

Abstract
Mating signals of many animal species are difficult to produce and thus should indicate signaler quality. Growing evidence suggests that receivers modulate their behaviour in response to signals with varying performance levels, although little is known about if and how responses are affected by receiver attributes. To explore this topic we conducted two experiments with swamp sparrows, Melospiza georgiana, in which we challenged territorial males with playback of songs with trill rates that were natural, digitally reduced, or digitally elevated (control-, low- and high-performance stimuli, respectively). In our first experiment, we found that males responded more aggressively to control songs than to low-performance stimuli, that low-performance stimuli with the most severe trill-rate reductions elicited the weakest aggressive responses, and that the subjects' own trill rates predicted aggressive responses. In our second experiment, we found that male responses to high-performance stimuli varied significantly, in ways predicted by two factors: the degree to which we had elevated stimulus performance levels of high-performance stimuli, and subjects' own vocal performance levels. Specifically, males were less aggressive towards stimuli for which we had elevated performance levels to higher degrees, and subject males with higher vocal performances themselves responded more aggressively. These findings together offer a novel illustration of how responses to aggressive signals may rely not just on signal attributes, but also on attributes of responding animals themselves.
Keywords: sexual selection, competition, performance, soft song, individual differences
1. Introduction
Competitive interactions among animals are often mediated by stereotyped signals of aggression, which can help defuse interactions before they lead to direct combat [1,2]. The dynamics of aggressive signalling have been studied from both theoretical and empirical perspectives, and two main predictions have emerged. The first is that aggressive signals should generally provide reliable indicators of signaller attributes (e.g. resource holding potential, aggressive motivation), including the risks they represent to potential rivals. The reliability of aggressive signalling is thought to be maintained by numerous mechanisms including production constraints and receiver retaliation [3,4]. Second, theory predicts that animals should evaluate aggressive signals with respect to their own attributes and adjust their responses accordingly [5,6]. For example, in snapping shrimp, Alpheus heterochaelis, chelae displays are thought to signal size and fighting ability, and male responses to open chelae display models depend on the size of their own chelae relative to that of the model [7]. Operationally, it is often difficult to measure and manipulate signal values, especially when experiments pairing known opponents are not possible. Much remains to be learned about how animals modulate their responses across a spectrum of available signal values, and how receiver attributes might interact with signal values in shaping receiver responses.
Significant advances in understanding the function of aggressive signals have emerged from studies of songbirds, in which acoustic signal parameters can be manipulated with fine precision. In a majority of songbird species, males produce stereotyped songs used for both mate attraction and territory defence [8]. In territorial interactions, certain song parameters provoke aggressive responses, presumably because they provide reliable indicators of threat levels posed by territorial rivals [9–11]. In general, low-threat rivals should be relatively unlikely to provoke attack, given that they should present few risks to territorial males. For example, male great tits (Parus major) respond with reduced aggression to playback of songs of ‘losers’, i.e. of simulated intruders that appeared to have lost in contests against other males [12]. By contrast, we expect truly threatening intruders to induce territory holders to respond aggressively, or else to retreat.
Comparatively overlooked here is the possibility that males' responses to song are also linked to variation in their own ‘quality’ attributes such as physiological condition, fighting prowess and the ability to defend preferred resources such as breeding territories. We presume that territory holders of high quality would be especially likely to respond aggressively to territorial challenges. While a high-quality territory owner would likely suffer large costs if he lost a challenge, he may have sufficient fighting ability to repel most competitors. By contrast, receivers of relatively low quality might be expected to avoid or retreat from high-threat competitors to prevent costly conflict and risk of injury, even it if means giving up an established territory. Operationally, in playback studies in which high-quality intruders are simulated, one might expect to see both types of responses, with some birds attacking and others retreating [13,14].
In some species, the aggressive content of song—i.e. the extent to which song indicates aggressive motivation or intent—appears to be encoded in measures of trill performance [15]. To elaborate, many songbirds sing trills, in which syllables are repeated in quick succession. Trills are difficult to produce, because the vocal apparatus needs to be actively reconfigured during frequency modulations, and to be ‘reset’ during brief intervals between syllables [15]. Evidence for performance constraints has come from several independent lines, including observations of trade-offs between two acoustic parameters: trill rate and frequency bandwidth [16–18]. Thus, for receivers of both sexes, trill performance may provide a reliable indicator of male quality. Supporting this interpretation, females of various species, including canaries (Serinus canaria), swamp sparrows (Melospiza georgiana) and Lincoln's sparrows (Melospiza lincolnii) respond preferentially to trills with higher vocal performance levels [19–21].
Available evidence suggests that trill performance is also attended to by rival males, but in ways that vary by species and playback context (reviewed by Podos et al. [15]). In field playback studies in which the performance of trilled stimuli varied (either naturally or through experimental manipulation), male aggressive responses to high-performance stimuli were diminished in red-winged blackbirds (Agelaius phoeniceus) [22], yet elevated in banded wrens (Thryothorus pleurostictus), at least in subjects' initial reactions [23]. Schmidt et al. [24], working with European nightingales (Luscinia megarhynchos), reported results similar to those for banded wrens [23], and also observed that males that remained unpaired during that breeding season showed low responses to fast trill playback, whereas males that did later pair with females maintained aggressive responses [24]. In another playback experiment on banded wrens, however, males were found to respond less aggressively to high-performance songs than to medium-performance songs [14]. These studies and others suggest that high-performance trills elicit wide-ranging responses, varying from increased aggression to avoidance. We know little about why some individuals respond to high-performance songs with aggression, whereas others respond with avoidance or retreat.
