

Abstract
Matrix metalloproteinases (MMPs) are involved in the pathogenesis of neuroinflammatory diseases (such as multiple sclerosis) as well as in the expansion of malignant gliomas because they facilitate penetration of anatomical barriers (such as the glia limitans) and migration within the neuropil. This review elucidates pathomechanisms and summarizes the current knowledge of the involvement of MMPs in neuroinflammation and glioma, invasion highlighting microglia as major sources of MMPs. The induction of expression, suppression, and multiple pathways of function of MMPs in these scenarios will also be discussed. Understanding the induction and action of MMPs might provide valuable information and reveal attractive targets for future therapeutic strategies.
1. Barriers from Blood to Brain
Influx of inflammatory cells into the neuropil is a hallmark of neuroinflammation (e.g., in multiple sclerosis (MS) [1], and respective mechanisms have been studied extensively in experimental autoimmune encephalomyelitis (EAE), an animal model for multiple sclerosis. Initially, leukocytes migrate across vascular walls and accumulate in the perivascular space. This perivascular “cuffing” [2], however, is only the first step in neuroinflammation because immune cells need to pass the glia limitans and its basement membrane to reach the parenchyma proper in a second, differentially regulated step [3]. While the endothelium does not provide an insurmountable barrier for activated T and B cells under certain (experimental) conditions [4, 5], the glia limitans and the parenchymal basal lamina represent more strictly regulated, secondary barriers [3]. Importantly, clinical symptoms only occur after the penetration of the parenchymal basal lamina (BM), which is formed by a variety of organized extracellular matrix (ECM) components build by astrocytic endfeet of the glia limitans.
There is strong evidence that inducible proteases, known as matrix metalloproteinases (MMPs), are involved in the second step of neuroinflammation [3, 6–10]. The unique features of different, highly specialized, basal laminae rely on their major constituents: collagen IV and laminin predominant are whereas collagen type V, proteoglycans, and glycoproteins are additional constituents [11]. Collagen types IV and V are, unlike other collagens, structurally organized in a nonfibrillar, multilayer network that is resistant to nonspecific proteolytic degradation. Noteworthy is the existence of different laminin isoforms in the specialized basement membrane (BM) of the vessel and the BM of the glia limitans. While the vascular BM exhibits laminin 8 and laminin 10, the BM of the glia limitans is characterized by laminin 1 and laminin 2 [12]. Dystroglycan is a transmembrane receptor that anchors astrocyte endfeet to the parenchymal BM [13–15] via high-affinity interactions with laminin 1 and 2. Dystroglycan was identified as a specific substrate of MMP-2 and MMP-9 [16]. Thus MMPs, secreted by juxtavascular microglia, might control the ECM composition, and as a consequence MMPs are involved in the integrity and function of the glia limitans.
2. The MMP Family
The MMPs are a family of zinc containing endoproteinases that share structural domains but differ in substrate specificity, cellular sources, and inducibility. The major function is the degradation and remodeling of all components of the ECM. As a group of more than 20 structurally related enzymes, they can be divided according to their substrate affinity profile: gelatinases (MMP-2 and -9), interstitial collagenases (MMP-1, -8, and -13), broad-specific stromelysins (MMP-7 and -13), and other variants (see Table 1) [17]. Together, the MMP substrate repertoire includes the extracellular matrix components, fibrillar collagens, elastin as well as matrix proteoglycan core proteins, and furthermore an expanding range of nonmatrix substrates [18, 19]. MMPs are synthesized in an inactive proform that is activated extracellularly by proteolytic cleavage under the regulation of several inflammatory mediators, including cytokines, chemokines, free radicals and steroids [20, 21]. Moreover certain MMPs are able to activate others; for example, MMP-12 was shown to activate MMP-2 and MMP-3, thereby leading to an exacerbation of proteolytic processes [22].
Table 1.
MMP overview. It was found that MMP-4, -5, and -6 were identical with MMP-2 or -3. Data compiled from Parks et al. 2004 [17] and Sbardella et al. 2012 [160].
| MMP group | MMP subgroup | Designation | Alternative name | Matrix substrate | Bioactive substrate |
|---|---|---|---|---|---|
|
Simple hemopexin-containing MMPs |
Collagenases | MMP-1 | Collagenase-1/ColA/ColB/Fibroblast collagenase/interstitial collagenase | Unclear: Type I and II fibrillar collagens; collagens I, II, III, VII,VIII, X; gelatin; aggrecan; link protein; entactin; tenascin; perlecan | a2-M; a-PI; a1-antichymotrypsin; IGFBP-2, 3, 5; proIL-1b; CTGF |
| MMP-8 | Neutrophil collagenase | Collagens I, II, III, V, VII, VIII, X; Fn; entactin; tenascin; gelatin; aggrecan; link protein, Mouse CXCL5 | a-PI | ||
| MMP-13 | Collagenase-3 | Collagens I, II, III, IV, VII, IX, X, XIV; aggrecan; gelatin; Fn; tenascin; osteonectin; Ln; perlecan | CTGF; ProTGF-b; MCP-3; a1- antichymotrypsin; plasminogen | ||
| Stromelysins | MMP-3 | Stromelysin-1 (transin-1) | E-cadherin, Laminin, type IV collagen, Latent TGF-β1; Aggrecan; decorin; gelatin; Fn; Ln; collagens III, IV, V, IX, X, XI; tenascin; link protein; perlecan; osteonectin; entactin | IGFBP-3; proIL-1b; HB-EGF; CTGF; Ecadherin; a1-antichymotrypsin; a1-PI; a2-M; plasminogen; uPA; pro-MMP-1, 7, 8, 9, 13 | |
| MMP-10 | Stromelysin-2 | Aggrecan; Ln; Fn; gelatin; collagens III, IV, V, IX, X, XI; tenascin; link protein; | Pro-1, 8, 10 | ||
| Others | MMP-12 | Macrophage metalloelastase | Latent TNF, Elastin; aggrecan; Fn; collagen IV; gelatin; vitronectin; entactin; osteonectin; Ln; nidogen | Plasminogen; apolipoprotein(a) | |
| MMP-19 | RASI-1 | Collagen IV; gelatin; Fn; tenascin; aggrecan; entactin; COMP; Ln; nidogen | IGFBP-3; proIL-1b; HB-EGF; CTGF; E-cadherin; a1-antichymotrypsin; a1-PI; a2-M; plasminogen; uPA; pro-MMP-1, 7, 8, 9, 13 | ||
| MMP-20 | Enamelysin | Amelogenin; aggrecan; gelatin; COMP | Unknown | ||
| MMP-27 | None | Unknown | Unknown | ||
|
| |||||
|
Gelatin-binding MMPs |
Gelatinases | MMP-2 | Gelatinase A/72kDa gelatinase | CCL-7/CXCL12, gelatin; collagens I, IV, V, VII, X, XI, XIV; Ln; Fn; elastin;aggrecan; osteonectin; link protein | ProTGF-b; FGF receptor I; MCP-3; IGFBP-5; proIL-1b; galectin-3; plasminogen; |
| MMP-9 | Gelatinase-B | Zona occludens1, α1-Antiproteinase, latent TGF-β1, latent VEGF, Fibrin, NG2 proteoglycan; gelatin; collagens I, III, IV, V, VII, X, XII; elastin; entactin; aggrecan; Fn; link protein; vitronectin; N-telopeptide of collagen I | ProTGF-b; IL-2 receptor a; Kit-L; IGFBP-3; proIL-1b; ICAM-1; a1-PI; galectin-3; plasminogen | ||
|
| |||||
| Furin-activated | MMP-11 | Stromelysin-3 | Fn; Ln; aggrecan; gelatins | a1-PI; a2-M; IGFBP-1 | |
| Secreted MMPs | MMP-28 | Epilysin | Unknown | Casein | |
|
| |||||
| Vitronectin-like insert MMPs | MMP-21 | None | Unknown | Unknown | |
|
| |||||
| Minimal domain MMPs |
Matrilysins | MMP-7 | Matrilysin-1 (Pump-1) | Pro-α-defensins, FAS ligand, latent TNF, syndecan-1, E-cadherin, Elastin; Aggrecan; gelatin; Fn; Ln; elastin; entactin; collagens, III, IV, V, IX, X, XI; tenascin; decorin; link protein; vitronectin | Both lack the hemopexin-line domain, they process collagen IV but not collagen I, Pro a-defensin; Fas-L; b4 integrin; E-cadherin; proTNF-a; CTGF; HB-EGF; RANKL; IGFBP-3; plasminogen |
| MMP-26 | Matrilysin-2 (endometase) | Gelatin; collagen IV; Fn; fibrinogen; vitronectin | pro-MMP-9; a1-PI | ||
|
| |||||
|
Type I transmembrane MMPs |
MTs-MMPs | MT1-MMP/MMP14 | Pro-MMP-2, fibrillar collagens, Fibrin, Syndecan-1, γ1-subunit of laminin-5, collagen I, II, III; gelatin; aggrecan; Fn; Ln; fibrin; vitronectin; entactin; proteoglycans; Ln-5 | Pro-MMP-2; Pro-MMP-13; CD44; MCP-3; tissue transglutaminase | |
| MT2-MMP/MMP-15 | Fibrin Fn; tenascin; nidogen; aggrecan; entactin; collagen; gelatin; perlecan; Ln; vitronectin | Pro-MMP-2; tissue transglutaminase | |||
| MT3-MMP/MMP-16 | Fibrin, Syndecan-1; collagen III; aggrecan; gelatin; Fn; vitronectin. | Pro-MMP-2; tissue transglutaminase | |||
| MT5-MMP/MMP-24 | Gelatin; fibronectin; vitronectin; collagen, aggrecan; PG | Pro-MMP-2 | |||
|
| |||||
| GPI-linked MMPs | MTs-MMPs | MT4-MMP/MMP-17 | Gelatin; fibrinogen | Unknown | |
| MT6-MMP/MMP-25 | Gelatin; collagen IV; fibrin; Fn; Ln | ProMMP-2 | |||
|
| |||||
| Type II transmembrane MMPs | MTs-MMPs | MMP-23A | Femalysin | Unknown | Unknown |
| MMP-23B | Gelatin | Unknown | |||
As proteolytic enzymes, MMPs have important roles in development and physiology. They are thus linked to physiological activities in the CNS, such as myelin formation, axonal growth, angiogenesis, and regeneration [23, 24]. In general, a deviant expression or overproduction of these MMPs leads to tissue destruction, and may contribute to brain pathologies such as Alzheimer's disease, ischemia, malignant glioma, and Parkinson's disease [25–29], when not counterbalanced by their physiological inhibitors, the tissue inhibitors of MMPs, TIMPs [18] (see Figure 1). Usually MMPs are under strict control at various levels: gene transcription, synthesis, secretion, activation, inhibition and glycosylation. Therefore, normal adult CNS contains low levels of most MMP members [30], in contrast to various neurological disorders of the CNS in which several MMPs are significantly upregulated [31].
Figure 1.
MMP inhibition is possible by targeted natural as well as synthetic inhibitors. References: Visse and Nagase [18]; Oh et al. [201]; Coussens et al. [202]; Overall and López-Otín [203].
The upregulated MMPs in the CNS have several potentially detrimental roles, including the promotion of neuroinflammation, disruption of the blood brain barrier (BBB) [20, 32], demyelination, and damage to axons and neurons (especially MMP-1 and MMP-2) [33]. MMPs also participate in the inflammatory cascade itself by actions on inflammatory mediators and their receptors [34, 35]. Thereby, several MMPs may act in concert in a so called MMP cascade [13]. Moreover, MMPs may contribute indirectly to the expansion of the inflammatory response and tissue damage by generating antigens through the breakdown of myelin or by conversion of inactive membrane bound TNF-α into the active myelinotoxic form [36]. Similar molecules (e.g., TNF receptors, L-selectin, TGF-β and FAS ligand) may, due to the action of MMPs, undergo analogous processes [37]. The definite sources of the activated MMPs are still to be determined: invading T cells may release proinflammatory cytokines that activate glia cells, which are in control of the expression, secretion and balance between MMPs, as well as the secretion of their natural and specific inhibitors (TIMPS).
