

Abstract
Ribonuclease P is an ancient enzyme that cleaves pre-tRNAs to generate mature 5′ ends. It contains an essential RNA subunit in Bacteria, Archaea, and Eukarya, but the degree to which the RNA subunit relies on proteins to supplement catalysis is highly variable. The eukaryotic nuclear holoenzyme has recently been found to contain almost twenty times the protein content of the bacterial enzymes, in addition to having split into at least two related enzymes with distinct substrate specificity. In this review, recent progress in understanding the molecular architecture and functions of nuclear forms of RNase P will be considered.
Nearly 20 years ago it was discovered that RNAs could have enzymatic activity, as well as merely assisting in enzymatic reactions catalyzed by proteins. An increasing number of different RNA catalysts, or ribozymes, have been identified in biological settings and characterized in detail (reviewed in Cech, 1993; McKay and Wedekind, 1999). Two examples of catalytic RNA, ribonuclease P and the ribosome are of particular interest, because they are found in virtually all living organisms and are likely to be holdovers from a very early biotic or pre-biotic world. The existence of such ancient RNA enzymes has stimulated the proposition of an early “RNA World,” where RNA (or something like RNA) both encoded its own reproduction and acted enzymatically to bring about that reproduction (Joyce and Orgel, 1993). Recently, the detailed crystal structures of several ribosome subunits have become available, allowing the conclusion that the catalytic centers of peptide synthesis are almost exclusively ribosomal RNA and transfer RNA, with the protein subunits excluded at more peripheral positions (Cate et al., 1999; Ban et al., 2000; Carter et al., 2000; Nissen et al., 2000; Wimberly et al., 2000). Thus, earlier speculations that ribosomal RNAs were descended from early RNA enzymes were strongly supported.
RNase P IS AN ANCIENT RIBOZYME
The RNA subunit of ribonuclease P (RNase P) was one of the first identified ribozymes, and remains the only non-ribosome enzyme found in all living organisms that contains a catalytic RNA subunit. RNase P activity has been identified in a wide variety of organisms including Bacteria, Archaea, and Eucarya, and in virtually all cases the enzyme has been shown to have an essential RNA subunit. Bacterial RNase P consists of an RNA and a small protein subunit, while the protein compositions of the Archaeal and Eucaryal holoenzymes are more complex (see below). The main function of the enzyme in vivo appears to be endonucleolytic cleavage of tRNA precursor transcripts (pre-tRNAs) to give mature 5′ termini. Substrate recognition is determined more by the tertiary structure than by specific sequences in pre-tRNAs.
Although crystal structures for the RNase P holoenzyme and RNA subunit are not yet available, it is clear that the catalytic center of at least the bacterial RNase P is contained entirely in the RNA subunit (Guerrier-Takada et al., 1983; Guerrier-Takada and Altman, 1984; Gardiner et al., 1985). The bacterial RNA subunits alone are capable of correctly recognizing and cleaving pre-tRNA substrates, although the small protein subunits aid both the folding of the RNA subunits (Reich et al., 1988; Talbot and Altman, 1994; Westhof et al., 1996) and recognition of the pre-tRNA (Kurz et al., 1998; Niranjanakumari et al., 1998). In eukaryotes, the RNA subunits have so far been proven incapable of cleaving substrates in the absence of proteins. Archaeal RNA subunits appear to exist somewhere between the catalytically competent and incompetent states in the absence of their presumed protein partners. Small amounts of substrate cleavage can be obtained at extreme salt concentrations from selected branches of the Archaeal RNase P RNA tree, but not others (Pannucci et al., 1999).
Variable forms of RNase P in eukaryotes
Several forms of RNase P activity have been identified in eukaryotes, including those found in nuclei, mitochondria and chloroplasts. Nuclear RNase P has been detected from a wide range of eukaryotes and isolated from both yeast and human (Eder et al., 1997; Chamberlain et al., 1998). The composition of the nuclear RNase P holoenzyme is far more complex than that of the bacterial enzyme, with one essential RNA subunit and several protein subunits. The increased complexity of the nuclear enzymes will be examined below in detail, but it is worth noting that separate RNase P activities have been described from mitochondria and chloroplasts, and the composition of these enzymes is somewhat controversial at this time.
Studies of RNase P in organelles, i.e., mitochondria and chloroplasts, suggest diversity in their composition and origin. The well-characterized mitochondrial RNase P in Saccharomyces cerevisiae clearly contains an RNA subunit, the RPM1 RNA, as well as at least one protein subunit, Rpm2p (Morales et al., 1992; Dang and Martin, 1993). The mitochondrial protein subunit is not present in the nuclear form of RNase P, yet there is evidence that it serves some cellular function outside the mitochondria (Kassenbrock et al., 1995; Groom et al., 1998; Lutz et al., 2000). The ribonucleoprotein nature of the cyanelle RNase P in the alga Cyanophora paradoxa has also been proven, and is suggested to be more like the nuclear enzymes than the bacterial enzymes (Cordier and Schon, 1999). In contrast, there are reports indicating that the RNase P activities in spinach chloroplasts and human mitochondria might be RNA-independent (Wang et al., 1988; Rossmanith and Karwan, 1998; Thomas et al., 2000a). Spinach chloroplast RNase P has been purified extensively, and has the properties of a solely protein enzyme (Wang et al., 1988; Thomas, 1996) with a pre-tRNA cleavage mechanism distinct from the RNA-based RNase Ps (Thomas et al., 2000a). Evidence concerning RNase P composition from mammalian mitochondria remains contradictory (Altman et al., 2000; Gegenheimer, 2000).
RNA-based RNase P cleavage
The cleavage reaction catalyzed by RNase P has been studied in Bacteria, Archaea and Eucarya, with the mechanism of the bacterial ribozyme reaction being the best characterized. Studies of the eukaryotic RNase P reaction have been primarily performed with partially purified nuclear holoenzyme from several sources (Carrara et al., 1995; Drainas, 1995; Levinger et al., 1995; Arends and Schon, 1997; Pfeiffer et al., 2000; Thomas et al., 2000a, 2000b). The RNase P cleavage reaction can be thought of as three consecutive steps: (1) pre-tRNA binding; (2) hydrolysis of the scissile phosphodiester bond, generating a 3′ hydroxyl group and a 5′ phosphate group, possibly via a ‘trigonal bipyramid’ intermediate (Warnecke et al., 1996); and (3) the dissociation of the mature tRNA and 5′-leader products.
The affinity of bacterial ribozyme for tRNA has been measured by a variety of methods including gel mobility shift, gel-exclusion chromatography and photoaffinity cross-linking (reviewed in Frank and Pace, 1998). The RNase P RNA from B. subtilis binds maturet RNA with a Kd of about 3nM under optimal salt conditions (Beebe and Fierke, 1994). In the case of the yeast nuclear holoenzyme, the substrate acquisition was determined under steady-state conditions giving a KM of approximately 55nM with a natural pre-tRNA (Ziehler et al., 2000a). However, it is worth noting that the many physiological pre-tRNA substrates for RNase P in vivo have widely varied properties that affect their relative rate of utilization by RNase P. Not only are the primary sequences of the mature domains, leaders, trailers and introns different, but the tertiary structure determinants are also slightly variable. Further, it is not clear at this time how many of the base and sugar modifications exist in the pre-tRNAs prior to RNase P cleavage, or whether the actual substrates for RNase P in nuclei are RNA-protein complexes.
The rate constants of different steps of the RNase P reaction have been determined for bacterial ribozymes by both steady-state and pre-steady-state kinetic studies. Under pre-steady-state conditions, the rate of scissile bond cleavage ranges from 180 min−1 to 360 min−1 for the bacterial ribozymes (Smith and Pace, 1993; Beebe and Fierke, 1994). In contrast, the kcat derived under multiple turnover conditions is about 100-fold slower, which appears to be determined by the rate of product release. Initial measurement of the rate constants for the yeast nuclear holoenzyme by steady state analyses give a kcat for the yeast holoenzyme of approximately 1.3 sec−1 (Ziehler et al., 2000a). The second-order rate constant, kcat/KM, for the yeast RNase P reaction is close to the predicted rate of diffusion 1.3×108M−1 sec−1 (Ziehler et al., 2000a).
Divalent metal ions, normally magnesium, are important co-factors for the RNase P reaction. Divalent metal ions strongly enhance the affinity of bacterial ribozyme for tRNA (Smith et al., 1992; Beebe et al., 1996) although tRNA can bind to the RNase P RNA in a similar fashion with and without metal ions as assayed by photoaffinity crosslinking (Smith et al., 1992). In contrast, catalysis by RNase P absolutely requires divalent metal ions. Manganese can substitute for Mg2+ in the ribozyme reaction with a slight defect in catalytic rate, while calcium substitution significantly reduces the catalytic rate of the reaction (Smith and Pace, 1993). These data further suggest the previous hypothesis that the active site of RNase P may form a highly ordered metal-binding pocket that is necessary for discriminating different metal ions (reviewed in Frank and Pace, 1998). The yeast nuclear RNase P also displays a high degree of specificity for magnesium as the divalent metal ion, but it has been very difficult to assess the precise role of the divalent cations because aggressive removal of enzyme-bound divalent cations with EDTA irreversibly inactivates the holoenzyme (Lee, 1991).
Eukaryotic RNase P differs significantly from the bacterial holoenzyme in composition and substrate specificity. The remainder of this review will focus on the current understanding on the structure, function, and subcellular localization of eukaryotic nuclear RNase P, especially from the yeast Saccharomyces cerevisiae.
