

Abstract
Amphibians, like other animals, generate corticosterone or cortisol glucocorticoid responses to stimuli perceived to be threatening. It is generally assumed that the corticosterone response of animals to capture and handling reflects the corticosterone response to stimuli such as the sight of a predator that are thought to be natural stressors. Fijian ground frogs ( Platymantisvitiana ) are preyed upon by the introduced cane toads ( Rhinellamarina ), and we used ground frogs to test the hypothesis that the sight of a predator will induce a corticosterone stress response in an amphibian. Urinary corticosterone metabolite concentrations increased in male ground frogs exposed to the sight of a toad for 1, 3 or 6 h, whereas corticosterone did not change in frogs exposed to another male ground frog, a ball, or when no stimulus was present in the test compartment. The frogs exposed to a toad initially moved towards the stimulus then moved away, whereas frogs exposed to another frog moved towards the test frog and remained closer to the frog than at the start of the test. Tonic immobility (TI) was measured as an index of fearfulness immediately after the test exposure of the frogs to a stimulus. The duration of TI was longer in frogs exposed to a toad than to another frog or to a ball. The results provide novel evidence that the sight of a predator can induce a corticosterone response and lead to increased fearfulness in amphibians. In addition, they show that endemic frogs can recognise an introduced predator as a threat.
Introduction
The hypothalamo-pituitary interrenal (HPI) axis is the central neuroendocrine pathway for physiological stress responses in amphibians. The glucocorticoid hormone corticosterone, the major stress hormone in amphibians (also found in birds, reptiles and many rodents), is secreted by the HPI-axis when individuals respond to a stressor [1–5]. This short-term increase in corticosterone promotes key changes in behaviour and physiology that enable individuals to cope with stress [6–10]. Some of the key behaviours affected by corticosterone in amphibians and also in other vertebrates (small mammals and lizards) include initiation of hiding and defensive behaviours [11–13], increased locomotor activity [14,15], and increased thermoregulatory behaviour [16,17]. The sight of a predator can induce a corticosterone response in birds [18] and exposure to predator odour can induce corticosterone responses in male Sprague–Dawley rats [19]. Thus, corticosterone measures could be used to explore the physiological sensitivity of amphibians to environmental stressors, such as predation and pathogenic infections [20] as well as experimental stressors, such as capture, transportation and toe-clipping [21–23]. Introduced species have had a significant impact on natural ecosystems worldwide, especially through predation [24,25]. Introduced species often cause native animal populations to decline because the native species are oblivious to the possible threat of predation therefore are unable to generate adequate physiological and behavioural responses towards the novel invader [26]. Naturally, prey species should be able to generate physiological and behavioural responses towards a visual predator by increasing stress hormone responses or by hiding, in order to increase its chances of escape and survival [27]. However, some endemic island species are unable to show any such behavioural adjustment due to lack of sufficient evolutionary time in residence with the introduced species [26,28,29]. Currently, there is no empirical evidence to show whether endemic amphibian species are able to recognize novel introduced predatory species as potential physiological stressors.
The Fijian ground frog ( Platymantisvitiana ) is currently listed as Endangered (B1ab (v) ver 3.1) by the International Union for Conservation of Nature (IUCN) 2008 (see [30]:). The endemic ground frog population on Viwa Island are preyed upon by the cane toad ( Rhinellamarina ) [31–33]. Evidence dating back 20 years highlights that cane toads predate upon froglets and juveniles [34]. The introduced cane toads have also been associated with mortality of native amphibian larvae in Australia [35]. It has also been shown that introduced cane toads have caused rapid alteration of both morphology and physiology of a native Australian species, the red-bellied black snake ( Pseudechisporphyriacus ) [36]. In this study, we investigated whether exposure of Fijian ground frogs to a cane toad induced a corticosterone response in the frogs. The degree of fear experienced by an animal when faced by a stimulus that represents a threat is generally correlated with the magnitude of the associated corticosterone response [37], and tests such as tonic immobility (TI) can be used to measure fearfulness in animals [38] including amphibians [39]. We used TI tests to measure fearfulness in Fijian ground frogs exposed to the sight of cane toads. We tested the hypothesis that the sight of a cane toad will induce a corticosterone stress response and fearfulness in the Fijian ground frog. Furthermore, we also investigated whether the presence of cane toads altered the movement of the ground frogs to provide an insight into the potential effects of toads on the locomotory behaviour of ground frogs.
Materials and Methods
(a) Ethics statement
All experiments were conducted according to relevant national and international guidelines. Ethical clearances were obtained from the Department of Environment, Fiji Islands for the collection of the endangered Fijian ground frogs on Viwa Island (18o00´S, 175o00´E), a small (60 ha) island located 900 m off the coast of mainland Viti Levu, Fiji. The traditional owner of the land (Chief of Viwa Island; Ratu Dovi Komaisavai) permitted us to undertake this research on Viwa through a traditional ceremony that was conducted prior to this study. The Griffith University Animal Ethics Committee (ENV/13/11/AEC) that approved the interventions with the amphibians in this study. All frogs were released back on into their natural habitat upon completion of the study in accordance with the Australian health check standards for amphibians [40].
