

Abstract
Recent studies in the SIV-macaque model of HIV infection suggest that Nef-specific CD8+ T-cell responses may mediate highly effective immune control of viraemia. In HIV infection Nef recognition dominates in acute infection, but in large cohort studies of chronically infected subjects, breadth of T cell responses to Nef has not been correlated with significant viraemic control. Improved disease outcomes have instead been associated with targeting Gag and, in some cases, Pol. However analyses of the breadth of Nef-specific T cell responses have been confounded by the extreme immunogenicity and multiple epitope overlap within the central regions of Nef, making discrimination of distinct responses impossible via IFN-gamma ELISPOT assays. Thus an alternative approach to assess Nef as an immune target is needed. Here, we show in a cohort of >700 individuals with chronic C-clade infection that >50% of HLA-B-selected polymorphisms within Nef are associated with a predicted fitness cost to the virus, and that HLA-B alleles that successfully drive selection within Nef are those linked with lower viral loads. Furthermore, the specific CD8+ T cell epitopes that are restricted by protective HLA Class I alleles correspond substantially to effective SIV-specific epitopes in Nef. Distinguishing such individual HIV-specific responses within Nef requires specific peptide-MHC I tetramers. Overall, these data suggest that CD8+ T cell targeting of certain specific Nef epitopes contributes to HIV suppression. These data suggest that a re-evaluation of the potential use of Nef in HIV T-cell vaccine candidates would be justified.
Introduction
The HIV-1 nef gene encodes a polymorphic 27kda protein, 200-215 amino acids in length [1,2]. Nef has a complex role in HIV pathogenicity via a number of mechanisms, including down-regulation of host CD4 and MHC cell surface expression, modulation of T cell function, and altering of macrophage signaling [2–4]. Nef is among the most diverse HIV proteins [5]. The greatest sequence variability is focused in the amino- (N-) and carboxy- (C-) terminal regions, while the central portion of the protein is substantially more conserved [6–8]. At least some of this sequence variability may be driven by Class I HLA selection pressure [9,10].
Nef has a high density of overlapping CD8+ T cell epitopes (see Nef epitope map at www.hiv.lanl.gov), and multiple copies of Nef are produced early in the HIV life cycle [11,12]. Nef is the most targeted protein in acute infection [13–15], accounting for 50% to 90% of CD8+ T cell responses in acutely infected subjects [16,17], as well as having the most CD8+ T cell responses and the highest magnitude IFN-gamma responses in chronic infection [7].
Large cross-sectional studies have shown no correlation between viraemic suppression and either the breadth of CD8+ T cell IFN-gamma responses to Nef [18], or the number of HLA-selected mutations in Nef [19]. However the immunogenicity of Nef and consequent density of overlapping epitopes confounds analyses of the distinct CTL responses targeting this protein that have typically depended on ELISpot assays using panels of overlapping peptides 15-18 amino acids in length. Other studies have attempted to address the issue of Nef diversity by using extended panels of overlapping peptides (e.g. 10-mers overlapping by 9 amino acids, with all common variants represented [20]) but this approach is costly and time-consuming.
The HLA Class I alleles that are most strongly associated with long-term suppression of viraemia (e.g. HLA-B*57, -B*58:01 and -B*27 [21–23]) present CD8+ T cell epitopes in Gag and Pol that have been especially well-studied [19,24,25]. The targeting of these highly conserved proteins is thought to underpin successful immunological control [26,27]. However these HLA class I molecules also present epitopes within Nef that may be important in mediating disease control. In a host expressing these protective HLA alleles, the virus adapts by selecting escape mutations in all Gag, Pol and Nef epitopes [28,29], underlining a survival benefit to the virus incurred through evasion of these responses. Indeed, some studies have suggested that viraemic suppression may be linked to specific Nef epitopes, including HLA-B*57/58:01-HW9 [30] and HLA-B*35: 01-VY8 [31].
Several further lines of evidence pointing towards the potential immunological benefit of targeting HIV-Nef have been reported. Many HLA-selected escape mutations in Nef revert to wild-type following transmission to an HLA-mismatched host, suggesting a fitness cost in association with the mutation [9,19]. This hypothesis is supported first by studies showing that Nef polymorphisms are more common among slow progressors than rapid progressors [6], and second by the finding that such sequence changes in elite controllers are associated with a detriment to Nef function [32].
Studies in the SIV model add further weight to the hypothesis that Nef responses can contribute to disease control. In the rhesus macaque, Mamu-B*08, which has a peptide-binding motif so similar to HLA-B*27 that it can bind the same peptides [33], is also associated with elite viraemic control [34–36]. However, unlike HLA-B*27 [25], the immunodominant responses are within accessory/regulatory proteins, and a vaccine induced Mamu-B*08-restricted response to the Nef RL10 epitope (SIV Nef 137–146, RRHRILDIYL) is correlated with viraemic control [34]. This epitope is homologous to the HIV-specific HLA-B*27: 05 RI10 epitope, (HIV Nef 105-114, RRQDILDLWI [37]). Likewise, macaques expressing another favourable allele, Mamu-B*17, also target Nef via two immunodominant epitopes, IW9 (SIV Nef 165-173, IRYPKTFGW) and MW9 (SIV Nef 195-203, MHPAQTSQW) [38,39]. In Mauritian Cynomolgus Macaques, controllers of SIV replication have been shown to target epitopes located near the carboxy terminus of the SIV Nef protein [40]. Recently, a novel protective MHC-I haplotype was described in Burmese macaques, 90-010-Id (D), which also mediates immune control against SIV through Nef responses [41].
The starting point for these current studies, therefore, was the conflict in evidence regarding the extent to which CD8+ T cell responses to Nef might impact upon HIV-1 disease control. On the one hand, large cross-sectional studies have suggested no benefit in extended breadth of Nef targeting [18,19], whilst on the other, recent data suggest that targeting particular regions of Nef might contribute fundamentally to viraemic control [29–32,34,38,39].
Most previous analyses of Nef diversity and HLA-selection have focused on B-clade infections [12,29,42,43], while studies of C-clade have often been limited by small sample numbers [44,45]. We focused here on an extended cohort of >700 C-clade infected adults to identify relationships between CD8+ T cell responses to Nef and HIV disease control. We hypothesized that certain specific Nef responses may help to underpin viraemic control, strengthening the argument for the inclusion of Nef as an immunogen in CD8+ T cell vaccines.
Materials and Methods
Recruitment and characterization of HIV-infected individuals
We studied 739 adults with chronic HIV-1 C-clade infection recruited from three cohorts: Durban, South Africa (n=436), Gaborone, Botswana (n=275), and Zimbabwean subjects recruited via the Thames Valley Cohort in the UK (n=28), as previously described [46,47]. Ethics approval was given by KwaZulu-Natal Review Board and the Massachusetts General Hospital Review Board (Durban cohort); the Office of Human Research Administration, Harvard School of Public Health and the Health Research Development Committee, Botswana Ministry of Health (Gaborone cohort); and the Oxford Research Ethics Committee (Durban, Gaborone and Thames Valley cohorts). All subjects provided written informed consent.
