D, Kimmig P, Nelson CS, El-Samad H, Walter P, DeRisi JL. 2012. Basic leucine zipper transcription factor Hac1 binds DNA in two distinct modes as revealed by microfluidic analyses. Proc. Natl. Acad. Sci. U. S. A. 109:E3084–3093 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Ito H, Fukuda Y, Murata K, Kimura A. 1983. Transformation of intact yeast cells treated with alkali cations. J. Bacteriol. 153:163–168 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Brachmann CB, Davies A, Cost GJ, Caputo E, Li J, Hieter P, Boeke JD. 1998. Designer deletion strains derived from Saccharomyces cerevisiae S288C: a useful set of strains and plasmids for PCR-mediated gene disruption and other applications. Yeast 14:115–132 [DOI] [PubMed] [Google Scholar]
- 49. Southern EM. 1975. Detection of specific sequences among DNA fragments separated by gel electrophoresis. J. Mol. Biol. 98:503–517 [DOI] [PubMed] [Google Scholar]
- 50. Bradford MM. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72:248–254 [DOI] [PubMed] [Google Scholar]
- 51. Roberts RL, Fink GR. 1994. Elements of a single MAP kinase cascade in Saccharomyces cerevisiae mediate two developmental programs in the same cell type: mating and invasive growth. Genes Dev. 8:2974–2985 [DOI] [PubMed] [Google Scholar]
- 52. Rose M, Botstein D. 1983. Construction and use of gene fusions to lacZ (β-galactosidase) that are expressed in yeast. Methods Enzymol. 101:167–180 [DOI] [PubMed] [Google Scholar]
- 53. Rupp S, Summers E, Lo HJ, Madhani H, Fink G. 1999. MAP kinase and cAMP filamentation signaling pathways converge on the unusually large promoter of the yeast FLO11 gene. EMBO J. 18:1257–1269 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Kleinschmidt M, Grundmann O, Bluthgen N, Mösch HU, Braus GH. 2005. Transcriptional profiling of Saccharomyces cerevisiae cells under adhesion-inducing conditions. Mol. Genet. Genomics 273:382–393 [DOI] [PubMed] [Google Scholar]
- 55. Klopotowski T, Wiater A. 1965. Synergism of aminotriazole and phosphate on the inhibition of yeast imidazole glycerol phosphate dehydratase. Arch. Biochem. Biophys. 112:562–566 [DOI] [PubMed] [Google Scholar]
- 56. Mortimer RK. 2000. Evolution and variation of the yeast (Saccharomyces) genome. Genome Res. 10:403–409 [DOI] [PubMed] [Google Scholar]
- 57. Albrecht G, Mösch HU, Hoffmann B, Reusser U, Braus GH. 1998. Monitoring the Gcn4 protein-mediated response in the yeast Saccharomyces cerevisiae. J. Biol. Chem. 273:12696–12702 [DOI] [PubMed] [Google Scholar]
- 58. Grundmann O, Mösch HU, Braus GH. 2001. Repression of GCN4 mRNA translation by nitrogen starvation in Saccharomyces cerevisiae. J. Biol. Chem. 276:25661–25671 [DOI] [PubMed] [Google Scholar]
- 59. Hinnebusch AG. 1985. A hierarchy of trans-acting factors modulates translation of an activator of amino acid biosynthetic genes in Saccharomyces cerevisiae. Mol. Cell. Biol. 5:2349–2360 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Octavio LM, Gedeon K, Maheshri N. 2009. Epigenetic and conventional regulation is distributed among activators of FLO11 allowing tuning of population-level heterogeneity in its expression. PLoS Genet. 5:e1000673. 10.1371/journal.pgen.1000673 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Nikolaev Y, Deillon C, Hoffmann SR, Bigler L, Friess S, Zenobi R, Pervushin K, Hunziker P, Gutte B. 2010. The leucine zipper domains of the transcription factors GCN4 and c-Jun have ribonuclease activity. PLoS One 5:e10765. 10.1371/journal.pone.0010765 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Wimalasena TT, Enjalbert B, Guillemette T, Plumridge A, Budge S, Yin Z, Brown AJ, Archer DB. 2008. Impact of the unfolded protein response upon genome-wide expression patterns, and the role of Hac1 in the polarized growth, of Candida albicans. Fungal Genet. Biol. 45:1235–1247 [DOI] [PubMed] [Google Scholar]