3. Microglia in Inflammation
The primary immune effector cells of the brain are microglia, which are activated in response to brain injury or inflammatory conditions. Most likely, they play a pivotal role during onset, maintenance, relapse and progression of an inflammatory state. In the course of activation, they do not only release neurotrophic factors (such as nerve growth factor and brain-derived neurotrophic factor), but also neurotoxic factors (e.g., nitric oxide) and proinflammatory cytokines (TNF-α and IL-1) [38, 39]. Thus microglial activation is necessary for host defense, but this comes at the prize of additional “bystander damage” [40]. There is evidence that microglia play a detrimental role in various neurodegenerative diseases [41, 42]. However, ample data demonstrate beneficial roles for microglia, for example, by stimulating myelin repair, removal of toxic proteins from the CNS, and the prevention of chronic neurodegeneration [43, 44]. Microglial activation can be caused by neuronal cell death leading to secretion of signaling molecules (including α-synuclein, neuromelanin, and active forms of MMP-3) [38, 39, 45, 46]. The expression of MMPs, produced in microglia at sites of inflammation upon activation (such as LPS and Con A [47, 48], could be shown in various studies [1, 49, 50]. Particularly the secreted MMP-2 and MMP-9 [16, 51] seem to be the key modulators (Figure 2).
Figure 2.
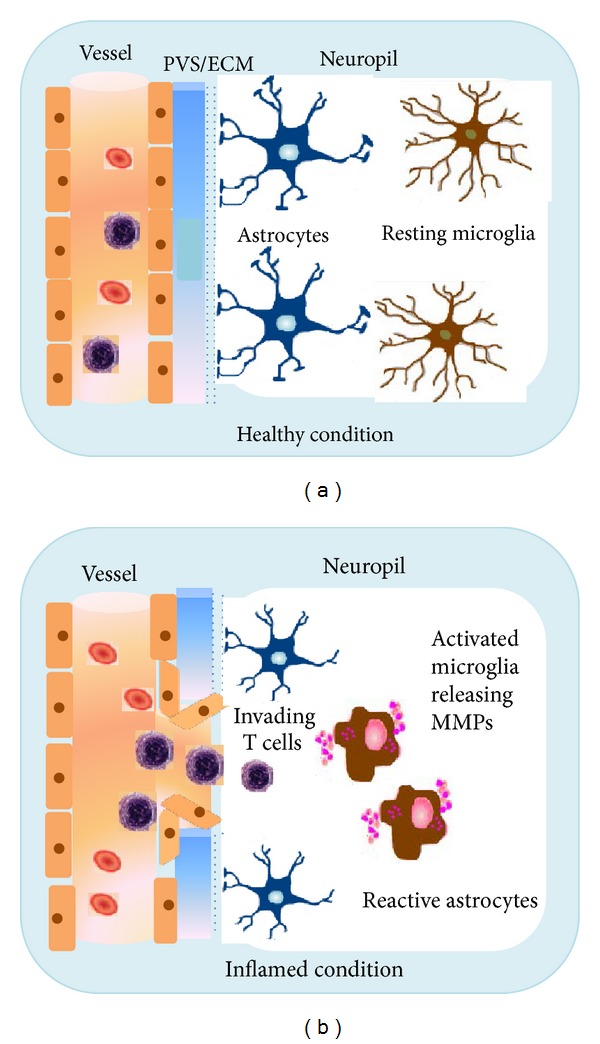
The blood brain barrier (BBB) in healthy and inflamed condition. (a) Vessel, endothelium, extracellular matrix (ECM), and glia limitans are intact. Microglia cells are in a resting state. (b) In the inflammed CNS breakdown of the BBB takes places. The glia limitans is opened, and astrocytic endfeet are drawn away. Reactive microglia secrete MMPs facilitating the opening of the BBB. Invading T cells migrate from the vessel via the perivascular space (PVS) into the neuropil.
4. MMP-2 and MMP-9 in Inflammation
MMP-2 and MMP-9 are structurally related and share the common feature of a zinc-binding domain. MMP-2 (gelatinase A a 72 kDa type IV collagenase) and MMP-9 (gelatinase B a 92 kDa type IV and type V collagenase) degrade collagens IV and V in their native forms [52]. Besides collagen, MMP-9 targets a variety of other substrates, for example, substance P [53] and b-amyloid (1–40) [54], and MMP-2 cleaves b-amyloid (1–40) and b-amyloid (1–42) [55]. MMP-9 can also degrade human myelin basic protein (MBP), thereby directly contributing to myelin damage [56–58].
MMP-9 was called a tuner and amplifier of immune functions [59], because of its assistance in peripheralization of leukocytes in response to chemokines [59] into sites of inflammation and by acting as switch and catalyst at the interplay between the innate and adaptive immune systems. MMP-9 has been implicated in opening the route for immune cells into the neuropil in various diseases, including not only MS but also strokes and brain injuries [1, 25, 31, 60–65]. In fact, the infarct size can be lessened by reducing the MMP-9 activity with a monoclonal antibody [66] or through enzymatic inhibition respectively, gene knockout [65].
Although the cause of MS remains unknown, MMPs are implicated in the pathology of MS. Focal BBB leakage and extravasation of immune cells into the brain parenchyma are the earliest steps in the pathogenesis of MS as mentioned above [67, 68]. MMPs are effectors of BBB disruption [69]; extensive studies in MS and EAE demonstrated especially activity of MMP-2 [70] and MMP-9 [71, 72]. MMP-9 around blood vessels suggest that MMP-9 might be pathologically involved in the disruption of the parenchymal basement membranes [73], paving the way for infiltrating cells of the immune system [1]. Within the CNS immune cells orchestrate myelin and axonal destruction resulting in severe destruction of normal CNS constituents. The histopathological hallmark of MS is the plaque, a well-demarcated white matter lesion characterized by demyelination and axonal loss. Expression of MMP-1, -2, -3, -7, and -9 in monocytes/macrophages, microglia, astrocytes, and lymphocytes around perivascular cuffs in MS lesions has been described [73–75]. We could also confirm by immunostaining that microglia are sources of MMP-2 and MMP-9 (see Figure 3).
Figure 3.
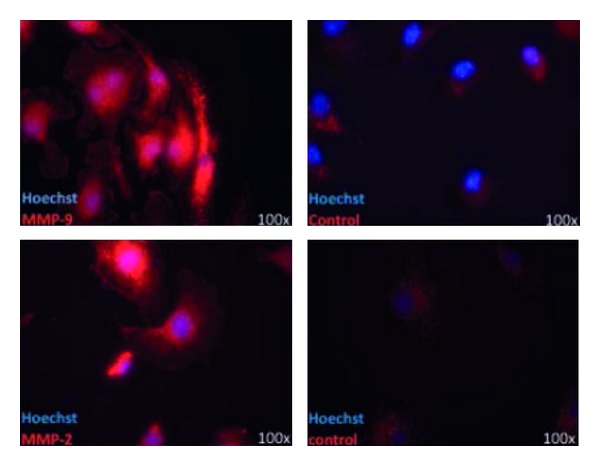
Immunohistochemistry of murine microglia, stained for MMP-2 and MMP-9. Microglia from CD11c GFP mice, fixed with PFA, treated with TBS and NGS, anti-MMP-2 (1 : 125) and anti-MMP-9 (1 : 500); control: BSA and secondary antibodies.
The secreted MMP-9 can cause demyelination and axonal injury [76, 77]. Axonal damage is considered to be a major cause of secondary progression (with irreversible neurological impairment) [78–80], which seems to be caused not only by T cells [81] but also by microglia/macrophages and their toxic products [75, 82, 83]. Cuzner et al. [84] could confirm enhanced MMP-9 expression in reactive microglia and astrocytes in autopsies from MS brains. Interestingly, intrathecal synthesis of MMP-9 appears to be specific for MS [85, 86]. Around the time of onset of the symptoms in EAE, elevated levels of MMP-9 can be found. The administration of GM6001 (a MMP inhibitor) improved the clinical condition by blocking the BBB injury [87].
The view that MMP-9 is a promoter of neuroinflammation has been additionally supported by the finding that young (3-4 weeks) but not older (7-8 weeks) MMP-9 null mice were less susceptible to development of EAE than wild type controls [71]. In addition, MMP-2 null mice were reported to have an earlier onset and more severe disease compared to wild type controls. Apparently this was related to a compensatory increase of MMP-9 in the MMP-2 null mice [71]. Enzyme inhibitors of MMPs have been shown to ameliorate the clinical course and reduce inflammatory cell infiltration in EAE [87–89]. Treatment of PTx-injected CCL2-overexpressing mice with the synthetic broad-spectrum inhibitor BB-94 (Batimastat) alleviated symptoms of neuroinflammation [90] and put blood-derived cells on hold in perivascular spaces. This was the first evidence that the second step of neuroinflammation, that is, passage of the glia limitans, but not the first, migration across the vascular wall, depends on MMPs.
The production of MMP-9 is negatively regulated by IL-4 [91], IL-10 [92], and interferon-β [93] whereas transforming growth factor-β was found to enhance the production of MMP-9 in transformed lymphocytes [94]. Furthermore it was shown that cytokines, chemokines [95, 96], eicosanoides and peptidoglycan, lectins, double-stranded RNS and endotoxin [31, 59, 65, 97, 98] are acting as soluble upregulators [99–101]. Potent stimulators of MMP-9 and MMP-2 expression in cultured astrocytes and microglia are the proinflammatory cytokines interleukin 1 (IL-1), tumor necrosis factor-alpha (TNF-α), and lipopolysaccharide (LPS) [102, 103].
Furthermore, interferon-β (an immune-modulator that is commonly used in MS) inhibits the expression of MMPs in glial cells. Liuzzi et al. [104] demonstrated that LPS treated microglia secreted higher levels of MMP-9. As soon as the microglia cells were pre-/treated with different doses of IFN-β they found dose-dependent inhibition of MMP-9. IFN-γ or IFN-β was also suggested to inhibit the expression of MMP-9 in human astroglioma and fibrosarcoma cell lines, as well as in primary astrocytes, supposable by the modulation of transcription factors that regulate the MMP-9 transcription [105, 106]. Still an indirect pathway cannot be excluded: IFN-β could regulate the MMP expression either through the reduction of proinflammatory cytokines or by the inhibition of the activity of enzymes involved in MMP activation [107]. IFN-β also reduces the production of MMP-9 by T cells and monocytes in vitro [8, 108, 109] leading to diminished MMP-9 levels in serum of multiple sclerosis patients [110, 111]. This was paralleled by the clinical recovery of the patients, presumably as a result of a significant reduction of T lymphocytes infiltrating in the brain. Besides interferon-β also increases gene transcription of TIMP-1, thus attenuating MMP overactivity in MS. Intravenous gamma globulins (IVIG) used in severe cases of MS were shown to diminish the amount of secreted MMP-9 and its mRNA expression [112].
In addition to their detrimental roles MMPs might also have a beneficial effect in MS, as they also have important functions in (the developmental) plasticity of the nervous system [70, 113, 114]: MMP-9 mediates the oligodendrocytes process outgrowth [115]. Cultured oligodendrocytes secrete MMP-9, and cell-associated gelatinases are found at the site of their growing tips of their processes [116].
Notably MMP-9 is significantly upregulated in the acute period of spinal cord injury [117, 118] which might promote the maturation of oligodendrocytes and their formation of myelin [119]. MMP-9 [115, 116] and MMP-12 [120] are expressed by oligodendrocytes and seem to be essential for regulating the extension of their processes. Remyelination was impaired in MMP-9 and MMP-9/-12-null mice, correlating with fewer mature oligodendrocytes [121]. Taking that into account the MMP-9 secretion by microglia might allow a microenvironment in lesions for better remyelination and repair [31]. MMP-2 levels increase between 7 and 14 days after spinal cord injury, and MMP-2 null mice do not recover equally well as wild type controls do suggesting that the delayed expression is necessary for ECM remodeling and functional recovery [122].