This study focuses on the swamp sparrow, a species in which males sing repertoires of trilled songs. Vocal performance can be assessed readily by measuring trill rate and frequency bandwidth, and trills for playback presentation can be digitally manipulated to various performance levels without changing other song features [16,17,25] (figure 1). Prior work with swamp sparrows indicates that males discriminate among songs with varying vocal performance levels, giving stronger flight and vocal responses when presented with song types of higher versus lower performance [26]. Additionally, males tend to slightly elevate vocal performance when presented with simulated territorial intrusion [27], although at levels that are not detected by other males in field conditions [26].
Figure 1.

(a) Example spectrograms of playback stimuli used in control and manipulated performance trials. Each subject was presented with two versions of one song type, one at its natural trill rate (left column) and the other version at either a reduced trill rate (experiment 1, right column rows (i) and (ii)) or an elevated trill rate (experiment 2, right column rows (iii) and (iv)). (b) Frequency bandwidth (kHz) plotted as a function of trill rate (Hz) for stimulus and subject songs. The family-wide ‘upper-bound regression’ (line of descending slope), from which ‘vocal deviation’ values are calculated, is provided for reference (see text and Podos [17]). Plotted are control stimuli and songs from subject males (circles), low-performance stimuli (triangles, experiment 1) and high-performance stimuli (diamonds, experiment 2). Open symbols correspond to the eight spectrograms shown above. (Online version in colour.)
We hypothesize that males respond to variation in vocal performance in accordance with the level of threat indicated thereby, and further that responses also vary with males' own vocal performance. To test this hypothesis we presented territorial males with playback of control songs and test songs in which trill rate, and thus perceived vocal performance, was digitally manipulated either to low- (experiment 1) or high-performance levels (experiment 2). For experiment 1, we predicted that responses would be greater to control songs than to low-performance songs, the latter of which should represent universally low levels of threat. For experiment 2, we predicted that responses to high-performance stimuli would vary depending on the vocal performance level of the stimulus. More specifically, we predicted that stimuli with performance levels increased slightly would be responded to aggressively, whereas stimuli increased to the highest performance levels would be avoided, because of the higher perceived risk. Furthermore, for both experiments, we predicted that subjects' tendencies to engage simulated intruders would vary positively with their own vocal performance.
2. Material and methods
(a). Study site and subjects
This study was conducted in a marsh habitat on the Prescott Peninsula of the Quabbin Reservoir in Franklin County, western Massachusetts, USA (42°25′ N, 72°20′ W). Swamp sparrows established territories of roughly 400–600 m2 in late April and early May, and actively defended these territories through late July. Beginning in 2005, we captured birds with mist-nets and gave males unique colour and United States Fish and Wildlife Service (FWS) aluminium band combinations (IACUC 28-10-02).
We mapped territories, determined nest locations when possible, and estimated breeding status. By estimating breeding status, we were able to conduct playback trials during a common stage, before initial clutches or between successive clutches.
(b). Song recording and vocal performance
Between 2005 and 2010, we compiled a library of song recordings from males in our population, made using Marantz PMD660 digital recorders and Sennheiser ME66 directional microphones. Swamp sparrows sing a modest repertoire of two to five song types, which can be classified by the number and category of notes in a syllable [28]. Spectrograms of recordings were reviewed on Audacity (http://audacity.sourceforge.net) for classification. For each male, the song type most commonly produced during playback trials was used to calculate his vocal performance [26,27].
For songs selected as playback stimuli, and for songs recorded from study subjects (n = 3–5 renditions per bird), we estimated vocal performance using measurements of trill rate (the average rate of syllable repetition in a song, in hertz) and frequency bandwidth (the range of frequencies represented in the song, in kilohertz) with the program Signal (Engineering Design 2003). Trill rate was measured across eight syllables in the middle of each song, and frequency bandwidth was calculated from amplitude spectra using a threshold of −24 dB relative to the peak amplitude [17]. A metric of vocal performance, ‘vocal deviation’, was calculated as in [29]: average trill rate and frequency bandwidth for each song type were plotted, and the distance from each point to the upper-bound regression for emberizids (y = –0.124x + 7.5, [17]) was calculated. Greater vocal deviation values represent lower performance, whereas smaller deviations represent higher performance [17,20] (figure 1 and electronic supplementary material, table S1). With this method, each male and playback stimulus was assigned a vocal performance score.
(c). Experiment 1: low- versus control-performance
The goal of our first experiment was to compare aggressive responses with low- versus control-performance songs. To prepare experimental stimuli, we selected songs from high-quality recordings from 11 males of known identity. Each song used to construct a playback stimulus was a unique combination of singer and song type, with no song used more than once. These songs varied in the number and category of notes per syllable, and represented natural variation in swamp sparrow song across our population.
Next, we created a ‘control-’ and a ‘low’-performance version of each of the 11 songs, resulting in 22 playback stimuli. All playback stimuli were constructed by concatenating a single syllable into a repeated sequence of syllables (a song) of 2 s in duration, following previous methods ([16,25]; figure 1). In constructing control stimuli, we preserved the original wild-recorded syllable timing and trill rate. Control stimuli naturally ranged from 4.35 to 10.61 Hz in trill rate, and from 1.82 to 3.19 in vocal deviation. Low-performance stimuli were constructed by digitally adding silent intervals between notes and syllables proportionally, thus adjusting syllable timing while preserving the original structure of individual notes (as in Lahti et al. [25]). Songs were slowed to 35–80% of their original trill rates, resulting in stimuli ranging from 2.19 to 7.49 Hz in trill rate, and 2.20–3.76 in vocal deviation. All stimuli were standardized to the same maximum amplitude, and played in trials at one song per 10 s for 3 min, which approximates the natural singing rate for this species [20,26,27].
