

Abstract
Background
CYP2C19 encodes a member of the cytochrome P450 superfamily of enzymes, which play a central role in activating and detoxifying many carcinogens and endogenous compounds thought to be involved in the development of cancer. In the past decade, two common polymorphisms among CYP2C19 (CYP2C19*2 and CYP2C19*3) that are responsible for the poor metabolizers (PMs) phenotype in humans and cancer susceptibility have been investigated extensively; however, these studies have yielded contradictory results.
Methods and Results
To investigate this inconsistency, we conducted a comprehensive meta-analysis of 11,554 cases and 16,592 controls from 30 case-control studies. Overall, the odds ratio (OR) of cancer was 1.52 [95% confidence interval (CI): 1.23–1.88, P<10-4] for CYP2C19 PMs genotypes. However, this significant association vanished when the analyses were restricted to 5 larger studies (no. of cases ≥ 500 cases). In the subgroup analysis for different cancer types, PMs genotypes had an effect of increasing the risks of esophagus cancer, gastric cancer, lung cancer and hepatocellular carcinoma as well as head neck cancer. Significant results were found in Asian populations when stratified by ethnicity; whereas no significant associations were found among Caucasians. Stratified analyses according to source of controls, significant associations were found only in hospital base controls.
Conclusions
Our meta-analysis suggests that the CYP2C19 PMs genotypes most likely contributes to cancer susceptibility, particularly in the Asian populations.
Introduction
Cancer is considered to be a multifactorial disease, in which multiple exposures to endogenous factors and dietary carcinogens interact with individual genetic background in a complex manner resulting in modulation of the risk. It has been reported that up to 80% of human cancers arise as a consequence of environmental exposure and host susceptibility factors [1]. Most pro-carcinogens exert their genotoxicity after undergoing metabolic activation by various enzymes [2]. Therefore, polymorphisms of the genes encoding for those enzymes involved in the activating and detoxifying carcinogens and endogenous compounds may be related to inter-individual differences in cancer susceptibility.
Individuals vary widely in their susceptibility to carcinogens. One attractive genetic mechanism to account for this variability is the activity of polymorphically expressed cytochrome P450 enzymes that activate procarcinogens or conversely detoxify carcinogens. Cytochrome P450 2C19 (CYP2C19) is an enzyme involved in the metabolism of an extensive range of clinical agents including diazepam, mephenytoin, proton-pump inhibitors and clopidogrel [3]. CYP2C19 also plays a crucial role in either the detoxification or inactivation of potential carcinogens, or the bioactivation of some environmental procarcinogens to reactive DNA-binding metabolites, such as nitrosamine [4]. Poor metabolizer (PM) and extensive metabolizer (EM) phenotypes has been demonstrated based on the ability to metabolize (S)-mephenytoin and other CYP2C19 substrates which have been assigned to genetic polymorphisms [5,6]. Several important single nucleotide polymorphisms have been identified in the CYP2C19 gene; however, most cases can be explained by only two kinds, CYP2C19*2 (rs4244285) and CYP2C19*3 (rs4986893) which are responsible for the PMs phenotype in humans, while EM is assigned to the CYP2C19*1 allele [5,6]. CYP2C19*2 shows a single-base mutation (G→A) in exon 5 of CYP2C19 which produces an aberrant splice site and is known to be present in both Japanese and Caucasian populations. CYP2C19*3 consists of a premature stop codon (G→A) in exon 4 and is reported in Oriental populations including both Japanese and Chinese but rare in the Caucasians [7].
Despite the biological plausibility of CYP2C19 region polymorphisms as a modulator of cancer susceptibility, previously inconsistent results have appeared in the literature. Such inconsistency could be due to the small effect of the polymorphism on cancer, sample size and ethnic diversity, and individual studies may have insufficient power to reach a comprehensive and reliable conclusion. We therefore performed a meta-analysis of the published studies to clarify this inconsistency and to establish a comprehensive picture of the relationship between CYP2C19 and cancer susceptibility.
Materials and Methods
Identification of eligible studies
A comprehensive literature search was performed using the PubMed, Web of Science, EMBASE and CNKI (Chinese National Knowledge Infrastructure) databases for relevant articles with a combination of the following keywords: ‘Cytochrome P450 2C19’, ‘CYP2C19’, ‘polymorphism’, ‘variation’, and ‘cancer’, or ‘tumor’, or ‘carcinoma’. Genetic association studies published before the end of December 2012 on cancer and polymorphisms in the CYP2C19 gene described above were retrieved, and their references were checked to identify other relevant publications. Review articles were also inspected to find additional eligible studies. All relevant reports identified were included without language restriction. As studies with the same population by different investigators or overlapping data by the same authors were found, the most recent or complete articles with the largest numbers of subjects were included.
Inclusion criteria and data extraction
The following criteria were used for the study selection: (1) evaluation of at least one of these two polymorphisms (CYP2C19*2 and CYP2C19*3) and cancer risks, (2) original papers containing independent data, (3) identification of cancer was confirmed pathologically or histologically, (4) genotype distribution information in cases and controls or odds ratio (OR) with its 95% confidence interval (CI) and P-value and (5) case–control or cohort studies. The major reasons for exclusion of studies were (1) overlapping data, (2) case-only studies and (3) family-based studies.