MMP-1, MMP-3, and MMP-8 were also reported to play a role in BBB disruption followed by a leukocyte infiltration into the brain [123, 124]. Woo et al. [125] demonstrated that the mRNA expression of MMP-1, -3, -8, and -9 in primary cultured microglia cells was significantly increased by LPS and other immunostimulants. Furthermore, the inhibition of MMP-3 and MMP-9 could suppress inflammatory reactions in activated microglia (such as iNOS, proinflammatory cytokine expression, and upstream signaling molecules such as MAPKs, which would amplify the inflammatory cascade by initiating the MMP production in an autocrine or paracrine way).
5. Conclusion
The data discussed beforehand reinforces the concept that leukocyte transmigration involves distinct molecular mechanisms. MMP-2 and MMP-9 are expressed by microglial cells which contribute to the formation of the glia limitans [126]. These cell types might be responsible for the opening of the glia limitans and the further progression of autoreactive immune cells into the neuropil. Besides the contribution of MMP-9 to BBB breakdown, it is also involved in the generation of autoimmune epitopes as well as the bioavailability of cytokines.
The treatment of neurological inflammation still remains a challenge today. Targeting MMPs in the CNS may serve as therapeutic option in autoimmune diseases. It is tempting to employ inhibitors of MMP activity to abrogate increased MMP expression (partially driven by microglial activation) within the inflamed CNS. However, MMPs also fulfill beneficial roles in the CNS, including mediation of tissue repair [31], synaptic plasticity [127], learning, and memory [128]. With respect to the multiple roles of MMPs, not only detrimental but also physiological, the need and the judicious application of specific inhibitors against individual MMPs should be highlighted. Due to their bifaced role, timing may also be crucial for therapeutic effects. There is the likelihood that nonspecific MMP inhibitors, although protecting against particular detrimental effects of some MMPs, could block useful actions of MMPs, thus slowing down disease recovery, too. It will be necessary to analyze further therapies aimed at decreasing MMP-2 and MMP-9 expression or activity.
6. Microglia and Gliomas
The most common brain tumors are malignant gliomas, infiltrating diffusely into normal brain parenchyma [129]. So far all current (multimodal) therapeutic approaches were ineffective, and life expectancy from the time of the diagnosis in glioblastoma multiforme is on average 14 months [130–133].
In 1921 Rio-Hortega [134], was the first to describe the presence of microglia cells in brain tumors. Microglia contribute substantially (at least 1/3) to the tumor mass of glioblastoma as they make up the largest population of tumor-infiltrating cells [135–138]. Microglia seem to possess a decisive tumor-supporting role by creating a microenvironment, which plays a critical role in tumor initiation and progression [139–144]. This special environment is also an immunosuppressive milieu, where, for example, IL-10 is released [135, 145, 146]. Microglia/macrophages play also an influential role in glioma invasion: there is a positive correlation in their density in gliomas with the invasiveness and grade of gliomas [139, 144]. The antitumor properties, namely glioma-cytotoxic effects of microglia, could only be shown in vitro so far [147, 148]. Favoring gliomas' growth could be due to a suppression and/or control of microglial cells by glioma cells and glioma-derived molecules (e.g., their loss of phagocytic function [149]. It was also suggested that, under the influence of glioma cells, microglia develop a different, noninflammatory phenotype suppressing their defense properties [139, 144, 150]. Instead of releasing pro-inflammatory cytokines, microglia upregulate enzymes that facilitate tumor invasion and proliferation. A key mechanism in the expansion and invasion of gliomas is the degradation of extracellular matrix by membrane-bound or secreted proteases such as MMPs [151], especially matrix-metalloproteinase-2 [152] and MMP-9 [151].
7. MMPs and Glioma Cells
Due to their ECM-degrading ability and confirmed upregulation in almost all cancer entities, MMP-2 has been linked to invasiveness and dissemination [153–155]. Because serum concentration of MMP-2 was significantly elevated in tumor patients, MMP-2 was suggested as a diagnostic and prognostic marker [156, 157]. On the other hand elevated MMP-9 levels in the serum seem to be even more relevant values, because in healthy individuals under physiological conditions MMP-9 is hardly detectable [158]. Abnormal MMP-9 concentrations in patients serum were also shown for brain cancers [159], and notably there is a positive correlation with poor prognosis [160].
So far, there is no evidence that links MMP-2 to a special phase of tumor development (in contrast to MMP-9): besides creating a microenvironment in the early phase favoring cancer growth (activation of growth factors), the transition into an undifferentiated phenotype permitting migration and invasiveness is also related to MMP-2 activity, for example, the proteolytic detachment of adhesion molecules like integrins or cadherins or cytoskeleton changes [161, 162]. MMP-2 acts in multiple ways on tumor cells by modulation of their metabolism, their receptor turnover [163], and their resistance to apoptosis [164]. In fact, anti-MMP-2 siRNA-treated glioma cells underwent apoptosis [165] and MMP-2 inhibition autophagy-associated cell death [166].
The expression of the MMP-2 gene in human glioma tissues was found to be upregulated in comparison to normal brain tissue, and dramatically increased in glioblastomas [167–169]. MMP-9 expression could be correlated with high malignancy and progression of gliomas [170, 171]. Various studies show that glioma and microglia cells both produce MMP-2 in vitro [144, 168] and in situ [172]. However, MMP-2 is released as an inactive profrom by glioma cells (especially at the invasive tumor zone), and glioma cells themselves are unable to activate pro-MMP-2. Since the extracellular activator MT1-MMP is highly upregulated in glioma infiltrating microglia [138, 151, 172, 173], glioma cells engage microglial cells to promote their spread and survival [174]. This concept of microglial “abuse” has been impressively demonstrated by the group of Kettenmann [138].
8. Pathways of MMP Induction and Suppression in Gliomas
Another key player of glioma motility and invasion seems to be FasL, which is expressed in tumor cells. It not only induces apoptosis in T cells thereby leading to local immunosuppression, but blockade of Fas signaling resulted in impaired MMP-2 activity with a subsequent reduction of glioma invasiveness and motility [175]. The expression of MMPs is also facilitated by glioma-derived TGF-β which suppresses the expression of TIMPs and also has an immunosuppressive role [176–178]. The inactive precursor of TGF-β can be processed by MMP-2 [179, 180], and TGF-β induces gene transcprition of MMP-2, thus generating a vicious circle leading to further tumor growth [181, 182] (Figure 4).
Figure 4.
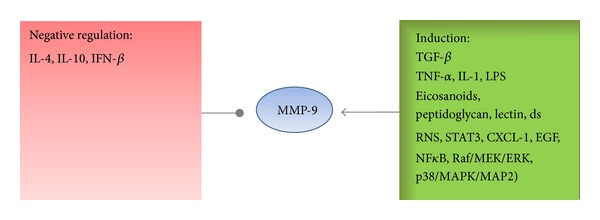
Regulation and induction of MMP-9: a variety of molecules are involved.
Another player in glioma growth, gliomagenesis, and progression is the activation of STAT3 [183]. This signal transducer and activator of transcription protein 3 is constitutively activated in glioblastoma cell lines [184–187] and increases MMP-9 expression and activation in human astrocytoma cell lines [188, 189]. The expression of MMP-2, -9, and -14 in microglia/macrophages was also shown to be enhanced by glioma-derived CX3CL1 (Chemokine (C-X3-C Motif) Ligand) and is significantly associated with the recruitment of microglia into the tumor [190]. Another role in the production of MMP-9 in glioma cells is played by protein kinase C (PKC) [191] and IL-6 is a confirmed growth factor for glioma stem cells, too [192]. The tumor-promoting role of IL-6 may be exerted via MMP-2, whose production is increased by IL-6 [193]. Glioma-induced MMP-2 activity in microglia could be significantly decreased by the A1AR (an adenosine receptor subtype, found on microglia and neurons) [194], which might explain the fact that adenosine treatment leads to decreased extracellular protease activity and thereby exerts its inhibitory effects on glioma invasion. Early studies in MS patients could also show that A1AR activation in microglia interfered with the MMPs production [195].
Although the substrate specifity of MMP-2 and MMP-9 overlaps, MMP-9 (in contrast to MMP-2) is highly inducible mostly by integrins, growth factors, and cytokines [13, 196] leading to a defined chronologically and spatially distribution. The expression of MMP-9 is further triggered by autocrine or paracrine mechanisms (IL-1β, TNF-α, and TGF-β), cell binding (to fibronectin or vitronectin), EGF release or distinct molecular pathways (transcription factors NF-κB, Raf/MEK/ERK cascade, or the p38 MAPK/MAPK2-signaling) [180, 196–200] (Figure 5).
Figure 5.
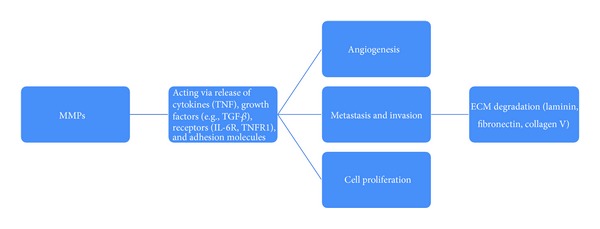
MMP in gliomas: the various roles of MMPs in promoting the growth of cancer cells.
In sum, ample data describe the communication between tumor cells and microglia. Microglia and their expression of MMPs could be a crucial target for future therapeutic options in gliomas, due to their profound involvement in tumor progression.