Nef sequences and HLA-types were generated as previously described [19,48]. Viral loads were performed using the Roche Amplicor v.1.5 assay. Viral load measurements were made in chronic disease (therefore reflecting set-point viraemia).
Identification of Nef sequence polymorphisms associated with Class I HLA genotype
We used computational methods, as previously described [19,49], to identify sites of HLA-selected polymorphism in Nef sequences, correcting for viral lineage and for linkage disequilibrium between HLA Class I alleles. We corrected for multiple comparisons using a q-value (false discovery rate, FDR [50]), and a cut-off value of q<0.05(5% FDR). Reversion was predicted, as previously, by the statistical association between the absence of a particular HLA allele with either (i) an increase in expression of the consensus amino acid, or (ii) a decrease in expression of a variant amino acid [19]. These associations demonstrate that a mutation selected by a particular HLA allele returns to consensus following transmission to an HLA-mismatched host, from which a fitness cost is inferred.
Data analysis
We computed amino acid variability using Shannon entropy with the on-line tool at Los Álamos HIV sequence database (http://www.hiv.lanl.gov/content/sequence/ENTROPY/entropy_one.html). Mean entropy scores were calculated for each overlapping peptide (OLP) spanning the Nef protein (OLPs 67-93) and ranged from 0.15 in a ‘conserved’ region (OLP-82) to 0.78 in a ‘non-conserved’ region (OLP-68).
Statistical analysis was performed using GraphPad Prism v.5.0a. We focused our attention particularly on the central, most conserved region of Nef (previously defined as residues 81-160 [8]), extended here to correspond to the most conserved overlapping peptides (OLPs) adopted in these and in previously published studies [51], namely OLPs 76-85.
IFN-gamma ELISpot studies to identify regions of the Nef protein associated with beneficial immune responses
In vitro HIV-specific CD8+ T-cell responses were determined in a cohort of 1010 subtype-C infected individuals using IFN-gamma ELISpot assays using a set of 410 overlapping 18-mer OLPs spanning the whole HIV-1 subtype C proteome (2001 consensus sequence). Overlapping peptides were arranged in a matrix system with 11-12 peptides in each pool. Responses to matrix pools were confirmed by subsequent testing with the individual 18-mer peptides within each pool, and the identity of the individual 18-mers recognized were thus confirmed, as previously described [46].
OLP responses that independently correlate with log10 VL were identified using stepwise linear regression (inclusion criterion: p<0.05; exclusion criterion: p>0.05). We included in analysis all OLPs for which responses were observed in ≥3 individuals, and HLA class I alleles expressed in ≥3 individuals. P-values were computed using likelihood ratio tests, and q-values were computed over all features, conditioned on the final model. For these studies, the analyses resulting in q values <0.2 (p<0.024) were defined as significant.
HLA Class I tetramer staining
Tetramers were generated as previously described [52]. We stained antigen-specific cells using peptide-MHC-I tetramer conjugated to phycoerythrin (PE) or allophycocyanin (APC). Peripheral blood mononuclear cells (PBMCs) were thawed in R10 medium (RPMI medium, 10% fetal calf serum [FCS], 1% penicillin/streptomycin, 1% glutamine); rested for 1 h at 37°C in 5% CO2; stained with HLA class I tetramer for 30 min at room temperature; washed; surface stained with CD8 Pacific Blue (BD Pharmingen), CD3 Pacific Orange (Invitrogen), and a live/dead violet cell stain kit (Invitrogen) for 30 min; washed in phosphate-buffered saline (PBS) and fixed in 2% paraformaldehyde. Data were acquired on an LSR II (BD) flow cytometer within 12 h of staining and analyzed using FlowJo version 8.8.6. The cells were hierarchically gated on singlets, lymphocytes, live cells, and distinct tetramer-specific CD8-positive cells.
Results
Over 50% of HLA-selected polymorphisms in Nef are predicted to reduce viral fitness
Sequence analysis identified 44 Nef sequence polymorphisms associated with Class I HLA expression, operating at 34 different amino acid residues (all associations corrected for multiple comparisons, q<0.05 [49]; Table 1). Reversion was predicted following transmission to an HLA-mismatched recipient in 24 of the 44 HLA-associated polymorphisms (54.5%) (Table 1), implying that these polymorphisms would occur at a cost to viral replicative or infective capacity [19,49].
Table 1. Amino acid residues in Nef at which amino acid polymorphism is associated with HLA Class I expression.
| HLA | HXB2 position | P value | q value | Reversion | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A*68:01 | 15 | W | S | K | S | S | I | V | G | W | P | A | V | R | E | R | I | R | R | T | E | P | 5.00E-07 | 0.00 | |
| A*24:02 | 135 | Q | N | Y | T | P | G | P | G | V | R | Y | P | L | T | F | G | W | C | F | K | L | 4.40E-05 | 0.02 | R |
| A*23 | 143 | V | R | Y | P | L | T | F | G | W | C | F | K | L | V | P | V | D | P | R | E | V | 8.52E-17 | 0.00 | R |
| A*66:01 | 156 | V | P | V | D | P | R | E | V | E | E | A | N | K | G | E | N | N | C | L | L | H | 2.87E-05 | 0.01 | R |
| A*02 | 176 | H | P | M | S | Q | H | G | M | E | D | E | D | R | E | V | L | K | W | K | F | D | 1.07E-05 | 0.01 | |
| A*66 | 184 | E | D | E | D | R | E | V | L | K | W | K | F | D | S | S | L | A | R | R | H | L | 6.70E-05 | 0.02 | R |
| A*74:01 | 192 | K | W | K | F | D | S | S | L | A | R | R | H | L | A | R | E | L | H | P | E | Y | 2.89E-06 | 0.00 | R |
| B*73:01 | 19 | S | I | V | G | W | P | A | V | R | E | R | I | R | R | T | E | P | A | A | E | G | 1.53E-04 | 0.05 | |
| B*42 | 20 | I | V | G | W | P | A | V | R | E | R | I | R | R | T | E | P | A | A | E | G | V | 7.22E-05 | 0.03 | |
