

Abstract
Circadian clocks are endogenous approximately 24 h oscillators that temporally regulate many physiological and behavioural processes. In order to be beneficial for the organism, these clocks must be synchronized with the environmental cycles on a daily basis. Both light : dark and the concomitant daily temperature cycles (TCs) function as Zeitgeber (‘time giver’) and efficiently entrain circadian clocks. The temperature receptors mediating this synchronization have not been identified. Transient receptor potential (TRP) channels function as thermo-receptors in animals, and here we show that the Pyrexia (Pyx) TRP channel mediates temperature synchronization in Drosophila melanogaster. Pyx is expressed in peripheral sensory organs (chordotonal organs), which previously have been implicated in temperature synchronization. Flies deficient for Pyx function fail to synchronize their behaviour to TCs in the lower range (16–20°C), and this deficit can be partially rescued by introducing a wild-type copy of the pyx gene. Synchronization to higher TCs is not affected, demonstrating a specific role for Pyx at lower temperatures. In addition, pyx mutants speed up their clock after being exposed to TCs. Our results identify the first TRP channel involved in temperature synchronization of circadian clocks.
Keywords: circadian clocks, Drosophila melanogaster, transient receptor potential channels, pyrexia, temperature synchronization, chordotonal organs
1. Introduction
Circadian clocks allow organisms to adjust their physiology and behaviour to the daily changes of light and temperature caused by the rotation of the Earth around its own axis and the concomitant cyclical exposure to sunlight [1,2]. For example, in addition to escape responses from excessive light and heat, animals are able to anticipate environmental changes thanks to their circadian timers [3]. Circadian clocks therefore contribute to the overall fitness of organisms, despite not being essential for the survival of an individual [4]. The combined functions of several so-called ‘clock genes’ constitute the molecular bases of circadian clocks [1,2,5]. In the fruitfly Drosophila melanogaster, for example, the clock genes period (per) and timeless (tim) are rhythmically activated by the transcription factors clock (Clk) and cycle (Cyc). PER and TIM proteins eventually move back into the nucleus where they inhibit their own transcription after blocking the activity of Clk and Cyc, forming a negative feedback loop. After degradation of PER and TIM, this repression is released so that the cycle can start again. Various post-translational modifications ensure that the period of the cycle is adjusted to approximately 24 h [5].
Despite having evolved in a continuously changing environment, circadian clocks are self-sustained oscillators, meaning that molecular and behavioural rhythms continue in constant conditions [1]. Synchronization of these oscillations with the environmental cycles (generally referred to as ‘entrainment’) involves specific light and temperature inputs to the clock cells [2,5]. Circadian clocks are extremely sensitive to the daily changes of light and temperature, as for example, a 2–3°C difference between ‘night’ and ‘day’ is sufficient to entrain the Drosophila clock [3]. Surprisingly, external temperature cycles (TCs) are also able to reset the circadian clocks of endothermic animals [6,7], while clock-controlled body temperature rhythms serve as internal synchronization signal [8,9]. Despite the exquisite temperature sensitivity with regard to entrainment, free-running properties of circadian clocks are temperature-compensated, which ensures that clock speed does not change at different temperatures. In other words, the period length of a rhythm observed under constant conditions varies little in relation to the ambient (constant) temperature [1]. It is not understood how this apparent conundrum (i.e. sensitivity versus compensation) is resolved by the circadian system.
The temperature receptor(s) for synchronization of the Drosophila clock have not been identified. It is known that peripheral (non-neuronal) clock cells can be synchronized in a tissue-autonomous manner, while the central clock neurons in the brain receive temperature signals from peripheral sense organs [10–12]. Temperature-sensitive transient receptor potential (TRP) channels are activated at defined temperature values [13,14]. Because temperature entrainment in Drosophila can be achieved with TCs covering their whole physiological temperature range (15–30°C), it is possible that different TRP channels are involved in temperature entrainment at different temperature ranges [10,15,16]. The TRP channel encoded by the pyrexia (pyx) gene is directly activated by temperatures above 38°C and protects flies from high temperature stress [17]. Surprisingly, pyx is also responsible for the response of temperature-sensitive anterior cell (AC) brain neurons, which regulate the temperature preference behaviour of adult fruitflies [18,19]. Pyx mediates the AC neuron response between 27°C and 33°C, suggesting that the Pyx channel may also be activated indirectly as has been shown for dTRPA1 [19,20]. Here, we investigated the possibility that Pyx contributes to temperature entrainment of the fly clock. We find that Pyx mediates temperature entrainment at low temperature intervals (16–20°C) but not at higher temperatures (21–29°C). We therefore define a low temperature range for this TRP channel, and Pyx is the first member of this protein family shown to be involved in thermal circadian clock synchronization.
2. Results
To monitor the ability of flies to synchronize to TCs, we recorded locomotor behaviour under the following conditions: initially flies were synchronized to a 12 L : 12 D cycle at constant temperature. This was followed by a TC (TC1; lasting 5 days) that was phase-delayed by 6 h compared with the initial light : dark (LD), and a second TC2 (5 days) that was 6 h delayed compared with TC1. The temperatures of TC1 and TC2 in each experiment cycled between 16 : 20°C, 20 : 25°C or 25 : 29°C. TCs were either accompanied by constant illumination (LL) or constant darkness (DD), and in the DD experiments, fly activity was recorded at the constant lower temperature for several days after the last day of TC2.