References
- 1.Rosenberg GA. Matrix metalloproteinases in neuroinflammation. GLIA. 2002;39(3):279–291. doi: 10.1002/glia.10108. [DOI] [PubMed] [Google Scholar]
- 2.Tanaka R, Iwasaki Y, Koprowski H. Ultrastructural studies of perivascular cuffing cells in multiple sclerosis brain. American Journal of Pathology. 1975;81(3):467–478. [PMC free article] [PubMed] [Google Scholar]
- 3.Bechmann I, Galea I, Perry VH. What is the blood-brain barrier (not)? Trends in Immunology. 2007;28(1):5–11. doi: 10.1016/j.it.2006.11.007. [DOI] [PubMed] [Google Scholar]
- 4.Uccelli A, Aloisi F, Pistoia V. Unveiling the enigma of the CNS as a B-cell fostering environment. Trends in Immunology. 2005;26(5):254–259. doi: 10.1016/j.it.2005.02.009. [DOI] [PubMed] [Google Scholar]
- 5.Hickey WF. Basic principles of immunological surveillance of the normal central nervous system. GLIA. 2001;36(2):118–124. doi: 10.1002/glia.1101. [DOI] [PubMed] [Google Scholar]
- 6.Goetzl EJ, Banda MJ, Leppert D. Matrix metalloproteinases in immunity. Journal of Immunology. 1996;156(1):1–4. [PubMed] [Google Scholar]
- 7.Madri JA, Graesser D. Cell migration in the immune system: the evolving inter-related roles of adhesion molecules and proteinases. Developmental Immunology. 2000;7(2–4):103–116. doi: 10.1155/2000/79045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Leppert D, Waubant E, Galardy R, Bunnett NW, Hauser SL. T cell gelatinases madiate basement membrane transmigration in vitro. Journal of Immunology. 1995;154(9):4379–4389. [PubMed] [Google Scholar]
- 9.Xia M, Sreedharan SP, Dazin P, Damsky CH, Goetzl EJ. Integrin-dependent role of human T cell matrix metalloproteinase activity in chemotaxis through a model basement membrane. Journal of Cellular Biochemistry. 1996;61(3):452–458. doi: 10.1002/(SICI)1097-4644(19960601)61:3%3C452::AID-JCB12%3E3.0.CO;2-L. [DOI] [PubMed] [Google Scholar]
- 10.Graesser G. Integrins, proteinases, and EAE: Explaining a complex web of interactions. Laboratory Investigation. 1998;78(12):p. 1445. [Google Scholar]
- 11.Laurie GW, Leblond CP, Martin GR. Localization of type IV collagen, laminin, heparan sulfate proteoglycan, and fibronectin to the basal lamina of basement membranes. Journal of Cell Biology. 1982;95(1):340–344. doi: 10.1083/jcb.95.1.340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Sixt M, Engelhardt B, Pausch F, Hallmann R, Wendler O, Sorokin LM. Endothelial cell laminin isoforms, laminins 8 and 10, play decisive roles in T cell recruitment across the blood-brain barrier in experimental autoimmune encephalomyelitis. Journal of Cell Biology. 2001;153(5):933–945. doi: 10.1083/jcb.153.5.933. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.van den Steen PE, Dubois B, Nelissen I, Rudd PM, Dwek RA, Opdenakker G. Biochemistry and molecular biology of gelatinase B or matrix metalloproteinase-9 (MMP-9) Critical Reviews in Biochemistry and Molecular Biology. 2002;37(6):375–536. doi: 10.1080/10409230290771546. [DOI] [PubMed] [Google Scholar]
- 14.Tian M, Jacobson C, Gee SH, Campbell KP, Carbonetto S, Jucker M. Dystroglycan in the cerebellum is a laminin alpha 2-chain binding protein at the glial-vascular interface and is expressed in Purkinje cells. European Journal of Neuroscience. 1996;8(12):2739–2747. doi: 10.1111/j.1460-9568.1996.tb01568.x. [DOI] [PubMed] [Google Scholar]
- 15.Zaccaria ML, Di Tommaso F, Brancaccio A, Paggi P, Petrucci TC. Dystroglycan distribution in adult mouse brain: a light and electron microscopy study. Neuroscience. 2001;104(2):311–324. doi: 10.1016/s0306-4522(01)00092-6. [DOI] [PubMed] [Google Scholar]
- 16.Agrawal S, Anderson P, Durbeej M, et al. Dystroglycan is selectively cleaved at the parenchymal basement membrane at sites of leukocyte extravasation in experimental autoimmune encephalomyelitis. Journal of Experimental Medicine. 2006;203(4):1007–1016. doi: 10.1084/jem.20051342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Parks WC, Wilson CL, López-Boado YS. Matrix metalloproteinases as modulators of inflammation and innate immunity. Nature Reviews Immunology. 2004;4(8):617–629. doi: 10.1038/nri1418. [DOI] [PubMed] [Google Scholar]
- 18.Visse R, Nagase H. Matrix metalloproteinases and tissue inhibitors of metalloproteinases: structure, function, and biochemistry. Circulation Research. 2003;92(8):827–839. doi: 10.1161/01.RES.0000070112.80711.3D. [DOI] [PubMed] [Google Scholar]
- 19.Woessner JF., Jr. Matrix metalloproteinases and their inhibitors in connective tissue remodeling. FASEB Journal. 1991;5(8):2145–2154. [PubMed] [Google Scholar]
- 20.Sternlicht MD, Werb Z. How matrix metalloproteinases regulate cell behavior. Annual Review of Cell and Developmental Biology. 2001;17:463–516. doi: 10.1146/annurev.cellbio.17.1.463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Alexander CM, Werb Z. Proteinases and extracellular matrix remodeling. Current Opinion in Cell Biology. 1989;1(5):974–982. doi: 10.1016/0955-0674(89)90068-9. [DOI] [PubMed] [Google Scholar]
- 22.Matsumoto SI, Kobayashi T, Katoh M, et al. Expression and localization of matrix metalloproteinase-12 in the aorta of cholesterol-fed rabbits: relationship to lesion development. American Journal of Pathology. 1998;153(1):109–119. doi: 10.1016/s0002-9440(10)65551-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Yong VW, Power C, Forsyth P, Edwards DR. Metalloproteinases in biology and pathology of the nervous system. Nature Reviews Neuroscience. 2001;2(7):502–511. doi: 10.1038/35081571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Agrawal SM, Lau L, Yong VW. MMPs in the central nervous system: where the good guys go bad. Seminars in Cell and Developmental Biology. 2008;19(1):42–51. doi: 10.1016/j.semcdb.2007.06.003. [DOI] [PubMed] [Google Scholar]
- 25.Rosenberg GA, Dencoff JE, McGuire PG, Liotta LA, Stetler-Stevenson WG. Injury-induced 92-kilodalton gelatinase and urokinase expression in rat brain. Laboratory Investigation. 1994;71(3):417–422. [PubMed] [Google Scholar]
- 26.Yong VW, Krekoski CA, Forsyth PA, Bell R, Edwards DR. Matrix metalloproteinases and diseases of the CNS. Trends in Neurosciences. 1998;21(2):75–80. doi: 10.1016/s0166-2236(97)01169-7. [DOI] [PubMed] [Google Scholar]
- 27.Ramos-DeSimone N, Hahn-Dantona E, Sipley J, Nagase H, French DL, Quigley JP. Activation of matrix metalloproteinase-9 (MMP-9) via a converging plasmin/stromelysin-1 cascade enhances tumor cell invasion. Journal of Biological Chemistry. 1999;274(19):13066–13076. doi: 10.1074/jbc.274.19.13066. [DOI] [PubMed] [Google Scholar]
- 28.Lorenzl S, Calingasan N, Yang L, et al. Matrix metalloproteinase-9 is elevated in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced parkinsonism in mice. NeuroMolecular Medicine. 2004;5(2):119–131. doi: 10.1385/NMM:5:2:119. [DOI] [PubMed] [Google Scholar]
- 29.Yin KJ, Cirrito JR, Yan P, et al. Matrix metalloproteinases expressed by astrocytes mediate extracellular amyloid-β peptide catabolism. Journal of Neuroscience. 2006;26(43):10939–10948. doi: 10.1523/JNEUROSCI.2085-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Weaver A, Goncalves da Silva A, Nuttall RK, et al. An elevated matrix metalloproteinase (MMP) in an animal model of multiple sclerosis is protective by affecting Th1/Th2 polarization. The FASEB Journal. 2005;19(12):1668–1670. doi: 10.1096/fj.04-2030fje. [DOI] [PubMed] [Google Scholar]
- 31.Yong VW. Metalloproteinases: mediators of pathology and regeneration in the CNS. Nature Reviews Neuroscience. 2005;6(12):931–944. doi: 10.1038/nrn1807. [DOI] [PubMed] [Google Scholar]
- 32.Woessner JF., Jr. Quantification of matrix metalloproteinases in tissue samples. Methods in Enzymology. 1995;248:510–528. doi: 10.1016/0076-6879(95)48033-1. [DOI] [PubMed] [Google Scholar]
- 33.Vos CMP, Gartner S, Ransohoff RM, et al. Matrix metalloprotease-9 release from monocytes increases as a function of differentiation: implications for neuroinflammation and neurodegeneration. Journal of Neuroimmunology. 2000;109(2):221–227. doi: 10.1016/s0165-5728(00)00308-8. [DOI] [PubMed] [Google Scholar]
- 34.Hu J, van den Steen PE, Sang QXA, Opdenakker G. Matrix metalloproteinase inhibitors as therapy for inflammatory and vascular diseases. Nature Reviews Drug Discovery. 2007;6(6):480–498. doi: 10.1038/nrd2308. [DOI] [PubMed] [Google Scholar]
- 35.Page-McCaw A, Ewald AJ, Werb Z. Matrix metalloproteinases and the regulation of tissue remodelling. Nature Reviews Molecular Cell Biology. 2007;8(3):221–233. doi: 10.1038/nrm2125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Gearing AJH, Beckett P, Christodoulou M, et al. Processing of tumour necrosis factor-α precursor by metalloproteinases. Nature. 1994;370(6490):555–557. doi: 10.1038/370555a0. [DOI] [PubMed] [Google Scholar]
- 37.Chandler S, Miller KM, Clements JM, et al. Matrix metalloproteinases, tumor necrosis factor and multiple sclerosis: an overview. Journal of Neuroimmunology. 1997;72(2):155–161. doi: 10.1016/s0165-5728(96)00179-8. [DOI] [PubMed] [Google Scholar]
- 38.Block ML, Zecca L, Hong JS. Microglia-mediated neurotoxicity: uncovering the molecular mechanisms. Nature Reviews Neuroscience. 2007;8(1):57–69. doi: 10.1038/nrn2038. [DOI] [PubMed] [Google Scholar]
- 39.Kim SU, de Vellis J. Microglia in health and disease. Journal of Neuroscience Research. 2005;81(3):302–313. doi: 10.1002/jnr.20562. [DOI] [PubMed] [Google Scholar]
- 40.Streit WJ, Conde JR, Fendrick SE, Flanary BE, Mariani CL. Role of microglia in the central nervous system’s immune response. Neurological Research. 2005;27(7):685–691. doi: 10.1179/016164105X49463a. [DOI] [PubMed] [Google Scholar]
- 41.McGeer PL, Itagaki S, Boyes BE, McGeer EG. Reactive microglia are positive for HLA-DR in the substantia nigra of Parkinson’s and Alzheimer’s disease brains. Neurology. 1988;38(8):1285–1291. doi: 10.1212/wnl.38.8.1285. [DOI] [PubMed] [Google Scholar]
- 42.Gao HM, Liu B, Hong JS. Critical role for microglial NADPH oxidase in rotenone-induced degeneration of dopaminergic neurons. Journal of Neuroscience. 2003;23(15):6181–6187. doi: 10.1523/JNEUROSCI.23-15-06181.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Glezer I, Simard AR, Rivest S. Neuroprotective role of the innate immune system by microglia. Neuroscience. 2007;147(4):867–883. doi: 10.1016/j.neuroscience.2007.02.055. [DOI] [PubMed] [Google Scholar]
- 44.Simard AR, Rivest S. Neuroprotective effects of resident microglia following acute brain injury. Journal of Comparative Neurology. 2007;504(6):716–729. doi: 10.1002/cne.21469. [DOI] [PubMed] [Google Scholar]