- 63. Richie DL, Hartl L, Aimanianda V, Winters MS, Fuller KK, Miley MD, White S, McCarthy JW, Latge JP, Feldmesser M, Rhodes JC, Askew DS. 2009. A role for the unfolded protein response (UPR) in virulence and antifungal susceptibility in Aspergillus fumigatus. PLoS Pathog. 5:e1000258. 10.1371/journal.ppat.1000258 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Krappmann S, Bignell EM, Reichard U, Rogers T, Haynes K, Braus GH. 2004. The Aspergillus fumigatus transcriptional activator CpcA contributes significantly to the virulence of this fungal pathogen. Mol. Microbiol. 52:785–799 [DOI] [PubMed] [Google Scholar]
- 65. Tripathi G, Wiltshire C, Macaskill S, Tournu H, Budge S, Brown AJ. 2002. Gcn4 co-ordinates morphogenetic and metabolic responses to amino acid starvation in Candida albicans. EMBO J. 21:5448–5456 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Gildor T, Shemer R, Atir-Lande A, Kornitzer D. 2005. Coevolution of cyclin Pcl5 and its substrate Gcn4. Eukaryot. Cell 4:310–318 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Saito S, Furuno A, Sakurai J, Sakamoto A, Park HR, Shin-Ya K, Tsuruo T, Tomida A. 2009. Chemical genomics identifies the unfolded protein response as a target for selective cancer cell killing during glucose deprivation. Cancer Res. 69:4225–4234 [DOI] [PubMed] [Google Scholar]
- 68. Wang G, Yang ZQ, Zhang K. 2010. Endoplasmic reticulum stress response in cancer: molecular mechanism and therapeutic potential. Am. J. Transl. Res. 2:65–74 [PMC free article] [PubMed] [Google Scholar]
- 69. Shajahan AN, Riggins RB, Clarke R. 2009. The role of X-box binding protein-1 in tumorigenicity. Drug News Perspect. 22:241–246 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Pan X, Heitman J. 2002. Protein kinase A operates a molecular switch that governs yeast pseudohyphal differentiation. Mol. Cell. Biol. 22:3981–3993 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Amoutzias GD, Robertson DL, Van de Peer Y, Oliver SG. 2008. Choose your partners: dimerization in eukaryotic transcription factors. Trends Biochem. Sci. 33:220–229 [DOI] [PubMed] [Google Scholar]
- 72. Strittmatter AW, Fischer C, Kleinschmidt M, Braus GH. 2006. FLO11 mediated filamentous growth of the yeast Saccharomyces cerevisiae depends on the expression of the ribosomal RPS26 genes. Mol. Genet. Genomics 276:113–125 [DOI] [PubMed] [Google Scholar]
- 73. Mueller PP, Hinnebusch AG. 1986. Multiple upstream AUG codons mediate translational control of GCN4. Cell 45:201–207 [DOI] [PubMed] [Google Scholar]
- 74. Goldstein AL, McCusker JH. 1999. Three new dominant drug resistance cassettes for gene disruption in Saccharomyces cerevisiae. Yeast 15:1541–1553 [DOI] [PubMed] [Google Scholar]
- 75. Mumberg D, Müller R, Funk M. 1994. Regulatable promoters of Saccharomyces cerevisiae: comparison of transcriptional activity and their use for heterologous expression. Nucleic Acids Res. 22:5767–5768 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Guarente L, Ptashne M. 1981. Fusion of Escherichia coli lacZ to the cytochrome c gene of Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. U. S. A. 78:2199–2203 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. Sikorski RS, Hieter P. 1989. A system of shuttle vectors and yeast host strains designed for efficient manipulation of DNA in Saccharomyces cerevisiae. Genetics 122:19–27 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.