

Abstract
This study investigated the effects of a long bout of aerobic exercise on hunger and energy intake and circulating levels of leptin and acylated ghrelin. Ten healthy male subjects undertook two, 4 h trials in a randomized crossover design. In the exercise trial subjects ran for 105 min at 50% of maximal oxygen uptake and the last 15 min at 70% of maximal oxygen uptake followed by a 120 min rest period. In the control trial, subjects rested for 4 h. Subjects consumed a buffet test meal at 180 min during each trial. Hunger ratings, acylated ghrelin, leptin, glucose and insulin concentrations were measured at 0, 1, 2, 3 and 4 h. No differences were found at baseline values for hunger, acylated ghrelin, leptin, insulin and glucose for both trials (p > 0.05). The estimated energy expenditure of the exercise trial was 1550 ± 136 kcal. Exercise did not change subsequent absolute energy intake, but produced a significant decrease (p < 0.05) in relative energy intake. A two-way ANOVA revealed a significant (p < 0. 05) interaction effect for hunger and acylated ghrelin. In conclusion, this exercise regimen had a positive effect on reducing appetite which is related to reduced acylated ghrelin responses over time. This finding lends support for a role of exercise in weight management.
Key points.
Physical exercise is a strategy used to counteract obesity, since it lowers the energetic balance by increasing energy expenditure. However, because any energy expended in exercise elevates the intensity of hunger and drives food consumption, it is pertinent to ask how effective exercise could be in helping people to lose weight or to prevent weight gain.
The effects of exercise on hunger sensations and food intake are fairly controversial and depend on the intensity and duration of exercise.
120 min prolonged treadmill exercise with mix intensity, temporarily decreased hunger sensations, acylated ghrelin and relative energy intake.
Variations in exercise intensity should theoretically be a useful means of weight loss.
Key words: Energy intake, energy expenditure, exercise, hunger, acylated ghrelin, weight loss, appetite
Introduction
Obesity is a global epidemic in developed countries that is associated with several serious medical conditions including diabetes mellitus, coronary heart disease and cancer (Adams et al., 2006; Hensrud et al., 2006). Physical exercise is a strategy used to counteract obesity, since it contributes to a negative energy balance by increasing energy expenditure. Because any energy expended in exercise elevates the intensity of hunger and drives food consumption (Bilski et al., 2009; Hopkins et al., 2010; Martins et al., 2008), it is pertinent to ask how effective exercise could be in helping people to lose weight or to prevent weight gain.
Mechanisms responsible for regulation of appetite and food intake are still under investigation. Over the past decade, our knowledge of this homeostatic system that regulates food intake and body weight has increased dramatically with the discovery of circulating peptide ghrelin that sends signals to the brain regarding the body’s nutritional status (Lely et al., 2004; Vriese et al., 2008).
Ghrelin was first discovered as an endogenous ligand for growth hormone secreting receptor (GHSR) (Kojima et al., 1999). Further studies showed that it strongly stimulated food intake and body mass (Mackelvie et al., 2007; Wren et al., 2000). Plasma ghrelin concentration increases before meals and decreases with feeding, achieving concentrations sufficient to stimulate hunger and food intake (Cummings, 2002; Wren, 2001). Because of such effects, existing studies have focused on the role of ghrelin on energy homeostasis.
Ghrelin has two forms in the circulation as acylated and des-acylated ghrelin (Mackelvie et al., 2007); however, only acylated ghrelin has an n-octanoyl group which is required for ghrelin to bind to the GHSR type 1a and for entry into the hypothalamus and pituitary gland to subsequently increase food intake (Kojima et al., 1999). At first, des-acyl ghrelin was regarded as a nonfunctional ghrelin ligand. However, it was later shown to affect food intake (Inhoff et al., 2008; Muscaritoli et al., 2007; Stengel et al., 2008), with recent evidence that des-acyl ghrelin inhibits acyl ghrelin (Kumar et al., 2010). Moreover, a recent study revealed that des-acyl ghrelin was secreted in a highly regulated manner in response to food deprivation in mice (Kirchner et al., 2009) and it has been suggested that des-acyl ghrelin binds to an additional, as yet unidentified receptor that is different from GHSR 1a.
There are limiting studies which investigated the effects of exercise on hunger, food intake and energy regulating hormones and the majority of these studies have focused on total ghrelin (Burns et al., 2007; Christ et al., 2006; Dall et al., 2002; Jürimae et al., 2007; Kraemer et al., 2004; Schmidt et al., 2004; Vestergaard et al., 2007). Burns et al., 2007 reported that the suppression of appetite during severe exercise was not related to the suppression of total ghrelin. There is some evidence that exercise at higher intensities for longer durations can suppresses acylated ghrelin and appetite (Broom et al., 2007; 2009).
Currently, little is known about the influence of exercise on acylated ghrelin and acylated ghrelin’s relationship with appetite and food intake after exercise. The limited number of studies are in contrast with each other and new studies are certainly needed in this field (Broom et al., 2007; 2009; Kim et al., 2008; King et al., 2010a; 2010b; Mackelvie et al., 2007; Marzullo et al., 2008; Unick et al., 2010).