COMPOSITION OF YEAST NUCLEAR RNase P
Although the RNA subunit of the yeast S. cerevisiae RNase P, RPR1 RNA, was identified almost a decade ago (Lee and Engelke, 1989; Lee et al., 1991a), it is not until recently that the holoenzyme has been biochemically purified to apparent homogeneity (Chamberlain et al., 1998). The size of RPR1 RNA is comparable to its bacterial counterpart, but the protein complement is substantially increased in the yeast holoenzyme. Early density measurements showed that protein contributes about 50% of the mass of the eukaryotic holoenzyme (Kline et al., 1981; Lawrence et al., 1987; Darr et al., 1990; Knap et al., 1990; Doria et al., 1991; Jayanthi and Van Tuyle, 1992), as compared to 10% in the bacterial enzyme (Akaboshi et al., 1980; Gardiner and Pace, 1980). The fact that micrococcal nuclease and proteinase treatments eliminate the yeast RNase P activity originally suggested that both the RNA and protein subunits are required for enzymatic function (Lee and Engelke, 1989). Consistent with this and the density measurements, purified holoenzyme contains RPR1 RNA and nine tightly associated protein subunits (Chamberlain et al., 1998) (Table 1). The identity of four protein subunits (Pop1p, Pop3p, Pop4p and Rpp1p) was previously identified by genetic and immunoprecipitation studies (Lygerou et al., 1994; Chuet al., 1997; Dichtl and Tollervey, 1997; Stolc and Altman, 1997). Enzyme purification confirmed the presence of all four proteins, as well as five additional essential polypeptides, namely Pop5p, Pop6p, Pop7p, Pop8p and Rpr2p.
TABLE 1.
Subunit composition of nuclear RNase P and RNase MRP from yeast (S. cerevisiae) and human
| Yeast gene
|
Subunit type | Molecular weight (kDA)a | Isoelectric point (pI)a | Human gene
|
References | ||
|---|---|---|---|---|---|---|---|
| RNase P | RNase MRP | RNase P | RNase MRP | ||||
| RPR1 | ...... | RNA | 120 | ...... | H1 | ...... | Bartkiewicz et al., 1989; Lee et al., 1991a. |
| ...... | NME1 | RNA | 112 | ...... | ...... | 7-2 | Schmitt and Clayton, 1992; Gold et al., 1989. |
| POP1 | POP1 | Protein | 100.5 | 9.84 | POP1 | POP1 | Lygerou et al., 1994; Lygerou et al., 1996. |
| POP3 | POP3 | Protein | 22.6 | 9.57 | ...... | ...... | Dichtl and Tollervey, 1997. |
| POP4 | POP4 | Protein | 32.9 | 9.26 | RPP29 | RPP29 | Jarrous et al., 1999a; Chu et al., 1997; Van Enennaam et al., 1999. |
| POP5 | POP5 | Protein | 19.6 | 7.79 | ...... | ...... | Chamberlain et al., 1998. |
| POP6 | POP6 | Protein | 18.2 | 9.28 | ...... | ...... | Chamberlain et al., 1998. |
| POP7b | POP7b | Protein | 15.8 | 9.34 | RPP20 | RPP20 | Stolc et al., 1998; Chamberlain et al., 1998. |
| POP8 | POP | Protein | 15.5 | 4.57 | ...... | ...... | Chamberlain et al., 1998. |
| RPP1 | RPP1 | Protein | 32.2 | 9.76 | RPP30 | RPP30 | Eder et al., 1997; Stolc and Altman, 1997. |
| RPR2 | Protein | 16.3 | 9.99 | ...... | ...... | Chamberlain et al., 1998. | |
| ...... | SNM1 | Protein | 22.5 | 9.81 | ...... | ...... | Schmitt and Clayton, 1994. |
| ...... | ...... | Protein | ...... | ...... | REP38c | ...... | Eder et al., 1997. |
| ...... | ...... | Protein | ...... | ...... | RPP40c | ...... | Jarrous et al., 1998. |
| ...... | ...... | Protein | ...... | ...... | RPP25c | ...... | Eder et al., 1997. |
| ...... | ...... | Protein | ...... | ...... | RPP14c | ...... | Jarrous et al., 1999a. |
Molecular weights are presented only for the yeast RNA and protein subunits. pis are presented only for the yeast protein subunits.
POP7 is also known as RPP2 (Stolc et al., 1998).
The relationship of these human proteins to the yeast protein subunits is currently unclear.
The RNA subunit
The RPR1 RNA gene is transcribed by RNA polymerase III (Lee et al., 1991b), and is essential for cell viability and pre-tRNA processing (Lee and Engelke, 1989; Lee et al., 1991a). The 486 nucleotide primary transcript (pre-RPR1 RNA) undergoes a single cleavage that removes the 84-nucleotide 5′ leader and a series of cleavages to remove the 33-nucleotide 3′ trailer sequence (Lee et al., 1991a). Ribonucleoprotein complexes containing the pre-RPR1 RNA and mature RPR1 RNA behave similarly during most of the purification steps (Chamberlain et al., 1996) and for the most part both RNAs can be immunoprecipitated using affinity tags on the protein subunits (Lygerou et al., 1994; Chu et al., 1997; Dichtl and Tollervey, 1997; Stolc and Altman, 1997; Chamberlain et al., 1998). These observations imply that the pre-RPR1 RNA is processed after the formation of ribonucleoprotein complexes that are similar to those formed in the mature holoenzyme.
A secondary structure of RPR1 RNA has been proposed based on phylogenetic studies and structure-sensitive RNA footprinting, and is shown in Figure 1 (Tranguch et al., 1994; Chen and Pace, 1997; Frank et al., 2000). Comparison of the RPR1 RNA structure to the bacterial consensus structure and other eukaryotic RNase P RNA structures reveals both similarities and differences. A core structure is conserved among RNase P RNA from different species (Chen and Pace, 1997; Pitulle et al., 1998; Frank et al., 2000). It contains five ‘critical regions’ carrying conserved nucleotides (CR I-V) and several helices (P1, P2, P3, P4 and P10/P11) at similar positions in different RNA structures. The P4 helix formed by base-pairing between CR I and CR V sequences, is postulated to be the catalytic center of the bacterial enzyme as suggested by site-directed mutagenesis, phosphorothioate substitution, and deletional analysis (Kazakov and Altman, 1991; Darr et al., 1992; Altman et al., 1993; Guerrier-Takada and Altman, 1993; Haas et al., 1994; Harris and Pace, 1995; Pagan-Ramos et al., 1996a).
Fig. 1.
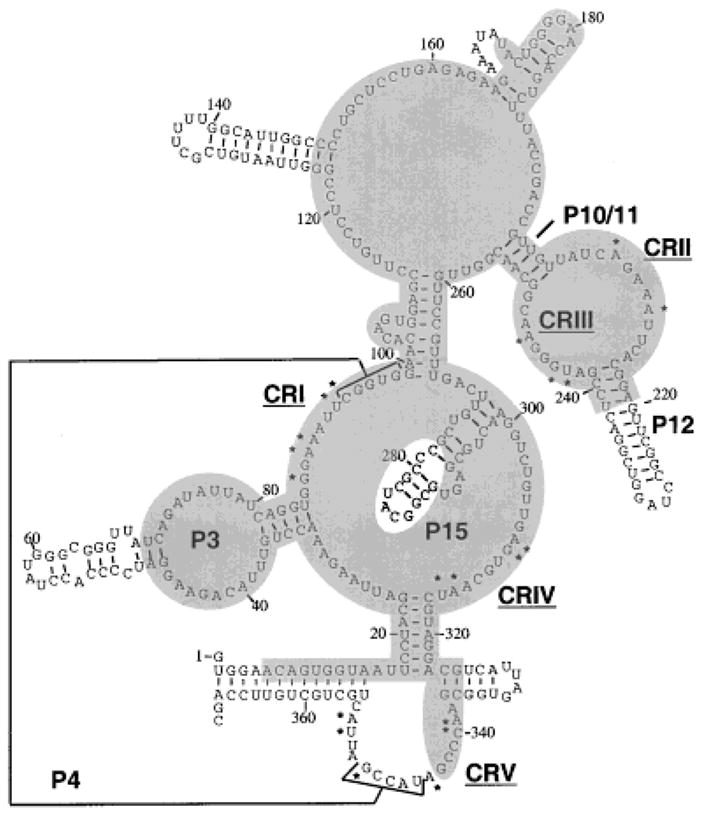
Predicted secondary structure of yeast nuclear RNase P RNA. Nucleotides that are most conserved in bacterial, archaeal and eukaryotic RNase P RNAs are marked with asterisks (Frank et al., 2000). Five critical regions (“CR”s) are numbered according to the Chen and Pace nomenclature (Chen and Pace, 1997), and correspond to clusters of the most conserved positions. “P” represents helical regions, with numbers assigned based on corresponding bacterial structures (Haas et al., 1994). The P4 helix is marked with brackets connected by a line. Nucleotides within the shaded regions are protected from nuclease attack in the holoenzyme compared to deproteinized RNA (Tranguch et al., 1994). For technical reasons it is not known whether the last 26 nucleotides are protected by protein in the holoenzyme. (Figure adapted from Tranguch et al., 1994).