(b) Field sampling and captive housing
We used adult male Fijian ground frogs for this study because of their easy accessibility compared to female ground frogs, and to reduce any confounding effects of reproductive condition on subsequent stress and behaviour measurements. Ground frogs and cane toads were caught at night (2000-2200h) on Viwa Island within shared terrestrial habitats in December 2011 during the annual breeding season of the ground frogs. Adult male frogs were identified by their distinct ‘stress call’ upon capture, as described earlier [41] while adult male cane toads were identified based on morphological characteristics [42] and a rough dorsal surface. Adult male ground frogs weighed 33.5 + 2.5 g, and had mean snout-vent length (SVL) of 35 mm (n = 112; 91 experimental frogs and 21 test stimulus frogs). Adult male cane toads weighed 55.0 + 1.5 g, and had mean SVL of 65 mm (n = 21 test stimulus toads). We used a urinary corticosterone metabolite enzyme-immunoassay (EIA) to measure urinary corticosterone metabolite concentrations (referred to hereafter as urinary corticosterone concentrations) using our established protocols for anuran amphibians [21,43–46]. Urine samples were collected from individual frogs on the night of capture on Viwa Island. For urine sampling, each frog was gently held above a sterile plastic cup (diameter = 100 mm) and its underbelly abdomen was briefly massaged to promote urination, which occurred always within 1 min. The volume of urine excreted by each frog ranged from 200 µL -3 mL. Labelled urine samples were kept in a freezer prior to assay (no longer than 1 month). Frogs and toads were transported overnight from Viwa Island to a nearby field station (within 5 km from Viwa Island) on mainland Viti Levu, where the experiments were conducted. At the field station, the frogs were housed individually in plastic “home containers” with meshed lids (10 x 35 x 10 cm). Frogs were provided with food (house crickets, Achetus domesticus ) every second day and distilled water was sprinkled twice daily into the home container to keep it moist. The arena containing the home containers had a similar photoperiod to Viwa Island. Frogs were adjusted to captivity for 9 days before the experiments began. This adjustment period was selected based on our earlier results on the physiological adjustment of amphibians to captivity [43,45]. Urine samples were collected from each frog after 0.5, 5 and 9 days to ensure that corticosterone concentrations had returned after 9 days to concentrations similar to the time of capture concentrations.
(c) Experimental design
The responses of adult male ground frogs to three experimental treatments were examined 10 days after capture on Viwa Island. Individual frogs were introduced to either a cane toad, another adult male ground frog or to a ball, or not exposed to any stimulus. The home container, which was also used as the experimental chamber, was divided in half using plastic wire mesh (15 mm x 15 mm) through which frogs could perceive (see and smell) the test stimulus. This was confirmed by placing a live or a dead house cricket in the test compartment and noticing the frog’s movement in the test chamber.
Male ground frogs (n = 21) and toads (n = 21) that were used as test stimuli were caught on Viwa Island on the same night in December 2011 as the experimental frogs, and were acclimated in separate home containers away from the arena where the experimental frogs were kept. On the day of the experiment, male frogs that were used as controls (n = 28) were sampled for urine 0, 1, 3 or 6 h (n = 7 frogs per time period) without any stimulus being present in the test compartment. Experimental male ground frogs (n = 63 frogs) were divided into groups and subjected to one of the three treatments (cane toad, another adult male ground frog or a ball), with a single urine sample collected 1, 3 or 6 h (n = 7 frogs sampled per time period for each treatment) after the experiment began. At the start of each experimental session a piece of black cardboard was placed in the middle of the test chamber and then a toad, frog or the ball was placed into the test compartment by the observer. The cardboard was removed so that the male ground frog could then see the test stimulus. The control frogs were also exposed to the placement and then urine was collected upon removal of the cardboard divider. Sampling time periods of 1, 3 or 6 h were used based on our previous works that showed that physiological stress responses of anurans (e.g. ground frogs) appeared in urinary corticosterone metabolites within 6 h after exposure to experimental stressor, such as adrenocorticotropic hormone (ACTH) injection [44].
(d) Behaviour
A rectangular sheet of white paper with lines drawn with a permanent marker across the sheet at 20, 40, 60 and 80% of the length of the area of each home container in which the ground frog was present was placed beneath the home container before the tests began so the observer could see the lines from a standard distance of 0.5 m. The lines divided the area in which the male frog was present in five locations which were termed 1, 2, 3, 4 and 5, with location 1 closest to the test compartment and location 5 furthest away from the test compartment at the start of the treatment. The location of the frog was recorded at the start of each test (0 min location). The location was then recorded 5, 15, 20, 40 and 60 min after the test began for frogs sampled for urine at 1 h; or 5, 15, 20, 40, 60, 120, 180 min after the test began for frogs sampled for urine at 3 h; or 5, 15, 20, 40, 60, 120, 180, 240 and 360 min after the test began for frogs sampled for urine at 6 h. Each frog was picked up for urine collection immediately after the final location observation was made for each treatment.
The duration of tonic immobility (TI, also known as the immobility reaction [47]; was measured after urine was collected from each frog. The tonic immobility test is a quantitative measure of fear that has been used in previous amphibian studies [47,48]. Each frog was removed from its home container and placed on its back on the test surface (flat plastic board 15 x 15 x 10 mm) and a timer was started immediately. The frog was gently held by the hands until the hands could be moved away and the frog remained on its back, following the procedure of [39]. The latency time (Sec) was the time when the hands could be moved away while the frog remained on its back. The time taken for the frog to upright was recorded as the duration of immobility. Frogs were returned into their home containers at the end of the test and returned to Viwa Island at the end of the experiment.