| B*44(03) | 64 | C | A | W | L | E | A | Q | E | E | E | E | V | G | F | P | V | R | P | Q | V | P | 2.52E-08 | 0.00 | R |
| B*4501 | 64 | C | A | W | L | E | A | Q | E | E | E | E | V | G | F | P | V | R | P | Q | V | P | 3.90E-08 | 0.00 | |
| B*4501 | 66 | A | W | L | E | A | Q | E | E | E | E | V | G | f | P | V | R | P | Q | V | P | L | 1.50E-26 | 0.00 | R |
| B*44(03) | 71 | E | E | E | E | E | V | G | F | P | V | R | P | Q | V | P | L | R | P | M | T | Y | 5.23E-06 | 0.00 | R |
| B*07 [Cw*07:02] | 71 | E | E | E | E | E | V | G | F | P | V | R | P | Q | V | P | L | R | P | M | T | Y | 2.16E-13 | 0.00 | R |
| B*57 | 76 | V | G | F | P | V | R | P | Q | V | P | L | R | P | M | T | Y | K | A | A | F | D | 5.55E-05 | 0.02 | R |
| B*81(01) | 76 | V | G | F | P | V | R | P | Q | V | P | L | R | P | M | T | Y | K | A | A | F | D | 2.71E-23 | 0.00 | |
| B*35(01) | 81 | R | P | Q | V | P | L | R | P | M | T | Y | K | A | A | F | D | L | S | F | F | L | 8.22E-11 | 0.00 | |
| B*57 | 83 | Q | V | P | L | R | P | M | T | Y | K | A | A | F | D | L | S | F | F | L | K | E | 2.41E-07 | 0.00 | |
| B*5801 [A*03] | 83 | Q | V | P | L | R | P | M | T | Y | K | A | A | F | D | L | S | F | F | L | K | E | 1.16E-12 | 0.00 | R |
| B*44(03) | 93 | A | A | F | D | L | S | F | F | L | K | E | K | G | G | L | E | G | L | I | Y | S | 2.48E-15 | 0.00 | R |
| B*44:03 | 102 | K | E | K | G | G | L | E | G | L | I | Y | S | K | K | R | Q | E | I | L | D | L | 1.57E-18 | 0.00 | R |
| B*18(01) | 108 | E | G | L | I | Y | S | K | K | R | Q | E | I | L | D | L | W | V | Y | H | T | Q | 1.36E-11 | 0.00 | |
| B*44:03 | 108 | E | G | L | I | Y | S | K | K | R | Q | E | I | L | D | L | W | V | Y | H | T | Q | 1.08E-09 | 0.00 | |
| B*44:03 | 115 | K | R | Q | E | I | L | D | L | W | V | Y | H | T | Q | G | F | F | P | D | W | Q | 1.25E-04 | 0.04 | R |
| B*57:03 | 116 | R | Q | E | I | L | D | L | W | V | Y | H | T | Q | G | F | F | P | D | W | Q | N | 8.93E-06 | 0.01 | |
| B*35(01) | 133 | D | W | Q | N | Y | T | P | G | P | G | V | R | Y | P | L | T | F | G | W | C | F | 3.32E-08 | 0.00 | |
| B*53(01) | 133 | D | W | Q | N | Y | T | P | G | P | G | V | R | Y | P | L | T | F | G | W | C | F | 3.12E-05 | 0.01 | |
| B*42 | 133 | D | W | Q | N | Y | T | P | G | P | G | V | R | Y | P | L | T | F | G | W | C | F | 4.75E-05 | 0.02 | R |
| B*14:01 | 156 | V | P | V | D | P | R | E | V | E | E | A | N | K | G | E | N | N | C | L | L | H | 5.46E-05 | 0.02 | |
| B*13 | 158 | V | D | P | R | E | V | E | E | A | N | K | G | E | N | N | C | L | L | H | P | M | 1.89E-05 | 0.01 | R |
| B*42 | 168 | K | G | E | N | N | C | L | L | H | P | M | S | Q | H | G | M | E | D | E | D | R | 1.75E-05 | 0.01 | |
| B*42 | 173 | C | L | L | H | P | M | S | Q | H | G | M | E | D | E | D | R | E | V | L | K | W | 4.11E-05 | 0.02 | R |
| B*15:01 | 176 | H | P | M | S | Q | H | G | M | E | D | E | D | R | E | V | L | K | W | K | F | D | 1.18E-05 | 0.01 | |
| B*53(01) | 176 | H | P | M | S | Q | H | G | M | E | D | E | D | R | E | V | L | K | W | K | F | D | 2.14E-05 | 0.01 | R |
| B*08:01 | 179 | S | Q | H | G | M | E | D | E | D | R | E | V | L | K | W | K | F | D | S | S | L | 1.24E-04 | 0.04 | |
| B*15:10 | 191 | L | K | W | K | F | D | S | S | L | A | R | R | H | L | A | R | E | L | H | P | E | 5.99E-06 | 0.00 | R |
| B*35(01) | 194 | K | F | D | S | S | L | A | R | R | H | L | A | R | E | L | H | P | E | Y | Y | K | 1.02E-05 | 0.01 | R |
| B*42:01 | 202 | R | H | L | A | R | E | L | H | P | E | Y | Y | K | D | C | 4.76E-05 | 0.02 | |||||||
| Cw*02:10 | 48 | K | H | G | A | L | T | S | S | N | T | A | H | N | N | A | D | C | A | W | L | E | 1.51E-04 | 0.05 | |
| Cw*08 | 85 | P | L | R | P | M | T | Y | K | A | A | F | D | L | S | F | F | L | K | E | K | G | 3.19E-05 | 0.01 | R |
| Cw*07:01 [B*080 | 105 | G | G | L | E | G | L | I | Y | S | K | K | R | Q | E | I | L | D | L | W | V | Y | 2.93E-14 | 0.00 | R |
| Cw*16:01 | 174 | L | L | H | P | M | S | Q | H | G | M | E | D | E | D | R | E | V | L | K | W | K | 1.13E-04 | 0.04 | |
| Cw*16 | 198 | S | L | A | R | R | H | L | A | R | E | L | H | P | E | Y | Y | K | D | C | 1.26E-05 | 0.01 | R | ||
| Cw*16 | 199 | L | A | R | R | H | L | A | R | E | L | H | P | E | Y | Y | K | D | C | 1.44E-05 | 0.01 | R |
Bold interface indicates site of polymorphism
Lineage-corrected analysis of sequences from 739 C-clade infected Southern African subjects. Amino acid residue at which polymorphism is identified is shown in bold, with ten amino acids flanking on either side (consensus sequence for this cohort). Known HLA-class I epitopes corresponding to the allele in question are underlined. Sites at which reversion is predicted are marked ‘R’ [19]. Round brackets indicate likely high resolution HLA type based on high population prevalence of this allele. Square brackets indicate alleles in linkage disequilibrium with selecting allele, with association also detected at this site.
Of the 44 HLA-associated polymorphisms identified, 25 (57%) were within or in the flanking regions (within 5 amino acids up- or down-stream) of known epitopes restricted by the HLA allele in question (Table 1). The majority of polymorphisms (n=31, 70%) were associated with HLA-B alleles, while 7 (16%) were associated with HLA-A and 6 (14%) with HLA-C (Table 1).
HLA-mediated selection pressure contributes to sequence diversity in conserved central Nef
Nef can be divided into three regions of differing sequence variability (Figure 1). The central region (residues 66-148, median residue entropy 0.135) is the most conserved, significantly more so than both the C-terminal region (residues 149-206, median residue entropy 0.491, p < 0.0001, Mann–Whitney U test), and the N-terminal region (residues 1-65, median residue entropy 0.829, p < 0.0001, Mann–Whitney U test). The C-terminal region is significantly more conserved than the N-terminal region (p=0.0056, Mann–Whitney U test).