(a). Pyrexia is required for synchronization to low (16 : 20°C) temperature cycles in constant light
Wild-type Canton S control flies synchronize their activity peak during 16 : 20°C TC in LL to the second part of the warm phase in TC1 (figure 1a). During TC2, they delay their activity onsets, so that at the end of TC2, the main activity peak occurs during the middle to second half of the warm phase. In addition, during both TCs wild-type flies react with a sharp increase of activity immediately after the transition to the lower temperature, and during the remaining periods of the warm and cold phase, they exhibit little activity (figure 1a). A very similar behavioural pattern has recently been described for 16 : 25°C TC using the same double delay conditions [15].
Figure 1.

Pyrexia is required for synchronization to 16 : 20°C TCs in constant light. Locomotor behaviour of (a) Canton S, (b) pyx3, (c) pyx3/pyxDf9 and (d) pyxGe pyx3 male flies was analysed in LD, followed by two 16 : 20°C TCs in constant light (LL) each of which was delayed by 6 h compared with the previous regime. Top graphs show double-plotted average actograms, depicting behavioural activity throughout the experiment. Below, for the LD and TC parts, the last 3 days were averaged and plotted as histograms. On the y-axis, average activity (beam crossings/30 min) is indicated, while x-axis represents the 24 h day, whereby each column represents 30 min. White and grey bars (histograms) or areas (actograms) indicate light and dark periods, respectively. Blue areas indicate cold, orange areas warm periods. Absolute temperatures are indicated to the left. The number of animals analysed is indicated in each histogram (average of several independent experiments). Average actograms are from one experiment in which 8–16 flies were tested for each genotype. (e) Plotting of the EI for TC1 and TC2 with error bars indicating s.e.m. Statistical significance was assessed by one-way ANOVA followed by Fisher's LSD post hoc test (*p-values < 0.05; ****p-values < 0.0001).
The pyx locus encodes two Pyx proteins (Pyx-PA and Pyx-PB), which presumably form heterodimers in order to build a functional channel [17]. If true, any mutant allele affecting either of the two proteins should interfere with channel function. We therefore tested four different alleles alone or in combination for their potential defects in temperature entrainment: pyx2 and pyx3 are P-element insertions that change the ratio of the pyx-PA and pyx-PB transcripts (pyx2) or eliminate them completely (pyx3), whereas the pyxDf4 (produce only pyx-PA) and pyxDf9(produce only pyx-PB) alleles are imprecise P-element excisions generated from the pyx2 allele [17]. All pyx mutant flies synchronize normally to LD but not to shifted 16 : 20°C TC in LL (figure 1b,c; electronic supplementary material S1). While pyx mutants do show a sharp activity increase after the temperature drop, they lack a synchronized activity peak in the warm phase, and their activity levels are evenly distributed over the cold and warm phase. Next, we tried to rescue the pyx3 mutation by introducing a pyx-encoding transgene (pyxGe pyx3) that rescues both the pyx3 heat-induced paralysis, as well as the AC neuron response at 27°C [17,19]. Although pyx3 rescue flies were still active during the cold phase, clear activity peaks are present towards the middle of the warm phase, indicating at least partial rescue of the temperature entrainment phenotype (figure 1d).
In order to quantify if and to which degree a genotype entrains to TC, we calculated an ‘entrainment index’ (EI) as previously described (briefly, we determined the ratio between the activity of Canton S flies occurring during the obvious peak times (from ZT5.5 to ZT11 for TC1 and TC2; figure 1a) and the total activity during the 12 h of the warm phase [15]). This quantification confirmed our interpretations with highest EI values for Canton S, lowest for the pyx mutants and significantly increased EI values for the pyxGe rescue flies compared with the mutants (figure 1e).
(b). Pyrexia is required for synchronization to low (16 : 20°C) temperature cycles in constant darkness
In contrast to LL, wild-type flies behave rhythmically in constant darkness (DD) in the absence of any Zeitgebers. We therefore performed the same TC experiments in DD, and released the flies into constant temperature and DD at the end of the TC entrainment to allow comparison of the activity phases of control flies with those of the pyx mutants. If flies entrain to TC, they should initiate their free run with a phase of activity similar to that of the last TC they were exposed to [16]. Compared with the LL conditions, Canton S flies show an earlier phased activity peak in TC1 and especially in TC2, where peak activity occurs already 2–3 h after transition to the warm phase (figure 2a; cf. [15]). The similar phasing of the activity peaks at the beginning of the constant conditions and at the end of TC2 as observed here for Canton S flies indicates synchronization to the TC (figure 2a).
Figure 2.

Pyrexia is required for synchronization to 16 : 20°C TCs in constant darkness. Locomotor behaviour of (a) Canton S, (b) pyx3, (c) pyx3/pyxDf9 and (d) pyxGe pyx3 male flies was analysed in LD, followed by two 16 : 20°C TCs in constant darkness (DD) as described in the legend to figure 1. After TC2, flies were released to constant 16°C in DD and the first 3 days were averaged to generate activity histograms. (e,f) Plotting of the EI for TC2 and free run with error bars indicating s.e.m. Statistical significance was assessed by one-way ANOVA followed by Fisher's LSD post hoc test (*p-values < 0.05; **p-values < 0.01; n.s., not significant).