Playback trials were conducted during June and July of 2009 and 2010. Each bird received two trials, and each trial consisted of either the control- or the low-performance version of the same song type. Song stimulus sets were selected randomly from banded males that were unfamiliar to the subject males, either recorded from a male out of earshot or from a male only present in a year in which the subject male was not. The order of presentation for control- versus low-performance was determined using a balanced design.
Each subject's two trials were separated by 1–3 days, and were conducted between 06.00 and 12.00. We placed a portable field speaker (SME-AFS speaker, Magnavox CD player) just within a subject's territory, in a bush or fixed to a tree at roughly 1.5 m height, in the same location for both trials. Two observers were present at all trials (D.L.M. and D.C.L.) to ensure continuous observation of playback subjects. One observer dictated response behaviours and recorded subjects' vocalizations with a Marantz digital recorder, whereas the other observer dictated and filmed behaviour with a digital video camera (Canon Handycam) from a distance of approximately 10 m from the speaker.
Before each trial, we located the subject male within the territory and recorded his behaviour. We recorded, filmed and annotated the behaviour of subjects during the 3 min stimulus playback as well as for an additional 7 min post-playback. Response behaviours during the 10 min block recorded included the production of broadcast and soft songs, flights, passes within 2 m of the speaker, closest approach to the speaker and latency to first song. The two observers conferred on distance estimates before and after each trial, and compared and reconciled differences between observations.
Responses to control- versus low-performance stimuli were quantified on a per-response variable basis, and then reduced using principal component analysis (PCA) to one overall measure of aggressive response, PC1. We used the raw and PC1 response scores for each male to compare responses between low- and control-performance trials, using Wilcoxon signed-ranks tests (JMP SAS and R).
We then conducted multiple regression analysis with model selection, in which we regressed subjects' overall aggressive responses to low-performance trials (PC1 scores) against selected predictor variables. We first tested our manipulation of the playback stimuli alone as a predictor, and then tested all possible models, including variables relating to the subject males. This analysis aimed to test whether quantitative variation in the stimulus manipulation would have a systematic effect on birds' responses (for a similar analysis, see [25]), and whether birds' own vocal performance levels predicted their responses to the playback stimuli. We evaluated the models using Akaike's information criteria (AIC), to identify the model with the best-fit.
(d). Experiment 2: high- versus control-performance
The goal of our second experiment was to test levels of threat signalled by high- versus control-performance songs. We predicted that males would respond more or less strongly to such stimuli, depending on the extent to which we increased the performance level of playback songs. The study subjects were 19 male swamp sparrows, none of which was used in experiment 1. Experiment 2 was conducted in June and July of 2006–2008, and followed the methods of experiment 1 except as indicated below.
We created 19 sets of ‘control’- and ‘high’-performance stimuli, and each subject male received a unique song type by singer combination. We prepared control stimuli as in experiment 1, using wild-recorded songs that ranged from 4.75 to 9.48 Hz in trill rate and 1.96–3.31 in vocal deviation. To create high-performance stimuli, we increased the trill rate by deleting silent intervals between notes and syllables proportionally, following procedures used previously [16,30]. Songs were increased to 115–155% of their wild-recorded rate, which resulted in high-performance stimuli with trill rates of 7.0–14.1 Hz and vocal deviations from 1.39 to 3.03. Each subject received two trials, one song type at control and one at high performance.
In addition to the response variables of experiment 1, we also measured the time subject males spent within 1 or 3 m of the speaker, to account for our observation that some males tended to flee or avoid the speaker location, whereas others made aggressive approaches and stayed in close proximity. Because some males fled during the initial stimulus period but returned later, we calculated broadcast song and flight rate for multiple time blocks including the 3 min playback period and the total trial period, and we calculated the post-playback period for song rate as males tended to increase song rate after cessation of the stimulus.
As in experiment 1, we initially examined differences between the two stimulus categories, control- and high-performance, for each response variable. Our next goal was to test the effect of both the stimulus manipulation and the subject male's vocal performance on response strength. Because some males responded more strongly to control stimuli, whereas others clearly responded more strongly to high-performance stimuli, we determined whether males were more aggressive or avoiding of the high-performance stimulus when compared with the control. To do so, we calculated, for each response variable, the difference between each male's response to high-performance and his response to control stimuli—positive values always indicated stronger response (more aggression) to high performance, whereas negative values indicated stronger response to control. We reduced these response differences with PCA, generating three PCs (table 1). We ran multiple regression analysis with model selection using PCs 1–3 factor scores as the response variables, first testing the effects of our stimulus manipulation alone and then all variables together (listed in table 2). Finally, we evaluated the explanatory power of all variables using model selection with AIC, comparing all combinations of predictor variables and identifying the best-fit, but not over-fit, model.
Table 1.
Principal component analysis of response differences between playback of high- and control-performance measured in experiment 2. Values reduced by PCA were the result of high minus control responses for all variables except closest approach and latency, for which we did the opposite subtraction, so that a larger number would similarly indicate a more aggressive response to the high-performance trial. PCA generated three PCs on which loaded singing variables (PC1), flying variables (PC2) and approach and proximity variables (PC3) with eigenvalues of 3.4 through 2.0. Italics indicate variables that loaded strongly onto each PC.
| principal component | PC1 | PC2 | PC3 |
|---|---|---|---|
| per cent | 31.0 | 25.9 | 17.8 |
| eigenvectors | loadings |
||
|---|---|---|---|
| initial songs | 0.50 | 0.00 | 0.13 |
| total songs | 0.48 | 0.03 | 0.24 |
| post-stimulus songs | 0.40 | 0.04 | 0.37 |
| latency to first song | 0.38 | 0.10 | 0.03 |
| soft songs | 0.35 | 0.16 | 0.03 |
| initial flights | 0.04 | 0.50 | 0.23 |
| total flights | 0.09 | 0.53 | 0.04 |
| passes | 0.04 | 0.52 | 0.08 |
| closest approach | 0.05 | 0.25 | 0.41 |
| time within 1 metre | 0.22 | 0.21 | 0.50 |
| time within 3 metres | 0.18 | 0.22 | 0.55 |
Table 2.