Figure 1. Variation in C-clade Nef sequences, and relationship between amino acid variability and presence of HLA-class I selection pressure.

[A] Nef consensus sequence (derived from 739 C-clade sequences from Southern African patients) plotted against Shannon entropy score. Residues at which there is an association with HLA-Class I expression are shown in grey. As previously observed, the sequence is most highly conserved in the central portion of the protein (residues 66-148) [8]..
[B] Entropy scores of residues associated with HLA-class I expression vs. those with no HLA association, showing significantly higher variability at sites at which HLA selection operates, particularly in the central conserved region. P-value by Mann Whitney U test.
There is a strong relationship between the presence of an HLA-selected polymorphism and amino acid variability, particularly in the central conserved region of Nef (p=0.0001, Mann–Whitney U test; Figure 1B). Although this type of analysis cannot determine the causation of amino acid diversity, this association may indicate that HLA is a significant factor in driving polymorphisms, as previously suggested with Gag sequences [10] and supported in studies of longitudinally sampled Nef sequences [53]. This illustrates the potential evolutionary influence of HLA-selection in Nef [29,54], 47], as a factor contributing to sequence diversity, together with other factors such as neutral ‘toggling’ between amino acids, founder effect and genetic drift [55].
HLA-B selected mutations in Nef are associated with reduction in viral set-point
We next investigated the relationship between the presence of polymorphisms selected by HLA-A, HLA-B and HLA-C alleles and viral load (VL) in chronic infection. HLA-B alleles driving the selection of Nef mutants are linked with lower VL than those that do not select Nef polymorphisms (median VL 21,175 RNA copies/ml versus 49,350 for HLA-B alleles not associated with any Nef selection; p=0.003, Mann Whitney U test; Figure 2). However, increasing the number of Nef mutations beyond 1 was not statistically associated with HLA-B alleles predictive of lower VL. There was no significant relationship between the selection of mutations and median VL for HLA-A and HLA-C alleles (p=0.2 and p=0.5, respectively, Figure 2).
Figure 2. Relationship between number of HLA associations with Nef sequence polymorphisms and median viral load for subjects expressing that allele, for HLA-A, -B and -C alleles.
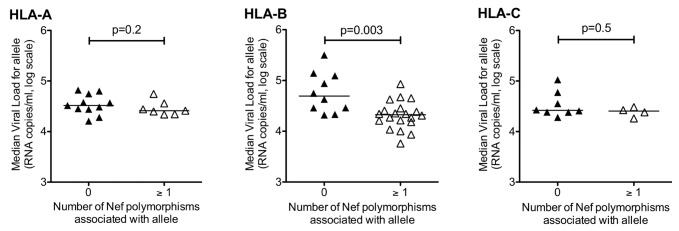
Data from lineage-corrected analysis of 739 C-clade Nef sequences. P-values by Mann–Whitney U test.
Given the relationship between HLA-B selection in Nef and reduced VL, we further scrutinized the pattern of HLA-B-associated polymorphisms. The commonest HLA-B alleles driving selection were HLA-B*44 (n=6; 14% of all associations), HLA-B*42 (n=5; 11%), HLA-B*57 (n=3; 7%) and HLA-B*35 (n=3; 7%) (Table 1). All of these alleles have been associated with favourable control of viraemia in previous studies of C-clade infected cohorts [31,56–58]. This includes HLA-B*35: 01, which is associated with rapid progression in B-clade infection but not in C-clade infection [31]. The presence of a strong CD8+ T cell response directed at Nef by all of these alleles is again consistent with the hypothesis that specific Nef responses may be a component of successful disease control.
Definition of novel HLA-B*44:03 restricted Nef epitopes
HLA-B*44 is responsible for the greatest number of HLA-selected polymorphisms in Nef (Table 1), but there are no epitopes restricted by HLA-B*44 that have been sufficiently well defined to appear in the Los Álamos epitope ‘A-list’ (http://www.hiv.lanl.gov/content/immunology/tables/optimal_ctl_summary.html). In Southern Africa, the predominant HLA-B*44 subtype is HLA-B*44: 03. For these reasons, we focused on further identification of HLA-B*44: 03 epitopes. The approach we used for guidance was, first, to identify the Nef residues at which HLA-B*44: 03 drives escape, then to predict the optimal epitope based on the peptide-binding motif for HLA-B*44: 03, which features glutamate (E) at position 2, and phenylalanine (F) or tyrosine (Y) at the C-terminus [59,60]. We thus identified Nef 93-102 (KEKGGLEGLIY; locations of HLA-escape associations underlined) and Nef 107-115 (QEILDLWVY) as putative HLA-B*44: 03-restricted epitopes. We subsequently confirmed these epitopes using HLA-B*44: 03 tetramers bound to KY11 or QY9 (Figure 3). In both epitopes, the putative escape mutations are located exclusively at the anchor residues, consistent with recent studies indicating a tendency for selection of escape at anchor residues [61], especially among protective alleles. The location of these epitopes, which overlap with others presented by highly protected MHC-I alleles (Figure 4), as well as with many others within the central highly immunogenic region of Nef (see epitope map at http://www.hiv.lanl.gov/content/immunology/maps/ctl/Nef.html) would make precise identification, quantification and analysis of such responses highly problematic without access to such tetrameric reagents.
Figure 3. Confirmation of Nef-KY11 (KEKGGLEGLIY) and Nef-QY9 (QEILDLWVY) as novel HLA-B*44: 03 restricted.

CD8 T cell epitopes using HLA-Class I tetramers. HLA-B*44: 03 tetramers were loaded with [A] Nef-KY11 and [B] Nef-QY9 optimal peptide and stained against donor PBMC’s from a C-clade infected B*44: 03 positive individual, subject R129 HLA-A*23: 01/30:04, B*08: 01/44:03, C*03:04/04:01. The number shown above each gate is the percentage of total lymphocytes that are tetramer-specific (tetramer positive cells expressed as a percentage of CD8+ cells were 1.48% for KY11 and 0.65% for QY9).
Figure 4. Map of central region of Nef showing sites of key epitopes and residues at which CD8+ selection pressure operates.

Central conserved region of HIV-1 Nef is shown previously defined as HXB2 residues 81-160 [8]. Corresponding B-clade and C-clade consensus sequences are shown along with SIVmac239 consensus. Positions of epitopes restricted by alleles HLA-B*27 [37], HLA-B*57 [28,64,65] and HLA-B*44 are highlighted in green, orange and blue respectively. Regions homologous to Mamu-B*08 [36] and Mamu-B*17 epitopes [66] are also marked (yellow and purple respectively). SIV 115-129 also highlighted as a region recently associated with SIV control in macaques [41]. Responses to overlapping peptides 79 and 85 are associated with viraemic control, q<0.2 (black boxes). Sites of mutations selected by HLA-B*57, HLA-B*27, HLA-B*44, Mamu-B*08 and Mamu-B*17 are marked with arrows [29,36,67]. This highlights the substantial overlap between HIV and SIV epitopes restricted by alleles that are associated with favourable immune control.