The pyx mutants showed again defects in synchronizing their locomotor behaviour to TC in DD. Mutant flies showed prominent reactions (activity increases) after the temperature step up, but otherwise keep their main activity peak at the same position as it was at the end of the LD entrainment, irrespective of the TC (figure 2b,c; electronic supplementary material S2). During TC1, all flies show the same activity profile, because the activity responses to the temperature increase cannot be distinguished from a synchronized behavioural peak. We therefore focused our analysis on TC2 and the subsequent free run in order to determine entrainment. During TC2, the mutant flies' main activity falls into the middle of the cold phase, because they fail to shift their main activity peak to the morning phase as control flies do. As in LL, the pyxGe pyx3 flies showed intermediate behaviour and resulted in a partial rescue of pyx3 behaviour. This is most obvious when comparing the phase of the onset of the free-running activity between mutant, rescue and control flies (lower panels in figure 2a–d). We calculated EI values for TC2 and the final free-run portion of the experiment, which showed significant differences between the pyx mutant and the pyxGe pyx3 rescue flies (figure 2e,f). Moreover, all pyx alleles and combinations differ significantly from Canton S in TC2, except for one, also in the free run (figure 2; electronic supplementary material S2). Our results show that the Pyx channel is required for synchronization to 16 : 20°C TC, and that both Pyx-PA and Pyx-PB are equally important (see electronic supplementary material, figure S2).
As independent demonstration that pyx mutants interfere with temperature entrainment, we also performed experiments where we directly compared the behaviour of flies that were exposed to shifted TC in DD with that of flies that were released into constant temperature after the first TC (see electronic supplementary material, figure S3). In Canton S and pyxGe pyx3 rescue flies, the activity peak phases during the last 4 days of the experiment occurred late or early in the subjective day when compared between the shifted and constant temperature control flies, demonstrating that the TC shifts stably resynchronized the flies' behaviour. By contrast, although pyx mutants did show activity increases in response to the higher temperature, their activity peak phases in the final free-run part were not aligned with those occurring at the previous temperature transitions but appeared to be in phase with the free-running component observable throughout the shifted TC regimes (see electronic supplementary material, figure S3). These results confirm that pyx mutants exhibit altered behavioural patterns during and after exposure to 16 : 20°C TC, indicating impaired temperature input to the circadian clock.
(c). Pyrexia is not involved in synchronization to higher temperature intervals
Based on previous reports [17,19], we expected to see pyx effects on temperature synchronization at higher temperatures. Surprisingly, both pyx2 and pyx3 showed normal synchronization to 20 : 25°C and to 25 : 29°C TCs in DD and LL (figure 3; electronic supplementary material S4; data not shown). These results suggest that Pyx-mediated activation of the AC neurons at 27°C is not required for temperature synchronization in the same temperature range and that other temperature receptors mediate synchronization to higher temperature intervals.
Figure 3.

Pyrexia is not required for synchronization to 25 : 29°C TCs. Locomotor behaviour of (a) Canton S, (b) pyx3, (c) pyx3/pyxDf9 and (d) pyxGe pyx3 male flies was analysed in LD, followed by two 25 : 29°C TC in DD as described in the legends to figures 1 and 2. Low and high temperatures are indicated in orange and red, respectively. After TC2, flies were released to constant 25°C in DD. Note that all genotypes show synchronized activity peaks during this temperature interval.
(d). Pyrexia is not required for temperature compensation
As in other organisms, the circadian clock of Drosophila is well compensated to non-rhythmic changes in ambient temperature, meaning that at different constant temperatures (i.e. in the absence of TC), the free-running period is very similar [21]. All TC synchronization mutants known so far do not affect temperature compensation [10], and we show here that the same is true for pyx mutations (figure 4a and table 1). Inspection of the free-running behaviour after TC entrainment (figures 2 and 3; electronic supplementary materials S3 and S4) indicated that impaired pyx function leads to shorter periods in constant conditions. Interestingly, with the exception of Canton S, free-running periods (in DD at constant 16°C and 20°C) of all pyx alleles tested were significantly shorter (between 0.5 and 1.3 h) after TC when compared with flies of the same genotype that were entrained in LD and released to 18°C (figure 4b and table 1). This is particularly obvious when flies were released into constant temperature after only one short TC (see electronic supplementary material, figure S3). Although these results may suggest a role for pyx in preventing short period rhythms after TC entrainment, the fact that pyxGe pyx3 rescue flies also show TC-dependent period shortening (figure 4b and table 1) argues against such a role (see Discussion).
Figure 4.