Multiple regression analysis of variables predicting the direction and degree of response to high- versus control-performance stimuli. Results determined by model selection (AIC) indicate the full model as best-fit, which includes the vocal performance of both playback stimuli and subject males as predictor variables for PC3 approach/duration (r2 = 0.725, n = 19, d.f. = 18, F = 4.15, p = 0.018). Asterisk (*) denotes individual variables that achieved significance within the model. See text and figure 2c,d for further explanation.
| predictor variables | β | F | p < |
|---|---|---|---|
| playback variables | |||
| % trill increased | –0.07 | 9.7 | 0.01* |
| trill rate (Hz) | 0.39 | 5.8 | 0.04* |
| notes per syllable | 0.81 | 3.9 | 0.08 |
| trill rate (Hz) × notes/syllable | –0.28 | 1.9 | 0.20 |
| subject male variables | |||
| vocal deviation | 274.3 | 8.1 | 0.02* |
| trill rate | 31.0 | 8.0 | 0.02* |
| frequency bandwidth | 0.27 | 8.0 | 0.02* |
3. Results
(a). Experiment 1: low- versus control-performance
Males responded less strongly to low- than to control-performance stimuli. All six raw response variables followed the same pattern of being weaker during playback of low- versus control-performance stimuli (see electronic supplementary material, figure S1), with the difference across treatments in soft song rate achieving significance (Wilcoxon signed-rank test, z = 2.72, p = 0.004). In PCA, all six raw response variables loaded positively and evenly (loadings between 0.35 and 0.44) onto PC1, which explained 55.9% of the overall variation. We found a significant difference in PC1 scores by condition, with higher responses to control- than to low-performance stimuli (Wilcoxon signed-rank test, n = 11, z = −2.66, p = 0.008, electronic supplementary material, figure S1).
When testing the stimulus manipulation alone, our regression analysis revealed a significant effect of stimulus vocal performance on male aggressive response (see electronic supplementary material, table S2). The best-fit model in our multiple regression was determined to include variables relating to the stimuli and a component of the subject males’ vocal performance, trill rate. The best-fit model included as predictor variables the per cent trill rate manipulation, the difference in vocal deviation between the low and control-performance stimuli, and subject males' trill rates (best-fit model r2 = 0.868, n = 11, d.f. = 3,7, F = 15.38, p = 0.002. AIC = 1.78 versus 3.4 and higher, see also electronic supplementary material, table S2). Specifically, the more we reduced stimulus trill rate from the wild-recorded rate, the less aggressive males were in response (figure 2a). Additionally, subject male trill rate was included as a positive predictor of aggressive response (figure 2b and electronic supplementary material, table S2), indicating that higher male trill rates tended to correspond to more aggressive responses. Results were robust to sequential-Bonferroni correction for multiple tests.
Figure 2.

Playback responses as a function of signal vocal performance (a,c) and subject vocal performance (b,d) relationships that were identified in the multiple regression analyses. Experiment 1 (a,b): the y-axis represents the aggressive response to the low-performance trial (PC1 scores of six response variables reduced). (a) The x-axis represents the percentage of the original rate for the low-performance playback stimuli (with stimuli closer to the original rate having percentages closer to 100%, and stimuli with comparatively lower trill rates at lower percentages). Males were more aggressive to stimuli at higher performance levels that more closely resembled the original trill rate. (b) The x-axis represents the subject male's trill rate. Males with higher trill rates (Hz) were more aggressive in response to playback (see electronic supplementary material, table S2). Experiment 2 (c,d): the y-axis represents the level of aggressive response differences (PC3 approach and time spent close to the speaker, table 1), with positive values indicating a more aggressive response to high-performance over control-performance stimuli, and negative values indicating more avoidance of high-performance stimuli and thus more aggression to control-performance stimuli. (c) The x-axis represents percentage of the original rate for the high-performance stimuli (increases from the original rate of 100%, to 115–155% for high performance; table 2 and electronic supplementary material, tables S1 and S3). The more the vocal performance of the playback stimulus was increased, the less likely males were to approach aggressively in terms of distance and time spent close to the speaker. (d) The x-axis represents the vocal performance of the subject males, with higher values indicating males with higher vocal performance (table 2). Males with higher vocal performance themselves approached the high-performance speaker more aggressively and remained longer in close proximity.
(b). Experiment 2: high- versus control-performance
On average, males appeared to respond as aggressively to control- as to high-performance stimuli (see electronic supplementary material, figure S1), but males varied individually as to which stimulus they responded to more aggressively. Specifically, males responded to high-performance trials with either elevated aggression or with diminished aggression and avoidance. Differences in responses reduced to three PCs and cumulatively explained 74.7% of the variation (table 1). Singing behaviours (songs, soft-songs) loaded most strongly onto PC1, flying behaviours (flights, passes) onto PC2, and approach behaviours (closest approach, time spent near the speaker) onto PC3. Another variable that loaded somewhat strongly on PC3 was the rate of singing after cessation of the playback stimulus.