CRII and CRIII, together with P10/11 and P12, form a domain whose function and accessibility to chemical reagents is magnesium-dependent in yeast (Pagan-Ramos et al., 1996b; Ziehier et al., 1998). Minimization experiments with the yeast RNase P RNA have shown that the helix P12 is dispensable in vivo, while the internal loop containing the conserved CAGARA (R represents purine) sequence is essential for viability and RNase P activity (Pagan-Ramos et al., 1994). Mutagenesis studies of the CAGAAA sequence in CR II of the RPR1 RNA (positions 206–211) indicate a function in magnesium utilization that primarily affects kcat of the holoenzyme (Pagan-Ramos et al., 1996b; Ziehler et al., 1998). Previous studies on bacterial RNase P RNA showed that substrate binding requires divalent cation-mediated interactions with the conserved loop, and that P10/11-P12 domain is subject to lead-induced hydrolysis (Ciesiolka et al., 1994) and Mg2+ cleavage at high pH (Kazakov and Altman, 1991). Thus, CR II and CR III appear to be positioned close to the active site of RNase P, facilitating pre-tRNA binding or catalysis.
The precise functions of CR IV in substrate binding and catalysis have not been delineated in any detail. Extensive mutagenesis of this region in yeast RPR1 RNA gave defects in both KM and kcat for the holoenzyme (Pagan-Ramos et al., 1996a).
RNA structural elements distinct to the bacterial or nuclear RNase P RNA subunits also exist, the most obvious being the P3 and P15 regions. Although RNAs from both kingdoms have the P3 region, the eukaryotic P3 helix is more complex than the bacterial counterpart (Fig. 1). The eukaryotic P3 region is composed of a helix-loop-helix structure as opposed to the continuous stem structure in the bacterial P3 (Schmitt et al., 1993; Tranguch and Engelke, 1993; Paluh and Clayton, 1995; Pitulle et al., 1998; Schmitt, 1999). The variation in structure is consistent with the possible functions of P3 in eukaryotes. In human cells, the P3 loop is required for the proper subcellular localization of the RNase P RNA (Jacobson et al., 1997), and is shown to be a binding site for proteins (Reddy et al., 1983; Liu et al., 1994; Eder et al., 1997). In the case of bacteria, the absence of an internal loop in P3 might be related to the lack of subcellular compartments. On the other hand, both the bacterial and eukaryotic P3 appear to be important for the assembly of RNase P holoenzymes. P3 was proposed to interact with the protein subunit in E. coli holoenzyme (Morse and Schmidt, 1993). In yeast RNase P, a mutated P3 loop causes the accumulation of pre-tRNA, and the failure to produce RPR1 RNA in vivo (Ziehler et al., 2000b).
In contrast to P3, P15 region is more complex in bacteria than in yeast, and is completely lost in human RNase P RNA. The function of P15 region is more pronounced in the RNase P ribozyme reaction than with the holoenzyme. An internal loop in the bacterial P15 stem contributes to substrate (and product) binding by base-pairing to the 3′ CCA of the substrate pre-tRNA (Kirsebom and Svard, 1994; Oh and Pace, 1994), and also binds catalytic metal ions (Burgin and Pace, 1990; Kufel and Kirsebom, 1996, 1998).
The presence of the conserved catalytic core structure of bacterial RNase P RNA (critical regions I–V) in the eukaryotic nuclear RNA subunits strongly suggests that the nuclear RNA is serving functions similar to the bacterial RNA subunit, even though some subdomains vary among the kingdoms. Affirmation of the essential nature of the critical regions in the yeast RPR1 RNA by mutagenesis (Pagan-Ramos et al., 1996a,b) confirms that the extensive protein contact has not supplanted these functions.
The Protein Subunits
A most striking difference between the bacterial and eukaryotic RNase P is the protein content. The bacterial RNase P holoenzyme contains only one protein subunit with the molecular weight of 14 kDa. In yeast, the molecular weight of the nine protein subunits ranges from 100.5 to 15.5 kDa and seven of the proteins are basic, with pIs higher than 9 (Table 1). The only acidic protein in RNase P is Pop8p, with a pI of 4.5. All nine proteins are essential for life and for RNase P activity (Lygerou et al., 1994; Chu et al., 1997; Dichtl and Tollervey, 1997; Stolc and Altman, 1997; Chamberlain et al., 1998), but little is known about what these proteins do. Possible multiple functions will be considered in a later section.
The yeast and human RNase P holoenzymes have similar subunit complexities (Table 1). Human RNase P from HeLa cells contains an RNA subunit, H1 RNA (Bartkiewicz et al, 1989), and at least seven protein subunits that co-purify with the enzymatic activity, namely Rpp14p, Rpp20p, Rpp25p, Rpp29p, Rpp30p, Rpp38p, and Rpp40p (Chu et al., 1997; Eder et al., 1997; Stoic and Altman, 1997; Chamberlain et al., 1998; Jarrous et al., 1998; Stolc et al., 1998; Jarrous et al., 1999a; van Eenennaam et al., 1999). In addition, the human homolog of yeast Pop1p (hPop1p) was discovered independently by database searches, although the overall sequence similarity between the two proteins is low (Lygerou et al., 1996). Since hPop1p antibody can immunoprecipitate the H1 RNA and RNase P activity from HeLa cell extracts, hPop1p is likely to be an integral component of human RNase P. Database searches have also revealed the human homologs of several other yeast RNase P proteins (see Table 1).
SUBSTRATE RECOGNITION BY YEAST RNase P
RNase P appears to be responsible for the 5′ end maturation of all the pre-tRNAs in a cell. The diversity of the sequence in tRNA genes requires the recognition of common structural features in pre-tRNAs. Nuclear pre-tRNAs contain 5′-leaders, the mature tRNA domains, 3′-trailer sequences, and sometimes introns. Sequence comparisons have not shown any common structure or sequence in the 5′-leader of the pre-tRNAs (reviewed in Lee et al., 1997; Frank and Pace, 1998) although the nature of the eukaryotic nuclear transcription unit tends to result in purine-rich 5′ leaders. Interestingly, synthetic substrates with a 5′-leader as small as a single nucleotide can be processed by both bacterial and yeast RNase P (Kline et al., 1981; Surratt et al., 1990; Smith and Pace, 1993; Ziehler et al., 2000a). These data suggest that any contribution of the 5′-leader to substrate binding is not critical.
Studies on bacterial and eukaryotic RNase P have demonstrated that essential recognition determinants are located in the consensus tertiary structure of the mature tRNA. RNase P reactions are strongly inhibited by the presence of mature tRNA products (Reich et al., 1988; Smith et al., 1992; Beebe and Fierke, 1994; Ziehler et al., 2000a). The use of small model substrates further delineates the minimal determinants to the TC and acceptor stems for bacterial RNase P. Bacterial enzymes cleave an RNA substrate that consists only of these two helices, although the KM is much higher than that of the natural substrates (McClain et al., 1987; Hardt et al., 1993; Schlegl et al., 1994). Yeast nuclear RNase P, in contrast, cannot efficiently recognize the RNA mini-helix substrate (J.Y. Lee and D.R. Engelke, unpublished results). The contact points in the mature tRNA domain for eukaryotic RNase P recognition have not yet been defined, although an intact tertiary structure is required (Hardt et al., 1993; Levinger et al., 1995).
In addition to the tertiary structure of the mature tRNA, 3′-trailer sequences also participate in binding to RNase P. Although there is no universal 3′-trailer element, there are conserved 3′-trailing sequences within individual kingdoms. In bacteria, a 3′-CCA sequence is immediately downstream of the aminoacyl stem in many tRNA primary transcripts. The substrate 3′-CCA sequence interacts with the internal loop of the P15 region of the bacterial RNase P RNA. Unlike the bacterial pre-tRNAs, eukaryotic pre-tRNAs have a polyuridine (poly-U) stretch as part of the trailing sequence because poly-U is the terminator for pre-tRNA transcription by RNA polymerase III. The P15 loop of the RNase P RNA is substantially reduced or missing in eukaryotes, but the 3′-trailer in yeast pre-tRNA still appears to be seen by the nuclear holoenzyme. It has been shown recently that a pre-tRNA containing single-stranded 3′-trailer sequences binds to the yeast RNase P more strongly than the mature tRNA (Ziehier et al., 2000a). It is not yet known whether the 3′ trailer interacts with an unidentified segment of the RNase P RNA subunit or one of the protein subunits.
The polyuridine patch at the 3′ end of nuclear pre-tRNAs has been shown to interact with the 5′-leader via base-pairing (Lee et al., 1997). In order for pre-tRNA to be efficiently processed by RNase P, a mismatched bulge at the −1/+1 position (relative to the cleavage site) is required, presumably to help destabilize any extended helix between the 5′ leader and 3′ trailer. Continuous extension of the aminoacyl stem into the leader and trailer reduces or abolishes pre-tRNA processing both in vivo and in vitro, affecting both the KM and kcat of cleavage by RNase P (Lee et al., 1997). It is possible that the leader/trailer stem structure melting is somehow facilitated in vivo, generating single-stranded 5′-leader and/or 3′-trailer for the recognition by RNase P. For example, some protein component, like La antigen (Lhp1p in S. cerevisiae), might bind to the 3′-trailer in vivo (Van Horn et al., 1997; Yoo and Wolin, 1997), generating an RNP complex as the physiological substrate for RNase P. Alternatively, the rate at which pre-tRNAs proceed through the early processing pathway could, to some degree, be controlled by the leader/trailer stem strength. There is also evidence that single-stranded 5′-leader sequences can modestly improve the substrate binding to the yeast nuclear RNase P (Ziehier et al., 2000a).