CD8+ T cell responses to certain Nef regions associated with lower viral load set-point
Our analyses of this extended dataset confirmed previous findings [51] of a statistically significant relationship between reduced VL and IFN-gamma ELISpot T cell responses to two OLPs in Nef, with q<0.2: OLP-79 SFFLKEKGGLEGLIYSKK (p=0.01, q=0.13), and OLP-85, RYPLTFGWCFKLVPV (p=0.002, q=0.04).
Examining these regions in more detail, the Nef regions spanned by OLP-79 and OLP-85 contain epitopes restricted by HLA-B*27, HLA-B*44 and HLA-B*57 [28] (http://www.hiv.lanl.gov/content/immunology/ctl_search and [14]) as well as by the protective macaque MHC-I alleles Mamu-B*08 and Mamu-B*17 and the protective Burmese rhesus monkey haplotype 90-010-Id (D) (Figure 4). Together, these data suggest that responses to these regions of Nef may be especially beneficial and contribute to MHC-I-associated protection against HIV/SIV disease.
Discussion
We have here shown, using several approaches, that CD8+ T cell responses to HIV Nef can be linked with improved viraemic suppression. First, HLA-selected escape mutations in Nef have a high rate of reversion to wild-type following transmission, suggesting a fitness cost imposed by the mutation in the presence of the selecting allele. Second, targeting of two specific OLPs in Nef is associated with reduction in viraemia in chronic infection, pointing to successful T cell responses in these regions. Third, there is substantial overlap between Nef regions targeted by beneficial HLA Class I epitopes and by simian Mamu-restricted responses that are associated with disease control.
The finding that HLA alleles predictive of lower viraemia are statistically associated with selection of one or more HLA-selected polymorphisms in Nef points to a contribution of CD8+ T cell responses to Nef in mediating disease control. It is problematic however to infer the impact of individual mutations by comparison of viral loads in subjects with and without those mutants. If the mutation is associated with a significantly increased viral load this may suggest that the CTL response contributes to immune control and if there is no significant increase that it does not. However, a lack of significant increase in viral load associated with selection of the escape mutant may reflect reduced replicative capacity resulting from that mutation and a fine balance between selection pressure driven by the CTL response and opposing purifying selection pressure exerted by the virus. The fact that 5 out of the 6 B*44: 03-associated Nef mutations were predicted to be revertants, that is, induced a cost to viral replicative capacity, underlines the intractability of such analyses. For these reasons, we cannot draw specific conclusions about the effects of selection of one or more Nef polymorphisms; however, these results do suggest that the ability to target Nef epitopes with sufficient strength that escape associations can be detected population-wide is a correlate of protection.
HLA-B*44 is most frequently associated with sequence polymorphism within Nef of any HLA allele and is also consistently associated with some tendency to protection against disease progression or control of viraemia, amounting to approximately a 0.25 log10 reduction in viral set point on average, compared to a ~0.75 log10 reduction for HLA-B*57: 03, the most protective HLA allele prevalent in sub-Saharan African populations [31,46,56]. We have here defined two novel HLA-B*44: 03 restricted epitopes within Nef, KY11 (KEKGGLEGLIY) and QY9 (QEILDLWVY) (Figure 3). KY11 is entirely contained within OLP-79 which has been associated with improved disease control overall. Due to the small number of HLA-B*44: 03 responders to this OLP in our cohort (2 responders of a total 91 subjects with HLA-B*44: 03), we have been unable to determine whether recognition of the KY11 epitope itself contributes to control of viraemia. These tetramer-based confirmations of the optimal epitopes in this central region of Nef highlight the necessity to use these reagents in order to characterize CTL responses that are potentially critically important to immune control.
The overall degree of sequence diversity in Nef might suggest that amino acid substitutions would be well tolerated without a significant detriment to viral infectivity or replicative capacity. However, a reversion rate of >50% (Table 1) suggests that, even in variable regions of the protein, a fitness cost to the virus is commonly imposed by the selection of HLA escape mutations. The polymorphisms we identified do not only occur in the epitope-rich central portion of Nef, but also in the terminal regions of the protein, particularly at the C-terminal region (Figure 1). This finding suggests that the C-terminal region of Nef as well as the central, highly conserved region, also contains CD8+ T cell epitopes. Potentially these may also be important, but the more variable regions of the viral proteome present a considerable challenge to epitope definition and characterization.
It is striking that SIV epitopes in Nef that are associated with immune control correspond so specifically to sites of confirmed or putative HIV HLA-B*57 and B*27 epitopes, and with HIV-OLPs that are associated with disease control (Figure 4). The substantial overlap between favourable responses in SIV and HIV highlights the utility of SIV studies in detecting regions of the HIV proteome that warrant further attention. However it is also notable that sequence conservation is not the only factor contributing to the potential efficacy of Nef-specific responses: disease protection was only associated with responses to OLPs-79 and -85, while responses to OLPs 80-84 which span this region - and are equally conserved and just as immunogenic as OLP-79 and OLP-85 - are not associated with lowered VL (Figure 4). This observation is also highlighted by other recent studies that report substantial variation in fitness costs associated with HLA-selected mutations irrespective of the degree of sequence conservation [62,63].
Overall, these results confirm the importance of Nef as a CD8+ T cell target with a potentially significant impact on HIV viraemic control, highlight the potential for HLA selection to drive evolutionary change, and corroborate the predominance of the HLA-B locus as a selection force. The high rate of reversion suggests that over half of the HLA selected polymorphisms are associated with a significant cost to viral fitness, underlining the usefulness of this protein as a vaccine target. Further studies to characterize the functional role of these regions of Nef and to define more CD8+ T cell epitopes will further inform the use of Nef as a vaccine immunogen.