(e) Corticosterone enzyme-immunoassay
A urinary corticosterone enzyme-immunoassay (EIA) that was previously validated for measuring conjugated corticosterone metabolites in frog urine was used for this study [21,43–45]. Urinary corticosterone metabolite concentrations in frog urine were determined using a polyclonal anti-corticosterone antiserum (CJM06, UC, Davis California) diluted 1: 45 000, horseradish peroxidise-conjugated corticosterone label diluted 1: 120 000 and corticosterone standards (1.56–400 pgwell-1). Cross reactivity of the CJM06 anti-corticosterone antiserum was reported earlier as 100% with corticosterone, 14.25% with desoxycorticosterone and 0.9% with tetrahydrocorticosterone [44]. Intra- (within) and inter- (between) assay coefficients of variation were determined from high- (~70%) and low- (~30%) binding internal controls run on all assays. Intra assay CVs were 3.9% and 4.4% for low, and high percentage bound controls, and inter-assay CVs were 4.2% and 6.8% for low, and high percentage bound-controls respectively. Assay sensitivity was calculated as the value 2 standard deviations from the mean response of the blank (zero binding) samples. Mean assay sensitivity for the corticosterone EIA was 1.16 + 0.27 pg/well (n = 24). Creatinine (Cr) was measured in each frog urine sample as an index of the urinary corticosterone metabolite concentrations because it is excreted at a constant rate and is therefore a good index of the amount of time over which hormones have been metabolized into the urine regardless of the volume of the sample [44]. Urinary corticosterone metabolite concentrations were expressed in relation to urinary creatinine concentrations as pg corticosterone metabolites/ug Cr.
(f) Statistical analysis
Statistical analyses were performed using Prism (GraphPad Software Inc.). Urinary corticosterone metabolite concentrations were transformed to logarithms, and D’Agostino & Pearson omnibus test confirmed normality and homogeneity of variances for log-transformed data. Urinary corticosterone metabolite concentrations in adult male ground frogs after they were captured from the wild and transferred into captivity were compared between days using one-way Repeated Measures ANOVA. Post-hoc comparisons were done between times (days 0, 0.5, 5 and 9) using Bonferroni’s Multiple Comparison Test.
All of the physiological and behavioural data (urinary corticosterone metabolite concentrations, mean locations of frogs in the test compartment and mean tonic immobility scores) at times 1, 3 or 6 h since the start of each treatment were compared between groups (control and treatments) using two-way ANOVA followed by post hoc comparisons.
Comparisons of the physiological and behavioural data between times (1, 3 or 6 h) within each frog group (control and treatments) were done using two-way ANOVA followed by post-hoc comparisons. Data are presented as individual points or mean + S.E.
Results
(a) Urinary corticosterone after capture
Urinary corticosterone concentrations in all male frogs (n = 91) had increased from 0.5 days after capture (Figure 1). Corticosterone decreased between 0.5 and 5 days and, in most frogs, decreased further from 5 to 9 days after capture (Figure 1). Mean urinary corticosterone concentrations in the frogs changed significantly after capture (One-way ANOVA, F 3,273 = 216.8, p < 0.0001, Figure 1). The mean urinary corticosterone concentration increased between the time of capture and day 0.5 (p < 0.05), decreased on day 5 (p < 0.05), and after 9 days in captivity did not differ from initial concentrations at the time of capture (p > 0.05).
Figure 1. Mean (+ S.E) urinary corticosterone metabolite concentrations at capture (0 days) and after 0.5, 5 and 9 days in captivity in 91 male Fijian ground frogs.

(b) Urinary corticosterone responses to test stimuli
A two-way ANOVA of mean urinary corticosterone metabolite concentration at 1, 3, or 6 h after exposure to a test stimulus showed that corticosterone metabolite concentrations varied with time (F 3,96 = 29.24, p < 0.001) and treatment (F 3,96 = 153.7, p < 0.001), with a significant interaction between time and treatment (F 9,96 = 26.04, p < 0.001). There were significant differences in mean urinary corticosterone metabolite concentrations between frogs in the control group and frogs that saw the cane toad 1, 3 and 6 h after exposure to the sight of the toad (p < 0.001; Figure 2). There were no significant differences in mean urinary corticosterone metabolite concentrations at these times between frogs that saw another frog or a ball compared with the control group (p > 0.05 for all comparisons; Figure 2). The mean urinary corticosterone concentrations were significantly different at times (1, 3 or 6 h) for frogs that saw the toad compared to those that saw the ball or another frog (p < 0.05 for all comparisons). Mean urinary corticosterone metabolite concentrations increased in frogs that saw a toad and were significantly higher compared with initial 0 h corticosterone concentrations at times 1, 3 and 6 h (p < 0.001; Figure 2). The highest mean urinary corticosterone metabolite concentration was at 6 h (105.14 + 5.4 pg/µg Cr) for the frogs that were exposed to the cane toad. Corticosterone did not increase from initial concentrations in frogs that saw another frog or a ball or for the control group (Figure 2).
Figure 2. Mean (+ S.E.) urinary corticosterone metabolite concentrations in male Fijian ground frogs during exposure to a cane toad, a ball, another male ground frog or control (no stimulus).