For each included study, the following information was extracted independently by two investigators (Checklist S1): first author’s surname, publication year, ethnicity, cancer type, number of cases and controls, genotyping method, Hardy-Weinberg equilibrium (HWE) status, source of control groups (population-based controls and hospital-based controls), and genotype frequency in cases and controls. For studies including subjects of different ethnic groups, data were extracted separately and categorized as Asians (e.g. Chinese, Japanese), and Caucasians (i.e. people of European origin). Meanwhile, studies investigating more than one kind of cancer were counted as individual data set only in subgroup analyses by cancer type. Cases that were homozygous for either the CYP2C19*2 or CYP2C19*3 mutation (*2/*2 or *3/*3) and heterozygous for CYP2C19*2 and CYP2C19*3 (*2/*3) were categorized as PMs. Cases that were homozygous for the WT (*1/*1) or heterozygous for the WT and mutation (*1/*2 or *1/*3) were categorized as extensive metabolizers (EMs) [3]. The results were compared and disagreements were discussed and resolved with consensus among all authors. Where essential information was not presented in articles, every effort was made to contact the authors.
Statistical methods
Crude ORs with 95% CIs were used to assess the strength of the association between the CYP2C19 polymorphism and cancer risks. For the CYP2C19 polymorphisms, we estimated the risks of the PMs genotypes on cancer compared with EMs genotypes under recessive model. HWE in the control group was assessed using Fisher’s exact test. Heterogeneity across individual studies was calculated using the Cochran chi-square Q test and I2 followed by subsidiary analysis or by random-effects regression models with restricted maximum likelihood estimation. Random-effects and fixed-effect summary measures were calculated as inverse variance-weighted average of the log OR. The results of random-effects summary were reported in the text because it takes into account the variation between studies [8]. In addition, ethnicity, cancer type, source of controls, and sample size were analyzed as covariates in meta-regression. The significance of the overall OR was determined by the Z-test. Funnel plots and Egger’s linear regression test were used to assess evidence for potential publication bias. In order to assess the stability of the result, sensitivity analyses were performed, each study in turn was removed from the total, and the remaining were reanalyzed. All Statistical analyses were done with Stata software version 10.0 (Stata Corporation, College Station, TX, USA). To assess the credibility of genetic associations, the Baysian false discovery probability (BFDP) was calculated [9]. We chose to calculate BFDP values for two levels of prior probabilities: at a medium or low prior level (0.05 to 10-3) that would be close to what would be expected for a candidate gene; and at a very low prior level (10-4 to 10-6) that would be close to what would be expected for a random SNP. The BFDP thresholds of noteworthiness are 0.80 [9]. P values are two-sided at the P = 0.05 level.
Results
Characteristics of studies
The combined search yielded 416 references. 387 articles were excluded because they clearly did not meet the criteria or overlapping references (Figure 1). Finally, a total of 30 studies with 11,554 cancer cases and 16,592 controls examining the association between the CYP2C19 polymorphism and cancer risk were included in the current meta-analysis [3,4,7,10–35]. The polymorphisms were found to occur in frequencies consistent with HWE in the control populations of the vast majority of the published studies. Of the cases, 74.4% were Caucasian, and 25.6% were Asian populations. The detailed characteristics of the studies included in this meta-analysis are presented in Table 1.
Figure 1. Flow diagram of the study selection process.
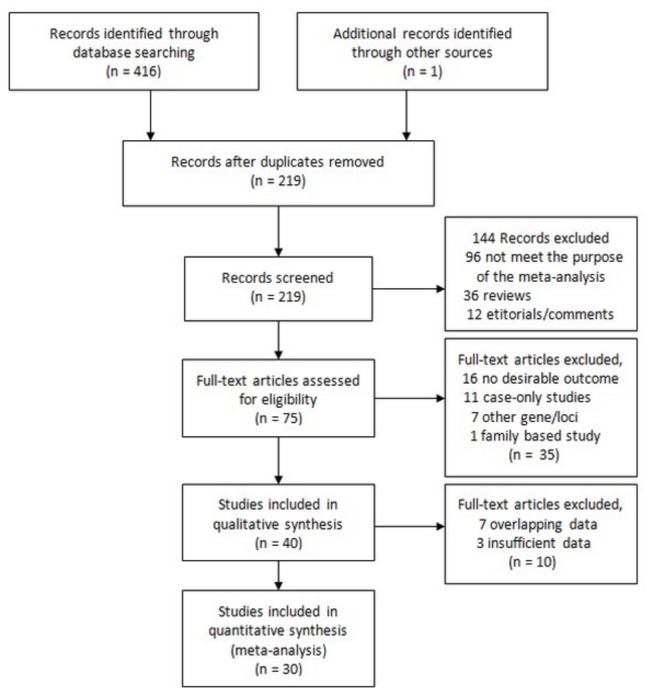
Table 1. Characteristics of the studies included in the meta-analysis.