- 45.Kim YS, Joh TH. Microglia, major player in the brain inflammation: their roles in the pathogenesis of Parkinson’s disease. Experimental and Molecular Medicine. 2006;38(4):333–347. doi: 10.1038/emm.2006.40. [DOI] [PubMed] [Google Scholar]
- 46.Kim YS, Choi DH, Block ML, et al. A pivotal role of matrix metalloproteinase-3 activity in dopaminergic neuronal degeneration via microglial activation. The FASEB Journal. 2007;21(1):179–187. doi: 10.1096/fj.06-5865com. [DOI] [PubMed] [Google Scholar]
- 47.Wahl LM, Lampel LL. Regulation of human peripheral blood monocyte collagenase by prostaglandins and anti-inflammatory drugs. Cellular Immunology. 1987;105(2):411–422. doi: 10.1016/0008-8749(87)90088-8. [DOI] [PubMed] [Google Scholar]
- 48.Lu Y, Wahl LM. Oxidative stress augments the production of matrix metalloproteinase-1, cyclooxygenase-2, and prostaglandin E2 through enhancement of NF-κB activity in lipopolysaccharide-activated human primary monocytes. Journal of Immunology. 2005;175(8):5423–5429. doi: 10.4049/jimmunol.175.8.5423. [DOI] [PubMed] [Google Scholar]
- 49.Liuzzi GM, Latronico T, Fasano A, Carlone G, Riccio P. Interferon-beta inhibits the expression of metallo proteinases in rat glial cell cultures: implications for multiple sclerosis pathogenesis and treatment. Multiple Sclerosis. 2004;10(3):290–297. doi: 10.1191/1352458504ms1016oa. [DOI] [PubMed] [Google Scholar]
- 50.Nuttall RK, Silva C, Hader W, et al. Metalloproteinases are enriched in microglia compared with leukocytes and they regulate cytokine levels in activated microglia. GLIA. 2007;55(5):516–526. doi: 10.1002/glia.20478. [DOI] [PubMed] [Google Scholar]
- 51.Webster NL, Crowe SM. Matrix metalloproteinases, their production by monocytes and macrophages and their potential role in HIV-related diseases. Journal of Leukocyte Biology. 2006;80(5):1052–1066. doi: 10.1189/jlb.0306152. [DOI] [PubMed] [Google Scholar]
- 52.Mignatti P, Rifkin DB. Biology and biochemistry of proteinases in tumor invasion. Physiological Reviews. 1993;73(1):161–195. doi: 10.1152/physrev.1993.73.1.161. [DOI] [PubMed] [Google Scholar]
- 53.Backstrom JR, Tokes ZA. The 84-kDa form of human matrix metalloproteinase-9 degrades substance P and gelatin. Journal of Neurochemistry. 1995;64(3):1312–1318. doi: 10.1046/j.1471-4159.1995.64031312.x. [DOI] [PubMed] [Google Scholar]
- 54.Backstrom JR, Lim GP, Cullen MJ, Tökés ZA. Matrix metalloproteinase-9 (MMP-9) is synthesized in neurons of the human hippocampus and is capable of degrading the amyloid-β peptide (1-40) Journal of Neuroscience. 1996;16(24):7910–7919. doi: 10.1523/JNEUROSCI.16-24-07910.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Roher AE, Kasunic TC, Woods AS, Cotter RJ, Ball MJ, Fridman R. Proteolysis of Aβ peptide from Alzheimer disease brain by gelatinase A. Biochemical and Biophysical Research Communications. 1994;205(3):1755–1761. doi: 10.1006/bbrc.1994.2872. [DOI] [PubMed] [Google Scholar]
- 56.Proost P, Van Damme J, Opdenakker G. Leukocyte gelatinase B cleavage releases encephalitogens from human myelin basic protein. Biochemical and Biophysical Research Communications. 1993;192(3):1175–1181. doi: 10.1006/bbrc.1993.1540. [DOI] [PubMed] [Google Scholar]
- 57.Gijbels K, Proost P, Masure S, Carton H, Billiau A, Opdenakker G. Gelatinase B is present in the cerebrospinal fluid during experimental autoimmune encephalomyelitis and cleaves myelin basic protein. Journal of Neuroscience Research. 1993;36(4):432–440. doi: 10.1002/jnr.490360409. [DOI] [PubMed] [Google Scholar]
- 58.Chandler S, Coates R, Gearing A, Lury J, Wells G, Bone E. Matrix metalloproteinases degrade myelin basic protein. Neuroscience Letters. 1995;201(3):223–226. doi: 10.1016/0304-3940(95)12173-0. [DOI] [PubMed] [Google Scholar]
- 59.Opdenakker G, van den Steen PE, van Damme J. Gelatinase B: a tuner and amplifier of immune functions. Trends in Immunology. 2001;22(10):571–579. doi: 10.1016/s1471-4906(01)02023-3. [DOI] [PubMed] [Google Scholar]
- 60.Lukes A, Mun-Bryce S, Lukes M, Rosenberg GA. Extracellular matrix degradation by metalloproteinases and central nervous system diseases. Molecular Neurobiology. 1999;19(3):267–284. doi: 10.1007/BF02821717. [DOI] [PubMed] [Google Scholar]
- 61.Rosenberg GA, Dencoff JE, Correa N, Jr., Reiners M, Ford CC. Effect of steroids on CSF matrix metalloproteinases in multiple sclerosis: relation to blood-brain barrier injury. Neurology. 1996;46(6):1626–1632. doi: 10.1212/wnl.46.6.1626. [DOI] [PubMed] [Google Scholar]
- 62.Rosenberg GA, Estrada EY, Dencoff JE. Matrix metalloproteinases and TIMPs are associated with blood-brain barrier opening after reperfusion in rat brain. Stroke. 1998;29(10):2189–2195. doi: 10.1161/01.str.29.10.2189. [DOI] [PubMed] [Google Scholar]
- 63.Anthony DC, Miller KM, Fearn S, et al. Matrix metalloproteinase expression in an experimentally-induced DTH model of multiple sclerosis in the rat CNS. Journal of Neuroimmunology. 1998;87(1-2):62–72. doi: 10.1016/s0165-5728(98)00046-0. [DOI] [PubMed] [Google Scholar]
- 64.Mun-Bryce S, Rosenberg GA. Gelatinase B modulates selective opening of the blood-brain barrier during inflammation. American Journal of Physiology. 1998;274(5):R1203–R1211. doi: 10.1152/ajpregu.1998.274.5.R1203. [DOI] [PubMed] [Google Scholar]
- 65.Asahi M, Asahi K, Jung JC, del Zoppo GJ, Fini ME, Lo EH. Role for matrix metalloproteinase 9 after focal cerebral ischemia: effects of gene knockout and enzyme inhibition with BB-4. Journal of Cerebral Blood Flow & Metabolism. 2000;20(12):1681–199. doi: 10.1097/00004647-200012000-00007. [DOI] [PubMed] [Google Scholar]
- 66.Romanic AM, White RF, Arleth AJ, Ohlstein EH, Barone FC. Matrix metalloproteinase expression increases after cerebral focal ischemia in rats: inhibition of matrix metalloproteinase-9 reduces infarct size. Stroke. 1998;29(5):1020–1030. doi: 10.1161/01.str.29.5.1020. [DOI] [PubMed] [Google Scholar]
- 67.Kermode AG, Thompson AJ, Tofts P, et al. Breakdown of the blood-brain barrier precedes symptoms and other MRI signs of new lesions in multiple sclerosis. Pathogenic and clinical implications. Brain. 1990;113(part 5):1477–1489. doi: 10.1093/brain/113.5.1477. [DOI] [PubMed] [Google Scholar]
- 68.Kwon EE, Prineas JW. Blood-brain barrier abnormalities in longstanding multiple sclerosis lesions. An immunohistochemical study. Journal of Neuropathology and Experimental Neurology. 1994;53(6):625–636. doi: 10.1097/00005072-199411000-00010. [DOI] [PubMed] [Google Scholar]
- 69.Rosenberg GA. Matrix metalloproteinases in brain injury. Journal of Neurotrauma. 1995;12(5):833–842. doi: 10.1089/neu.1995.12.833. [DOI] [PubMed] [Google Scholar]
- 70.Romanic AM, Madri JA. Extracellular matrix-degrading proteinases in the nervous system. Brain Pathology. 1994;4(2):145–156. doi: 10.1111/j.1750-3639.1994.tb00825.x. [DOI] [PubMed] [Google Scholar]
- 71.Dubois B, Masure S, Hurtenbach U, et al. Resistance of young gelatinase B-deficient mice to experimental autoimmune encephalomyelitis and necrotizing tail lesions. Journal of Clinical Investigation. 1999;104(11):1507–1515. doi: 10.1172/JCI6886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Kieseier BC, Kiefer R, Clements JM, et al. Matrix metalloproteinase-9 and -7 are regulated in experimental autoimmune encephalomyelitis. Brain. 1998;121(part 1):159–166. doi: 10.1093/brain/121.1.159. [DOI] [PubMed] [Google Scholar]
- 73.Lindberg RL, de Groot CJ, Montagne L, et al. The expression profile of matrix metalloproteinases (MMPs) and their inhibitors (TIMPs) in lesions and normal appearing white matter of multiple sclerosis. Brain. 2001;124(9):1743–1753. doi: 10.1093/brain/124.9.1743. [DOI] [PubMed] [Google Scholar]
- 74.Maeda A, Sobel RA. Matrix metalloproteinases in the normal human central nervous system, microglial nodules, and multiple sclerosis lesions. Journal of Neuropathology and Experimental Neurology. 1996;55(3):300–309. doi: 10.1097/00005072-199603000-00005. [DOI] [PubMed] [Google Scholar]
- 75.Cossins JA, Clements JM, Ford J, et al. Enhanced expression of MMP-7 and MMP-9 in demyelinating multiple sclerosis lesions. Acta Neuropathologica. 1997;94(6):590–598. doi: 10.1007/s004010050754. [DOI] [PubMed] [Google Scholar]
- 76.Diaz-Sanchez M, Williams K, DeLuca GC, Esiri MM. Protein co-expression with axonal injury in multiple sclerosis plaques. Acta Neuropathologica. 2006;111(4):289–299. doi: 10.1007/s00401-006-0045-0. [DOI] [PubMed] [Google Scholar]
- 77.Newman TA, Woolley ST, Hughes PM, Sibson NR, Anthony DC, Perry VH. T-cell- and macrophage-mediated axon damage in the absence of a CNS-specific immune response: involvement of metalloproteinases. Brain. 2001;124(11):2203–2214. doi: 10.1093/brain/124.11.2203. [DOI] [PubMed] [Google Scholar]
- 78.Bitsch A, Schuchardt J, Bunkowski S, Kuhlmann T, Brück W. Acute axonal injury in multiple sclerosis. Correlation with demyelination and inflammation. Brain. 2000;123(part 6):1174–1183. doi: 10.1093/brain/123.6.1174. [DOI] [PubMed] [Google Scholar]
- 79.DeLuca GC, Ebers GC, Esiri MM. Axonal loss in multiple sclerosis: a pathological survey of the corticospinal and sensory tracts. Brain. 2004;127(5):1009–1018. doi: 10.1093/brain/awh118. [DOI] [PubMed] [Google Scholar]
- 80.Trapp BD, Peterson J, Ransohoff RM, Rudick R, Mörk S, Bö L. Axonal transection in the lesions of multiple sclerosis. New England Journal of Medicine. 1998;338(5):278–285. doi: 10.1056/NEJM199801293380502. [DOI] [PubMed] [Google Scholar]
- 81.Kuhlmann T, Lingfeld G, Bitsch A, Schuchardt J, Brück W. Acute axonal damage in multiple sclerosis is most extensive in early disease stages and decreases over time. Brain. 2002;125(10):2202–2212. doi: 10.1093/brain/awf235. [DOI] [PubMed] [Google Scholar]
- 82.Bitsch A, Da Costa C, Bunkowski S, Weber F, Rieckmann P, Brück W. Identification of macrophage populations expressing tumor necrosis factor-α mRNA in acute multiple sclerosis. Acta Neuropathologica. 1998;95(4):373–377. doi: 10.1007/s004010050813. [DOI] [PubMed] [Google Scholar]
- 83.Koeberle PD, Ball AK. Nitric oxide synthase inhibition delays axonal degeneration and promotes the survival of axotomized retinal ganglion cells. Experimental Neurology. 1999;158(2):366–381. doi: 10.1006/exnr.1999.7113. [DOI] [PubMed] [Google Scholar]
- 84.Cuzner ML, Gveric D, Strand C, et al. The expression of tissue-type plasminogen activator, matrix metalloproteases and endogenous inhibitors in the central nervous system in multiple sclerosis: comparison of stages in lesion evolution. Journal of Neuropathology and Experimental Neurology. 1996;55(12):1194–1204. doi: 10.1097/00005072-199612000-00002. [DOI] [PubMed] [Google Scholar]