In our multiple regression analysis with model selection, models with PC1 or PC2 as response variables were not significant (p > 0.05), but models testing PC3 were significant (see electronic supplementary material, table S3). In model selection analysis for PC3 (approach, time spent close), the best-fit model was determined to be the full model and included predictor variables of both playback stimuli and subject males (table 2, n = 19, r2 = 0.725, F = 4.15, p = 0.018, AIC = 3.23 versus 4.06 and larger; sum of squared errors = 9.7). There was a significant negative effect of the per cent stimulus manipulation indicating that higher stimulus performance levels were associated with males staying farther away and spending less time close to the speaker (figure 2c). For predictor variables relating to the subject males, there was a significant positive effect; the higher a male's own vocal performance (high trill rate and frequency bandwidth), the more likely he was to approach and spend time closer to the speaker during high-performance stimulus playback (figure 2d).
4. Discussion
As expected, responses to song playback in both experiments varied with stimulus attributes. In our first experiment, territorial male swamp sparrows responded significantly less strongly to low-performance than to control-performance playback stimuli, consistent with our hypothesis that receivers should attribute limited threat to low-performance songs (see electronic supplementary material, figure S1). Additionally, males' reduced aggression to low-performance stimuli was more pronounced for playback stimuli that had been lowered further from their control trill rates (figure 2a). Notably, males sang significantly fewer soft songs to low-performance stimuli (see electronic supplementary material, figure S1); singing softly in response to simulated territorial intrusion is known to be a highly reliable predictor of attack in swamp sparrows [31], song sparrows (Melospiza melodia) [10] and black-throated blue warblers (Setophaga caerulescens) [11]. This suggests that males would be more likely to attack intruders singing control- versus low-performance songs.
In our second experiment, high-performance stimuli elicited aggressive responses in some individuals, yet avoidance behaviour in others, resulting in similar overall outcomes across treatments (see electronic supplementary material, figure S2). Subjects' tendencies to respond one way or the other varied consistently with performance levels of playback stimuli. Specifically, we found that birds did not approach as closely or remain near the speaker during presentation of the stimuli that we had manipulated to the greatest extent, as measured by the percentage increase of stimuli above their original trill rate (table 2 and figure 2c). Closest approach was previously indicated as a reliable predictor of attack in swamp sparrows [31]. Prior studies in banded wrens, red-winged blackbirds and nightingales showed differential male approaches to high-performance playback, which either deterred or provoked aggressive approach by subjects [14,22–24]. Similar to DuBois et al. [26], we found that swamp sparrows responded less to low-performance stimuli and responded strongly to stimuli ranging from control to slightly increased trill rates (higher performance). While their study tested songs within average inter-male differences, we expanded on this by testing a continuum of vocal performance levels, pushed to low and high extremes, in order to determine factors that would predict aggression versus avoidance particularly in response to high performance.
A key additional finding in both our experiments was that responses varied in accordance with the vocal performance of study subjects themselves. In experiment 1, birds' tendencies to respond with greater strength were predicted by their own trill rates, with birds singing higher trill rates tending to respond comparatively more aggressively with playback (figure 2b and electronic supplementary material, table S2). In experiment 2, subjects that possessed higher vocal performance levels approached high-performance stimuli more closely and spent more time close to the playback speaker, whereas subject males singing lower performance songs remained at a farther distance from the playback speaker (table 2 and figure 2d).
How variation in subject attributes (e.g. vocal parameters, overall quality) shape responses to territorial playback has been virtually overlooked in studies of signal function in songbirds. Our data suggest that subject attributes can be important predictors of responses to territorial intrusion. Available evidence from several songbird species indicates that vocal performance can correlate with age, size or other quality attributes [32,33], and males with high-performance songs would therefore be better positioned to ward off the challenge of high-performance intruders. In agonistic contests, the importance of subjects' own attributes as predictors of behaviour has been established in other taxa [34], but fewer studies have shown the connection to receiver signal values specifically [7,35]. For example, in a study of hermit crabs (Pagurus berhardus), model intruders that signalled aggressively were seen to elicit varied responses, with the likelihood of different responses (attack or retreat) depending on whether study subjects had initially signalled with threat displays themselves [36]. Focusing on signal values specifically, a recent study of poison frogs, Dendrobates pumilio, showed that males responded more aggressively (quicker approaches and more frequent calls) to stimulus frogs with brighter dorsal colours, and moreover subjects' own brightness predicted their aggressive calling and approach [35]. Similarities between these studies and ours support the generality of the interpretation that receivers with high signal values themselves are more aggressive in their responses to opponents, especially those that are highly threatening.
Returning to our data, we offer a possible explanation for the main difference between our two experiments, i.e. why average response scores differed between control- versus low-performance stimuli but not between control- versus high-performance stimuli. On the one hand, disregarding low-performance signals should not carry high costs for any males, even those who themselves are of low quality. Males on territories would likely suffer little impact on their reproductive success if they were to tolerate younger or poorer-quality males on their territories, as it is unlikely that females would select males with low-performance songs for extra-pair mating [20,32,37,38] (D. L. Moseley 2012, unpublished data). It thus may not be surprising that responses to low-performance stimuli were uniformly low. On the other hand, failing to respond to high-performance signals, even ones that push the bounds of species-typical acoustic structure, could readily result in the loss of a territory to a higher-quality opponent [39], loss of paternity from eavesdropping by females and other rivals [40], or in direct conflict. If a perceived threat is too great, however, territorial males may benefit from avoiding such costly conflicts [41], and we often observed that the immediate response of some males to some high-performance stimuli was to flee.