| Study | Year | Nationality | Ethnicity | Cancer types | No. of case/control | Source of control | Genotyping method |
|---|---|---|---|---|---|---|---|
| Brockmöller [10] | 1996 | German | Caucasian | BLC | 355/340 | Hospital | PCR-RFLP |
| Tsuneoka [11] | 1996 | Japanese | Asian | HCC, LC | 30/64 | Population | PCR-RFLP |
| Wadelius [12] | 1999 | Swedish, Danish | Caucasian | PC | 178/160 | Population | PCR-RFLP |
| Chau [13] | 2000 | Japanese | Asian | HCC | 29/186 | Population | PCR-RFLP |
| Roddam [14] | 2000 | British | Caucasian | Leukaemia | 557/952 | Population | Taqman |
| Sachse [15] | 2002 | British | Caucasian | CRC | 490/592 | Population | PCR-RFLP |
| Shi [16] | 2004 | Chinese | Asian | EC, GC, LC BLC | 607/372 | Population | PCR-RFLP |
| Mochizuki [17] | 2005 | Japanese | Asian | HCC | 44/843 | Population | PCR-RFLP |
| Sugimoto [18] | 2005 | Japanese | Asian | GC | 111/315 | Hospital | PCR-RFLP |
| Landi [19] | 2005 | Spanish | Caucasian | CRC | 351/321 | Population | APEX |
| Zhou [20] | 2006 | Chinese | Asian | EC | 127/254 | Hospital | PCR-RFLP |
| Tamer [21] | 2006 | Turkish | Caucasian | CRC, GC | 182/105 | Hospital | RT-PCR |
| Xing [22] | 2006 | Chinese | Asian | BLC | 108/112 | Hospital | PCR-RFLP |
| Gemignani [23] | 2007 | European | Caucasian | LC | 245/275 | Hospital | Microarray |
| Jiang [24] | 2008 | Chinese | Asian | HCC | 48/88 | Hospital | PCR-RFLP |
| Yang [25] | 2008 | Chinese | Asian | CRC | 83/112 | Hospital | PCR-RFLP |
| Gra [26] | 2008 | Russian | Caucasian | Leukemia | 83/177 | Population | Microarray |
| Yadav [7] | 2008 | Indian | Asian | HNC | 300/300 | Hospital | PCR-RFLP |
| Khedhaier [27] | 2008 | Tunisian | Caucasian | BRC | 304/240 | Population | PCR-RFLP |
| Sameer [28] | 2009 | Palestinian | Caucasian | Leukaemia | 52/200 | Population | PCR-RFLP |
| Justenhoven [29] | 2009 | German | Caucasian | BC | 969/991 | Population | MassARRAY |
| Zhang [30] | 2009 | Chinese | Asian | EC | 46/38 | Hospital | PCR-RFLP |
| Wen [31] | 2009 | Chinese | Asian | BLC | 87/298 | Hospital | Taqman |
| Isomura [3] | 2010 | Japanese | Asian | BTC | 65/566 | Hospital | PCR-RFLP |
| Chang-Claude [4] | 2010 | German | Caucasian | BC | 3131/5478 | Population | MassARRAY |
| Chang [32] | 2010 | Chinese | Asian | HCC | 68/254 | Population | Allele-specific PCR |
| Gan [33] | 2011 | Chinese | Asian | BC | 600/600 | Population | PCR-RFLP |
| Sainz [34] | 2011 | German | Caucasian | CRC | 1759/1776 | Population | SNPlex |
| Feng [35] | 2011 | Chinese | Asian | HNC | 300/300 | Hospital | PCR-RFLP |
| Unpublished data | / | Chinese | Asian | HCC, CRC | 245/283 | Hospital | PCR-RFLP |
CRC: colorectal cancer; LC: lung cancer; BC: breast cancer; BLC: bladder cancer; EC: esophagus cancer; HCC: hepatocellular carcinoma; GC: gastric cancer; PC: prostate cancer; BTC; biliary tract cancer; HNC: head neck cancer
Quantitative Data Synthesis
Significant heterogeneity was present among the 36 data sets from 30 studies (P<10-5). In meta-regression analysis, source of controls (P = 0.13), and HWE status among controls (P = 0.55) did not significantly explain such heterogeneity. By contrast, cancer type (P = 0.02), ethnicity (P = 0.006) and sample size (P = 0.01) were significantly correlated with the magnitude of the genetic effect, explaining 11%, 17% and 15% of the heterogeneity, respectively. Overall, significant associations were found between CYP2C19 PMs genotypes cancer risk when all studies pooled into the meta-analysis. Using random effect model, the summary OR of PMs for cancer was 1.52 [95% CI: 1.23-1.88, P(Z) <10-4, P(Q) <10-5; Figure 2].
Figure 2. Forest plot from the meta-analysis of CYP2C19 PMs genotypes and cancer risk.

In the stratified analysis by cancer type, we found that PMs with the minor variant genotypes had a higher risk of esophagus cancer (OR=2.93, 95%CI: 2.06-4.17, P<10-5), gastric cancer (OR=2.19, 95%CI: 1.47-3.26, P<10-4) and hepatocellular carcinoma (OR=1.66, 95%CI: 1.15-2.39, P=0.006). Furthermore, marginally significant association was also observed for lung cancer and head neck cancer with OR of 2.38 (95%CI: 1.06-5.36, P=0.03) and of 2.40 (95%CI: 1.46-3.93, P=0.001), respectively. However, no significant association was found for other types of cancer (Table 2). This analysis is based on pooling of data from a number of different ethnic populations. When stratifying for ethnicity, an OR of 1.84 (95% CI: 1.44-2.35, P <10-4) and 1.11 (95% CI: 0.87-1.42, P =0.40) resulted for PMs genotype, among Asian and Caucasian populations, respectively. By considering control source subgroups, the OR was 1.38 (95% CI: 0.96-1.98, P =0.08) in population-based controls compared to 1.56 (95% CI: 1.22-1.98, P <10-4) in hospital controls. Subsidiary analyses of HWE status yielded an OR for controls consistent to HWE of 1.42 (95% CI: 1.11-1.80), while similar result was also found for controls deviated from HWE. Analysis restricted to the 5 studies with at least 500 cases, which should be less prone to selective publication than smaller studies, yielded an OR of 1.10 (95% CI: 0.78-1.57, P =0.58) without significant between-study heterogeneity. After applying the BFDP, the polymorphism was identified as a credible positive association (Table S1).