- 85.Liuzzi GM, Trojano M, Fanelli M, et al. Intrathecal synthesis of matrix metalloproteinase-9 in patients with multiple sclerosis: implication for pathogenesis. Multiple Sclerosis. 2002;8(3):222–228. doi: 10.1191/1352458502ms800oa. [DOI] [PubMed] [Google Scholar]
- 86.Leppert D, Ford J, Stabler G, et al. Matrix metalloproteinase-9 (gelatinase B) is selectively elevated in CSF during relapses and stable phases of multiple sclerosis. Brain. 1998;121(part 12):2327–2334. doi: 10.1093/brain/121.12.2327. [DOI] [PubMed] [Google Scholar]
- 87.Gijbels K, Galardy RE, Steinman L. Reversal of experimental autoimmune encephalomyelitis with a hydroxamate inhibitor of matrix metalloproteases. Journal of Clinical Investigation. 1994;94(6):2177–2182. doi: 10.1172/JCI117578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Hewson AK, Smith T, Leonard JP, Cuzner ML. Suppression of experimental allergic encephalomyelitis in the Lewis rat by the matrix metalloproteinase inhibitor Ro31-9790. Inflammation Research. 1995;44(8):345–349. doi: 10.1007/BF01796266. [DOI] [PubMed] [Google Scholar]
- 89.Clements JM, Cossins JA, Wells GM, et al. Matrix metalloproteinase expression during experimental autoimmune encephalomyelitis and effects of a combined matrix metalloproteinase and tumour necrosis factor-a inhibitor. Journal of Neuroimmunology. 1997;74(1):85–94. doi: 10.1016/s0165-5728(96)00210-x. [DOI] [PubMed] [Google Scholar]
- 90.Toft-Hansen H, Buist R, Sun XJ, Schellenberg A, Peeling J, Owens T. Metalloproteinases control brain inflammation induced by pertussis toxin in mice overexpressing the chemokine CCL2 in the central nervous system. Journal of Immunology. 2006;177(10):7242–7249. doi: 10.4049/jimmunol.177.10.7242. [DOI] [PubMed] [Google Scholar]
- 91.Corcoran ML, Stetler-Stevenson WG, Brown PD, Wahl LM. Interleukin 4 inhibition of prostaglandin E2 synthesis blocks interstitial collagenase and 92-kDa type IV collagenase/gelatinase production by human monocytes. Journal of Biological Chemistry. 1992;267(1):515–519. [PubMed] [Google Scholar]
- 92.Arthur FE, Shivers RR, Bowman PD. Astrocyte-mediated induction of tight junctions in brain capillary endothelium: an efficient in vitro model. Brain Research. 1987;433(1):155–159. doi: 10.1016/0165-3806(87)90075-7. [DOI] [PubMed] [Google Scholar]
- 93.Malik N, Greenfield BW, Wahl AF, Kiener PA. Activation of human monocytes through CD40 induces matrix metalloproteinases. Journal of Immunology. 1996;156(10):3952–3960. [PubMed] [Google Scholar]
- 94.Zhou H, Bernhard EJ, Fox FE, Billings PC. Induction of metalloproteinase activity in human T-lymphocytes. Biochimica et Biophysica Acta. 1993;1177(2):174–178. doi: 10.1016/0167-4889(93)90037-p. [DOI] [PubMed] [Google Scholar]
- 95.Harkness KAC, Adamson P, Sussman JD, Davies-Jones GAB, Greenwood J, Woodroofe MN. Dexamethasone regulation of matrix metalloproteinase expression in CNS vascular endothelium. Brain. 2000;123(part 4):698–709. doi: 10.1093/brain/123.4.698. [DOI] [PubMed] [Google Scholar]
- 96.Owens T, Renno T, Taupin V, Krakowski M. Inflammatory cytokines in the brain: does the CNS shape immune responses? Immunology Today. 1994;15(12):566–571. doi: 10.1016/0167-5699(94)90218-6. [DOI] [PubMed] [Google Scholar]
- 97.Opdenakker G, Masure S, Grillet B, van Damme J. Cytokine-mediated regulation of human leukocyte gelatinases and role in arthritis. Lymphokine and Cytokine Research. 1991;10(4):317–324. [PubMed] [Google Scholar]
- 98.Opdenakker G, Masure S, Proost P, Billiau A, van Damme J. Natural human monocyte gelatinase and its inhibitor. FEBS Letters. 1991;284(1):73–78. doi: 10.1016/0014-5793(91)80765-u. [DOI] [PubMed] [Google Scholar]
- 99.Johnatty RN, Taub DD, Reeder SP, et al. Cytokine and chemokine regulation of proMMP-9 and TIMP-1 production by human peripheral blood lymphocytes. Journal of Immunology. 1997;158(5):2327–2333. [PubMed] [Google Scholar]
- 100.Leppert D, Hauser SL, Kishiyama JL, An S, Zeng L, Goetzl EJ. Stimulation of matrix metalloproteinase-dependent migration of T cells by eicosanoids. FASEB Journal. 1995;9(14):1473–1481. doi: 10.1096/fasebj.9.14.7589989. [DOI] [PubMed] [Google Scholar]
- 101.Schrijver IA, Melief MJ, Markusse HM, et al. Peptidoglycan from sterile human spleen induces T-cell proliferation and inflammatory mediators in rheumatoid arthritis patients and healthy subjects. Rheumatology. 2001;40(4):438–446. doi: 10.1093/rheumatology/40.4.438. [DOI] [PubMed] [Google Scholar]
- 102.Gottschall PE, Yu X. Cytokines regulate gelatinase A and B (matrix metalloproteinase 2 and 9) activity in cultured rat astrocytes. Journal of Neurochemistry. 1995;64(4):1513–1520. doi: 10.1046/j.1471-4159.1995.64041513.x. [DOI] [PubMed] [Google Scholar]
- 103.Gottschall PE, Yu X, Bing B. Increased production of gelatinase B (matrix metalloproteinase-9) and interleukin-6 by activated rat microglia in culture. Journal of Neuroscience Research. 1995;42(3):335–342. doi: 10.1002/jnr.490420307. [DOI] [PubMed] [Google Scholar]
- 104.Liuzzi GM, Latronico T, Fasano A, Carlone G, Riccio P. Interferon-beta inhibits the expression of metalloproteinases in rat glial cell cultures: implications for multiple sclerosis pathogenesis and treatment. Multiple Sclerosis. 2004;10(3):290–297. doi: 10.1191/1352458504ms1016oa. [DOI] [PubMed] [Google Scholar]
- 105.Ma Z, Qin H, Benveniste EN. Transcriptional suppression of matrix metalloproteinase-9 gene expression by IFN-γ and IFN-β: critical role of STAT-1α . Journal of Immunology. 2001;167(9):5150–5159. doi: 10.4049/jimmunol.167.9.5150. [DOI] [PubMed] [Google Scholar]
- 106.Sancéau J, Boyd DD, Seiki M, Bauvois B. Interferons inhibit tumor necrosis factor-α-mediated matrix metalloproteinase-9 activation via interferon regulatory factor-1 binding competition with NF-κB. Journal of Biological Chemistry. 2002;277(38):35766–35775. doi: 10.1074/jbc.M202959200. [DOI] [PubMed] [Google Scholar]
- 107.Cuzner ML, Opdenakker G. Plasminogen activators and matrix metalloproteases, mediators of extracellular proteolysis in inflammatory demyelination of the central nervous system. Journal of Neuroimmunology. 1999;94(1-2):1–14. doi: 10.1016/s0165-5728(98)00241-0. [DOI] [PubMed] [Google Scholar]
- 108.Stuve Dooley NP O, Dooley NP, Uhm JH, et al. Interferon beta-1b decreases the migration of T lymphocytes in vitro: effects on matrix metalloproteinase-9. Annals of Neurology. 1996;40(6):853–863. doi: 10.1002/ana.410400607. [DOI] [PubMed] [Google Scholar]
- 109.Bartholomé EJ, van Aelst I, Koyen E, et al. Human monocyte-derived dendritic cells produce bioactive gelatinase B: inhibition by IFN-beta. Journal of Interferon & Cytokine Research. 2001;21(7):495–501. doi: 10.1089/10799900152434367. [DOI] [PubMed] [Google Scholar]
- 110.Trojano M, Avolio C, Liuzzi GM, et al. Changes of serum sICAM-1 and MMP-9 induced by rIFNβ-1b treatment in relapsing-remitting MS. Neurology. 1999;53(7):1402–1408. doi: 10.1212/wnl.53.7.1402. [DOI] [PubMed] [Google Scholar]
- 111.Ozenci Kouwenhoven M V, Teleshova N, Pashenkov M, Fredrikson S, Link H. Multiple sclerosis: pro- and anti-inflammatory cytokines and metalloproteinases are affected differentially by treatment with IFN-beta. Journal of Neuroimmunology. 2000;108(1):236–243. doi: 10.1016/s0165-5728(00)00281-2. [DOI] [PubMed] [Google Scholar]
- 112.Shapiro S, Shoenfeld Y, Gilburd B, Sobel E, Lahat N. Intravenous gamma globulin inhibits the production of matrix metalloproteinase-9 in macrophages. Cancer. 2002;95(9):2032–2037. doi: 10.1002/cncr.10905. [DOI] [PubMed] [Google Scholar]
- 113.Vaillant C, Didier-Bazès M, Hutter A, Belin MF, Thomasset N. Spatiotemporal expression patterns of metalloproteinases and their inhibitors in the postnatal developing rat cerebellum. Journal of Neuroscience. 1999;19(12):4994–5004. doi: 10.1523/JNEUROSCI.19-12-04994.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Soler RC, Gui YH, Linask KK, Muschel RJ. MMP-9 (gelatinase B) mRNA is expressed during mouse neurogenesis and may be associated with vascularization. Developmental Brain Research. 1995;88(1):37–52. doi: 10.1016/0165-3806(95)00079-s. [DOI] [PubMed] [Google Scholar]
- 115.Uhm JH, Dooley NP, Oh LY, Yong VW. Oligodendrocytes utilize a matrix metalloproteinase, MMP-9, to extend processes along an astrocyte extracellular matrix. Glia. 1998;22(1):53–63. doi: 10.1002/(sici)1098-1136(199801)22:1<53::aid-glia5>3.0.co;2-9. [DOI] [PubMed] [Google Scholar]
- 116.Oh LYS, Larsen PH, Krekoski CA, et al. Matrix metalloproteinase-9/gelatinase B is required for process outgrowth by oligoclendrocytes. Journal of Neuroscience. 1999;19(19):8464–8475. doi: 10.1523/JNEUROSCI.19-19-08464.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Yu F, Kamada H, Niizuma K, Endo H, Chan PH. Induction of MMP-9 expression and endothelial injury by oxidative stress after spinal cord injury. Journal of Neurotrauma. 2008;25(3):184–195. doi: 10.1089/neu.2007.0438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Buss Pech K A, Kakulas BA, Martin D, Schoenen J, Noth J, Brook GA. Matrix metalloproteinases and their inhibitors in human traumatic spinal cord injury. BMC Neurology. 2007;7(article 17) doi: 10.1186/1471-2377-7-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Larsen PH, Wells JE, Stallcup WB, Opdenakker G, Yong VW. Matrix metalloproteinase-9 facilitates remyelination in part by processing the inhibitory NG2 proteoglycan. Journal of Neuroscience. 2003;23(35):11127–11135. doi: 10.1523/JNEUROSCI.23-35-11127.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Larsen PH, Yong VW. The expression of matrix metalloproteinase-12 by oligodendrocytes regulates their maturation and morphological differentiation. Journal of Neuroscience. 2004;24(35):7597–7603. doi: 10.1523/JNEUROSCI.2092-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Larsen PH, DaSilva AG, Conant K, Yong VW. Myelin formation during development of the CNS is delayed in matrix metalloproteinase-9 and -12 null mice. Journal of Neuroscience. 2006;26(8):2207–2214. doi: 10.1523/JNEUROSCI.1880-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Hsu JYC, McKeon R, Goussev S, et al. Matrix metalloproteinase-2 facilitates wound healing events that promote functional recovery after spinal cord injury. Journal of Neuroscience. 2006;26(39):9841–9850. doi: 10.1523/JNEUROSCI.1993-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Haorah J, Ramirez SH, Schall K, Smith D, Pandya R, Persidsky Y. Oxidative stress activates protein tyrosine kinase and matrix metalloproteinases leading to blood-brain barrier dysfunction. Journal of Neurochemistry. 2007;101(2):566–576. doi: 10.1111/j.1471-4159.2006.04393.x. [DOI] [PubMed] [Google Scholar]