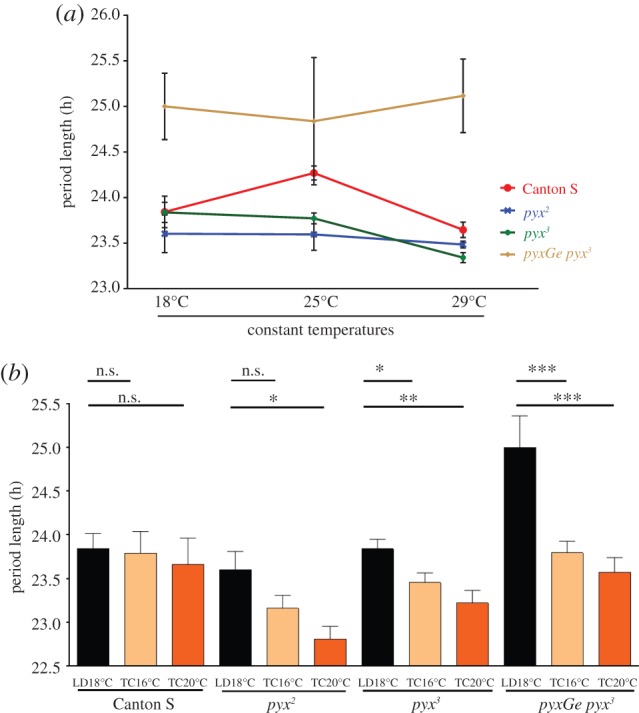
Pyrexia mutants do not affect temperature compensation but potentially shorten period length after temperature entrainment. (a) Free-running periods of male flies of the indicated genotypes at constant temperatures of 18°C, 25°C and 29°C in DD, after initial LD entrainment at the same temperature. All genotypes show robust temperature compensation. (b) Free-running periods in DD after LD entrainment and release to constant 18°C (first column) compared with periods after 16 : 20°C (second column) and 20 : 25°C (third column) TC entrainment at constant temperatures of 16°C and 20°C, respectively. Note that all genotypes containing either the pyx2 or pyx3 allele show significant period shortening, including the pyxGe pyx3 rescue flies.
Table 1.
Rhythmicity and period length in DD at constant temperatures and after TC entrainment. Flies of the indicated genotypes were entrained to 12 L :12 D or TC cycles, as indicated in the ‘entrainment condition’ column, followed by exposure to constant darkness and temperature, indicated in the ‘free run’ column. Flies in the last group (TC 16–20°C and free-run at 20°C) were entrained to one TC only and monitored in free run for approximately two weeks (see the electronic supplementary material, figure S3, right panels). Period length, s.e.m. and ryhthmicity statistics (RS) were calculated as described in Materials and methods.
| genotype | entrainment condition | free run | n | rhythmic (%) | period length (h) | s.e.m. | RS |
|---|---|---|---|---|---|---|---|
| Canton S | LD | 18°C | 66 | 54.5 | 23.8 | 0.17 | 2.2 |
| pyx2 | LD | 18°C | 61 | 67.2 | 23.6 | 0.21 | 2.4 |
| pyx3 | LD | 18°C | 56 | 80.4 | 23.8 | 0.11 | 3.3 |
| pyxGe pyx3 | LD | 18°C | 46 | 60.9 | 25.0 | 0.36 | 2.9 |
| Canton S | LD | 25°C | 41 | 70.7 | 24.3 | 0.08 | 2.9 |
| pyx2 | LD | 25°C | 31 | 74.2 | 23.6 | 0.17 | 2.9 |
| pyx3 | LD | 25°C | 22 | 95.5 | 23.8 | 0.06 | 3.3 |
| pyxGe pyx3 | LD | 25°C | 13 | 61.5 | 24.8 | 0.70 | 2.0 |
| Canton S | LD | 29°C | 46 | 76.1 | 23.6 | 0.08 | 2.9 |
| pyx2 | LD | 29°C | 32 | 96.9 | 23.5 | 0.04 | 4.0 |
| pyx3 | LD | 29°C | 38 | 86.8 | 23.3 | 0.06 | 3.2 |
| pyxGe pyx3 | LD | 29°C | 29 | 62.1 | 25.1 | 0.40 | 2.3 |
| Canton S | TC 16–20°C | 16°C | 68 | 47.1 | 23.8 | 0.25 | 2.2 |
| pyx2 | TC 16–20°C | 16°C | 69 | 60.9 | 23.2 | 0.15 | 2.4 |
| pyx3 | TC 16–20°C | 16°C | 96 | 70.8 | 23.5 | 0.11 | 2.6 |
| pyxGe pyx3 | TC 16–20°C | 16°C | 62 | 66.1 | 23.8 | 0.13 | 2.7 |
| Canton S | TC 20–25°C | 20°C | 32 | 68.8 | 23.7 | 0.30 | 4.2 |
| pyx2 | TC 20–25°C | 20°C | 32 | 68.8 | 22.8 | 0.15 | 3.5 |
| pyx3 | TC 20–25°C | 20°C | 44 | 75.0 | 23.2 | 0.14 | 2.8 |
| pyxGe pyx3 | TC 20–25°C | 20°C | 30 | 80.0 | 23.6 | 0.17 | 3.0 |
| Canton S | TC 16–20°C | 20°C | 25 | 100 | 23.7 | 0.11 | 4.1 |
| pyx3 | TC 16–20°C | 20°C | 18 | 88.9 | 23.3 | 0.19 | 4.5 |
| pyx3/pyxDf9 | TC 16–20°C | 20°C | 20 | 100 | 23.4 | 0.15 | 5.4 |
| pyxGe pyx3 | TC 16–20°C | 20°C | 11 | 100 | 23.8 | 0.14 | 4.5 |
(e). Pyrexia is expressed in attachment cells of femur chordotonal organs
Because chordotonal organs (cho) have been implicated in temperature entrainment [12], we investigated whether pyx is also expressed in these sensory structures. Previous studies had shown pyx expression in the antennal cho, most probably in the attachment cells, although neuronal expression could not be excluded [19,22]. To distinguish more clearly between potential neuronal and/or attachment cells expression, we crossed pyx-Gal4 flies to flies carrying a nuclear RFP and mCD8-GFP, and inspected expression in the larger femur cho. We observed prominent expression in the attachment cells but no expression within cho neurons (figure 5a–c).
Figure 5.