Table 2. Results of meta-analysis and subgroup analysis.
| Sub-group analysis | No. of data sets | No. cases/controls | OR (95%CI) | P(Z) | P(Q)a | I2 | P(Q)b |
|---|---|---|---|---|---|---|---|
| Overall | 36 | 11554/16592 | 1.52 (1.23-1.88) | <10-4 | <10-5 | 60.0% | |
| Cancer type | <10-4 | ||||||
| Colorectal cancer | 6 | 2913/3189 | 1.25 (0.88-1.77) | 0.21 | 0.23 | 27.2% | |
| Breast cancer | 4 | 5004/7309 | 1.02 (0.69-1.50) | 0.92 | 0.27 | 23.7% | |
| Esophagus cancer | 3 | 308/664 | 2.93 (2.06-4.17) | <10-5 | 0.79 | 0% | |
| Hepatocellular carcinoma | 6 | 325/1718 | 1.66 (1.15-2.39) | 0.006 | 0.57 | 0% | |
| Gastric cancer | 3 | 336/792 | 2.19 (1.47-3.26) | <10-4 | 0.28 | 20.7% | |
| Lung cancer | 3 | 471/711 | 2.82 (1.58-5.04) | <10-4 | 0.36 | 1.5% | |
| Leukemia | 3 | 692/1329 | 1.10 (0.37-3.24) | 0.86 | 0.24 | 30.7% | |
| Head neck cancer | 2 | 600/600 | 2.40 (1.46-3.93) | 0.001 | 0.88 | 0% | |
| Bladder cancer | 4 | 662/1122 | 0.67 (0.27-1.66) | 0.39 | 0.005 | 76.5% | |
| Prostate cancer | 1 | 178/160 | 1.13 (0.30-4.27) | 0.86 | NA | NA | |
| Biliary tract cancer | 1 | 65/566 | 0.86 (0.30-2.49) | 0.78 | NA | NA | |
| Ethnicity | <10-4 | ||||||
| Caucasian | 14 | 8656/11607 | 1.11 (0.87-1.42) | 0.40 | 0.17 | 26.9% | |
| Asian | 22 | 2898/4975 | 1.84 (1.44-2.35) | <10-4 | 0.002 | 50.6% | |
| Control source | 0.35 | ||||||
| Population | 19 | 8901/12885 | 1.38 (0.96-1.98) | 0.08 | <10-4 | 72.0% | |
| Hospital | 17 | 2653/3707 | 1.56 (1.22-1.98) | <10-4 | 0.14 | 28.0% | |
| Sample size | 0.0001 | ||||||
| No. cases <500 | 31 | 4538/6795 | 1.64 (1.32-2.05) | <10-5 | 0.002 | 53.4% | |
| No. cases ≥500 | 5 | 7016/9797 | 1.10 (0.78-1.57) | 0.58 | 0.07 | 48.1% | |
| HWE status for controls | 0.37 | ||||||
| Yes | 31 | 9999/14502 | 1.49 (1.19-1.88) | 0.001 | <10-4 | 64.6% | |
| No | 5 | 1555/2090 | 1.82 (1.17-2.85) | 0.008 | 0.75 | 0% |
NA: not available P(Z): Z test used to determine the significance of the overall OR.
aCochran’s chi-square Q statistic test used to assess the heterogeneity in subgroups.
bCochran’s chi-square Q statistic test used to assess the heterogeneity between subgroups.
Sensitivity analyses and Publication bias
A single study involved in the meta-analysis was deleted each time to reflect the influence of the individual dataset to the pooled ORs, and the corresponding pooled ORs were not qualitatively altered (data not shown). The shape of the funnel plot did not indicate any evidence of obvious asymmetry (Figure 3), thus suggesting no publication bias among the studies included. Egger’s test was used to provide further statistical evidence; similarly, the results showed no significant publication bias in this meta-analysis (Egger test, t= 0.08, P = 0.93).
Figure 3. Begg’s funnel plot of CYP2C19 and cancer risk.

Discussion
Cancer is a major public health problem in the world. Despite much investigation, detailed pathogenesis mechanisms of cancer remain a matter of speculation. Large sample and unbiased epidemiological studies of predisposition genes polymorphisms could provide insight into the in vivo relationship between candidate genes and complex diseases. This is the first comprehensive meta-analysis that examined the CYP2C19 polymorphisms and the relationship to cancer susceptibility. Its strength was based on the accumulation of published data giving greater information to detect significant differences. In total, the meta-analysis involved 30 studies for cancer, which provided 11,554 cases and 16,592 controls. Our results indicated that the PMs genotypes of CYP2C19 is a risk factor for developing cancer.
In the stratified analysis by ethnicity, significant associations were found in Asians, while no associations were found in Caucasians. Several factors may contribute to the results that the same polymorphism plays different roles in cancer risk among different ethnic populations. Above all, ethnic differences may attribute to these different results, since the distributions of the CYP2C19 polymorphism were different between various ethnic populations. For instance, the frequencies of CYP2C19*3 polymorphism allele differs from less than 0.5% in Caucasian population [36], 12% in Chinese population [22], to 16% in Japanese populations [17]. However, several studies conducted among Middle East populations which shown a similar PMs genotypes prevalence as Caucasians found a significant association between CYP2C19 PMs genotype and cancer susceptibility [21,27]. It is possible that variation at this locus has modest effects on cancer, but environmental factors may predominate in the progress of cancer, and mask the effects of this variation. On the other hand, study design or small sample size or some environmental factors may affect the results. Most of these included studies did not consider most of the important environmental factors. Thus, the effect of single genetic factor on the risk of cancer may be more pronounced in the presence of other common genetic or environmental risk factors such as smoking, hepatitis virus infection, H. pylori infection. Another explanation of no association between CYP2C19 polymorphism and cancer risk in Caucasians may be that different linkage disequilibrium patterns usually exist in different populations.