Sample sizes at each time point were n = 7. Mean urinary corticosterone data for each four group (control and three treatments) were used to obtain the data for time = 0 h.
(c) Behaviour
Adult male ground frogs were usually sitting in the middle of their home container (location number 3) before being subjected to the test stimulus. The locations of the ground frogs changed when the frogs were exposed to some of the test stimuli. Frogs exposed to cane toads moved towards the test compartment within the first 15-20 min and then moved away and stayed as far away as possible for the remainder of the test period (Figure 3). Frogs exposed to a male ground frog in the test compartment moved towards the compartment within the first 15-20 min and stayed close to the test compartment throughout the remainder of the test period (Figure 3). There was little or no change in the locations of the ground frogs in the control group and in the presence of a ball. There was a significant effect of test stimulus (F 3,96 = 94.98, p < 0.0001) and time (F 3,96 = 3.22, p = 0.026) on the location of the male ground frogs, and a significant interaction between test stimulus and time (F 9,96 = 11.39, p < 0.0001). There was a significant difference in locations of frogs over 1, 3 or 6 h between frogs subjected to the toad or another frog (p < 0.05; Figure 3A–C). There was no significant difference in locations of the frogs between those subjected to a ball or a control group (p > 0.05).
Figure 3. Mean (+ S.E.) locations of the Fijian ground frogs during exposure to a cane toad or another frog.

Location 1 was closest to the test compartment and location 6 was furthest away from the test compartment. The location of the frog was recorded at 5, 15, 20, 40 and 60 min after the test began (Fig. 3A, frogs sampled up to 60 min) or 5, 20, 40, 60, 120, 180 (Fig. 3B, frogs sampled up to 180 min) or 5, 20, 40, 60, 120, 180, 240 and 360 min (Fig. 3C, frogs sampled up to 360 min). Sample sizes at each time point were n = 7.
The mean duration of tonic immobility (TI) of the male ground frogs differed significantly between stimuli (F 3,72 = 56.09, p < 0.0001, Figure 4) and time (F 3,72 = 11.78, p < 0.0001, Figure 4). There was no significant interaction between treatment and time (F 3,72 = 1.97, p = 0.080, Figure 4). The frogs that were subjected to a toad had the longest durations of TI at 1, 3 and 6 h (Figure 4). The highest mean TI duration (75.0 + 13.2 seconds) was recorded at 3 h for the frog group that was subjected to the sight of a toad. There was no significant difference between time in mean TI durations for frogs that saw a frog or ball (Figure 4; p > 0.05 for all time comparisons). There was a significant difference between the times in mean TI durations of the control frogs and frogs that had perceived the cane toad (Figure 4; p < 0.05 for all time comparisons). The mean TI durations of the frogs that saw another frog were significantly shorter than those of frogs that saw a toad, and significantly longer than the mean TI durations of frogs that saw a ball 1, 3 or 6 h (Figure 4; p < 0.05).
Figure 4. Histogram showing mean durations of tonic immobility (seconds) of male Fijian ground frogs after exposure to a cane toad, a ball, another frog or no stimulus.
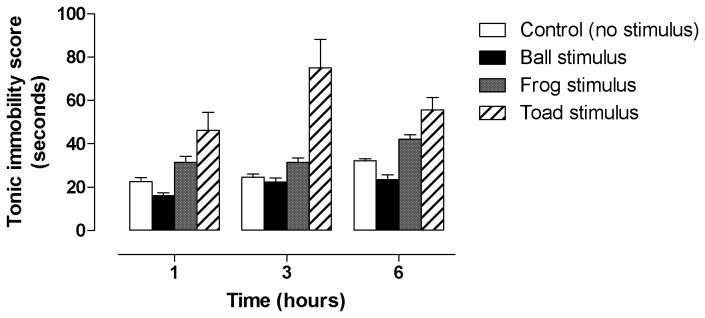
Sample sizes at each time point were n = 7.
Discussion
This is the first study to show that amphibians have a corticosterone response to the sight of an invasive predator. Frogs that saw the predator also had greater fearfulness, as measured by the duration of tonic immobility (TI) immediately after exposure to the predator, than frogs that saw another frog or saw a ball. These results show that the Fijian ground frogs perceived the sight of a cane toad to be a threat, and experienced fearfulness at the same time as their hypothalamo-pituitary interrenal (HPI) axis was activated and corticosterone secretion increased.
The adult male ground frogs experienced greater fearfulness, as measured by higher durations of TI in the presence of cane toads than in the presence of another frog or a novel object (ball). Tonic immobility has been demonstrated in a wide range of animals, including birds, fish, rabbits and guinea pigs, to reduce the probability of continued attacks by an intruder [49]. Experimental elevation of corticosterone has also been shown to increase the duration of immobility in birds [50]. Furthermore, the male ground frogs altered their behaviour (moved further away from the test chamber) following visual detection of the cane toad in the experimental chamber. Ground frogs are often found in areas of minimal cover, such as open-exposed bare ground and exposed perches on tree trunks at night-time [51]. Cane toad encounters with ground frogs are not uncommon and the frogs usually evade the toads by retreating into terrestrial substrates, such as leaf litter, or moving up into the vegetation [41]. In their study [52], demonstrated that behaviour towards any biological stressor (such a predator) could differ between frog species depending on micro-habitat preferences. For example, more terrestrial dwelling northern leopard frogs ( Rana pipiens ) escaped towards land while their aquatic dwelling congener, the green frog ( R . clamitans ), escaped towards the water when approached by snakes or humans around natural ponds. Seeking refuge following a stressful encounter could be beneficial if the intruder is a visual hunter, but may prove detrimental when the predator is able to enter these substrates. There is empirical evidence that the spatial distributions of the cane toads and Fijian ground frogs on Viwa Island overlap quite significantly [31]. We studied the male ground frogs during breeding thus future studies should investigate whether responses (fearfulness and corticosterone responses) of male frogs are similar outside the breeding season, including their correlation with testosterone [53,54] and body condition [7]. Similar testing could be done on female frogs to test if physiological responsiveness to the cane toad is affected by sex, condition and reproductive stage [55].