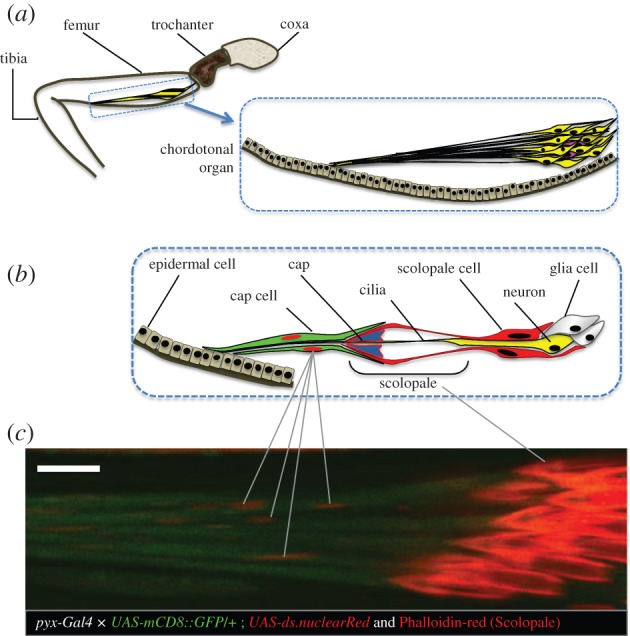
Pyrexia expression in the cho cap cells. (a) Schematic of the general anatomy of the fly's leg with cho neurons indicated in yellow. (b) Detailed representation of the cho showing only one neuron and its associated cells, for simplification purpose. (c) Confocal microscopy of femoral cho of pyx-Gal4 flies expressing mCD8 :: GFP and Nuclear Red. Membrane of Cap cells are shown in green and their nuclei in red. Phalloidin Red was used to stain the scolopales. Scale bar is 10 µm.
3. Discussion
(a). Role of Pyx in temperature entrainment
Temperature entrainment relevant for the synchronization of behavioural rhythms of fruitflies involves input from internal mechanosensory structures called cho [12]. Although largely known for their function as mechanoreceptors, cho have also been linked to temperature sensation in Drosophila [23–25]. nocte mutants exhibit cho structural defects in the dendritic cap of the scolopale cell, which connects the cilia of the cho neuron with the cap cell (also called attachment cell; figure 5) [12]. Consistent with a previous study [22], we find that pyx is expressed in cho cap cells, which connect the cho to the body wall (figure 5). In conjunction with the lack of pyx expression in the CNS [22], we interpret the temperature synchronization defects of pyx mutants as evidence for the involvement of the cap cells in the process of temperature sensation.
We previously showed that different groups of clock neurons in the fly brain mediate synchronization to low and high temperature intervals, with PER expression in the more ventrally located LNv clock neurons being sufficient for synchronization to 16 : 25°C TC [15]. Our finding that pyx mutants specifically affect entrainment to lower TC therefore suggests Pyx's involvement in a thermal input pathway to the LNv and, at the same time, that other channels and receptors mediate synchronization at higher temperatures involving different clock neurons.
AC neurons express the thermo-sensitive channel dTRPA1 and regulate Drosophila temperature preference behaviour [18]. The dTRPA1 channel is activated at temperatures above 25°C, which is the preferred temperature of wild-type flies, and dTRPA1 mutants prefer temperatures above 25°C [18,26]. In addition, AC neurons respond to slightly higher temperatures of 27°C, and this response is mediated by Pyx [19]. The AC neurons do not express Pyx, but synaptic connections from unidentified Pyx-expressing neurons in the second segment of the antennae to the AC neurons have been visualized [19]. Interestingly, pyx loss-of-function mutant flies do not show any defects in temperature preference behaviour, suggesting an additional role for the AC neurons and pyx [17,19]. In fact, Tang et al. [19] proposed a role for both Pyx and the AC neurons in temperature entrainment. We were therefore surprised that pyx mutant showed normal entrainment in the 25–29°C interval and that we identified a third temperature range for this channel (16–20°C). Temperature preference behaviour also has a clock-controlled circadian component (temperature preference rhythm, or TPR), whereby the preferred temperature gradually increases during the day, reaching almost 27°C in the evening [27]. It will be interesting to see whether Pyx is required for TPR.
Flies from which the antennae have been completely removed are able to synchronize to TCs [12]. Although this does not rule out a contribution of antennal thermo-sensors, it suggests that sensory receptors in the body signal temperature information to the clock neurons in the brain, and/or that thermo-receptors for clock resetting are expressed within the brain. We did not find any effects of dtrpA1 mutants on temperature entrainment (data not shown), suggesting the involvement of other TRP channels or receptors responsible for synchronization to temperatures cycles above 20°C. Fly Rhodopsins contribute to temperature preference behaviour in larvae, and it will be interesting to see whether they are involved in temperature entrainment [28].
(b). Indirect activation of Pyx
Based on our mutant analysis, both Pyx isoforms are required for temperature entrainment in agreement with the idea that Pyx-PA and Pyx-PB subunits form functional heterodimers for mediating tolerance to high temperatures [17]. By contrast, the ankyrin-repeat containing Pyx-PA subunit is sufficient for mediating normal negative geotaxis behaviour [22], suggesting different mechanisms for thermal and mechanical Pyx activation. The high direct activation temperature of Pyx (greater than 38°C) suggests that indirect activation accounts for Pyx function in temperature entrainment. Direct (greater than 25°C) and indirect (19–24°C) modes of activation have also been shown for dTRPA1, whereby the latter involves phospholipase C-β (PLC-β) signalling [18,20,26]. Interestingly, PLC-β signalling has also been implicated in temperature entrainment of peripheral and central clocks [10,11,29]. It is therefore likely that the actual temperature receptor(s) mediating clock synchronization remain to be identified.