In another subgroup analysis by cancer types, we found that CYP2C19 PMs genotypes led to an increased incidence of esophagus cancer, gastric cancer, lung cancer and head neck cancer as well as hepatocellular carcinoma, but not for breast cancer, colorectal cancer, leukemia, prostate cancer, bladder cancer and biliary tract cancer. However, in our meta-analysis, only one or two studies were available for some specific cancers, and they had limited sample size, and hence the results may be capricious and should be interpreted with caution. It should also be considered that the apparent inconsistency of these results may underlie differences in ethnicity, lifestyle and disease prevalence as well as possible limitations due to the relatively small sample size. The current knowledge of carcinogenesis indicates a multi-factorial and multi-step process that involves various genetic alterations and several biological pathways. Thus, it is unlikely that risk factors of cancer work in isolation from each other. And the same polymorphism may play different roles in cancer susceptibility, because cancer is a complicated multi-genetic disease, and different genetic backgrounds may contribute to the discrepancy. And even more importantly, the low penetrance genetic effects of single polymorphism may largely depend on interaction with other polymorphisms and/or a particular environmental exposure.
After stratification by sample size, the association became non-significant when the meta-analysis was restricted to larger studies (at least 500 cancer cases), suggesting a potential small study effects with an overestimate of the true association by smaller studies. Even though the use of a statistical test did not show publication bias among included studies, both theoretical arguments and empirical studies (including surveys and simulations) have demonstrated that the Egger’s test is not powerful enough to be used in publication bias assessment. Therefore, additional studies with much larger sample size are warranted to further validate our results.
When stratified by the source of controls, our results indicated a significantly increased risk among studies using hospital-based controls but not for population-based controls. The reason may be that the hospital-based studies have some biases because such controls may just represent a sample of ill-defined reference population, and may not be representative of the general population very well, particularly when the genotypes under investigation were associated with the disease conditions that the hospital-based controls may have. Therefore, using a proper and representative population-based control subjects is very important to reduce biases in such genetic association studies.
Though polymorphism in CYP2C19 largely accounts for the poor metabolizing status, it has also been reported to influence the metabolism, particularly detoxification of the carcinogens [37]. Using hepatic microsomal preparations, CYP2C19 was shown to metabolize both aromatic amines (AA, nitrosamines) and polycyclic aromatic hydrocarbons (PAHs), found in tobacco smoke and smokeless tobacco [38–40]. Therefore, CYP2C19 polymorphism is considered as one of the factors that determine an individual’s cancer susceptibility by the interindividually different ability of detoxification of carcinogen(s) and/or activation of procarcinogen(s) [41,42]. Homozygous EMs may have higher carcinogen level and potent cell toxicity by the higher ability for bioactivating procarcinogens, whereas PMs may have a higher carcinogen level and potent cell toxicity by the lower ability for detoxificating carcinogens. As for gastric cancer, most patients are infected with H. pylori and have severe active gastritis or atrophic gastritis, suggesting that candidate carcinogens metabolized by CYP2C19 require severe inflammation or atrophic changes induced by H. pylori infection in order to initiate cancerous transformation in gastric epithelial cells [18]. In animal models, chemical carcinogen-induced gastric cancer development was enhanced in the presence of atrophic gastritis caused by chronic H. pylori infection [43,44]. Therefore, it was assumed that the direct effect of candidate carcinogen(s) metabolized by CYP2C19 on the gastric epithelial cells is enhanced in the presence of H. pylori infection [18]. Recently, Wu et al. found that CYP2C19 mRNA expression is highest in hepatocarcinoma tissue, moderate in adjacent normal liver tissue [45]. The significantly elevated expression of CYP2C19 mRNA in hepatocarcinoma suggests an association between the occurrence of hepatocarcinoma and the expression and/or turnover of CYP2C19 mRNA [44].
As like any other meta-analysis, limitations also inevitably existed in the present study. Firstly, our results were based on unadjusted estimates, while a more precise analysis should be conducted if all individual raw data were available, which would allow for the adjustment by other co-variants including age, sex, drinking status, cigarette consumption and other lifestyle. Secondly, the subgroup meta-analyses considering different type of cancer and CYP2C19 were performed on the basis of a fraction of all the possible data to be pooled, so selection bias may have occurred and our results may be over inflated. Since studies among some specific types of cancer are currently limited, further studies including a wider spectrum of subjects should be carried out to investigate the role of the enzyme in different types of cancer. Thirdly, lacking the original data for the included studies limited our further evaluation of potential interactions among gene–gene, gene–environment, or even different polymorphism loci of the same gene, which all may affect cancer risk. Fourthly, hampered by limited number of studies available currently prevent us from making further analysis to identify any interactions between genetic variation and cancer risk as well as ethnic diversity. Finally, some genetic polymorphisms were not in HWE in the current meta-analysis, which may affect the validity of conclusion. These considerations may distort our results.