An important assumption we are making in interpreting our data is that males are able to recognize the performance levels of song stimuli, and to modulate their responses accordingly. An alternative possible explanation for reduced responses to manipulated songs is that males failed to recognize such songs as conspecific. Indeed, a few of our manipulated stimuli featured trill rates of less than 4.3 Hz and more than 12.5 Hz, which are outside the natural range of swamp sparrows' trill rates (although not vocal deviations), but these made up less than one-third of all stimuli. However, this alternative explanation is countered by song learning studies that have shown that male swamp sparrows perceive and successfully memorize songs manipulated to much higher and lower performance levels, treating these songs as viable learning models [16,25,30]. Hand-reared male swamp sparrows were found to be able to memorize with precision song models of extremely high-performance levels (trill rates increased to 160–187%) in the early sensory-motor phase [16,30]. At the other end of the spectrum, young male swamp sparrows were found to be able to learn song models reduced to as little as 55% of their original trill rates, albeit with reduced copying accuracy [25]. Also consistent with our inference that all stimuli were recognized as conspecific is the observation that, in this study, males responded with some aggressive behaviour, including broadcast songs and passes in close range of the speaker, even to stimuli that were altered considerably, for example, to 35% or 155% of the original trill rate. In experiment 1, none of the 11 males completely ignored the low-performance stimuli, but instead responded with some aggressive singing, flying and approach behaviour. We thus infer that low-performance songs were recognized as relevant, yet did not represent threats sufficient to elicit strong responses [10]. In experiment 2, as mentioned above, some individuals reserved their most aggressive responses for high-performance stimuli. It is also notable that males possessing songs of lower performance, who were initially deterred during playback during the high-performance trial, often returned after playback ended, and then sang at rates comparable with those of other males that had responded strongly during playback. This recovery of responses is consistent with behaviours of males interacting with a conspecific intruder [8].
Taken together, our results provide a novel line of support for the hypothesis that vocal performance provides a reliable signal of aggressive threat. Searcy & Nowicki [3] outlined three criteria needed to demonstrate that a signal is reliable: (i) some aspect of the signal must correlate with signaller attributes; (ii) receivers should respond to the signal in ways consistent with its presumed function; and (iii) receivers should respond in ways that provide them with overall fitness benefits. Considering the potential reliability of vocal performance in songbirds, the first criterion had been addressed in previous studies, including in swamp sparrows [32,33]. These studies showed that older or larger males tend to sing at higher vocal performance levels, thus presumably presenting greater threats to territorial rivals. Similarly, in our study, higher vocal performance or trill rate correlated with increased approach and aggression (figure 2b,d). This helps to explain why males with higher vocal performance responded more aggressively to high-performance stimuli. Our data also support the second reliability criterion, by showing that male swamp sparrows respond to playback stimuli in degrees consistent with presumed levels of threat (low responses to low performance and aggressive or avoidance behaviour in response to high performance). Finally, although not tested explicitly here, our data are consistent with the third criterion. Low-quality males should benefit by retreating from high-threat signals to avoid the risk of costly combat, whereas high-quality males should benefit by being aggressive, so as to limit the risk of losing a territory or paternity. The potential fitness benefits of singing higher-performance songs could be assessed most directly in field studies of territorial males, in which vocal behaviour, territorial interactions and breeding success are all quantified.
Most broadly, our data contribute to a general understanding of how animals respond to signals or signallers that are threatening. Traditionally, researchers predicted that playback of threatening signals should result in defensive responses proportional to the level of threat signals represent [42], a prediction supported in empirical studies including some that have focused on trilled vocalizations [25]. More recent studies, however, report circumstances in which high-performance signals elicit less of an aggressive response than do signals of lower or more intermediate levels of performance [14,22]. A possible explanation for this outcome is that intruders signalling at particularly high-threat levels might present too great a risk for a territory holder to engage in conflict [13,14]. Our study extends this suggestion by showing that birds' responses to high-threat stimuli correspond not just to the degree of threat presented by an intruder, but also to their own vocal performance, as predicted more generally by Maynard-Smith & Parker [5]. We do not yet know whether this correspondence is best explained as self- or mutual-assessment [34], nor do we know whether there is an underlying physiological relationship between vocal performance and male attributes such as resource holding potential. In figure 3, we present a schematic model that summarizes male responses to territorial intrusion (y-axis) as it varies with both factors (see legend for more detail). One could potentially test this model using a factorial design that presents subject males of known vocal performance with playbacks of much higher and much lower vocal performances.
Figure 3.
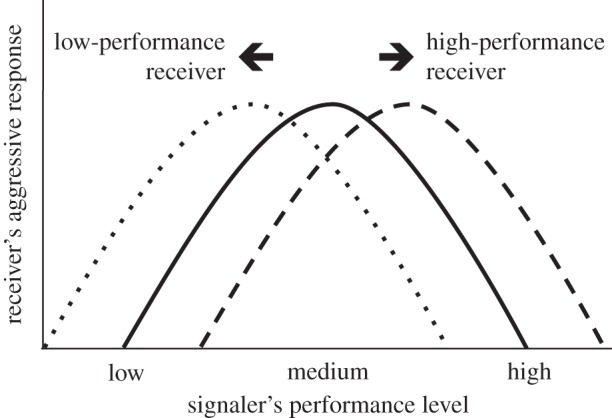
A conceptual model describing the intensity of responses by territorial males to simulated territorial intrusion, as we expect it to vary with two factors: the level of threat signaled by an intruder and the subjects' own vocal performance levels. The x-axis represents the level of threat signalled by the intruder as the level of vocal performance, and the y-axis represents the aggressive response intensity of the receiver (the territorial male). Response intensity has recently been modelled as a parabolic function, to account for the expectation that territorial males should avoid rather than confront intruders with high-threat signals (solid line, from de Kort et al. [14], modified from Collins [13]). We extend this model by accounting for expected variation in the quality of territorial subjects, here reflected in the vocal performance of the songs of playback subjects. As illustrated in this model, we predict that the parabolic function would shift to the right for territory owners producing high-performance signals (dashed line), and to the left for territory owners producing low-performance signals (dotted line). According to this model, as intruder signals increase in performance (levels of threat), low-performance territory holders would switch from aggression to avoidance sooner than would high-performance territory holders. These expectations are consistent with the outcomes of our multiple regression models and the relationships illustrated in figure 2.