(c). Pyx influence on clock speed
We were surprised to see significant reductions of period lengths in pyx mutant genotypes after temperature entrainment compared with LD entrainment (figure 4b and table 1). Effects of entrainment conditions on the subsequent free-running period are known as ‘aftereffects’, but to our knowledge, they have not been described to occur after 12 L : 12 D TCs. For example, aftereffects occur when using long photoperiods (e.g. 18 L : 6 D) for entrainment, resulting in short free-running periods [30]. Also, phase-delay paradigms of LD cycles similar to the ones applied here with TCs cause an increase in free-running period length [30], but we observed period shortenings. Moreover, wild-type flies do not show a significant change of free-running period after temperature entrainment (figure 4b and table 1; electronic supplementary material S3). Our results indicate the possibility that Pyx may help buffering the clock against TC-induced aftereffects. Even though we could not rescue the period shortenings with our rescue construct, it seems rather unlikely that another mutation in the background of our pyx stocks exhibits a temperature-dependent (but Pyx-independent) phenotype, but we cannot exclude this possibility. Because the pyxGe pyx3 flies did not fully restore the AC neuron response [19], we favour the idea that suboptimal function of our rescue construct (also indicated by the partial rescue of the pyx temperature entrainment phenotypes) is not sufficient for protecting the pyx mutants from the TC-induced aftereffects.
In summary, we demonstrate that the Pyx TRP channel regulates temperature synchronization in Drosophila to lower temperature intervals. Pyx activation for this function must be indirect and requires the function of as yet unidentified receptors. In agreement with previous studies, we find pyx expression in the cap cells of cho, which therefore seem to be part of at least one of the input routes to a subset of the circadian clock neurons in the fly brain. Because pyx mutants synchronize normally to higher temperature intervals, other receptors must exist for mediating entrainment to higher temperatures.
4. Material and methods
(a). Fly stocks
Wild-type Canton S was used as control. The pyx2, pyx3, pyxDf4, pyxDf9 and pyxGe pyx3 rescue flies have previously been described [17,19,22]. Only male flies were used in the current study. pyx-Gal4 contains 1 kb genomic DNA sequence upstream of the pyx translation start fused to Gal4 coding sequences, and was generated and described in [22]. The UAS-dsRED, UAS-mCD8 reporter has been described in [12].
(b). Analysis of activity behaviour
Male flies were initially synchronized to three LD cycles at constant temperatures as indicated in the figures. After the last dark period of the final day in LD, lights were turned on (LL) or left off (DD) for the rest of the experiment. For the 16 : 20°C TC, temperature remained at 20°C (LD entrainment temperature) for 18 h, before being dropped to 16°C (effectively inducing a 6 h delay compared with the previous LD cycle). For the 20 : 25°C and 25 : 29°C TCs, temperature remained at 20°C and 25°C (respective LD entrainment temperature) for 6 h, before raising to 25°C or 29°C, also inducing a 6 h delay compared with the initial LD cycle. The initial TC1 continued for 5 days and was then shifted by delaying the temperature increase by 6 h, and flies were tested for resynchronization to this shifted TC2 for another 5 days. For DD experiments, behaviour was also monitored after the second TC in DD and constant temperature (as indicated in the figures). For the experiment in the electronic supplementary material, figure S3, flies of the same genotype were either exposed to three 16 : 20°C TCs as indicated, followed by release into constant 20°C (shift group) or released to constant 20°C after TC1 (constant temperature control group). Daily average histograms were generated in Excel, and actograms were plotted using the fly toolbox and MATLab software [31]. For the daily average histograms showing behaviour during TC only, the last 3 days of each TC were averaged in order to avoid inclusion of transient behaviour. To quantify synchronization during the 16 : 20°C TC, we calculated an EI [15]. For the LL experiment (figure 1), the EI indicates the ratio of total activity occurring during a 6 h window of TC1 and TC2 (ZT5.5 to ZT11, activity peak of Canton S) over the activity during the entire warm phase; for the DD experiment (figure 2), the EI indicates the ratio of activity within the ZT(CT) 20.5 to ZT(CT)2 window during TC2 (figure 2e) and the first 3 days of free run (figure 2f) over the entire 24 h (because here the main activity peak spanned the transition from warm to cold). Values closer to 1 show that most of the activity occurred in the specified window suggesting entrainment. To avoid interference of activity increases at the two temperature transitions (startle response) with the EI calculation, we also applied a simple smoothing filter for the four activity bins during the 2 h following each temperature transition (activity was collected in 30 min bins): if bin activity was more than 1.5 times that of the last bin before the transition, it was replaced by the average activity values of the two bins before the transition and the two bins following the bin to be replaced. In this way, only data from genotypes and conditions that did produce a startle response were filtered. Note that the histograms shown in the figures contain unfiltered data and that the filter was only applied to calculate EI. We first calculated an EI for each one of the last 3 days of a TC shift (or the first 3 days of the free run). A total of six EIs were obtained for each genotype and condition (TC1 and TC2 in LL; TC2 and free run in DD), with which the average EI, standard error of the mean (s.e.m.) and statistics were calculated. We tested and confirmed for the normality of the distribution and performed one-way analysis of variance (ANOVA) followed by Fisher's least significant difference (LSD) post hoc test to analyse differences between groups. Statistical analysis was performed in GraphPad Prism 6. Free-running periods for temperature compensation experiments and for DD TC experiments (table 1) were calculated using the autocorrelation function. In this study, all flies with a rhythmicity statistics (RS) value of more than 1.5 were considered as rhythmic (see [31] for how this cut-off was determined).