It is currently unclear what part of the yeast nuclear RNase P interacts with the 5′-leader or the 3′-trailer. The protein subunit of B. subtilis RNase P possesses a single-stranded RNA binding cleft that interacts with the unpaired 5′-leader (Crary et al., 1998; Kurz et al., 1998; Niranjanakumari et al., 1998; Stams et al., 1998). Considering that seven of the nuclear protein subunits are rich in basic amino acids and at least six have non-specific RNA binding capabilities, the additional protein content of the nuclear holoenzyme could provide the capacity to interact with multiple sites in pre-tRNAs (or other substrates).
Pre-tRNA is the only known physiological substrate for eukaryotic RNase P to date. In the case of bacterial RNase P, the natural substrates include pre-tRNAs, precursors to 4.55 rRNA (Bothwell et al., 1976), 10Sa RNA (Komine et al., 1994), and the polycistronic his operon mRNA (Alifano et al., 1994). The bacterial RNase P protein plays an important role in broadening the substrate specificity of the holoenzyme. Several mutations in the protein subunit block the cleavage of pre-4.5S RNA, but not pre-tRNA (Gopalan et al., 1997). Thus, with the increased amount of protein content, eukaryotic nuclear RNase P may also have other non-tRNA substrates in vivo. No such alternative nuclear substrates have been definitely identified to date, although individual mRNA or other RNA cleavages could easily have been overlooked. One interesting observation in this regard is that a temperature-sensitive mutation in the RNA subunit of yeast nuclear RNase P leads to accumulation of a pre-rRNA intermediate in vivo (Chamberlain et al., 1996). However, the sites that the nuclear RNase P cleaves in naked pre-rRNA substrates do not correspond in any obvious way to the processing intermediate that accumulates in vivo, so the effect may be indirect rather than suggesting pre-rRNA is a direct substrate for RNase P in vivo.
RELATIONSHIP BETWEEN RNase P AND RNase MRP
Genetic and biochemical studies have indicated that RNase P is closely related to another ribonucleoprotein, RNase MRP, which has only been found in eukaryotes. RNase MRP was first discovered as an RNA-containing enzyme that was proposed to cleave mitochondrial RNA primers for DNA replication (Chang and Clayton, 1987). Later the majority of RNase MRP RNA was found to be in the nucleolus, where the enzyme plays an important role in pre-rRNA maturation (reviewed in Reilly and Schmitt, 1995). Whether there are two pools of RNase MRP RNA, one in the nucleolus and one in the mitochondria, and whether these pools associate with the same protein subunits, needs further investigation.
Nuclear RNase P and nucleolar RNase MRP show a significant degree of overlap in subunit composition. Structural analyses including phylogenetic studies provide a working structural model for RNase MRP RNA (Forster and Altman, 1990; Schmitt et al., 1993; Schmitt, 1999). Comparison of the secondary structures of yeast nuclear RNase P and MRP RNAs has revealed a clear evolutionary relatedness. The two share at least one strikingly similar subdomain not found in bacterial RNase P RNA–the P3 internal loop, as well as the CR I, CRIV, and CRV regions conserved in all RNase P RNAs. At this time it is not clear whether equivalents of the CR II or CRIII subdomains will also be found in RNase MRP RNA.
The P3 region is of interest because it has been shown to be needed for protein binding in human RNase P and MRP (Yuan et al., 1991; Liu et al., 1994). Moreover, the P3 internal loop sequence is better conserved between RNase P and MRP within the same organism than it is among RNase Ps from different eukaryotic species. Recently, it has been demonstrated that the P3 regions in the yeast nuclear RNases P and MRP are functionally equivalent (Lindahl et al., 2000; Shadel et al., 2000; Ziehler et al., 2000b), suggesting interactions with protein subunit(s) common to the two enzymes that has co-evolved with the P3 domain. Several studies have implicated different proteins as requiring P3 for binding or being close enough to P3 to allow cross-linking (Liu et al., 1994; Pluk et al., 1999). However, recent data from this laboratory indicated that Pop1p is probably the subunit that specifically recognizes P3(Ziehler et al., 2000b).
Eight of the nine protein subunits of the yeast nuclear RNase P also appear to associate with RNase MRP (Table 1). Extensive protein subunit overlap has been seen between human RNases P and MRP as well (Lygerou et al., 1996; Jarrous et al., 1999a; Pluk et al., 1999; van Eenennaam et al., 1999), although the characterization of protein subunits is not yet complete. In yeast, each enzyme has at least one unique protein subunit. Rpr2p seems to associate only with RNase P, while Snm1p appears distinctive to RNase MRP (Schmitt and Clayton, 1994; Chamberlain et al., 1998). It remains unclear whether the differences in the RNA subunits, the unique protein subunit, or both are required for the substrate specificity of each enzyme. Despite the different substrate specificity, these two enzymes catalyze a common reaction – hydrolysis of a phosphodiester bond leaving a 3′-hydroxyl and a 5′-phosphate. RNase P and RNase MRP are therefore likely to be descendents of a single ancestral enzyme. In retrospect, this makes a great deal of sense. In bacteria, RNase P often serves as a pre-rRNA processing enzyme because the rRNA subunits in the large transcription unit are often separated by a pre-tRNA. When the pre-tRNA is cleaved by RNase P the large bacterial rRNA subunits are separated. In eukaryotes, RNase MRP cuts in the internal transcribed spacer 1 (ITS 1) region, separating the two large rRNA subunits in nuclear primary transcripts.
No extensive study has been done to identify all the substrates and functions of RNases P and MRP in eukaryotes. However, a recent report has linked the yeast RNase MRP deficiency to plasmid segregation (Cai et al., 1999). Considering the high degree of similarity in the composition of RNase P and MRP, it would not be surprising that eukaryotic RNase P also has essential functions in addition to pre-tRNA maturation.
LOCALIZATION OF NUCLEAR RNase P
Eukaryotes have multiple defined subcellular compartments. Most ribosome biosynthesis, including processing of pre-rRNA transcripts by RNase MRP, takes place in dense, subnuclear regions termed nucleoli. It was previously assumed that pre-tRNA biosynthesis, including RNase P cleavage, would be distributed throughout the nucleoplasm, since tRNA genes are distributed throughout the linear genome maps of eukaryotes. Recent studies have suggested this might not be the case.
The exact location of nuclear RNase P in metazoans is controversial. Studies on the localization of the RNA subunit have suggested that some of the nuclear RNase P resides in the perinucleolar compartment (PNC) on the surface of some nucleoli (Matera et al., 1995; Lee et al., 1996). By the use of slightly different methods, the majority of nuclear RNase P in HeLa cells was seen as nucleoplasmic, with only transient localization to the nucleolus (Jacobson et al., 1997). It has recently been shown that some of the protein subunits of human RNase P are localized in the nucleolus of cultured mammalian cells (Jarrous et al., 1999b). However, since these proteins might also be integral subunits of human RNase MRP in the nucleolus, it is difficult to assess what proportion of the nucleolar signal is from RNase MRP, as opposed to RNase P.
In the yeast S. cerevisiae, the 5′-leader cleavage of pre-tRNAs by RNase P is an early step in the nuclear tRNA maturation pathway. Using fluorescence in situ hybridization, nuclear pre-tRNAs, as well as the RNase P RNA, were shown to be primarily nucleolar with additional foci in the nucleoplasm (Bertrand et al., 1998) (Fig. 2). The results suggest that the early events in tRNA maturation pathway are compartmentalized rather than distributed evenly in the nucleoplasm. The co-localization of much of the pre-tRNA pathway with nucleoli may reflect the possible interplay between tRNA and rRNA biogenesis.
Fig. 2.
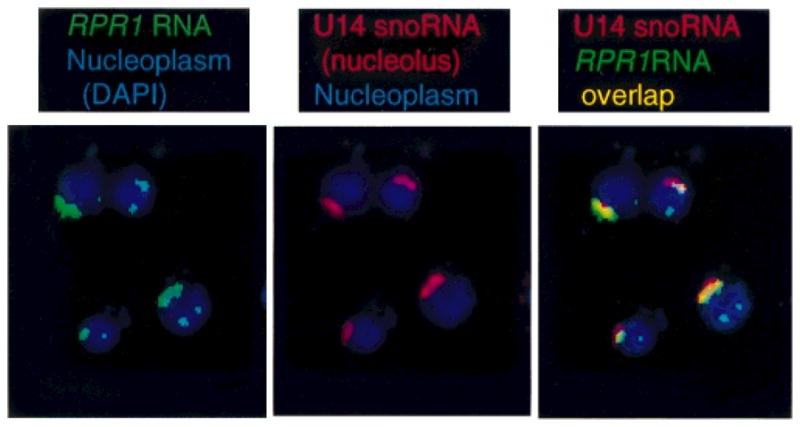
Subnuclear localization of yeast nuclear RNase P. A fluorescent oligonucleotide probe complementary to the RPR1 RNA subunit (green) was hybridized to fixed, permeabilized yeast. Yeast were stained with DAPI (blue in aH panels) to show the nucleoplasm and a fluorescent oligonucleotide to a small nucleolar RNA (U14, red in the second and third panels). The DAPI nucleoplasm signal and position of U14 in the nucleolus are as expected. The RPRJ RNA probe localizes more than 70% in or near the nucleolus, with secondary punctate loci that are somewhat variable among strains. Cytoplasmic features are not visible with these fluorescent tags. (Figure adapted from Bertrand et al., 1998).