To conclude, responses of territorial males to varying vocal performances of rivals correlate not just with threat levels posed by intruders (as shown in prior studies), but also by subjects' own vocal performance capabilities (novel to this study). Researchers seeking to identify signalling traits that mediate aggressive contests thus should take into consideration not only the properties of signal features, but also the individual quality and signalling capabilities of their study subjects, and potential interactions among signaller and receiver attributes.
Acknowledgements
We thank the Massachusetts Department of Conservation and Recreation (DCR) for access to our field site; Christine Rega, Greg LeBouef, and Chrissy Rivera for help with data management; Steve Johnson for his expert help in the field. We greatly appreciate Bruce Byers, Beth Jakob, Sarah Goodwin, David Hof, and three reviewers who gave feedback that improved the manuscript.
Funding statement
Funding provided by the NSF DDIG no. 1011241 and Cooper Ornithological Society Joseph Grinnell Award to D.L.M., NIH NRSA grant to D.C.L. and J.P. and NSF grant no. IBN-0347291 to J.P.
References
- 1.Maynard-Smith J, Price GR. 1973. Logic of animal conflict. Nature 246, 15–18 (doi:10.1038/246015a0) [Google Scholar]
- 2.Andersson M. 1994. Sexual selection. Princeton, NJ: Princeton University Press [Google Scholar]
- 3.Searcy W, Norwicki S. 2005. The evolution of animal communication: reliability and deception in signaling systems (monographs in behavior and ecology). Princeton, NJ: Princeton University Press [Google Scholar]
- 4.Maynard-Smith J, Harper D. 2003. Animal signals. Oxford, UK: Oxford University Press [Google Scholar]
- 5.Maynard-Smith J, Parker GA. 1976. Logic of asymmetric contests. Anim. Behav. 24, 159–175 (doi:10.1016/S0003-3472(76)80110-8) [Google Scholar]
- 6.Enquist M. 1985. Communication during aggressive interactions with particular reference to variation in choice of behavior. Anim. Behav. 33, 1152–1161 (doi:10.1016/S0003-3472(85)80175-5) [Google Scholar]
- 7.Hughes M. 1996. Size assessment via a visual signal in snapping shrimp. Behav. Ecol. Sociobiol. 38, 51–57 (doi:10.1007/s002650050216) [Google Scholar]
- 8.Catchpole C, Slater P. 2008. Bird song. Cambridge, UK: Cambridge University Press [Google Scholar]
- 9.Burt JM, Campbell SE, Beecher MD. 2001. Song type matching as threat: a test using interactive playback. Anim. Behav. 62, 1163–1170 (doi:10.1006/anbe.2001.1847) [Google Scholar]
- 10.Searcy WA, Anderson RC, Nowicki S. 2006. Bird song as a signal of aggressive intent. Behav. Ecol. Sociobiol. 60, 234–241 (doi:10.1007/s00265-006-0161-9) [Google Scholar]
- 11.Hof D, Hazlett N. 2010. Low-amplitude song predicts attack in a North American wood warbler. Anim. Behav. 80, 821–828 (doi:10.1016/j.anbehav.2010.07.017) [Google Scholar]
- 12.Peake TM, Terry AMR, McGregor PK, Dabelsteen T. 2002. Do great tits assess rivals by combining direct experience with information gathered by eavesdropping? Proc. R. Soc. Lond. B 269, 1925–1929 (doi:10.1098/rspb.2002.2112) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Collins S. 2004. Vocal fighting and flirting: the functions of birdsong. In Nature‘s music: the science of bird song (eds Marler P, Slabbekoorn H.), pp. 39–79 San Diego, CA: Elsevier [Google Scholar]
- 14.de Kort SR, Eldermire ERB, Cramer ERA, Vehrencamp SL. 2009. The deterrent effect of bird song in territory defense. Behav. Ecol. 20, 200–206 (doi:10.1093/beheco/arn135) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Podos J, Lahti DC, Moseley DL. 2009. Vocal performance and sensorimotor learning in songbirds. Adv. Study Behav. 40, 159–195 (doi:10.1016/S0065-3454(09)40005-6) [Google Scholar]
- 16.Podos J. 1996. Motor constraints on vocal development in a songbird. Anim. Behav. 51, 1061–1070 (doi:10.1006/anbe.1996.0107) [Google Scholar]
- 17.Podos J. 1997. A performance constraint on the evolution of trilled vocalizations in a songbird family (Passeriformes: Emberizidae). Evolution 51, 537–551 (doi:10.2307/2411126) [DOI] [PubMed] [Google Scholar]
- 18.Suthers RA, Vallet E, Kreutzer M. 2012. Bilateral coordination and the motor basis of female preference for sexual signals in canary song. J. Exp. Biol. 215, 2950–2959 (doi:10.1242/jeb.071944) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Draganoiu TI, Nagle L, Kreutzer M. 2002. Directional female preference for an exaggerated male trait in canary (Serinus canaria) song. Proc. R. Soc. Lond. B 269, 2525–2531 (doi:10.1098/rspb.2002.2192) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ballentine B, Hyman J, Nowicki S. 2004. Vocal performance influences female response to male bird song: an experimental test. Behav. Ecol. 15, 163–168 (doi:10.1093/beheco/arg090) [Google Scholar]