In summary, our meta-analysis demonstrated an association between CYP2C19 polymorphism and cancer risk among Asian populations, but not among Caucasians. As a statistically significant 1.52-fold increased risk for cancer appeared for individuals with PM genotypes, this result suggests that in the presence of both of the two risk factors, an important number of cancer cases would occur. For future association studies, strict selection of patients, much larger sample size will be required. More studies should also be carried out to examine the impact of CYP2C19 on cancer risk, especially in Caucasian populations. Moreover, gene–gene and gene–environment interactions should also be considered in future studies.
Supporting Information
Summary crude odds ratios (ORs) and 95% Confidence Intervals (95% CI) after applying the BFDP.
(DOCX)
(DOC)
Funding Statement
The authors have no funding or support to report.
References
- 1. Saadat I, Saadat M (2001) Glutathione S-transferase M1 and T1 null genotypes and the risk of gastric and colorectal cancers. Cancer Lett 169: 21–26. doi:10.1016/S0304-3835(01)00550-X. PubMed: 11410321. [DOI] [PubMed] [Google Scholar]
- 2. Guengerich FP (1992) Metabolic activation of carcinogens. Pharmacol Ther 54: 17-61. doi:10.1016/0163-7258(92)90050-A. PubMed: 1528954. [DOI] [PubMed] [Google Scholar]
- 3. Isomura Y, Yamaji Y, Ohta M, Seto M, Asaoka Y et al. (2010) A genetic polymorphism of CYP2C19 is associated with susceptibility to biliary tract cancer. J Gastroenterol 45: 1045-1052. doi:10.1007/s00535-010-0246-0. PubMed: 20549256. [DOI] [PubMed] [Google Scholar]
- 4. Chang-Claude J, Beckmann L, Corson C, Hein R, Kropp S et al. (2010) Genetic polymorphisms in phase I and phase II enzymes and breast cancer risk associated with menopausal hormone therapy in postmenopausal women. Breast Cancer Res Treat 119: 463-474. doi:10.1007/s10549-009-0407-0. PubMed: 19424794. [DOI] [PubMed] [Google Scholar]
- 5. Kubota T, Chiba K, Ishizaki T (1996) Genotyping of S-mephenytoin 4’-hydroxylation in an extended Japanese population. Clin Pharmacol Ther 60: 661–666. doi:10.1016/S0009-9236(96)90214-3. PubMed: 8988068. [DOI] [PubMed] [Google Scholar]
- 6. Goldstein JA (2001) Clinical relevance of genetic polymorphisms in the human CYP2C subfamily. Br J Clin Pharmacol 52: 349-355. doi:10.1046/j.0306-5251.2001.01499.x. PubMed: 11678778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Yadav SS, Ruwali M, Shah PP, Mathur N, Singh RL et al. (2008) Association of poor metabolizers of cytochrome P450 2C19 with head and neck cancer and poor treatment response. Mutat Res 644: 31-37. doi:10.1016/j.mrfmmm.2008.06.010. PubMed: 18644391. [DOI] [PubMed] [Google Scholar]
- 8. DerSimonian R, Laird N (1986) Meta-analysis in clinical trials. Control Clin Trials 7: 177–188. doi:10.1016/0197-2456(86)90046-2. PubMed: 3802833. [DOI] [PubMed] [Google Scholar]
- 9. Wakefield J (2007) A Bayesian measure of the probability of false discovery in genetic epidemiology studies. Am J Hum Genet 81: 208-227. doi:10.1086/519024. PubMed: 17668372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Brockmöller J, Cascorbi I, Kerb R, Roots I (1996) Combined analysis of inherited polymorphisms in arylamine N-acetyltransferase 2, glutathione S-transferases M1 and T1, microsomal epoxide hydrolase, and cytochrome P450 enzymes as modulators of bladder cancer risk. Cancer Res 56: 3915-3925. PubMed: 8752158. [PubMed] [Google Scholar]
- 11. Tsuneoka Y, Fukushima K, Matsuo Y, Ichikawa Y, Watanabe Y (1996) Genotype analysis of the CYP2C19 gene in the Japanese population. Life Sci 59: 1711–1715. doi:10.1016/S0024-3205(96)00507-3. PubMed: 8890945. [DOI] [PubMed] [Google Scholar]
- 12. Wadelius M, Autrup JL, Stubbins MJ, Andersson SO, Johansson JE et al. (1999) Polymorphisms in NAT2, CYP2D6, CYP2C19 and GSTP1 and their association with prostate cancer. Pharmacogenetics 9: 333-340. doi:10.1097/00008571-199906000-00008. PubMed: 10471065. [DOI] [PubMed] [Google Scholar]
- 13. Chau TK, Marakami S, Kawai B, Nasu K, Kubota T et al. (2000) Genotype analysis of the CYP2C19 gene in HCVseropositive patients with cirrhosis and hepatocellular carcinoma. Life Sci 67: 1719–1724. doi:10.1016/S0024-3205(00)00757-8. PubMed: 11021356. [DOI] [PubMed] [Google Scholar]