The fact that both RNase P and RNase MRP are nucleolar enzymes in yeast suggests that one or more of the eight shared protein subunits might provide a means for subnuclear localization of both enzymes. Studies on human RNases MRP and P have shown that the P3 region, a common subdomain in RNase P and MRP RNA, is required for the nuclear localization of the RNA subunits (Jacobson et al., 1995, 1997). Since P3 is required for specific binding of several protein subunits to the RNA subunit, mislocalization when P3 is deleted is likely to be a result of protein subunit loss, rather than direct localization signals in the P3 RNA subdomain. Which of the proteins might specify subnuclear localization is unknown at this time.
POSSIBLE FUNCTIONS OF THE PROTEIN SUBUNITS OF EUKARYOTIC RNase P
Little is known about the function of the protein subunits in eukaryotic RNase P. The nine polypeptides in yeast nuclear RNase P bear no obvious resemblance to the single, small bacterial protein. RNA footprinting of the yeast holoenzyme has shown that most of the RNA subunit is covered by proteins, although the critical regions are accessible to small, chemical probes (Tranguch et al., 1994). Why are there so many protein subunits in eukaryotic holoenzymes? Depletion of any protein subunit of yeast RNase P except Pop3p reduces or eliminates the ability to produce RNase P with a mature-size RPR1 RNA subunit (Lygerou et al., 1994; Chu et al., 1997; Dichtl and Tollervey, 1997: Stolc and Altman, 1997; Chamberlain et al., 1998). This might reflect incorrect assembly of the yeast holoenzyme since, as noted above, the RNA subunit seems to be assembled into a ribonucleoprotein similar to the mature form before the RNA subunit is fully processed. Thus, the absolute requirement for all nine protein subunits to assemble viable RNase P in vivo might simply reflect bad assembly in the absence of any one protein, but might also suggest that some important functions have migrated from the bacterial ribozyme to eukaryotic protein subunits.
Inhibition experiments of RNase P with different synthetic pre-tRNAs show that the nuclear holoenzyme binds far more aggressively to single-stranded RNAs than the bacterial holoenzyme. This applies to both pre-tRNA substrates with single-stranded termini and other non-specific, single-stranded RNAs. Some protein subunits might interact with the 5′-leader, 3′-trailer, or other regions of the pre-tRNAs (e.g., the anticodon stem). It is also possible that one or more subunits are specifically needed to interact with entirely different substrate RNAs that are yet to be discovered. As noted above, the bacterial protein is responsible in part for substrate discrimination, and it would be expected that the nuclear enzyme proteins also fulfill this role. In addition, the physiological substrates of RNase P in nuclei might well be RNP complexes rather than naked RNAs. In this case, the huge increased eukaryotic substrate complexity might discriminate among a variety of protein, as well as RNA determinants for efficient substrate acquisition.
The possibility should be noted that some proteins might act to prevent interaction with inappropriate substrates. We previously determined that the highly purified nuclear holoenzyme, unlike the bacterial ribozyme, could efficiently cleave deproteinized pre-rRNA substrates at a large number of sites that are not likely to have physiological significance (Chamberlain et al., 1996). The protein subunits might well function within the cell to prevent the holoenzyme from seeing inappropriate RNAs, either through subnuclear sequestration or rejection of inappropriate interactions with nuclear RNP complexes.
The existence of subcellular compartments is a distinctive feature in eukaryotic cells, and yeast nuclear RNase P is found in subnuclear foci, primarily the nucleolus. In addition to more biochemical functions, some proportion of the large RNase P/RNase MRP protein content almost certainly specifies location of the enzyme within the spatially organized pre-tRNA and pre-rRNA biosynthetic pathways. The molecular interactions that lead to this spatial organization are currently under investigation.
CONCLUSIONS AND FUTURE DIRECTIONS
RNase P is an ancient RNA enzyme that has been found in Bacteria, Archaea and Eukarya. Although one essential function of the enzymes remains the same, to mature the 5′ end of tRNA, it has evolved from an active bacterial ribozyme to a protein-dependent eukaryotic holoenzyme. Substantial progress has been made in understanding the bacterial RNase P reaction, predominantly the ribozyme-catalyzed reaction. With the discovery of a more complex eukaryotic RNase P composition, the involvement of the protein subunits in RNase P activity has raised several interesting issues. Current knowledge of RNase P suggests that the protein subunits are required for the assembly, efficient substrate recognition, and subcellular localization of the holoenzyme. Other new functions of the protein subunits are expected to eventually emerge.
In bacteria, RNase P cleaves not only pre-tRNAs but also other small RNAs, ribosomal RNA precursors, and an unknown number of messenger RNAs. In eukaryotes, pre-tRNA is currently the only known substrate for RNase P. The sibling enzyme, RNase MRP, has taken over the function of rRNA processing, but no systematic search has been made to determine whether there are other, non-tRNA substrates for RNase P in eukaryotes. The complexity of the nuclear RNase P suggests a variety of additional functions might await discovery.
Acknowledgments
Contract grant sponsor: NIH; Contract grant number: GM34869.
We thank Edouard Bertrand and Robert Singer for their collaboration on the localization of yeast nuclear RNase P, and Drs. Daniel Frank and Norman Pace for communicating results prior to publication. Felicia Houser-Scott was a fellow of the UNCF-Merck Postdoctoral Science Initiative and the Ford Foundation.
LITERATURE CITED
- Akaboshi E, Guerrier-Takada C, Altman S. Veal heart ribonuclease P has an essential RNA component. Biochem Biophys Res Commun. 1980;96:831–837. doi: 10.1016/0006-291x(80)91430-8. [DOI] [PubMed] [Google Scholar]
- Alifano P, Rivellini F, Piscitelli C, Arraiano CM, Bruni CB, Carlomagno MS. Ribonuclease E provides substrates for ribonuclease P-dependent processing of a polycistronic mRNA. Genes Dev. 1994;8:3021–3031. doi: 10.1101/gad.8.24.3021. [DOI] [PubMed] [Google Scholar]
- Altman S, Gopalan V, Vioque A. Varieties of RNase P: a nomenclature problem? RNA. 2000;6:1689–1694. doi: 10.1017/s1355838200001783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Altman S, Wesolowski D, Puranam RS. Nucleotide sequences of the RNA subunit of RNase P from several mammals. Genomics. 1993;18:418–422. doi: 10.1006/geno.1993.1488. [DOI] [PubMed] [Google Scholar]
- Arends S, Schon A. Partial purification and characterization of nuclear ribonuclease P from wheat. Eur J Biochem. 1997;244:635–645. doi: 10.1111/j.1432-1033.1997.t01-1-00635.x. [DOI] [PubMed] [Google Scholar]
- Ban N, Nissen P, Hansen J, Moore PB, Steitz TA. The complete atomic structure of the large ribosomal subunit at 2.4 A resolution. Science. 2000;289:905–920. doi: 10.1126/science.289.5481.905. [DOI] [PubMed] [Google Scholar]
- Bartkiewicz M, Gold H, Altman S. Identification and characterization of an RNA molecule that copurifies with RNase P activity from HeLa cells. Genes Dev. 1989;3:488–499. doi: 10.1101/gad.3.4.488. [DOI] [PubMed] [Google Scholar]
- Beebe JA, Fierke CA. A kinetic mechanism for cleavage of precursor tRNA(Asp) catalyzed by the RNA component of Bacillus subtilis ribonuclease P. Biochemistry. 1994;33:10294–10304. doi: 10.1021/bi00200a009. [DOI] [PubMed] [Google Scholar]
- Beebe JA, Kurz JC, Fierke CA. Magnesium ions are required by Bacillus subtilis ribonuclease P RNA for both binding and cleaving precursor tRNAAsp. Biochemistry. 1996;35:10493–10505. doi: 10.1021/bi960870m. [DOI] [PubMed] [Google Scholar]
- Bertrand E, Houser-Scott F, Kendall A, Singer RH, Engelke DR. Nucleolar localization of early tRNA processing. Genes Dev. 1998;12:2463–2468. doi: 10.1101/gad.12.16.2463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bothwell ALM, Garber RL, Altman S. Nucleotide sequence and in vitro processing of a precursor molecule to Escherichia coli 4.5 S RNA. J Biol Chem. 1976;251:7709–7716. [PubMed] [Google Scholar]