- 21.Caro SP, Sewall KB, Salvante KG, Sockman KW. 2010. Female Lincoln's sparrows modulate their behavior in response to variation in male song quality. Behav. Ecol. 21, 562–569 (doi:10.1093/beheco/arq022) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Cramer ERA, Price JJ. 2007. Red-winged blackbirds Ageliaus phoeniceus respond differently to song types with different performance levels. J. Avian Biol. 38, 122–127 (doi:10.1111/j.2006.0908-8857.03839.x) [Google Scholar]
- 23.Illes AE, Hall ML, Vehrencamp SL. 2006. Vocal performance influences male receiver response in the banded wren. Proc. R. Soc. B 273, 1907–1912 (doi:10.1098/rspb.2006.3535) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Schmidt R, Kunc HP, Amrhein V, Naguib M. 2008. Aggressive responses to broadband trills are related to subsequent pairing success in nightingales. Behav. Ecol. 19, 635–641 (doi:10.1093/beheco/arn021) [Google Scholar]
- 25.Lahti DC, Moseley DL, Podos J. 2011. A tradeoff between performance and accuracy in bird song learning. Ethology 117, 802–811 (doi:10.1111/j.1439-0310.2011.01930.x) [Google Scholar]
- 26.DuBois AL, Nowicki S, Searcy WA. 2011. Discrimination of vocal performance by male swamp sparrows. Behav. Ecol. Sociobiol. 65, 717–726 (doi:10.1007/s00265-010-1073-2) [Google Scholar]
- 27.DuBois AL, Nowicki S, Searcy WA. 2009. Swamp sparrows modulate vocal performance in an aggressive context. Biol. Lett. 5, 163–165 (doi:10.1098/rsbl.2008.0626) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Marler P, Pickert R. 1984. Species-universal microstructure in the learned song of the swamp sparrow (Melospiza georgiana). Anim. Behav. 32, 673–689 (doi:10.1016/S0003-3472(84)80143-8) [Google Scholar]
- 29.Podos J. 2001. Correlated evolution of morphology and vocal signal structure in Darwin's finches. Nature 409, 185–188 (doi:10.1038/35051570) [DOI] [PubMed] [Google Scholar]
- 30.Podos J, Nowicki S, Peters S. 1999. Permissiveness in the learning and development of song syntax in swamp sparrows. Anim. Behav. 58, 93–103 (doi:10.1006/anbe.1999.1140) [DOI] [PubMed] [Google Scholar]
- 31.Ballentine B, Searcy WA, Nowicki S. 2008. Reliable aggressive signaling in swamp sparrows. Anim. Behav. 75, 693–703 (doi:10.1016/j.anbehav.2007.07.025) [Google Scholar]
- 32.Ballentine B. 2009. The ability to perform physically challenging songs predicts age and size in male swamp sparrows, Melospiza georgiana. Anim. Behav. 77, 973–978 (doi:10.1016/j.anbehav.2008.12.027) [Google Scholar]
- 33.de Kort SR, Eldermire ERB, Valderrama S, Botero CA, Vehrencamp SL. 2009. Trill consistency is an age-related assessment signal in banded wrens. Proc. R. Soc. B 276, 2315–2321 (doi:10.1098/rspb.2009.0127) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Elwood RW, Arnott G. 2012. Understanding how animals fight with Lloyd Morgan's canon. Anim. Behav. 84, 1095–1102 (doi:10.1016/j.anbehav.2012.08.035) [Google Scholar]
- 35.Crothers L, Gering E, Cummings M. 2011. Aposematic signal variation predicts male–male interactions in a polymorphic poison frog. Evolution 65, 599–605 (doi:10.1111/j.1558-5646.2010.01154.x) [DOI] [PubMed] [Google Scholar]
- 36.Laidre ME. 2009. How often do animals lie about their intentions? An experimental test. Am. Nat. 173, 337–346 (doi:10.1086/596530) [DOI] [PubMed] [Google Scholar]
- 37.Searcy WA, Peters S, Kipper S, Nowicki S. 2010. Female response to song reflects male developmental history in swamp sparrows. Behav. Ecol. Sociobiol. 64, 1343–1349 (doi:10.1007/s00265-010-0949-5) [Google Scholar]
- 38.Byers J, Hebets E, Podos J. 2010. Female mate choice based upon male motor performance. Anim. Behav. 79, 771–778 (doi:10.1016/j.anbehav.2010.01.009) [Google Scholar]
- 39.Searcy WA, Brenowitz EA. 1988. Sexual differences in species recognition of avian song. Nature 332, 152–154 (doi:10.1038/332152a0) [Google Scholar]
- 40.Mennill DJ, Ratcliffe LM, Boag PT. 2002. Female eavesdropping on male song contests in songbirds. Science 296, 873 (doi:10.1126/science.296.5569.873) [DOI] [PubMed] [Google Scholar]
- 41.Hof D, Hazlett N. 2012. Mortal combat: an apparent intraspecific killing by a male black-capped Chickadee. J. Field Ornithol. 83, 290–294 (doi:10.1111/j.1557-9263.2012.00377.x) [Google Scholar]
- 42.Falls JB, Krebs JR, McGregor PK. 1982. Song matching in the great tit (Parus major): the effect of similarity and familiarity. Anim. Behav. 30, 997–1009 (doi:10.1016/S0003-3472(82)80188-7) [Google Scholar]