(c). Visualization of pyx expression in the femur chordotonal organ
Transgenic flies expressing mCD8 :: GFP and nuclear RFP under the control of pyx-Gal4 were fixed in 4% paraformaldehyde for 2 h, then washed three times for 10 min in 1% Triton X-100 (PBS-T) at room temperature. Dissection of the legs was performed in the same washing solution, then legs were cut at the proximal end of the femur. Samples were then incubated with Phalloidin-TRITC (1 : 1000, Sigma) in 1% PBS-T overnight at 4°C, then washed three times for 10 min in 1% PBS-T at room temperature, mounted in Vectashield medium (Vector laboratories, Burlingame, CA) and examined under a Leica SP5 confocal microscope (Leica, Germany). In control assessments, the parental line carrying UAS-mCD8::GFP and UAS-nuclearRed was used to verify the absence of Gal4-independent signals (data not shown).
Note added in proof
While this manuscript was under review, the Montell group published a paper implicating the TRPA1 channel in temperature synchronization of the Drosophila circadian clock: Lee Y, Montell C. (2012). J Neurosci 33(16):6716–6725.
Acknowledgements
We thank Joanna Szular for help in performing the behavioural assays and Jaesob Kim for pyrexia flies. We also thank Fumika Hamada, Patrick Emery and members of our laboratory for discussions.
Funding statement
This work was supported by BBSRC grant no. BB/H001204/1 given to R.S. and the EU: EUCLOCK, an Integrated Project of FP6.
References
- 1.Dunlap JC, Loros JJ, DeCoursey PJ. 2004. Chronobiolgy: biological timekeeping. Sunderland, MA: Sinauer Associates, Inc [Google Scholar]
- 2.Peschel N, Helfrich-Förster C. 2011. Setting the clock: by nature: circadian rhythm in the fruitfly Drosophila melanogaster. FEBS Lett. 585, 1435–1442 (doi:10.1016/j.febslet.2011.02.028) [DOI] [PubMed] [Google Scholar]
- 3.Wheeler DA, Hamblen-Coyle MJ, Dushay MS, Hall JC. 1993. Behavior in light–dark cycles of Drosophila mutants that are arrhythmic, blind, or both. J. Biol. Rhythms 8, 67–94 (doi:10.1177/074873049300800106) [DOI] [PubMed] [Google Scholar]
- 4.Woelfle MA, Ouyang Y, Phanvijhitsiri K, Johnson CH. 2004. The adaptive value of circadian clocks: an experimental assessment in cyanobacteria. Curr. Biol. 14, 1481–1486 (doi:10.1016/j.cub.2004.08.023) [DOI] [PubMed] [Google Scholar]
- 5.Hardin PE. 2011. Molecular genetic analysis of circadian timekeeping in Drosophila. Adv. Genet. 74, 141–173 (doi:10.1016/B978-0-12-387690-4.00005-2) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Refinetti R. 2010. Entrainment of circadian rhythm by ambient temperature cycles in mice. J. Biol. Rhythms 25, 247–256 (doi:10.1177/0748730410372074) [DOI] [PubMed] [Google Scholar]
- 7.El Allali K, Achaaban MR, Bothorel B, Piro M, Bouaouda H, El Allouchi M, Ouassat M, Malan A, Pevet P. 2013. Entrainment of the circadian clock by daily ambient temperature cycles in the camel (Camelus dromedarius). Am. J. Physiol. Regul. Integr. Comp. Physiol. 304, R1044–R1052 (doi:10.1152/ajpregu.00466.2012) [DOI] [PubMed] [Google Scholar]
- 8.Buhr ED, Yoo SH, Takahashi JS. 2010. Temperature as a universal resetting cue for mammalian circadian oscillators. Science 330, 379–385 (doi:10.1126/science.1195262) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Brown SA, Zumbrunn G, Fleury-Olela F, Preitner N, Schibler U. 2002. Rhythms of mammalian body temperature can sustain peripheral circadian clocks. Curr. Biol. 12, 1574–1583 (doi:10.1016/S0960-9822(02)01145-4) [DOI] [PubMed] [Google Scholar]
- 10.Glaser FT, Stanewsky R. 2005. Temperature synchronization of the Drosophila circadian clock. Curr. Biol. 15, 1352–1363 (doi:10.1016/j.cub.2005.06.056) [DOI] [PubMed] [Google Scholar]
- 11.Glaser FT, Stanewsky R. 2007. Synchronization of the Drosophila circadian clock by temperature cycles. Cold Spring Harb. Symp. Quant. Biol. 72, 233–242 (doi:10.1101/sqb.2007.72.046) [DOI] [PubMed] [Google Scholar]
- 12.Sehadova H, Glaser FT, Gentile C, Simoni A, Giesecke A, Albert JT, Stanewsky R. 2009. Temperature entrainment of Drosophila’s circadian clock involves the gene nocte and signaling from peripheral sensory tissues to the brain. Neuron 64, 251–266 (doi:10.1016/j.neuron.2009.08.026) [DOI] [PubMed] [Google Scholar]