- Burgin AB, Pace NR. Mapping the active site of ribonuclease P RNA using a substrate containing a photoaffinity agent. EMBO J. 1990;9:4111–4118. doi: 10.1002/j.1460-2075.1990.tb07633.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cai T, Reilly TR, Cerio M, Schmitt ME. Mutagenesis of SNM1, which encodes a protein component of the yeast RNase MRP, reveals a role for this ribonucleoprotein endoribonuclease in plasmid segregation. Mol Cell Biol. 1999;19:7857–7869. doi: 10.1128/mcb.19.11.7857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carrara G, Calandra P, Fruscoloni P, Tocchini-Valentini GP. Two helices plus a linker: a small model substrate for eukaryotic RNase P. Proc Natl Acad Sci USA. 1995;92:2627–2631. doi: 10.1073/pnas.92.7.2627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carter AP, Clemons WM, Brodersen DE, Morgan-Warren RJ, Wimberly BT, Ramakrishnan V. Functional insights from the structure of the 30S ribosomal subunit and its interactions with antibiotics. Nature. 2000;407:340–348. doi: 10.1038/35030019. [DOI] [PubMed] [Google Scholar]
- Cate JH, Yusupov MM, Yusupova GZ, Earnest TN, Noller HF. X-ray crystal structures of 70S ribosome functional complexes. Science. 1999;285:2095–2104. doi: 10.1126/science.285.5436.2095. [DOI] [PubMed] [Google Scholar]
- Cech TR. Structure and mechanism of the large catalytic RNAs: group I and group II introns and ribonuclease P. In: Gesteland RF, Atkins JF, editors. The RNA World. New York: CSHL Press; 1993. pp. 239–269. [Google Scholar]
- Chamberlain JR, Lee Y, Lane WS, Engelke DR. Purification and characterization of the nuclear RNase P holoenzyme complex reveals extensive subunit overlap with RNase MRP. Genes Dev. 1998;12:1678–1690. doi: 10.1101/gad.12.11.1678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chamberlain JR, Pagan R, Kindelberger DW, Engelke DR. An RNase P RNA subunit mutation affects ribosomal RNA processing. Nucleic Acids Res. 1996;24:3158–3166. doi: 10.1093/nar/24.16.3158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang DD, Clayton DA. A novel endoribonuclease cleaves at a priming site of mouse mitochondrial DNA replication. EMBO J. 1987;6:409–417. doi: 10.1002/j.1460-2075.1987.tb04770.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen JL, Pace NR. Identification of the universally conserved core of ribonuclease P RNA (letter) RNA. 1997;3:557–560. [PMC free article] [PubMed] [Google Scholar]
- Chu S, Zengel JM, Lindahl L. A novel protein shared by RNase MRP and RNase P. RNA. 1997;3:382–391. [PMC free article] [PubMed] [Google Scholar]
- Ciesiolka J, Hardt WD, Schlegl J, Erdmann VA, Hartmann RK. Lead-ion-induced cleavage of RNase P RNA. Eur J Biochem. 1994;219:49–56. doi: 10.1111/j.1432-1033.1994.tb19913.x. [DOI] [PubMed] [Google Scholar]
- Cordier A, Schon A. Cyanelle RNase P: RNA structure analysis and holoenzyme properties of an organellar ribonucleoprotein enzyme. J Mol Biol. 1999;289:9–20. doi: 10.1006/jmbi.1999.2762. [DOI] [PubMed] [Google Scholar]
- Crary SM, Niranjanakumari S, Fierke CA. The protein component of Bacillus subtilis ribonuclease P increases catalytic efficiency by enhancing interactions with the 5′ leader sequence of pre-tRNAAsp. Biochemistry. 1998;37:9409–9416. doi: 10.1021/bi980613c. [DOI] [PubMed] [Google Scholar]
- Dang YL, Martin NC. Yeast mitochondrial RNase P. Sequence of the RPM2 gene and demonstration that its product is a protein subunit of the enzyme. J Biol Chem. 1993;268:19791–19796. [PubMed] [Google Scholar]
- Darr SC, Pace B, Pace NR. Characterization of ribonuclease P from the archaebacterium Sulfolobus solfataricus. J Biol Chem. 1990;265:12927–12932. [PubMed] [Google Scholar]
- Darr SC, Zito K, Smith D, Pace NR. Contributions of phylogenetically variable structural elements to the function of the ribozyme ribonuclease P. Biochemistry. 1992;31:328–333. doi: 10.1021/bi00117a003. [DOI] [PubMed] [Google Scholar]
- Dichtl B, Tollervey D. Pop3p is essential for the activity of the RNase MRP and RNase P ribonucleoproteins in vivo. EMBO J. 1997;16:417–429. doi: 10.1093/emboj/16.2.417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doria M, Carrara G, Calandra P, Tocchini-Valentini GP. An RNA molecule copurifies with RNase P activity from Xenopus laevis oocytes. Nucleic Acids Res. 1991;19:2315–2320. doi: 10.1093/nar/19.9.2315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drainas D. The RNase P of Dictyostelium discoideum. Mol Biol Rep. 1995;22:135–138. doi: 10.1007/BF00988718. [DOI] [PubMed] [Google Scholar]
- Eder PS, Kekuda R, Stolc V, Altman S. Characterization of two scleroderma autoimmune antigens that copurify with human ribonuclease P. Proc Natl Acad Sci USA. 1997;94:1101–1106. doi: 10.1073/pnas.94.4.1101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Forster AC, Altman S. Similar cage-shaped structures for the RNA components of all ribonuclease P and ribonuclease MRP enzymes. Cell. 1990;3:407–409. doi: 10.1016/0092-8674(90)90003-w. [DOI] [PubMed] [Google Scholar]
- Frank DN, Adamidi C, Ehringer M, Pitulle C, Pace N. Phylogenetic-comparative analysis of the eucaryal ribonuclease P RNA. RNA. 2000;6:1895–1904. doi: 10.1017/s1355838200001461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frank DN, Pace NR. Ribonuclease P: unity and diversity in a tRNA processing ribozyme. Annu Rev Biochem. 1998;67:153–180. doi: 10.1146/annurev.biochem.67.1.153. [DOI] [PubMed] [Google Scholar]
- Gardiner K, Pace NR. RNase P of Bacillus subtilis has a RNA component. J Biol Chem. 1980;255:7507–7509. [PubMed] [Google Scholar]
- Gardiner KJ, Marsh TL, Pace NR. Ion dependence of the Bacillus subtilis RNase P reaction. J Biol Chem. 1985;260:5415–5419. [PubMed] [Google Scholar]
- Gegenheimer P. Enzyme nomenclature: functional or structural? RNA. 2000;6:1695–1697. doi: 10.1017/s1355838200001850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gopalan V, Baxevanis AD, Landsman D, Altman S. Analysis of the functional role of conserved residues in the protein subunit of ribonuclease P from Escherichia coli. J Mol Biol. 1997;267:818–829. doi: 10.1006/jmbi.1997.0906. [DOI] [PubMed] [Google Scholar]
- Groom KR, Heyman HC, Steffen MC, Hawkins L, Martin NC. Kluyveromyces lactis SEF1 and its Saccharomyces cerevisiae homologue bypass the unknown essential function, but not the mitochondrial RNase P function, of the S. cerevisiae RPM2 gene. Yeast. 1998;14:77–87. doi: 10.1002/(SICI)1097-0061(19980115)14:1<77::AID-YEA201>3.0.CO;2-P. [DOI] [PubMed] [Google Scholar]
- Guerrier-Takada C, Altman S. Catalytic activity of an RNA molecule prepared by transcription in vitro. Science. 1984;223:285–286. doi: 10.1126/science.6199841. [DOI] [PubMed] [Google Scholar]
- Guerrier-Takada C, Altman S. A physical assay for and kinetic analysis of the interactions between Ml RNA and tRNA precursor substrates. Biochemistry. 1993;32:7152–7161. doi: 10.1021/bi00079a012. [DOI] [PubMed] [Google Scholar]
- Guerrier-Takada C, Gardiner K, Marsh T, Pace N, Altman S. The RNA moiety of ribonuclease P is the catalytic subunit of the enzyme. Cell. 1983;35:849–857. doi: 10.1016/0092-8674(83)90117-4. [DOI] [PubMed] [Google Scholar]
- Haas ES, Brown JW, Pitulle C, Pace NR. Further perspective on the catalytic core and secondary structure of ribonuclease P RNA. Proc Natl Acad Sci USA. 1994;91:2527–2531. doi: 10.1073/pnas.91.7.2527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hardt WD, Schlegl J, Erdmann VA, Hartmann RK. Role of the D arm and the anticodon arm in tRNA recognition by eubacterial and eukaryotic RNase P enzymes. Biochemistry. 1993;32:13046–13053. doi: 10.1021/bi00211a014. [DOI] [PubMed] [Google Scholar]
- Harris ME, Pace NR. Identification of phosphates involved in catalysis by the ribozyme RNase P RNA. RNA. 1995;1:210–218. [PMC free article] [PubMed] [Google Scholar]
- Jacobson MR, Cao L-G, Wang Y-L, Pederson T. Dynamic localization of RNase MRP RNA in the nucleolus observed by fluorescent RNA cytochemistry in living cells. J Cell Biol. 1995;131:1649–1658. doi: 10.1083/jcb.131.6.1649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jacobson MR, Cao LG, Taneja K, Singer RH, Wang YL, Pederson T. Nuclear domains of the RNA subunit of RNase P. J Cell Sci. 1997;110:829–837. doi: 10.1242/jcs.110.7.829. [DOI] [PubMed] [Google Scholar]
- Jarrous N, Eder PS, Guerrier-Takada C, Hoog C, Altman S. Autoantigenic properties of some protein subunits of catalytically active complexes of human ribonuclease P. RNA. 1998;4:407–417. [PMC free article] [PubMed] [Google Scholar]