- 124.Luckow B, Joergensen J, Chilla S, et al. Reduced intragraft mRNA expression of matrix metalloproteinases Mmp3, Mmp12, Mmp13 and Adam8, and diminished transplant arteriosclerosis in Ccr5-deficient mice. European Journal of Immunology. 2004;34(9):2568–2578. doi: 10.1002/eji.200324776. [DOI] [PubMed] [Google Scholar]
- 125.Woo MS, Park JS, Choi IY, Kimf WK, Kim HS. Inhibition of MMP-3 or -9 suppresses lipopolysaccharide-induced expression of proinflammatory cytokines and iNOS in microglia. Journal of Neurochemistry. 2008;106(2):770–780. doi: 10.1111/j.1471-4159.2008.05430.x. [DOI] [PubMed] [Google Scholar]
- 126.Lassmann H, Zimprich F, Vass K, Hickey WF. Microglial cells are a component of the perivascular glia limitans. Journal of Neuroscience Research. 1991;28(2):236–243. doi: 10.1002/jnr.490280211. [DOI] [PubMed] [Google Scholar]
- 127.Ethell IM, Ethell DW. Matrix metalloproteinases in brain development and remodeling: synaptic functions and targets. Journal of Neuroscience Research. 2007;85(13):2813–2823. doi: 10.1002/jnr.21273. [DOI] [PubMed] [Google Scholar]
- 128.Nagy Bozdagi O V, Bozdagi O, Bozdagi O, et al. Matrix metalloproteinase-9 is required for hippocampal late-phase long-term potentiation and memory. Journal of Neuroscience. 2006;26(7):1923–1934. doi: 10.1523/JNEUROSCI.4359-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Holland EC. Glioblastoma multiforme: the terminator. Proceedings of the National Academy of Sciences. 2000;97(12):6242–6244. doi: 10.1073/pnas.97.12.6242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Takakura K, Abe H, Tanaka R. Effects of ACNU and radiotherapy on malignant glioma. Journal of Neurosurgery. 1986;64(1):53–57. doi: 10.3171/jns.1986.64.1.0053. [DOI] [PubMed] [Google Scholar]
- 131.Shapiro WR, Green SB, Burger PC, et al. A randomized comparison of intra-arterial versus intravenous BCNU, with or without intravenous 5-fluorouracil, for newly diagnosed patients with malignant glioma. Journal of Neurosurgery. 1992;76(5):772–781. doi: 10.3171/jns.1992.76.5.0772. [DOI] [PubMed] [Google Scholar]
- 132.Stupp R, Mason WP, van den Bent MJ, et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. New England Journal of Medicine. 2005;352(10):987–996. doi: 10.1056/NEJMoa043330. [DOI] [PubMed] [Google Scholar]
- 133.van Meir EG, Hadjipanayis CG, Norden AD, Shu HK, Wen PY, Olson JJ. Exciting new advances in neuro-oncology: the avenue to a cure for malignant glioma. CA Cancer Journal for Clinicians. 2010;60(3):166–193. doi: 10.3322/caac.20069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Rio-hortega P. Sobre la fagocitosis en los tumores y en otros procesos patológicos/P. del Río-Hortega. Archivos de Cardiología y Hematología. 1921;2(5):161–220. [Google Scholar]
- 135.Graeber MB, Scheithauer BW, Kreutzberg GW. Microglia in brain tumors. GLIA. 2002;40(2):252–259. doi: 10.1002/glia.10147. [DOI] [PubMed] [Google Scholar]
- 136.Badie B, Schartner J. Role of microglia in glioma biology. Microscopy Research and Technique. 2001;54(2):106–113. doi: 10.1002/jemt.1125. [DOI] [PubMed] [Google Scholar]
- 137.Ribot E, Bouzier-Sore AK, Bouchaud V, et al. Microglia used as vehicles for both inducible thymidine kinase gene therapy and MRI contrast agents for glioma therapy. Cancer Gene Therapy. 2007;14(8):724–737. doi: 10.1038/sj.cgt.7701060. [DOI] [PubMed] [Google Scholar]
- 138.Markovic DS, Vinnakota K, Chirasani S, et al. Gliomas induce and exploit microglial MT1-MMP expression for tumor expansion. Proceedings of the National Academy of Sciences of the United States of America. 2009;106(30):12530–12535. doi: 10.1073/pnas.0804273106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Markovic DS, Glass R, Synowitz M, van Rooijen N, Kettenmann H. Microglia stimulate the invasiveness of glioma cells by increasing the activity of metalloprotease-2. Journal of Neuropathology and Experimental Neurology. 2005;64(9):754–762. doi: 10.1097/01.jnen.0000178445.33972.a9. [DOI] [PubMed] [Google Scholar]
- 140.Watters JJ, Schartner JM, Badie B. Microglia function in brain tumors. Journal of Neuroscience Research. 2005;81(3):447–455. doi: 10.1002/jnr.20485. [DOI] [PubMed] [Google Scholar]
- 141.Kerber M, Reiss Y, Wickersheim A, et al. Flt-1 signaling in macrophages promotes glioma growth in vivo. Cancer Research. 2008;68(18):7342–7351. doi: 10.1158/0008-5472.CAN-07-6241. [DOI] [PubMed] [Google Scholar]
- 142.Zhai H, Heppner FL, Tsirka SE. Microglia/macrophages promote glioma progression. GLIA. 2011;59(3):472–485. doi: 10.1002/glia.21117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Sliwa M, Markovic D, Gabrusiewicz K, et al. The invasion promoting effect of microglia on glioblastoma cells is inhibited by cyclosporin A. Brain. 2007;130(2):476–489. doi: 10.1093/brain/awl263. [DOI] [PubMed] [Google Scholar]
- 144.Grasbon-Frodl EMFA WP, Klinkert WEF, Kreutzberg GW, Graeber MB. Jahrestagung der Neuroonkologischen Arbeitsgemeinschaft der Deutschen Gesellschaft fuer Neurochirurgie in Dresden. 1998. [Google Scholar]
- 145.Huettner C, Czub S, Kerkau S, Roggendorf W, Tonn JC. Interleukin 10 is expressed in human gliomas in vivo and increases glioma cell proliferation and motility in vitro. Anticancer Research. 1997;17(5 A):3217–3224. [PubMed] [Google Scholar]
- 146.Wagner S, Czub S, Greif M, et al. Microglial/macrophage expression of interleukin 10 in human glioblastomas. International Journal of Cancer. 1999;82(1):12–16. doi: 10.1002/(sici)1097-0215(19990702)82:1<12::aid-ijc3>3.0.co;2-o. [DOI] [PubMed] [Google Scholar]
- 147.Frei K, Siepl C, Groscurth P, Bodmer S, Schwerdel C, Fontana A. Antigen presentation and tumor cytotoxicity by interferon-γ-treated microglial cells. European Journal of Immunology. 1987;17(9):1271–1278. doi: 10.1002/eji.1830170909. [DOI] [PubMed] [Google Scholar]
- 148.Sutter A, Hekmat A, Luckenbach GA. Antibody-mediated tumor cytotoxicity of microglia. Pathobiology. 1991;59(4):254–258. doi: 10.1159/000163657. [DOI] [PubMed] [Google Scholar]
- 149.Voisin P, Bouchaud V, Merle M, et al. Microglia in close vicinity of glioma cells: correlation between phenotype and metabolic alterations. Frontiers in Neuroenergetics. 2010;2(article 131) doi: 10.3389/fnene.2010.00131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Hussain SF, Yang D, Suki D, Aldape K, Grimm E, Heimberger AB. The role of human glioma-infiltrating microglia/macrophages in mediating antitumor immune responses. Neuro-Oncology. 2006;8(3):261–279. doi: 10.1215/15228517-2006-008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Rao JS. Molecular mechanisms of glioma invasiveness: the role of proteases. Nature Reviews Cancer. 2003;3:489–501. doi: 10.1038/nrc1121. [DOI] [PubMed] [Google Scholar]
- 152.Hanisch UK, Kettenmann H. Microglia: active sensor and versatile effector cells in the normal and pathologic brain. Nature Neuroscience. 2007;10(11):1387–1394. doi: 10.1038/nn1997. [DOI] [PubMed] [Google Scholar]
- 153.Liotta LA, Tryggvason K, Garbisa S. Metastatic potential correlates with enzymatic degradation of basement membrane collagen. Nature. 1980;284(5751):67–68. doi: 10.1038/284067a0. [DOI] [PubMed] [Google Scholar]
- 154.Bjorklund M, Koivunen E. Gelatinase-mediated migration and invasion of cancer cells. Biochimica et Biophysica Acta. 2005;1755(1):37–69. doi: 10.1016/j.bbcan.2005.03.001. [DOI] [PubMed] [Google Scholar]
- 155.Fingleton B. Matrix metalloproteinases: roles in cancer and metastasis. Frontiers in Bioscience. 2006;11:479–491. doi: 10.2741/1811. [DOI] [PubMed] [Google Scholar]
- 156.Colotti C, Angeli V, del Ry S, Maltinti M, Vittorini S, Giannessi D. Matrix metalloprotease-2 and -9 concentration and activity in serum and culture medium samples: a methodological reappraisal. Clinical Chemistry and Laboratory Medicine. 2007;45(10):1292–1298. doi: 10.1515/CCLM.2007.283. [DOI] [PubMed] [Google Scholar]
- 157.Patel S, Sumitra G, Koner BC, Saxena A. Role of serum matrix metalloproteinase-2 and -9 to predict breast cancer progression. Clinical Biochemistry. 2011;44(10-11):869–872. doi: 10.1016/j.clinbiochem.2011.04.019. [DOI] [PubMed] [Google Scholar]
- 158.Turpeenniemi-Hujanen T. Gelatinases (MMP-2 and -9) and their natural inhibitors as prognostic indicators in solid cancers. Biochimie. 2005;87(3-4):287–297. doi: 10.1016/j.biochi.2005.01.014. [DOI] [PubMed] [Google Scholar]
- 159.Liu L, Wu J, Ying Z, et al. Astrocyte elevated gene-1 upregulates matrix metalloproteinase-9 and induces human glioma invasion. Cancer Research. 2010;70(9):3750–3759. doi: 10.1158/0008-5472.CAN-09-3838. [DOI] [PubMed] [Google Scholar]
- 160.Sbardella D, Fasciglione GF, Gioia M, et al. Human matrix metalloproteinases: an ubiquitarian class of enzymes involved in several pathological processes. Molecular Aspects of Medicine. 2012;33(2):119–208. doi: 10.1016/j.mam.2011.10.015. [DOI] [PubMed] [Google Scholar]
- 161.Aresu L, Benali S, Garbisa S, Gallo E, Castagnaro M. Matrix metalloproteinases and their role in the renal epithelial mesenchymal transition. Histology and Histopathology. 2011;26(3):307–313. doi: 10.14670/HH-26.307. [DOI] [PubMed] [Google Scholar]
- 162.Ribeiro AS, Albergaria A, Sousa B, et al. Extracellular cleavage and shedding of P-cadherin: a mechanism underlying the invasive behaviour of breast cancer cells. Oncogene. 2010;29(3):392–402. doi: 10.1038/onc.2009.338. [DOI] [PubMed] [Google Scholar]
- 163.Levi E, Fridman R, Miao HQ, Ma YS, Yayon A, Vlodavsky I. Matrix metalloproteinase 2 releases active soluble ectodomain of fibroblast growth factor receptor 1. Proceedings of the National Academy of Sciences. 1996;93(14):7069–7074. doi: 10.1073/pnas.93.14.7069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Gondi CS, Dinh DH, Klopffenstein JD, Gujrati M, Rao JS. MMP-2 downregulation mediates differential regulation of cell death via ErbB-2 in glioma xenografts. International Journal of Oncology. 2009;35(2):257–263. [PMC free article] [PubMed] [Google Scholar]