Funding Statement
PCM is funded by Oxford University as an National Institute for Health Research Clinical Lecturer. PG is supported by grants from the Wellcome Trust (Senior Clinical Fellowship) and by the National Institutes of Health, grant R01 AI46995. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. O’Neill E, Kuo LS, Krisko JF, Tomchick DR, Garcia JV et al. (2006) Dynamic evolution of the human immunodeficiency virus type 1 pathogenic factor, Nef. J Virol 80: 1311-1320. doi:10.1128/JVI.80.3.1311-1320.2006. PubMed: 16415008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Foster JL, Garcia JV (2008) HIV-1 Nef: at the crossroads. Retrovirology 5: 84. doi:10.1186/1742-4690-5-84. PubMed: 18808677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Landi A, Iannucci V, Nuffel AV, Meuwissen P, Verhasselt B (2011) One protein to rule them all: modulation of cell surface receptors and molecules by HIV Nef. Curr HIV Res 9: 496-504. doi:10.2174/157016211798842116. PubMed: 22103833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Herbein G, Gras G, Khan KA, Abbas W (2010) Macrophage signaling in HIV-1 infection. Retrovirology 7: 34. doi:10.1186/1742-4690-7-S1-P34. PubMed: 20380698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Goulder PJ, Watkins DI (2008) Impact of MHC class I diversity on immune control of immunodeficiency virus replication. Nat Rev Immunol 8: 619-630. doi:10.1038/nri2357. PubMed: 18617886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Walker PR, Ketunuti M, Choge IA, Meyers T, Gray G et al. (2007) Polymorphisms in Nef associated with different clinical outcomes in HIV type 1 subtype C-infected children. AIDS Res Hum Retrovir 23: 204-215. doi:10.1089/aid.2006.0080. PubMed: 17331028. [DOI] [PubMed] [Google Scholar]
- 7. Almeida CA, Bronke C, Roberts SG, McKinnon E, Keane NM et al. (2011) 2011) Translation of HLA-HIV associations to the cellular level: HIV adapts to inflate CD8 T cell responses against Nef and HLA-adapted variant epitopes. J Immunol 187: 2502-2513. doi:10.4049/jimmunol.1100691. PubMed: 21821798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Mashishi T, Loubser S, Hide W, Hunt G, Morris L et al. (2001) Conserved domains of subtype C nef from South African HIV type 1-infected individuals include cytotoxic T lymphocyte epitope-rich regions. AIDS Res Hum Retrovir 17: 1681-1687. doi:10.1089/088922201753342103. PubMed: 11779357. [DOI] [PubMed] [Google Scholar]
- 9. Brumme ZL, Brumme CJ, Carlson J, Streeck H, John M et al. (2008) Marked epitope and allele-specific differences in rates of mutation in HIV-1 Gag, Pol and Nef CTL epitopes in acute/early HIV-1 infection. J Virol. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Kawashima Y, Pfafferott K, Frater J, Matthews P, Payne R et al. (2009) Adaptation of HIV-1 to human leukocyte antigen class I. Nature 458: 641-645. doi:10.1038/nature07746. PubMed: 19242411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Shugars DC, Smith MS, Glueck DH, Nantermet PV, Seillier-Moiseiwitsch F et al. (1993) Analysis of human immunodeficiency virus type 1 nef gene sequences present in vivo. J Virol 67: 4639-4650. PubMed: 8043040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Tolstrup M, Laursen AL, Gerstoft J, Pedersen FS, Ostergaard L et al. (2006) Cysteine 138 mutation in HIV-1 Nef from patients with delayed disease progression. Sex Health 3: 281-286. doi:10.1071/SH06002. PubMed: 17112441. [DOI] [PubMed] [Google Scholar]
- 13. Wang YE, Li B, Carlson JM, Streeck H, Gladden AD et al. (2009) Protective HLA class I alleles that restrict acute-phase CD8+ T-cell responses are associated with viral escape mutations located in highly conserved regions of human immunodeficiency virus type 1. J Virol 83: 1845-1855. doi:10.1128/JVI.01061-08. PubMed: 19036810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Gray CM, Mlotshwa M, Riou C, Mathebula T, de Assis Rosa D et al. (2009) Human immunodeficiency virus-specific gamma interferon enzyme-linked immunospot assay responses targeting specific regions of the proteome during primary subtype C infection are poor predictors of the course of viremia and set point. J Virol 83: 470-478. doi:10.1128/JVI.01678-08. PubMed: 18945774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Mlotshwa M, Riou C, Chopera D, de Assis Rosa D, Ntale R et al. (2010) Fluidity of HIV-1-specific T-cell responses during acute and early subtype C HIV-1 infection and associations with early disease progression. J Virol 84: 12018-12029. doi:10.1128/JVI.01472-10. PubMed: 20826686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Lichterfeld M, Yu XG, Cohen D, Addo MM, Malenfant J et al. (2004) HIV-1 Nef is preferentially recognized by CD8 T cells in primary HIV-1 infection despite a relatively high degree of genetic diversity. AIDS 18: 1383-1392. doi:10.1097/01.aids.0000131329.51633.a3. PubMed: 15199314. [DOI] [PubMed] [Google Scholar]
- 17. Radebe M, Nair K, Chonco F, Bishop K, Wright JK et al. (2011) Limited immunogenicity of HIV CD8+ T-cell epitopes in acute Clade C virus infection. J Infect Dis 204: 768-776. doi:10.1093/infdis/jir394. PubMed: 21844303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Kiepiela P, Ngumbela K, Thobakgale C, Ramduth D, Honeyborne I et al. (2007) CD8+ T-cell responses to different HIV proteins have discordant associations with viral load. Nat Med 13: 46-53. doi:10.1038/nm1520. PubMed: 17173051. [DOI] [PubMed] [Google Scholar]
- 19. Matthews PC, Prendergast A, Leslie A, Crawford H, Payne R et al. (2008) Central role of reverting mutations in HLA associations with human immunodeficiency virus set point. J Virol 82: 8548-8559. doi:10.1128/JVI.00580-08. PubMed: 18596105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Rolland M, Frahm N, Nickle DC, Jojic N, Deng W et al. (2011) Increased breadth and depth of cytotoxic T lymphocytes responses against HIV-1-B Nef by inclusion of epitope variant sequences. PLOS ONE 6: e17969. doi:10.1371/journal.pone.0017969. PubMed: 21464919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Altfeld M, Addo MM, Rosenberg ES, Hecht FM, Lee PK et al. (2003) Influence of HLA-B57 on clinical presentation and viral control during acute HIV-1 infection. AIDS 17: 2581-2591. doi:10.1097/00002030-200312050-00005. PubMed: 14685052. [DOI] [PubMed] [Google Scholar]
- 22. Leslie AJ, Pfafferott KJ, Chetty P, Draenert R, Addo MM et al. (2004) HIV evolution: CTL escape mutation and reversion after transmission. Nat Med 10: 282-289. doi:10.1038/nm992. PubMed: 14770175. [DOI] [PubMed] [Google Scholar]