- Jarrous N, Eder PS, Wesolowski D, Altman S. Rppl4 and Rpp29, two protein subunits of human ribonuclease P. RNA. 1999a;5:153–157. doi: 10.1017/s135583829800185x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jarrous N, Wolenski JS, Wesolowski D, Lee C, Altman S. Localization in the nucleolus and coiled bodies of protein subunits of the ribonucleoprotein ribonuclease P. J Cell Biol. 1999b;146:559–572. doi: 10.1083/jcb.146.3.559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jayanthi GP, Van Tuyle GC. Characterization of ribonuclease P isolated from rat liver cytosol. Arch Biochem Biophys. 1992;296:264–270. doi: 10.1016/0003-9861(92)90571-d. [DOI] [PubMed] [Google Scholar]
- Joyce GF, Orgel LE. Prospects for understanding the origin of the RNA world. In: Gesteland RF, Atkins JF, editors. The RNA world. New York: CSHL Press; 1993. pp. 1–25. [Google Scholar]
- Kassenbrock CK, Gao GJ, Groom KR, Sulo P, Douglas MG, Martin NC. RPM2, independently of its mitochondrial RNase P function, suppresses an ISP42 mutant defective in mitochondrial import and is essential for normal growth. Mol Cell Biol. 1995;15:4763–4770. doi: 10.1128/mcb.15.9.4763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kazakov S, Altman S. Site-specific cleavage by metal ion cofactors and inhibitors of Ml RNA, the catalytic subunit of RNase P from Escherichia coli. Proc Natl Acad Sci USA. 1991;88:9193–9197. doi: 10.1073/pnas.88.20.9193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kirsebom LA, Svard SG. Base pairing between Escherichia coli RNase P RNA and its substrate. EMBO J. 1994;13:4870–4876. doi: 10.1002/j.1460-2075.1994.tb06814.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kline L, Nishikawa S, Soll D. Partial purification of RNase P from Schizosaccharomyces pombe. J Biol Chem. 1981;256:5058–5063. [PubMed] [Google Scholar]
- Knap AK, Wesolowski D, Altman S. Protection from chemical modification of nucleotides in complexes of Ml RNA, the catalytic subunit of RNase P from E coli, and tRNA precursors. Biochimie. 1990;72:779–790. doi: 10.1016/0300-9084(90)90187-l. [DOI] [PubMed] [Google Scholar]
- Komine Y, Kitabatake M, Yokogawa T, Nishikawa K, Inokuchi H. A tRNA-like structure is present in lOSa RNA, a small stable RNA from Escherichia coli. Proc Natl Acad Sci USA. 1994;91:9223–9227. doi: 10.1073/pnas.91.20.9223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kufel J, Kirsebom LA. Residues in Escherichia coli RNase P RNA important for cleavage site selection and divalent metal ion binding. J Mol Biol. 1996;263:685–698. doi: 10.1006/jmbi.1996.0608. [DOI] [PubMed] [Google Scholar]
- Kufel J, Kirsebom LA. The P15-loop of Escherichia coli RNase P RNA is an autonomous divalent metal ion binding domain. RNA. 1998;4:777–788. doi: 10.1017/s1355838298970923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kurz JC, Niranjanakumari S, Fierke CA. Protein component of Bacillus subtilis RNase P specifically enhances the affinity for precursor-tRNAASP. Biochemistry. 1998;37:2393–2400. doi: 10.1021/bi972530m. [DOI] [PubMed] [Google Scholar]
- Lawrence N, Wesolowski D, Gold H, Bartkiewicz M, Guerrier-Takada C, McClain WH, Altman S. Characteristics of ribonuclease P from various organisms. Cold Spring Harb Symp Quant Biol. 1987;52:233–238. doi: 10.1101/sqb.1987.052.01.028. [DOI] [PubMed] [Google Scholar]
- Lee B, Matera AG, Ward DC, Craft J. Association of RNase mitochondrial RNA processing enzyme with ribonuclease P in higher ordered structures in the nucleolus: A possible coordinate role in ribosome biogenesis. Proc Natl Acad Sci USA. 1996;93:11471–11476. doi: 10.1073/pnas.93.21.11471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee J-Y. PhD dissertation. University of Michigan; Ann Arbor, MI: 1991. Characterization of Saccharomyces cerevisiae nuclear RNase P and its RNA subunit. [Google Scholar]
- Lee J-Y, Engelke DR. Partial characterization of an RNA component that copurifies with Saccharomyces cerevisiae RNase P. Mol Cell Biol. 1989;9:2536–2543. doi: 10.1128/mcb.9.6.2536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee J-Y, Evans CF, Engelke DR. Expression of RNase P RNA in Saccharomyces cerevisiae is controlled by an unusual RNA polymerase III promoter. Proc Natl Acad Sci USA. 1991b;88:6986–6990. doi: 10.1073/pnas.88.16.6986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee J-Y, Rohlman CE, Molony LA, Engelke DR. Characterization of RPR1, an essential gene encoding the RNA component of Saccharomyces cerevisiae nuclear RNase P. Mol Cell Biol. 1991a;11:721–730. doi: 10.1128/mcb.11.2.721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee Y, Kindelberger DW, Lee J-Y, McClennen S, Chamberlain J, Engelke DR. Nuclear pre-tRNA terminal structure and RNase P recognition. RNA. 1997;3:175–185. [PMC free article] [PubMed] [Google Scholar]
- Levinger L, Vasisht V, Greene V, Bourne R, Birk A, Kolla S. Sequence and structure requirements for Drosophila tRNA 5′- and 3′-end processing. J Biol Chem. 1995;270:18903–18909. doi: 10.1074/jbc.270.32.18903. [DOI] [PubMed] [Google Scholar]
- Lindahl L, Fretz S, Epps N, Zengel JM. Functional equivalence of hairpins in the RNA subunits of RNase MRP and RNase P in Saccharomyces cerevisiae. RNA. 2000;6:653–658. doi: 10.1017/s1355838200992574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu MH, Yuan Y, Reddy R. Human RNase P RNA and nucleolar 7-2 RNA share conserved ‘To’ antigen-binding domains. Mol Cell Biochem. 1994;130:75–82. doi: 10.1007/BF01084270. [DOI] [PubMed] [Google Scholar]
- Lutz MS, Ellis SR, Martin NC. Proteasome mutants, pre4-2 and ump1–2, suppress the essential function but not the mitochondrial RNase P function of the Saccharomyces cerevisiae gene RPM2. Genetics. 2000;154:1013–1023. doi: 10.1093/genetics/154.3.1013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lygerou Z, Mitchell P, Petfalski E, Seraphin B, Tollervey D. The POP1 gene encodes a protein component common to the RNase MRP and RNase P ribonucleoproteins. Genes Dev. 1994;8:1423–1433. doi: 10.1101/gad.8.12.1423. [DOI] [PubMed] [Google Scholar]
- Lygerou Z, Pluk H, van Venrooij WJ, Seraphin B. hPopl: an autoantigenic protein subunit shared by the human RNase P and RNase MRP ribonucleoproteins. EMBO J. 1996;15:5936–5948. [PMC free article] [PubMed] [Google Scholar]
- Matera AG, Frey MR, Margelot K, Wolin SL. A perinucleolar compartment contains several RNA polymerase III transcripts as well as the polypyrimidine tract-binding protein, hnRNP I. J Cell Biol. 1995;129:1181–1193. doi: 10.1083/jcb.129.5.1181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McClain WH, Guerrier-Takada C, Altman S. Model substrates for an RNA enzyme. Science. 1987;238:527–530. doi: 10.1126/science.2443980. [DOI] [PubMed] [Google Scholar]
- McKay DB, Wedekind JE. Small Ribozymes. In: Gesteland RF, Cech TR, Atkins JF, editors. The RNA world. 2. New York: CSHL Press; 1999. pp. 265–286. [Google Scholar]
- Morales MJ, Dang YL, Lou YC, Sulo P, Martin NC. A 105-kDa protein is required for yeast mitochondrial RNase P activity. Proc Natl Acad Sci USA. 1992;89:9875–9879. doi: 10.1073/pnas.89.20.9875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morse DP, Schmidt FJ. Suppression of loss-of-function mutations in Escherichia coli ribonuclease P RNA (Ml RNA) by a specific base-pair disruption. J Mol Biol. 1993;230:11–14. doi: 10.1006/jmbi.1993.1120. [DOI] [PubMed] [Google Scholar]
- Niranjanakumari S, Stams T, Crary SM, Christianson DW, Fierke CA. Protein component of the ribozyme ribonuclease P alters substrate recognition by directly contacting precursor tRNA. Proc Nati Acad Sci USA. 1998;95:15212–15217. doi: 10.1073/pnas.95.26.15212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nissen P, Hansen J, Ban N, Moore PB, Steitz TA. The structural basis of ribosome activity in peptide bond synthesis. Science. 2000;289:920–930. doi: 10.1126/science.289.5481.920. [DOI] [PubMed] [Google Scholar]
- Oh BK, Pace NR. Interaction of the 3′-end of tRNA with ribonuclease P RNA. Nucleic Acids Res. 1994;22:4087–4094. doi: 10.1093/nar/22.20.4087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pagan-Ramos E, Lee Y, Engelke DR. Mutational analysis of Saccharomyces cerevisiae nuclear RNase P: randomization of universally conserved positions in the RNA subunit. RNA. 1996a;2:441–451. [PMC free article] [PubMed] [Google Scholar]
- Pagan-Ramos E, Lee Y, Engelke DR. A conserved RNA motif involved in divalent cation utilization by nuclear RNase P. RNA. 1996b;2:1100–1109. [PMC free article] [PubMed] [Google Scholar]
- Pagan-Ramos E, Tranguch AJ, Kindelberger DW, Engelke DR. Replacement of the Saccharomyces cerevisiae RPR1 gene with heterologous RNase P RNA genes. Nucleic Acids Res. 1994;22:200–207. doi: 10.1093/nar/22.2.200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paluh JL, Clayton DA. Schizosaccharomyces pombe RNase MRP RNA is homologous to metazoan RNase MRP RNAs and may provide clues to interrelationships between RNase MRP and RNase P. Yeast. 1995;11:1249–1264. doi: 10.1002/yea.320111305. [DOI] [PubMed] [Google Scholar]