- 165.Badiga AV, Chetty C, Kesanakurti D, et al. MMP-2 siRNA inhibits radiation-enhanced invasiveness in glioma cells. PLoS ONE. 2011;6(6) doi: 10.1371/journal.pone.0020614.e20614 [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 166.Jo YK, Park SJ, Shin JH, et al. ARP101, a selective MMP-2 inhibitor, induces autophagy-associated cell death in cancer cells. Biochemical and Biophysical Research Communications. 2011;404(4):1039–1043. doi: 10.1016/j.bbrc.2010.12.106. [DOI] [PubMed] [Google Scholar]
- 167.Lampert K, Machein U, Machein MR, Conca W, Peter HH, Volk B. Expression of matrix metalloproteinases and their tissue inhibitors in human brain tumors. American Journal of Pathology. 1998;153(2):429–437. doi: 10.1016/S0002-9440(10)65586-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Yamamoto M, Mohanam S, Sawaya R, et al. Differential expression of membrane-type matrix metalloproteinase and its correlation with gelatinase A activation in human malignant brain tumors in vivo and in vitro. Cancer Research. 1996;56(2):384–392. [PubMed] [Google Scholar]
- 169.Forsyth PA, Wong H, Laing TD, et al. Gelatinase-A (MMP-2), gelatinase-B (MMP-9) and membrane type matrix metalloproteinase-1 (MT1-MMP) are involved in different aspects of the pathophysiology of malignant gliomas. British Journal of Cancer. 1999;79(11-12):1828–1835. doi: 10.1038/sj.bjc.6990291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Estève PO, Chicoine E, Robledo O, et al. Protein kinase C-zeta regulates transcription of the matrix metalloproteinase-9 gene induced by IL-1 and TNF-alpha in glioma cells via NF-kappa B. Journal of Biological Chemistry. 2002;277(38):35150–35155. doi: 10.1074/jbc.M108600200. [DOI] [PubMed] [Google Scholar]
- 171.Rao JS, Yamamoto M, Mohaman S, et al. Expression and localization of 92 kDa type IV collagenase/gelatinase B (MMP-9) in human gliomas. Clinical & Experimental Metastasis. 1996;14(1):12–18. doi: 10.1007/BF00157681. [DOI] [PubMed] [Google Scholar]
- 172.Guo P, Imanishi Y, Cackowski FC, et al. Up-regulation of angiopoietin-2, matrix metalloprotease-2, membrane type 1 metalloprotease, and laminin 5 gamma 2 correlates with the invasiveness of human glioma. The American Journal of Pathology. 2005;166(3):877–890. doi: 10.1016/s0002-9440(10)62308-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Yamada T, Yoshiyama Y, Sato H, Seiki M, Shinagawa A, Takahashi M. White matter microglia produce membrane-type matrix metalloprotease, an activator of gelatinase A, in human brain tissues. Acta Neuropathologica. 1995;90(5):421–424. doi: 10.1007/BF00294800. [DOI] [PubMed] [Google Scholar]
- 174.Ghosh A, Chaudhuri S. Microglial action in glioma: a boon turns bane. Immunology Letters. 2010;131(1):3–9. doi: 10.1016/j.imlet.2010.03.003. [DOI] [PubMed] [Google Scholar]
- 175.Wisniewski P, Ellert-Miklaszewska A, Kwiatkowska A, Kaminska B. Non-apoptotic Fas signaling regulates invasiveness of glioma cells and modulates MMP-2 activity via NFκB-TIMP-2 pathway. Cellular Signalling. 2010;22(2):212–220. doi: 10.1016/j.cellsig.2009.09.016. [DOI] [PubMed] [Google Scholar]
- 176.Baumann F, Leukel P, Doerfelt A, et al. Lactate promotes glioma migration by TGF-β2–dependent regulation of matrix metalloproteinase-2. Neuro-Oncology. 2009;11(4):368–380. doi: 10.1215/15228517-2008-106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Wick W, Platten M, Weller M. Glioma cell invasion: regulation of metalloproteinase activity by TGF-β . Journal of Neuro-Oncology. 2001;53(2):177–185. doi: 10.1023/a:1012209518843. [DOI] [PubMed] [Google Scholar]
- 178.Nakao A, Imamura T, Souchelnytskyi S, et al. TGF-β receptor-mediated signalling through Smad2, Smad3 and Smad4. EMBO Journal. 1997;16(17):5353–5362. doi: 10.1093/emboj/16.17.5353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Imai K, Hiramatsu A, Fukushima D, Pierschbacher MD, Okada Y. Degradation of decorin by matrix metalloproteinases: identification of the cleavage sites, kinetic analyses and transforming growth factor-β1 release. Biochemical Journal. 1997;322(part 3):809–814. doi: 10.1042/bj3220809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Yu Q, Stamenkovic I. Cell surface-localized matrix metalloproteinase-9 proteolytically activates TGF-β and promotes tumor invasion and angiogenesis. Genes and Development. 2000;14(2):163–176. [PMC free article] [PubMed] [Google Scholar]
- 181.Binker MG, Binker-Cosen AA, Gaisano HY, de Cosen RH, Cosen-Binker LI. TGF-β1 increases invasiveness of SW1990 cells through Rac1/ROS/NF-κB/IL-6/MMP-2. Biochemical and Biophysical Research Communications. 2011;405(1):140–145. doi: 10.1016/j.bbrc.2011.01.023. [DOI] [PubMed] [Google Scholar]
- 182.Kim ES, Sohn YW, Moon A. TGF-β-induced transcriptional activation of MMP-2 is mediated by activating transcription factor (ATF)2 in human breast epithelial cells. Cancer Letters. 2007;252(1):147–156. doi: 10.1016/j.canlet.2006.12.016. [DOI] [PubMed] [Google Scholar]
- 183.Dasgupta A, Raychaudhuri B, Haqqi T, et al. Stat3 activation is required for the growth of U87 cell-derived tumours in mice. European Journal of Cancer. 2009;45(4):677–684. doi: 10.1016/j.ejca.2008.11.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Lo HW, Cao X, Zhu H, Ali-Osman F. Constitutively activated STAT3 frequently coexpresses with epidermal growth factor receptor in high-grade gliomas and targeting STAT3 sensitizes them to iressa and alkylators. Clinical Cancer Research. 2008;14(19):6042–6054. doi: 10.1158/1078-0432.CCR-07-4923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Mizoguchi M, Betensky RA, Batchelor TT, Bernay DC, Louis DN, Nutt CL. Activation of STAT3, MAPK, and AKT in malignant astrocytic gliomas: correlation with EGFR status, tumor grade, and survival. Journal of Neuropathology and Experimental Neurology. 2006;65(12):1181–1188. doi: 10.1097/01.jnen.0000248549.14962.b2. [DOI] [PubMed] [Google Scholar]
- 186.Rahaman SO, Harbor PC, Chernova O, Barnett GH, Vogelbaum MA, Haque SJ. Inhibition of constitutively active Stat3 suppresses proliferation and induces apoptosis in glioblastoma multiforme cells. Oncogene. 2002;21(55):8404–8413. doi: 10.1038/sj.onc.1206047. [DOI] [PubMed] [Google Scholar]
- 187.Schaefer LK, Ren Z, Fuller GN, Schaefer TS. Constitutive of Stat3α in brain tumors: localization to tumor endothelial cells and activation by the endothelial tyrosine kinase receptor (VEGFR-2) Oncogene. 2002;21(13):2058–2065. doi: 10.1038/sj.onc.1205263. [DOI] [PubMed] [Google Scholar]
- 188.Chen SH, Gillespie GY, Benveniste EN. Divergent effects of oncostatin M on astroglioma cells: influence on cell proliferation, invasion, and expression of matrix metalloproteinases. GLIA. 2006;53(2):191–200. doi: 10.1002/glia.20264. [DOI] [PubMed] [Google Scholar]
- 189.Repovic P, Fears CY, Gladson CL, Benveniste EN. Oncostatin-M induction of vascular endothelial growth factor expression in astroglioma cells. Oncogene. 2003;22(50):8117–8124. doi: 10.1038/sj.onc.1206922. [DOI] [PubMed] [Google Scholar]
- 190.Held-Feindt J, Hattermann K, Müerköster SS, et al. CX3CR1 promotes recruitment of human glioma-infiltrating microglia/macrophages (GIMs) Experimental Cell Research. 2010;316(9):1553–1566. doi: 10.1016/j.yexcr.2010.02.018. [DOI] [PubMed] [Google Scholar]
- 191.Yabkowitz R, Meyer S, Black T, Elliott G, Merewether LA, Yamane HK. Inflammatory cytokines and vascular endothelial growth factor stimulate the release of soluble tie receptor from human endothelial cells via metalloprotease activation. Blood. 1999;93(6):1969–1979. [PubMed] [Google Scholar]
- 192.Wang H, Lathia JD, Wu Q, et al. Targeting interleukin 6 signaling suppresses glioma stem cell survival and tumor growth. Stem Cells. 2009;27(10):2393–2404. doi: 10.1002/stem.188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Li R, Li G, Deng L, et al. IL-6 augments the invasiveness of U87MG human glioblastoma multiforme cells via up-regulation of MMP-2 and fascin-1. Oncology Reports. 2010;23(6):1553–1559. doi: 10.3892/or_00000795. [DOI] [PubMed] [Google Scholar]
- 194.Johnston JB, Silva C, Gonzalez G, et al. Diminished adenosine A1 receptor expression on macrophages in brain and blood of patients with multiple sclerosis. Annals of Neurology. 2001;49(5):650–658. [PubMed] [Google Scholar]
- 195.Tsutsui S, Schnermann J, Noorbakhsh F, et al. A1 Adenosine Receptor Upregulation and Activation Attenuates Neuroinflammation and Demyelination in a Model of Multiple Sclerosis. Journal of Neuroscience. 2004;24(6):1521–1529. doi: 10.1523/JNEUROSCI.4271-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Beliveau A, Mott JD, Lo A, et al. Raf-induced MMP9 disrupts tissue architecture of human breast cells in three-dimensional culture and is necessary for tumor growth in vivo. Genes and Development. 2010;24(24):2800–2811. doi: 10.1101/gad.1990410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Lin CC, Kuob CT, Cheng CY, et al. IL-1β promotes A549 cell migration via MAPKs/AP-1- and NF-κB-dependent matrix metalloproteinase-9 expression. Cellular Signalling. 2009;21(11):1652–1662. doi: 10.1016/j.cellsig.2009.07.002. [DOI] [PubMed] [Google Scholar]
- 198.Coulson-Thomas VJ, Gesteira TF, Coulson-Thomas YM, et al. Fibroblast and prostate tumor cell cross-talk: fibroblast differentiation, TGF-β, and extracellular matrix down-regulation. Experimental Cell Research. 2010;316(19):3207–3226. doi: 10.1016/j.yexcr.2010.08.005. [DOI] [PubMed] [Google Scholar]
- 199.Kumar B, Koul S, Petersen J, et al. p38 mitogen-activated protein kinase-driven MAPKAPK2 regulates invasion of bladder cancer by modulation of MMP-2 and MMP-9 activity. Cancer Research. 2010;70(2):832–841. doi: 10.1158/0008-5472.CAN-09-2918. [DOI] [PubMed] [Google Scholar]
- 200.Ehrenfeld P, Conejeros I, Pavicic MF, et al. Activation of kinin B1 receptor increases the release of metalloproteases-2 and -9 from both estrogen-sensitive and -insensitive breast cancer cells. Cancer Letters. 2011;301(1):106–118. doi: 10.1016/j.canlet.2010.09.020. [DOI] [PubMed] [Google Scholar]
- 201.Oh J, Takahashi R, Kondo S, et al. The membrane-anchored MMP inhibitor RECK is a key regulator of extracellular matrix integrity and angiogenesis. Cell. 2001;107(6):789–800. doi: 10.1016/s0092-8674(01)00597-9. [DOI] [PubMed] [Google Scholar]
- 202.Coussens LM, Fingleton B, Matrisian LM. Matrix metalloproteinase inhibitors and cancer: trials and tribulations. Science. 2002;295(5564):2387–2392. doi: 10.1126/science.1067100. [DOI] [PubMed] [Google Scholar]
- 203.Overall CM, López-Otín C. Strategies for MMP inhibition in cancer: innovations for the post-trial era. Nature Reviews Cancer. 2002;2(9):657–672. doi: 10.1038/nrc884. [DOI] [PubMed] [Google Scholar]