Behavioural and physiological stress responses are under some level of genetic control [56], and responsiveness could increase from natural selection pressures, such as that imposed by predation [57]. We have shown that ground frogs perceive cane toads as a threat and move away from toads. On Viwa Island other potential predators of ground frogs, such as the introduced small Indian mongoose ( Herpestes javanicus ) have never been present, while rats ( Rattus spp. ) and feral cats (Felis catus) have already been eradicated from the island [31]. It could be that there has been selection pressure in the frogs to recognise toads as predators, which is also not known. In future, it would be worthwhile to investigate whether the ground frogs are able to show similar physiological and behavioural responses in the presence of potential native predators such snakes ( Candoia sp. ) as well as in the presence of invasive species (such as rats and feral cats). Certain native species that have evolved in the absence of any intruders could also display specific behaviours in response to the intruder. A classic example was an earlier study by [58] in which the marine iguana ( Amblyrhynchus cristatus ) that has been isolated from mainland populations for over 15 million years was tested for behavioural and physiological stress hormone responses to some novel predators. Their study demonstrated that the ‘naïve’ marine iguana were able to generate anti-predator responses (both stress hormonal and behavioural responses) in the presence of novel predators (dogs and cats) [58]. Both this study and another by [36] emphasized that physiological stress and behavioural responses of native species against novel introduced species demonstrate cases of rapid evolution under the influence of strong predatory selection pressure. Earlier [18], found a marked increase in plasma corticosterone in captive great tits ( Parus major ) that saw an owl, which is a natural predator. In another study [59], studied the stress hormone responses of free-living male pied flycatchers ( Ficedula hypoleuca ) that were exposed for 10 min to a stuffed weasel ( Mustela sp. ) model, a stuffed great spotted woodpecker ( Dendrocopos major ), or a stuffed flycatcher that was placed on or near their nest box to act as a territorial intruder. Exposure to a weasel or to a territorial intruder elevated plasma corticosterone concentrations in flycatchers caught 10-25 min after the beginning of a test stimulus.
Overall, this study has provided novel evidence that amphibians can show a corticosterone response to the sight of a predator. Determining whether the physiological stress and behavioural responses of ground frogs towards cane toads are a result of natural selection or learnt responses could be tested by replicating our experimental protocols with a naïve population of ground frogs from neighbouring Gau Island (located at 18.00° S and 179.30 ° E), where there are currently no cane toads present [2,60].
Acknowledgments
We wish to thank the youth of Viwa Island for providing moral support to our research team and for assisting with the field work.
Funding Statement
This research was supported through a Griffith University postdoctoral Research Fellowship awarded to E.J.N. and financial support was provided by the Rufford Small Grants Foundation. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Cockrem JF, Barrett DP, Candy EJ, Potter MA (2009) Corticosterone responses in birds: individual variation and repeatability in Adelie penguins (Pygoscelis adeliae) and other species, and the use of power analysis to determine sample sizes. Gen Comp Endocrinol 163: 158-168. doi:10.1016/j.ygcen.2009.03.029. PubMed: 19351536. [DOI] [PubMed] [Google Scholar]
- 2. Narayan EJ, Cockrem JF, Hero J-M (2012) Individual variation in urinary corticosterone metabolite responses in two closely related species of Fijian frogs. Gen Comp Endocrinol 177: 55-61. doi:10.1016/j.ygcen.2012.02.007. PubMed: 22387322. [DOI] [PubMed] [Google Scholar]
- 3. Graham C, Narayan E, McCallum H, Hero J-M (2013) Non-invasive monitoring of stress physiology within free-living highland and lowland populations of native Australian Great Barred Frogs (Mixophyes fasciolatus). Gen Comp Endocrinol 191: 24-30. doi:10.1016/j.ygcen.2013.05.012. [DOI] [PubMed] [Google Scholar]
- 4. Narayan EJ, Molinia FC, Cockrem JF, Hero J-M (2012) Individual variation and repeatability in urinary corticosterone metabolite responses to capture in the cane toad (Rhinella marina). Gen Comp Endocrinol 175: 284-289. doi:10.1016/j.ygcen.2011.11.023. PubMed: 22137908. [DOI] [PubMed] [Google Scholar]