- Pannucci JA, Haas ES, Hall TA, Harris JK, Brown JW. RNase P RNAs from some Archaea are catalytically active. Proc Natl Acad Sci USA. 1999;96:7803–7808. doi: 10.1073/pnas.96.14.7803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pfeiffer T, Tekos A, Warnecke JM, Dramas D, Engelke DR, Seraphin B, Hartmann RK. Effects of phosphorothioate modifications on precursor tRNA processing by eukaryotic RNase P enzymes. J Mol Biol. 2000;298:559–565. doi: 10.1006/jmbi.2000.3655. [DOI] [PubMed] [Google Scholar]
- Pitulle C, Garcia-Paris M, Zamudio KR, Pace NR. Comparative structure analysis of vertebrate ribonuclease P RNA. Nucleic Acids Res. 1998;26:3333–3339. doi: 10.1093/nar/26.14.3333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pluk H, van Eenennaam H, Rutjes SA, Pruijn GJ, van Venrooij WJ. RNA-protein interactions in the human RNase MRP ribonucleoprotein complex. RNA. 1999;5:512–524. doi: 10.1017/s1355838299982079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reddy R, Tan EM, Henning D, Nohga K, Busch H. Detection of a nucleolar 7-2 ribonucleoprotein and a cytoplasmic 8-2 ribonuceoprotein with autoantibodies from patients with scleroderma. J Biol Chem. 1983;258:1383–1386. [PubMed] [Google Scholar]
- Reich C, Olsen GJ, Pace B, Pace NR. Role of the protein moiety of ribonuclease P, a ribonucleoprotein enzyme. Science. 1988;239:178–181. doi: 10.1126/science.3122322. [DOI] [PubMed] [Google Scholar]
- Reilly TH, Schmitt ME. The yeast, Saccharomyces cerevisiae, RNase P/MRP ribonucleoprotein endoribonuclease family. Mol Biol Rep. 1995;22:87–93. doi: 10.1007/BF00988711. [DOI] [PubMed] [Google Scholar]
- Rossmanith W, Karwan RM. Characterization of human mitochondrial RNase P: novel aspects in tRNA processing. Biochem Biophys Res Commun. 1998;247:234–241. doi: 10.1006/bbrc.1998.8766. [DOI] [PubMed] [Google Scholar]
- Schlegl J, Hardt WD, Erdmann VA, Hartmann RK. Contribution of structural elements to Thermus thermophilus ribonuclease P RNA function. EMBO J. 1994;13:4863–4869. doi: 10.1002/j.1460-2075.1994.tb06813.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmitt ME. Molecular modeling of the three-dimensional architecture of the RNA component of yeast RNase MRP. J Mol Biol. 1999;292:827–836. doi: 10.1006/jmbi.1999.3116. [DOI] [PubMed] [Google Scholar]
- Schmitt ME, Bennett JL, Dairaghi DJ, Clayton DA. Secondary structure of RNase MRP RNA as predicted by phylogenetic comparison. FASEB J. 1993;7:208–213. doi: 10.1096/fasebj.7.1.7678563. [DOI] [PubMed] [Google Scholar]
- Schmitt ME, Clayton DA. Characterization of a unique protein component of yeast RNase MRP: An RNA-binding protein with a zinc-cluster domain. Genes Dev. 1994;8:2617–2628. doi: 10.1101/gad.8.21.2617. [DOI] [PubMed] [Google Scholar]
- Shadel GS, Buckenmeyer GA, Clayton DA, Schmitt ME. Mutational analysis of the RNA component of Saccharomyces cerevisiae RNase MRP reveals distinct nuclear phenotypes. Gene. 2000;245:175–184. doi: 10.1016/s0378-1119(00)00013-5. [DOI] [PubMed] [Google Scholar]
- Smith D, Burgin AB, Haas ES, Pace NR. Influence of metal ions on the ribonuclease P reaction. Distinguishing substrate binding from catalysis. J Biol Chem. 1992;267:2429–2436. [PubMed] [Google Scholar]
- Smith D, Pace NR. Multiple magnesium ions in the ribonuclease P reaction mechanism. Biochemistry. 1993;32:5273–5281. doi: 10.1021/bi00071a001. [DOI] [PubMed] [Google Scholar]
- Stams T, Niranjanakumari S, Fierke CA, Christianson DW. Ribonuclease P protein structure: evolutionary origins in the translational apparatus. Science. 1998;280:752–755. doi: 10.1126/science.280.5364.752. [DOI] [PubMed] [Google Scholar]
- Stolc V, Altman S. Rppl, an essential protein subunit of nuclear RNase P required for processing of precursor tRNA and 35S precursor rRNA in Saccharomyces cerevisiae. Genes Dev. 1997;11:2926–2937. doi: 10.1101/gad.11.21.2926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stolc V, Katz A, Altman S. Rpp2, an essential protein subunit of nuclear RNase P, is required for processing of precursor tRNAs and 35S precursor rRNA in Saccharomyces cerevisiae. Proc Natl Acad Sci USA. 1998;95:6716–6721. doi: 10.1073/pnas.95.12.6716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Surratt CK, Lesnikowski Z, Schifman AL, Schmidt FJ, Hecht SM. Construction and processing of transfer RNA precursor models. J Biol Chem. 1990;265:22506–22512. [PubMed] [Google Scholar]
- Talbot SJ, Altman S. Gel retardation analysis of the interaction between C5 protein and Ml RNA in the formation of the ribonuclease P holoenzyme from Escherichia coli. Biochemistry. 1994;33:1399–1405. doi: 10.1021/bi00172a016. [DOI] [PubMed] [Google Scholar]
- Thomas BC. PhD thesis. University of Kansas; Lawrence, KS: 1996. Structural and mechanistic differences between a protein and an RNA-containing RNase P. [Google Scholar]
- Thomas BC, Li X, Gegenheimer P. Chloroplast ribonuclease P does not utilize the ribozyme-type pre-tRNA cleavage mechanism. RNA. 2000a;6:545–553. doi: 10.1017/s1355838200991465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomas BC, Chamberlain J, Engelke DR, Gegenheimer P. Evidence for an RNA-based catalytic mechanism in eukaryotic nuclear ribonuclease P. RNA. 2000b;6:554–562. doi: 10.1017/s1355838200991477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tranguch AJ, Engelke DR. Comparative structural analysis of nuclear RNase P RNAs from yeast. J Biol Chem. 1993;268:14045–14055. [PubMed] [Google Scholar]
- Tranguch AJ, Kindelberger DW, Rohlman CE, Lee J-Y, Engelke DR. Structure-sensitive RNA footprinting of yeast nuclear ribonuclease P. Biochemistry. 1994;33:1778–1787. doi: 10.1021/bi00173a022. [DOI] [PubMed] [Google Scholar]
- van Eenennaam H, Pruijn GJ, van Venrooij WJ. hPop4: a new protein subunit of the human RNase MRP and RNase P ribonucleoprotein complexes. Nucleic Acids Res. 1999;27:2465–2472. doi: 10.1093/nar/27.12.2465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Horn DJ, Yoo CJ, Xue D, Shi H, Wolin SL. The La protein in Schizosaccharomyces pombe: a conserved yet dispensable phosphoprotein that functions in tRNA maturation. RNA. 1997;3:1434–1443. [PMC free article] [PubMed] [Google Scholar]
- Wang MJ, Davis NW, Gegenheimer PA. Novel machanisms for maturation of chloroplast transfer RNA precursors. EMBO J. 1988;7:1567–1574. doi: 10.1002/j.1460-2075.1988.tb02981.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Warnecke JM, Furste JP, Hardt WD, Erdmann VA, Hartmann RK. Ribonuclease P (RNase P) RNA is converted to a Cd(2+)- ribozyme by a single Rp-phosphorothioate modification in the precursor tRNA at the RNase P cleavage site. Proc Natl Acad Sci USA. 1996;93:8924–8928. doi: 10.1073/pnas.93.17.8924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Westhof E, Wesolowski D, Altman S. Mapping in three dimensions of regions in a catalytic RNA protected from attack by an Fe(II)-EDTA reagent. J Mol Biol. 1996;258:600–613. doi: 10.1006/jmbi.1996.0272. [DOI] [PubMed] [Google Scholar]
- Wimberly BT, Brodersen DE, Clemons WM, Jr, Morgan-Warren RJ, Carter AP, Vonrhein C, Hartsch T, Ramakrishnan V. Structure of the 30S ribosomal subunit. Nature. 2000;407:327–339. doi: 10.1038/35030006. [DOI] [PubMed] [Google Scholar]
- Yoo CJ, Wolin SL. The yeast La protein is required for the 3′ endonucleolytic cleavage that matures tRNA precursors. Cell. 1997;89:393–402. doi: 10.1016/s0092-8674(00)80220-2. [DOI] [PubMed] [Google Scholar]
- Yuan Y, Tan E, Reddy R. The 40-kilodalton to autoantigen associates with nucleotides 21 to 64 of human mitochondrial RNA processing/7-2 RNA in vitro. Mol Cell Biol. 1991;11:5266–5274. doi: 10.1128/mcb.11.10.5266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ziehler WA, Day JJ, Fierke CA, Engelke DR. Effects of 5′ leader and 3′ trailer structures on pre-tRNA processing by nuclear RNase P. Biochemistry. 2000a;39:9909–9916. doi: 10.1021/bi000603n. [DOI] [PubMed] [Google Scholar]
- Ziehler WA, Morris J, Houser-Scott F, Millikin CE, Engelke DR. An essential protein-binding domain of nuclear RNase P RNA. RNA. 2000b doi: 10.1017/s1355838201001996. (in press) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ziehler WA, Yang J, Kurochkin AV, Sandusky P0, Zuiderweg ER, Engelke DR. Structural analysis of the P10/11-P12 RNA domain of yeast RNase P RNA and its interaction with magnesium. Biochemistry. 1998;37:3549–3557. doi: 10.1021/bi972886y. [DOI] [PubMed] [Google Scholar]