- 23. Goulder PJ, Phillips RE, Colbert RA, McAdam S, Ogg G et al. (1997) Late escape from an immunodominant cytotoxic T-lymphocyte response associated with progression to AIDS. Nat Med 3: 212-217. doi:10.1038/nm0297-212. PubMed: 9018241. [DOI] [PubMed] [Google Scholar]
- 24. Brockman MA, Schneidewind A, Lahaie M, Schmidt A, Miura T et al. (2007) Escape and compensation from early HLA-B57-mediated cytotoxic T-lymphocyte pressure on human immunodeficiency virus type 1 Gag alter capsid interactions with cyclophilin A. J Virol 81: 12608-12618. doi:10.1128/JVI.01369-07. PubMed: 17728232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Payne RP, Kløverpris H, Sacha JB, Brumme Z, Brumme C et al. (2010) Efficacious early antiviral activity of HIV Gag- and Pol-specific HLA-B*2705-restricted CD8+ T-cells. J Virol 84: 10543-10557. doi:10.1128/JVI.00793-10. PubMed: 20686036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Briggs JA, Simon MN, Gross I, Kräusslich HG, Fuller SD et al. (2004) The stoichiometry of Gag protein in HIV-1. Nat Struct Mol Biol 11: 672-675. doi:10.1038/nsmb785. PubMed: 15208690. [DOI] [PubMed] [Google Scholar]
- 27. Sacha JB, Chung C, Rakasz EG, Spencer SP, Jonas AK et al. (2007) Gag-specific CD8+ T lymphocytes recognize infected cells before AIDS-virus integration and viral protein expression. J Immunol 178: 2746-2754. PubMed: 17312117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Bailey JR, Brennan TP, O’Connell KA, Siliciano RF, Blankson JN (2009) Evidence of CD8+ T-cell-mediated selective pressure on human immunodeficiency virus type 1 nef in HLA-B*57+ elite suppressors. J Virol 83: 88-97. doi:10.1128/JVI.01958-08. PubMed: 18945771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Carlson JM, Brumme CJ, Martin E, Listgarten J, Brockman MA et al. (2012) Correlates of protective cellular immunity revealed by analysis of population-level immune escape pathways in HIV-1. J Virol 86: 13202-13216. doi:10.1128/JVI.01998-12. PubMed: 23055555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Navis M, Schellens IM, van Swieten P, Borghans JA, Miedema F et al. (2008) A nonprogressive clinical course in HIV-infected individuals expressing human leukocyte antigen B57/5801 is associated with preserved CD8+ T lymphocyte responsiveness to the HW9 epitope in Nef. J Infect Dis 197: 871-879. doi:10.1086/528695. PubMed: 18279072. [DOI] [PubMed] [Google Scholar]
- 31. Matthews PC, Koyanagi M, Kløverpris HN, Harndahl M, Stryhn A et al. (2012) Differential clade-specific HLA-B*3501 association with HIV-1 disease outcome is linked to immunogenicity of a single Gag epitope. J Virol 86: 12643-12654. doi:10.1128/JVI.01381-12. PubMed: 22973023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Mwimanzi P, Markle TJ, Martin E, Ogata Y, Kuang XT et al. (2013) Attenuation of multiple Nef functions in HIV-1 elite controllers. Retrovirology 10: 1. doi:10.1186/1742-4690-10-1. PubMed: 23289738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Loffredo JT, Sidney J, Bean AT, Beal DR, Bardet W et al. (2009) Two MHC class I molecules associated with elite control of immunodeficiency virus replication, Mamu-B*08 and HLA-B*2705, bind peptides with sequence similarity. J Immunol 182: 7763-7775. doi:10.4049/jimmunol.0900111. PubMed: 19494300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Mudd PA, Ericsen AJ, Burwitz BJ, Wilson NA, O’Connor DH et al. (2012) Escape from CD8+ T Cell Responses in Mamu-B*00801+ Macaques Differentiates Progressors from Elite Controllers. J Immunol 188: 3364-3370. doi:10.4049/jimmunol.1102470. PubMed: 22387557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Loffredo JT, Bean AT, Beal DR, León EJ, May GE et al. (2008) Patterns of CD8+ immunodominance may influence the ability of Mamu-B*08-positive macaques to naturally control simian immunodeficiency virus SIVmac239 replication. J Virol 82: 1723-1738. doi:10.1128/JVI.02084-07. PubMed: 18057253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Loffredo JT, Friedrich TC, León EJ, Stephany JJ, Rodrigues DS et al. (2007) CD8+ T cells from SIV elite controller macaques recognize Mamu-B*08-bound epitopes and select for widespread viral variation. PLOS ONE 2: e1152. doi:10.1371/journal.pone.0001152. PubMed: 18000532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Goulder PJ, Edwards A, Phillips RE, McMichael AJ (1997) Identification of a novel HLA-B*2705-restricted cytotoxic T-lymphocyte epitope within a conserved region of HIV-1 Nef. AIDS 11: 536-538. PubMed: 9084804. [PubMed] [Google Scholar]
- 38. O’Connor DH, Mothe BR, Weinfurter JT, Fuenger S, Rehrauer WM et al. (2003) Major histocompatibility complex class I alleles associated with slow simian immunodeficiency virus disease progression bind epitopes recognized by dominant acute-phase cytotoxic-T-lymphocyte responses. J Virol 77: 9029-9040. doi:10.1128/JVI.77.16.9029-9040.2003. PubMed: 12885919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Mothé BR, Sidney J, Dzuris JL, Liebl ME, Fuenger S et al. (2002) Characterization of the peptide-binding specificity of Mamu-B*17 and identification of Mamu-B*17-restricted epitopes derived from simian immunodeficiency virus proteins. J Immunol 169: 210-219. PubMed: 12077247. [DOI] [PubMed] [Google Scholar]
- 40. Budde ML, Greene JM, Chin EN, Ericsen AJ, Scarlotta M et al. (2012) Specific CD8+ T cell responses correlate with control of simian immunodeficiency virus replication in Mauritian cynomolgus macaques. J Virol 86: 7596-7604. doi:10.1128/JVI.00716-12. PubMed: 22573864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Takahashi N, Nomura T, Takahara Y, Yamamoto H, Shiino T et al. (2013) A Novel Protective MHC-I Haplotype Not Associated with Dominant Gag-Specific CD8(+) T-Cell Responses in SIVmac239 Infection of Burmese Rhesus Macaques. PLOS ONE 8: e54300. doi:10.1371/journal.pone.0054300. PubMed: 23342126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Kirchhoff F, Easterbrook PJ, Douglas N, Troop M, Greenough TC et al. (1999) Sequence variations in human immunodeficiency virus type 1 Nef are associated with different stages of disease. J Virol 73: 5497-5508. PubMed: 10364298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Miura T, Brockman MA, Brumme CJ, Brumme ZL, Carlson JM et al. (2008) Genetic characterization of human immunodeficiency virus type 1 in elite controllers: lack of gross genetic defects or common amino acid changes. J Virol 82: 8422-8430. doi:10.1128/JVI.00535-08. PubMed: 18562530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Bredell H, Martin DP, Van Harmelen J, Varsani A, Sheppard HW et al. (2007) HIV type 1 subtype C gag and nef diversity in Southern Africa. AIDS Res Hum Retrovir 23: 477-481. doi:10.1089/aid.2006.0232. PubMed: 17411382. [DOI] [PubMed] [Google Scholar]