- 5. Narayan EJ, Cockrem JF, Hero J-M (2013) Are baseline and short-term corticosterone stress responses in free-living amphibians repeatable? Comp Biochem Physiol A Mol Integr Physiol 164: 21-28. doi:10.1016/j.cbpa.2012.10.001. PubMed: 23047053. [DOI] [PubMed] [Google Scholar]
- 6. Cockrem JF (2007) Stress, corticosterone responses and avian personalities. J Ornithol 148: S169-S178. doi:10.1007/s10336-007-0175-8. [Google Scholar]
- 7. Narayan EJ, Cockrem JF, Hero J-M (2013) Repeatability of baseline corticosterone and short-term corticosterone stress responses, and their correlation with testosterone and body condition in a terrestrial breeding anuran (Platymantis vitiana). Comp Biochem Physiol A Mol Integr Physiol 165: 304-312. doi:10.1016/j.cbpa.2013.03.033. PubMed: 23562802. [DOI] [PubMed] [Google Scholar]
- 8. Narayan EJ, Webster K, Nicolson V, Mucci A, Hero J-M (2013) Non-invasive evaluation of physiological stress in an iconic Australian marsupial: the Koala (Phascolarctos cinereus). Gen Comp Endocrinol 187: 39-47. doi:10.1016/j.ygcen.2013.03.021. PubMed: 23583768. [DOI] [PubMed] [Google Scholar]
- 9. Narayan E, Cockrem JF, Hero J-M (2013) Changes in serum and urinary corticosterone and testosterone in adult male cane toad (Rhinella marina) during short-term capture and handling Gen Comp Endocrinol 191: 225–230. doi:10.1016/j.ygcen.2013.06.018. [DOI] [PubMed] [Google Scholar]
- 10. Kindermann C, Narayan EJ, Wild F, Wild CH, Hero J-M (2013) The effect of stress and stress hormones on dynamic colour-change in a sexually dichromatic Australian frog. Comp Biochem Physiol A Mol Integr Physiol 165: 223-227. doi:10.1016/j.cbpa.2013.03.011. PubMed: 23507571. [DOI] [PubMed] [Google Scholar]
- 11. Kalynchuk LE, Gregus A, Boudreau D, Perrot-Sinal TS (2004) Corticosterone increases depression-like behavior, with some effects on predator odor-induced defensive behavior, in male and female rats. Behav Neurosci 118: 1365-1377. doi:10.1037/0735-7044.118.6.1365. PubMed: 15598145. [DOI] [PubMed] [Google Scholar]
- 12. Thaker M, Lima SL, Hews DK (2009) Acute corticosterone elevation enhances antipredator behaviors in male tree lizard morphs. Horm Behav 56: 51-57. doi:10.1016/j.yhbeh.2009.02.009. PubMed: 19281811. [DOI] [PubMed] [Google Scholar]
- 13. Narayan E, Hero JM, Evans N, Nicolson V, Mucci A (2012) Non-invasive evaluation of physiological stress hormone responses in a captive population of the greater bilby (Macrotis lagotis). Endangered Species Res 18: 279-289. doi:10.3354/esr00454. [Google Scholar]
- 14. Belliure J, Clobert J (2004) Behavioral sensitivity to corticosterone in juveniles of the wall lizard, Podarcis muralis . Physiol Behav 81: 121-127. doi:10.1016/j.physbeh.2004.01.008. PubMed: 15059691. [DOI] [PubMed] [Google Scholar]
- 15. Ricciardella LF, Bliley JM, Feth CC, Woodley SK (2010) Acute stressors increase plasma corticosterone and decrease locomotor activity in a terrestrial salamander (Desmognathus ochrophaeus). Physiol Behav 101: 81-86. doi:10.1016/j.physbeh.2010.04.022. PubMed: 20434476. [DOI] [PubMed] [Google Scholar]
- 16. Preest MR, Alison C (2008) Corticosterone treatment has subtle effects on thermoregulatory behavior and raises metabolic rate in the New Zealand common gecko, Hoplodactylus maculatus . Physiol Biochem Zool 81: 641-650. doi:10.1086/590371. PubMed: 18752421. [DOI] [PubMed] [Google Scholar]
- 17. Narayan EJ, Cockrem JF, Hero JM (2012) Effects of temperature on urinary corticosterone metabolite responses to short-term capture and handling stress in the cane toad (Rhinella marina). Gen Comp Endocrinol 178: 301-305. doi:10.1016/j.ygcen.2012.06.014. PubMed: 22728158. [DOI] [PubMed] [Google Scholar]
- 18. Cockrem JF, Silverin B (2002) Sight of a predator can stimulate a corticosterone response in the great tit (Parus major). Gen Comp Endocrinol 125: 248-255. doi:10.1006/gcen.2001.7749. PubMed: 11884070. [DOI] [PubMed] [Google Scholar]
- 19. Muñoz-Abellán C, Daviu N, Rabasa C, Nadal R, Armario A (2009) Cat odor causes long-lasting contextual fear conditioning and increased pituitary-adrenal activation, without modifying anxiety. Horm Behav 56: 465-471. doi:10.1016/j.yhbeh.2009.08.002. PubMed: 19699202. [DOI] [PubMed] [Google Scholar]
- 20. Kindermann C, Narayan EJ, Hero J-M (2012) Urinary corticosterone metabolites and chytridiomycosis disease prevalence in a free-living population of male Stony Creek frogs (Litoria wilcoxii). Comp Biochem Physiol A Mol Integr Physiol 162: 171-176. doi:10.1016/j.cbpa.2012.02.018. PubMed: 22387450. [DOI] [PubMed] [Google Scholar]