- 14. Roddam PL, Rollinson S, Kane E, Roman E, Moorman A et al. (2000) Poor metabolizers at the cytochrome P450 2D6 and 2C19 loci are at increased risk of developing adult acute leukaemia. Pharmacogenetics 10: 605-615. doi:10.1097/00008571-200010000-00004. PubMed: 11037802. [DOI] [PubMed] [Google Scholar]
- 15. Sachse C, Smith G, Wilkie MJ, Barrett JH, Waxman R et al. (2002) A pharmacogenetic study to investigate the role of dietary carcinogens in the etiology of colorectal cancer. Carcinogenesis 23: 1839-1849. doi:10.1093/carcin/23.11.1839. PubMed: 12419832. [DOI] [PubMed] [Google Scholar]
- 16. Shi WX, Chen SQ (2004) Frequencies of poor metabolizers of cytochrome P450 2C19 in esophagus cancer, stomach cancer, lung cancer and bladder cancer in Chinese population. World J Gastroenterol 10: 1961-1963. PubMed: 15222046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Mochizuki J, Murakami S, Sanjo A, Takagi I, Akizuki S et al. (2005) Genetic polymorphisms of cytochrome P450 in patients with hepatitis C virus-associated hepatocellular carcinoma. J Gastroenterol Hepatol 20: 1191-1197. doi:10.1111/j.1440-1746.2005.03808.x. PubMed: 16048566. [DOI] [PubMed] [Google Scholar]
- 18. Sugimoto M, Furuta T, Shirai N, Nakamura A, Kajimura M et al. (2005) Poor metabolizer genotype status of CYP2C19 is a risk factor for developing gastric cancer in Japanese patients with Helicobacter pylori infection. Aliment Pharmacol Ther 22: 1033-1040. doi:10.1111/j.1365-2036.2005.02678.x. PubMed: 16268979. [DOI] [PubMed] [Google Scholar]
- 19. Landi S, Gemignani F, Moreno V, Gioia-Patricola L, Chabrier A et al. (2005) A comprehensive analysis of phase I and phase II metabolism gene polymorphisms and risk of colorectal cancer. Pharmacogenet Genomics 15: 535-546. doi:10.1097/01.fpc.0000165904.48994.3d. PubMed: 16006997. [DOI] [PubMed] [Google Scholar]
- 20. Zhou YL, Chen HF, Shi XS, Zhou ZJ, Li GL et al. (2006) A case-control study on the polymorphisms of NQO1 and susceptibility of esophageal cancer. Zhongguo Zhong Liu 15: 659-663. [Google Scholar]
- 21. Tamer L, Ercan B, Ercan S, Ateş N, Ateş C et al. (2006) CYP2C19 Polymorphisms in Patients with Gastric and Colorectal Carcinoma. Int J Gastrointest Cancer 37: 1-5. doi:10.1385/IJGC:37:1:1. PubMed: 17290075. [DOI] [PubMed] [Google Scholar]
- 22. Xing DL, Song DK, Zhang LR (2006) Polymorphism of CYP2C19 gene in patients with bladder cancer. Zhengzhou Da Xue Xue Bao 41: 285-287. [Google Scholar]
- 23. Gemignani F, Landi S, Szeszenia-Dabrowska N, Zaridze D, Lissowska J et al. (2007) Development of lung cancer before the age of 50: the role of xenobiotic metabolizing genes. Carcinogenesis 28: 1287-1293. doi:10.1093/carcin/bgm021. PubMed: 17259654. [DOI] [PubMed] [Google Scholar]
- 24. Jiang W, Shao JG, Lu JR (2008) Relationship between cytochrome P450 2C19 gene polymorphism and primary hepatocellular carcinoma. Wei Chang Bing Xue 13: 39-41. [Google Scholar]
- 25. Yang ZF, Bi LF, Su XL (2008) Cytochrome P450 and tumor susceptibility. Zhongliu Yan Jiu Yu Lin Chuang 20:65-7. [Google Scholar]
- 26. Gra OA, Glotov AS, Nikitin EA, Glotov OS, Kuznetsova VE et al. (2008) Polymorphisms in xenobiotic-metabolizing genes and the risk of chronic lymphocytic leukemia and non-Hodgkin’s lymphoma in adult Russian patients. Am J Hematol 83: 279-287. doi:10.1002/ajh.21113. PubMed: 18061941. [DOI] [PubMed] [Google Scholar]
- 27. Khedhaier A, Hassen E, Bouaouina N, Gabbouj S, Ahmed SB et al. (2008) Implication of Xenobiotic Metabolizing Enzyme gene (CYP2E1, CYP2C19, CYP2D6, mEH and NAT2) Polymorphisms in Breast Carcinoma. BMC Cancer 8: 109. doi:10.1186/1471-2407-8-109. PubMed: 18423013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Sameer AE, Amany GM, Abdela AA, Fadel SA (2009) CYP2C19 genotypes in a population of healthy volunteers and in children with hematological malignancies in Gaza Strip. Can J Clin Pharmacol 16: e156-e162. PubMed: 19193970. [PubMed] [Google Scholar]
- 29. Justenhoven C, Hamann U, Pierl CB, Baisch C, Harth V et al. (2009) CYP2C19*17 is associated with decreased breast cancer risk. Breast Cancer Res Treat 115: 391-396. doi:10.1007/s10549-008-0076-4. PubMed: 18521743. [DOI] [PubMed] [Google Scholar]
- 30. Zhang WL, Sheyhidin I, Wu MB, Zhang Z, Li DS et al. (2009) Study on relations between genetic polymorphisms in CYP2C19, GSTT1 and risk of Kazakh’s esophageal cancer in Xinjiang. Shiyong Zhongliu Za Zhi 24: 232-236. [Google Scholar]