- 45. van Harmelen J, Williamson C, Kim B, Morris L, Carr J et al. (2001) Characterization of full-length HIV type 1 subtype C sequences from South Africa. AIDS Res Hum Retrovir 17: 1527-1531. doi:10.1089/08892220152644232. PubMed: 11709097. [DOI] [PubMed] [Google Scholar]
- 46. Kiepiela P, Leslie AJ, Honeyborne I, Ramduth D, Thobakgale C et al. (2004) Dominant influence of HLA-B in mediating the potential co-evolution of HIV and HLA. Nature 432: 769-775. doi:10.1038/nature03113. PubMed: 15592417. [DOI] [PubMed] [Google Scholar]
- 47. Matthews PC, Adland E, Listgarten J, Leslie A, Mkhwanazi N et al. (2011) HLA-A*7401-Mediated Control of HIV Viremia Is Independent of Its Linkage Disequilibrium with HLA-B*5703. J Immunol 186: 5675-5686. doi:10.4049/jimmunol.1003711. PubMed: 21498667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Leslie A, Kavanagh D, Honeyborne I, Pfafferott K, Edwards C et al. (2005) Transmission and accumulation of CTL escape variants drive negative associations between HIV polymorphisms and HLA. J Exp Med 201: 891-902. doi:10.1084/jem.20041455. PubMed: 15781581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Carlson JM, Brumme ZL, Rousseau CM, Brumme CJ, Matthews P et al. (2008) Phylogenetic dependency networks: inferring patterns of CTL escape and codon covariation in HIV-1 Gag. PLOS Comput Biol 4: e1000225 PubMed: 19023406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Storey JD, Tibshirani R (2003) Statistical significance for genomewide studies. Proc Natl Acad Sci U S A 100: 9440-9445. doi:10.1073/pnas.1530509100. PubMed: 12883005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Mothe B, Llano A, Ibarrondo J, Daniels M, Miranda C et al. (2011) Definition of the viral targets of protective HIV-1-specific T cell responses. J Transl Med 9: 208. doi:10.1186/1479-5876-9-208. PubMed: 22152067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Leisner C, Loeth N, Lamberth K, Justesen S, Sylvester-Hvid C et al. (2008) One-pot, mix-and-read peptide-MHC tetramers. PLOS ONE 3: e1678. doi:10.1371/journal.pone.0001678. PubMed: 18301755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Brumme ZL, Brumme CJ, Carlson J, Streeck H, John M et al. (2008) Marked epitope- and allele-specific differences in rates of mutation in human immunodeficiency type 1 (HIV-1) Gag, Pol, and Nef cytotoxic T-lymphocyte epitopes in acute/early HIV-1 infection. J Virol 82: 9216-9227. doi:10.1128/JVI.01041-08. PubMed: 18614631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Matthews PC, Leslie AJ, Katzourakis A, Crawford H, Payne R et al. (2009) HLA footprints on human immunodeficiency virus type 1 are associated with interclade polymorphisms and intraclade phylogenetic clustering. J Virol 83: 4605-4615. doi:10.1128/JVI.02017-08. PubMed: 19244334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Bhattacharya T, Daniels M, Heckerman D, Foley B, Frahm N et al. (2007) Founder effects in the assessment of HIV polymorphisms and HLA allele associations. Science 315: 1583-1586. doi:10.1126/science.1131528. PubMed: 17363674. [DOI] [PubMed] [Google Scholar]
- 56. Leslie A, Matthews PC, Listgarten J, Carlson JM, Kadie C et al. (2010) Additive contribution of HLA class I alleles in the immune control of HIV-1 infection. J Virol 84: 9879-9888. doi:10.1128/JVI.00320-10. PubMed: 20660184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Matthews PC, Listgarten J, Carlson JM, Payne R, Huang KH et al. (2012) Co-operative additive effects between HLA alleles in control of HIV-1. PLOS ONE 7: e47799. doi:10.1371/journal.pone.0047799. PubMed: 23094091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Kløverpris HN, Harndahl M, Leslie AJ, Carlson JM, Ismail N et al. (2012) HIV control through a single nucleotide on the HLA-B locus. J Virol 86: 11493-11500. doi:10.1128/JVI.01020-12. PubMed: 22896606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Fleischhauer K, Avila D, Vilbois F, Traversari C, Bordignon C et al. (1994) Characterization of natural peptide ligands for HLA-B*4402 and -B*4403: implications for peptide involvement in allorecognition of a single amino acid change in the HLA-B44 heavy chain. Tissue Antigens 44: 311-317. doi:10.1111/j.1399-0039.1994.tb02401.x. PubMed: 7878657. [DOI] [PubMed] [Google Scholar]
- 60. DiBrino M, Parker KC, Margulies DH, Shiloach J, Turner RV et al. (1995) Identification of the peptide binding motif for HLA-B44, one of the most common HLA-B alleles in the Caucasian population. Biochemistry 34: 10130-10138. doi:10.1021/bi00032a005. PubMed: 7543776. [DOI] [PubMed] [Google Scholar]
- 61. Bronke C, Almeida CA, McKinnon E, Roberts SG, Keane NM et al. (2012) HIV escape mutations occur preferentially at HLA-binding sites of CD8 T cell epitopes. AIDS. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Boutwell CL, Carlson JM, Lin TH, Seese A, Power KA et al. (2013) Frequent and variable cytotoxic-T-lymphocyte escape-associated fitness costs in the human immunodeficiency virus type 1 subtype B Gag proteins. J Virol 87: 3952-3965. doi:10.1128/JVI.03233-12. PubMed: 23365420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Rolland M, Manocheewa S, Swain JV, Lanxon-Cookson EC, Kim M et al. (2013) HIV-1 conserved elements vaccines: Relationship between sequence conservation and replicative capacity. J Virol. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Goulder PJ, Bunce M, Krausa P, McIntyre K, Crowley S et al. (1996) Novel, cross-restricted, conserved, and immunodominant cytotoxic T lymphocyte epitopes in slow progressors in HIV type 1 infection. AIDS Res Hum Retrovir 12: 1691-1698. doi:10.1089/aid.1996.12.1691. PubMed: 8959245. [DOI] [PubMed] [Google Scholar]
- 65. Draenert R, Brander C, Yu XG, Altfeld M, Verrill CL et al. (2004) Impact of intrapeptide epitope location on CD8 T cell recognition: implications for design of overlapping peptide panels. AIDS 18: 871-876. doi:10.1097/00002030-200404090-00004. PubMed: 15060434. [DOI] [PubMed] [Google Scholar]
- 66. Evans DT, O’Connor DH, Jing P, Dzuris JL, Sidney J et al. (1999) Virus-specific cytotoxic T-lymphocyte responses select for amino-acid variation in simian immunodeficiency virus Env and Nef. Nat Med 5: 1270-1276. doi:10.1038/15224. PubMed: 10545993. [DOI] [PubMed] [Google Scholar]
- 67. Minang JT, Trivett MT, Coren LV, Barsov EV, Piatak M Jr. et al. (2008) The Mamu B 17-restricted SIV Nef IW9 to TW9 mutation abrogates correct epitope processing and presentation without loss of replicative fitness. Virology 375: 307-314. doi:10.1016/j.virol.2008.02.005. PubMed: 18328525. [DOI] [PMC free article] [PubMed] [Google Scholar]