- 21. Narayan EJ, Molinia FC, Christina K, Cockrem JF, Hero J-M (2011) Urinary corticosterone responses to capture and toe-clipping in the cane toad (Rhinella marina) indicate that toe-clipping is a stressor for amphibians. Gen Comp Endocrinol 174: 238-245. doi:10.1016/j.ygcen.2011.09.004. PubMed: 21945118. [DOI] [PubMed] [Google Scholar]
- 22. Narayan EJ, Cockrem JF, Hero JM (2011) Urinary corticosterone metabolite responses to capture and captivity in the cane toad (Rhinella marina). Gen Comp Endocrinol 173: 371-377. doi:10.1016/j.ygcen.2011.06.015. PubMed: 21756910. [DOI] [PubMed] [Google Scholar]
- 23. Narayan E, Hero J-M (2011) Urinary corticosterone responses and haematological stress indicators in the endangered Fijian ground frog (Platymantis vitiana) during transportation and captivity. Aust J Zool 59: 79-85. doi:10.1071/ZO11030. [Google Scholar]
- 24. Clavero M, García-Berthou E (2005) Invasive species are a leading cause of animal extinctions. Tree 20: 110 PubMed: 16701353. [DOI] [PubMed] [Google Scholar]
- 25. Shine R (2010) The ecological impact of invasive cane toads (Bufo marinus) in Australia Q Rev Biol 85: 253-291. doi:10.1086/655116. PubMed: 20919631. [DOI] [PubMed] [Google Scholar]
- 26. Rodl T, Berger S, Romero LM, Wikelski M (2007) Tameness and stress physiology in a predator-naive island species confronted with novel predation threat. Proc R Soc Lond B 274: 577-582. doi:10.1098/rspb.2006.3755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Kiesecker JM, Blaustein AR (1997) Population differences in responses of Red-Legged frogs (Rana aurora) to introduced Bull frogs. Ecology 78: 1752-1760. doi:10.1890/0012-9658(1997)078[1752:PDIROR]2.0.CO;2. [Google Scholar]
- 28. Narayan E (2009) Biology of the Endangered Fijian Ground Frog (Platymantis vitiana): Breeding and Reproductive Endocrinology. Suva Univ South Pacific: 289. [Google Scholar]
- 29. Narayan E (2011) Biology and conservation of Fiji’s iconic native amphibians. FrogLog 99: 30-31. [Google Scholar]
- 30. Zug G, Watling D, Morrison C (2004) Platymantis vitianus . In: IUCN 2008. 2008 IUCN Red; List: of Threatened Species. Available: www iucnredlistorg [Google Scholar]
- 31. Thomas N, Morrison C, Winder L, Morley C (2011) Spatial distribution and habitat preferences of co-occurring vertebrate species: Case study of an endangered frog and an introduced toad in Fiji. Pac Conserv Biol 17: 68-77. [Google Scholar]
- 32. Narayan E, Hero J-M (2010) Platymantis vitiana; Froglets Morphology. Herpetol Rev 41: 478-479. [Google Scholar]
- 33. Narayan E, Christi K, Morley C (2008) Ecology and reproduction of the endangered Fijian Ground Frog Platymantis vitianus–Fiji Islands. South Pacific. J Nat Sciences 26: 28-32. [Google Scholar]
- 34. Ryan PA (1984) Fiji amphibia. Domodomo 11: 87-98. [Google Scholar]
- 35. Crossland MR, Brown GP, Anstis M, Shilton CM, Shine R (2008) Mass mortality of native anuran tadpoles in tropical Australia due to the invasive cane toad (Bufo marinus). Biol Conserv 141: 2387-2394. doi:10.1016/j.biocon.2008.07.005. [Google Scholar]
- 36. Phillips BL, Shine R (2006) An invasive species induces rapid adaptive change in a native predator: cane toads and black snakes in Australia. Proc R Soc Lond B 273: 1545-1550. doi:10.1098/rspb.2006.3479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Fraisse F, Cockrem JF (2006) Corticosterone and fear behaviour in white and brown caged laying hens. Br Poult Sci 47: 110-119. doi:10.1080/00071660600610534. PubMed: 16641020. [DOI] [PubMed] [Google Scholar]
- 38. Jones RB (1996) Fear and adaptability in poultry: insights, implications and imperatives. World's Poult Science J 52: 131-174. doi:10.1079/WPS19960013. [Google Scholar]
- 39. Lupo C, Lodi L, Paluffi G, Viti A (1991) Central and peripheral endocrine correlates of the immobility reaction in the toad Bufo bufo . Behav Processes 24: 1-7. doi:10.1016/0376-6357(91)90082-B. [DOI] [PubMed] [Google Scholar]
- 40. Phillott AD, Speare R, Hines HB, Skerratt LF, Meyer E, McDonald KR, Cashins SD, Mendez D, Berger L (2010) Minimising exposure of amphibians to pathogens during field studies. Dis Aquat Organ 92: 175-185. doi:10.3354/dao02162. PubMed: 21268979. [DOI] [PubMed] [Google Scholar]
- 41. Narayan E, Christi K, Morley C (2008) Ecology and reproduction of endangered Fijian ground frog Platymantis vitianus (Anura, Ranidae). South Pacific. J Nat Sciences 26: 28-32. [Google Scholar]