- 31. Wen H, Ding Q, Fang ZJ, Xia GW, Fang J (2009) Population study of genetic polymorphisms and superficial bladder cancer risk in Han-Chinese smokers in Shanghai. Int Urol Nephrol 41: 855-864. doi:10.1007/s11255-009-9560-y. PubMed: 19350405. [DOI] [PubMed] [Google Scholar]
- 32. Chang FH, Zhang ZX, Bai TY, Wang MJ, Fan L et al. (2010) The study on the polymorphisms of CYP2C19 genes associated with susceptibility to liver cancer. Zhongguo Yao Li Tong Xun 27:22-3. [Google Scholar]
- 33. Gan CQ, Wang XY, Cao YD, Ye WX, Liu H et al. (2011) Association of CYP2C19*3 gene polymorphism with breast cancer in Chinese women. Genet Mol Res 10: 3514-3519. doi:10.4238/2011.December.5.3. PubMed: 22180071. [DOI] [PubMed] [Google Scholar]
- 34. Sainz J, Rudolph A, Hein R, Hoffmeister M, Buch S et al. (2011) Association of genetic polymorphisms in ESR2, HSD17B1, ABCB1, and SHBG genes with colorectal cancer risk. Endocr Relat Cancer 18: 265-276. doi:10.1530/ERC-10-0264. PubMed: 21317201. [DOI] [PubMed] [Google Scholar]
- 35. Feng J, Li L, Zhao YS, Tang SQ, Yang HB et al. (2011) Interaction between CYP 2C19*3 polymorphism and smoking in relation to laryngeal carcinoma in the Chinese Han population. Genet Mol Res 10: 3331-3337. doi:10.4238/2011.December.5.9. PubMed: 22183943. [DOI] [PubMed] [Google Scholar]
- 36. De Morais SM, Wilkinson GR, Blaisdell J, Meyer UA, Nakamura K et al. (1994) Identification of a new genetic defect responsible for the polymorphism of (S)-mephenytoin metabolism in Japanese. Mol Pharmacol 46: 594-598. PubMed: 7969038. [PubMed] [Google Scholar]
- 37. Kappers WA, Edwards RJ, Murray S, Boobis AR (2001) Diazinon is activated by CYP2C19 in human liver. Toxicol Appl Pharmacol 177: 68-76. doi:10.1006/taap.2001.9294. PubMed: 11708902. [DOI] [PubMed] [Google Scholar]
- 38. Sugimoto M, Furuta T, Shirai N, Kajimura M, Hishida A et al. (2004) Different dosage regimens of rabeprazole for nocturnal gastric acid inhibition in relation to cytochrome P450 2C19 genotype status. Clin Pharmacol Ther 76: 290-301. doi:10.1016/j.clpt.2004.06.008. PubMed: 15470328. [DOI] [PubMed] [Google Scholar]
- 39. Yamazaki Y, Fujita K, Nakayama K, Suzuki A, Nakamura K et al. (2004) Establishment of ten strains of genetically engineered Salmonella typhimurium TA1538 each co-expressing a form of human cytochrome P450 with NADPH-cytochrome P450 reductase sensitive to various promutagens. Mutat Res 562: 151-162. doi:10.1016/j.mrgentox.2004.06.003. PubMed: 15279838. [DOI] [PubMed] [Google Scholar]
- 40. Fujita K, Kamataki T (2001) Predicting the mutagenicity of tobacco-related N-nitrosamines in humans using 11 strains of Salmonella typhimurium YG7108, each coexpressing a form of human cytochrome P450 along with NADPH-cytochrome P450 reductase. Environ Mol Mutagen 38: 339-346. doi:10.1002/em.10036. PubMed: 11774366. [DOI] [PubMed] [Google Scholar]
- 41. Suzuki S, Muroishi Y, Nakanishi I, Oda Y (2004) Relationship between genetic polymorphisms of drug-metabolizing enzymes (CYP1A1, CYP2E1, GSTM1, and NAT2), drinking habits, histological subtypes, and p53 gene point mutations in Japanese patients with gastric cancer. J Gastroenterol 39: 220-230. doi:10.1007/s00535-003-1281-x. PubMed: 15064998. [DOI] [PubMed] [Google Scholar]
- 42. Gao C, Takezaki T, Wu J, Li Z, Wang J et al. (2002) Interaction between cytochrome P-450 2E1 polymorphisms and environmental factors with risk of esophageal and stomach cancers in Chinese. Cancer Epidemiol Biomarkers Prev 11: 29-34. PubMed: 11815398. [PubMed] [Google Scholar]
- 43. Sugiyama A, Maruta F, Ikeno T, Ishida K, Kawasaki S et al. (1998) Helicobacter pylori infection enhances N-methyl-N-nitrosourea-induced stomach carcinogenesis in the Mongolian gerbil. Cancer Res 58: 2067-2069. PubMed: 9605743. [PubMed] [Google Scholar]
- 44. Watanabe T, Tada M, Nagai H, Sasaki S, Nakao M (1998) Helicobacter pylori infection induces gastric cancer in mongolian gerbils. Gastroenterology 115: 642-648. doi:10.1016/S0016-5085(98)70143-X. PubMed: 9721161. [DOI] [PubMed] [Google Scholar]
- 45. Wu M, Chen S, Wu X (2006) Differences in cytochrome P450 2C19 (CYP2C19) expression in adjacent normal and tumor tissues in Chinese cancer patients. Med Sci Monit 12: BR174–8:BR174- 8. PubMed: 16641871 [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Summary crude odds ratios (ORs) and 95% Confidence Intervals (95% CI) after applying the BFDP.
(DOCX)
(DOC)