- 13.Garrity PA, Goodman MB, Samuel AD, Sengupta P. 2010. Running hot and cold: behavioral strategies, neural circuits, and the molecular machinery for thermotaxis in C. elegans and Drosophila. Genes Dev. 24, 2365–2382 (doi:10.1101/gad.1953710) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Dhaka A, Viswanath V, Patapoutian A. 2006. Trp ion channels and temperature sensation. Annu. Rev. Neurosci. 29, 135–161 (doi:10.1146/annurev.neuro.29.051605.112958). [DOI] [PubMed] [Google Scholar]
- 15.Gentile C, Sehadova H, Simoni A, Chen C, Stanewsky R. 2013. Cryptochrome antagonizes synchronization of Drosophila's circadian clock to temperature cycles. Curr. Biol. 23, 185–195 (doi:10.1016/j.cub.2012.12.023) [DOI] [PubMed] [Google Scholar]
- 16.Busza A, Murad A, Emery P. 2007. Interactions between circadian neurons control temperature synchronization of Drosophila behavior. J. Neurosci. 27, 10 722–10 733 (doi:10.1523/JNEUROSCI.2479-07.2007) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Lee Y, et al. 2005. Pyrexia is a new thermal transient receptor potential channel endowing tolerance to high temperatures in Drosophila melanogaster. Nat. Genet. 37, 305–310 (doi:10.1038/ng1513) [DOI] [PubMed] [Google Scholar]
- 18.Hamada FN, Rosenzweig M, Kang K, Pulver SR, Ghezzi A, Jegla TJ, Garrity PA. 2008. An internal thermal sensor controlling temperature preference in Drosophila. Nature 454, 217–220 (doi:10.1038/nature07001). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Tang X, Platt MD, Lagnese CM, Leslie JR, Hamada FN. 2013. Temperature integration at the AC thermosensory neurons in Drosophila. J. Neurosci. 33, 894–901 (doi:10.1523/JNEUROSCI.1894-12.2013) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kwon Y, Shim HS, Wang X, Montell C. 2008. Control of thermotactic behavior via coupling of a TRP channel to a phospholipase C signaling cascade. Nat. Neurosci. 11, 871–873 (doi:10.1038/nn.2170) [DOI] [PubMed] [Google Scholar]
- 21.Konopka RJ, Pittendrigh C, Orr D. 1989. Reciprocal behaviour associated with altered homeostasis and photosensitivity of Drosophila clock mutants. J. Neurogenet. 6, 1–10 (doi:10.3109/01677068909107096) [DOI] [PubMed] [Google Scholar]
- 22.Sun Y, Liu L, Ben-Shahar Y, Jacobs JS, Eberl DF, Welsh MJ. 2009. TRPA channels distinguish gravity sensing from hearing in Johnston's organ. Proc. Natl Acad. Sci. USA 106, 13 606–13 611 (doi:10.1073/pnas.0906377106) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kwon Y, Shen WL, Shim HS, Montell C. 2010. Fine thermotactic discrimination between the optimal and slightly cooler temperatures via a TRPV channel in chordotonal neurons. J. Neurosci. 30, 10 465–10 471 (doi:10.1523/JNEUROSCI.1631-10.2010) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kernan MJ. 2007. Mechanotransduction and auditory transduction in Drosophila. Pflugers Arch. 454, 703–720 (doi:10.1007/s00424-007-0263-x) [DOI] [PubMed] [Google Scholar]
- 25.Liu L, Yermolaieva O, Johnson WA, Abboud FM, Welsh MJ. 2003. Identification and function of thermosensory neurons in Drosophila larvae. Nat. Neurosci. 6, 267–273 (doi:10.1038/nn1009) [DOI] [PubMed] [Google Scholar]
- 26.Viswanath V, Story GM, Peier AM, Petrus MJ, Lee VM, Hwang SW, Patapoutian A, Jegla T. 2003. Opposite thermosensor in fruitfly and mouse. Nature 423, 822–823 (doi:10.1038/423822a) [DOI] [PubMed] [Google Scholar]
- 27.Kaneko H, Head LM, Ling J, Tang X, Liu Y, Hardin PE, Emery P, Hamada FN. 2012. Circadian rhythm of temperature preference and its neural control in Drosophila. Curr. Biol. 22, 1851–1857 (doi:10.1016/j.cub.2012.08.006) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Shen WL, Kwon Y, Adegbola AA, Luo J, Chess A, Montell C. 2011. Function of rhodopsin in temperature discrimination in Drosophila. Science 331, 1333–1336 (doi:10.1126/science.1198904) [DOI] [PubMed] [Google Scholar]
- 29.Ito C, Goto SG, Tomioka K, Numata H. 2011. Temperature entrainment of the circadian cuticle deposition rhythm in Drosophila melanogaster. J. Biol. Rhythms 26, 14–23 (doi:10.1177/0748730410391640) [DOI] [PubMed] [Google Scholar]
- 30.Moore-Ede MC, Sulzman FM, Fuller CA. 1982. The clocks that time us. Cambridge, MA: Harvard University Press [Google Scholar]
- 31.Levine JD, Funes P, Dowse HB, Hall JC. 2002. Signal analysis of behavioral and molecular cycles. BMC Neurosci. 3, 1 (doi:10.1186/1471-2202-3-1) [DOI] [PMC free article] [PubMed] [Google Scholar]