- 42. Narayan E, Christi K, Morley C, Trevenen P (2008) Sexual dimorphism in the cane toad Bufo marinus: a quantitative comparison of visual inspection methods for sexing individuals. Herpetol J 18: 63-65. [Google Scholar]
- 43. Narayan E, Hero J-M (2011) Urinary corticosterone responses and haematological stress indicators in the endangered Fijian ground frog (Platymantis vitiana) during transportation and captivity. Aust J Zool 59: 79-85. doi:10.1071/ZO11030. [Google Scholar]
- 44. Narayan E, Molinia F, Christi K, Morley C, Cockrem J (2010) Urinary corticosterone metabolite responses to capture, and annual patterns of urinary corticosterone in wild and captive endangered Fijian ground frogs (Platymantis vitiana). Aust J Zool 58: 189-197. doi:10.1071/ZO10010. [DOI] [PubMed] [Google Scholar]
- 45. Narayan EJ, Cockrem JF, Hero J-M (2011) Urinary corticosterone metabolite responses to capture and captivity in the cane toad (Rhinella marina). Gen Comp Endocrinol 173: 371-377. doi:10.1016/j.ygcen.2011.06.015. PubMed: 21756910. [DOI] [PubMed] [Google Scholar]
- 46. Narayan E (2013) Non-invasive reproductive and stress endocrinology in amphibian conservation physiology. Cons Physiol 1: 1-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Dabrowska B, Manikowski S (1982) Temperature and immobility reaction in Rana temporaria . Behav Processes 7: 179-182. doi:10.1016/0376-6357(82)90026-2. [DOI] [PubMed] [Google Scholar]
- 48. Taigen TL, Pough FH (1985) Metabolic correlates of anuran behavior. Am Zool 25: 987-997. [Google Scholar]
- 49. Thompson RKR, Foltin RW, Boylan RJ, Sweet A, Graves CA et al. (1981) Tonic immobility in Japanese quail can reduce the probability of sustained attack by cats. Anim Learn Behav 9: 145-149. doi:10.3758/BF03212037. [Google Scholar]
- 50. Jones RB, Beuving G, Blokhuis HJ (1988) Tonic immobility and heterophil/lymphocyte responses of the domestic fowl to corticosterone infusion. Physiol Behav 42: 249-253. doi:10.1016/0031-9384(88)90078-9. PubMed: 3406152. [DOI] [PubMed] [Google Scholar]
- 51. Young JE, Tracy CR, Christian KA, McArthur JL (2006) Rates of cutaneous evaporative water loss of native Fijian frogs. Copeia 1: 83-88. [Google Scholar]
- 52. Heinen JT, Hammond G (1997) Antipredator behaviors of newly metamorphosed green frogs (Rana clamitans) and leopard frogs (R. pipiens) in encounters with eastern garter snakes (Thamnophis s. sirtalis). Am Midl Nat 137: 136-144. doi:10.2307/2426762. [Google Scholar]
- 53. Narayan EJ, Hero J-M, Cockrem JF (2012) Inverse urinary corticosterone and testosterone responses to different durations of restraint in the cane toad (Rhinella marina). Gen Comp Endocrinol 179: 345-349. doi:10.1016/j.ygcen.2012.09.017. PubMed: 23036735. [DOI] [PubMed] [Google Scholar]
- 54. Narayan E, Molinia FC, Cockrem JF, Hero J-M (2012) Changes in urinary testosterone and corticosterone metabolites during short-term confinement with repeated handling in wild male cane toads (Rhinella marina). Aust J Zool 59: 264-269. [Google Scholar]
- 55. Narayan EJ, Hero J-M (2013) Repeatability of baseline corticosterone and acute stress responses to capture, and patterns of reproductive hormones in vitellogenic and non-vitellogenic female Fijian ground frog (Platymantis vitiana). J Exp Zool – A: Ecol Gen Physiol 9999A: 1–11. PubMed: 23847023. [DOI] [PubMed] [Google Scholar]
- 56. Cockrem JF, Candy EJ, Castille SA, Satterlee DG (2010) Plasma corticosterone responses to handling in Japanese quail selected for low or high plasma corticosterone responses to brief restraint. Br Poult Sci 51: 453-459. doi:10.1080/00071668.2010.503637. PubMed: 20680881. [DOI] [PubMed] [Google Scholar]
- 57. Archard GA, Earley RL, Hanninen AF, Braithwaite VA (2012) Correlated behaviour and stress physiology in fish exposed to different levels of predation pressure. Func Ecol 26: 637-645. doi:10.1111/j.1365-2435.2012.01968.x. [Google Scholar]
- 58. Berger S, Wikelski M, Romero LM, Kalko EKV, Rödl T (2007) Behavioral and physiological adjustments to new predators in an endemic island species, the Galapagos marine iguana. Horm Behav 52: 653-663. doi:10.1016/j.yhbeh.2007.08.004. PubMed: 17904141. [DOI] [PubMed] [Google Scholar]
- 59. Silverin B (1998) Behavioural and hormonal responses of the pied flycatcher to environmental stressors. Anim Behav 55: 1411-1420. doi:10.1006/anbe.1997.0717. PubMed: 9641986. [DOI] [PubMed] [Google Scholar]
- 60. Morrison C (2006) Brief overview of status and principal threats to amphibians in the Pacific. Pacific Ecol 13: 67-69. [Google Scholar]