Previous studies actually demonstrated suppression of appetite during and briefly after intense (>60% of VO2max) bouts of activity (Blundel and King, 2000; Blundell et al., 2003; Broom et al., 2007) but not after low and moderate intensity exercise (Erdmann et al., 2007; Imbeault et al., 1997; Thompson et al., 1988). This evidence suggests that the acute effects of exercise on hunger and food intake largely depend on the intensity of exercise. Although the common belief among health professionals is that low to moderate intensity (40-59% VO2max) exercise is the most appropriate exercise prescription for weight loss, and correspondingly appropriate for the prevention and treatment of obesity, (Carey, 2009; Imbeault et al., 1997; Kissileff et al., 1990) the majority of acylated ghrelin studies have focused on high intensity (≥ 70% VO2max) exercise (Broom et al., 2007; 2009; King et al., 2010b; Marzullo et al., 2008). These studies showed that intense (≥ 70% VO2max) exercise caused a post exercise suppression of hunger and acylated ghrelin although low intensity exercise did not cause the same effect on acylated ghrelin (King et al., 2010a) and hunger (Imbeault et al., 1997; King et al., 2010a; Thompson et al., 1988). Since previous studies have shown that exercise at higher intensities affect appetite and acylated ghrelin but lower intensities do not, a combination of mostly low intensity combined with some high intensity exercise may have positive effects on hunger and acylated ghrelin.
Glucose, leptin and insulin have suppressive effects on ghrelin and are therefore important for ghrelin regulation (Flanagan et al., 2003; Murdolo et al., 2003; Shintani et al., 2001; Shiya et al., 2002). Insulin, like leptin, is thought to inhibit neuropeptide Y (NPY) / agouti-related protein (AgRP) neurons in the arcuate region of the hypothalamus and thus reduce food intake (Air et al., 2002).
Therefore the objective of this study was to investigate the subjective feelings of hunger sensations, energy intake, glucose, insulin, leptin and acylated ghrelin concentrations after energy deficit created by long duration acute low intensity exercise followed by a short moderately high intensity exercise which could be more easily sustained in weight loss and weight control programs than solely high intensity exercise.
Methods
Study design
Ratings of subjective feelings of hunger, energy intake (EI), acylated ghrelin, leptin, insulin and glucose levels in response to rest and exercise were investigated using a randomized crossover design. Subjects acted as their own controls and were assigned to the two experimental conditions (resting and exercise), 1 week apart, in a counter-balanced order.
Subjects
Ten elite, male soccer players participated voluntarily in the study and all subjects were well trained and had a history of regular exercise for at least 5 years with an exercise frequency of at least 3 times a week. Written informed consent was obtained from all subjects before participation. The participants completed a health screen and a physical activity questionnaire. Individuals were recruited only if they met the following criteria: were non-smokers, were not currently on a weight gain/weight loss diet, had not been on any diet during the previous six month, had maintained a stable weight in the previous six month, had no gastric or digestive problems, and had no known history of cardiovascular disease. The experimental protocol was approved by the ethical board of the Abant Izzet Baysal University School of Medicine Clinical Laboratory Research, Bolu, Turkey, and the Institutional Review Board of Lehman College, The City University of New York, USA and was performed in accordance with the principles of the Declaration of Helsinki. The characteristics of the subjects at baseline are shown in Table 1.
Table 1.
Descriptive characteristics of subjects at baseline. Data are means (±SD).
| Age (years) | 20.12 (.17) |
| Body weight(kg) | 70.4 (3.0) |
| Height (m) | 1.76 (.83) |
| BMI (kg·m-2) | 22.03 (.44) |
| Waist/hip ratio(cm) | .86 (.03) |
| Waist circumference (cm) | 79.18 (2.34) |
| VO2max (ml ·kg-1 ·min -1) | 62.74 (5.05) |
| Body fat (%) | 16.51 (3.17) |
| REE (kcal/24 h) | 2713 (252) |
Specific procedures: Dietary protocol
Participants were asked to maintain their normal dietary and physical activity programs throughout the duration of the study. Subjects refrained from exercise, alcohol and caffeine 24 h before testing. Because energy balance and macronutrient intake can influence hunger, satiety, and feeding behavior, it was important to provide the same diet during the 24 h before each testing day. Throughout the day before the first main trial, participants' weight and food intake were recorded. Participants then replicated this food intake during the day before the second main trial. Participants were also provided with a standardized evening meal and asked to eat it at around 08:00 pm on the day preceding each of the study days. The dietary intake was analyzed by using a diet analysis software (Diet Analysis Plus©, Version 4.0, ESHA Research, Salem, OR).
Preliminary test: Resting energy expenditure
Resting energy expenditure (REE) was measured for the purpose of estimating daily energy requirements. On trial days, participants arrived at the laboratory at 08:00 am. REE was measured between 08:15 and 09:00 am after an overnight fast. Upon arriving at the laboratory, the subject would lie in a supine position on a bed for 30 min to acclimatize to room temperature and undergo familiarization with the equipment and procedures. REE measurements were performed for 10 min after a 30 min rest period in a lit room at a comfortable temperature setting (20-24 °C). Oxygen consumption (VO2, mL·min-1) and carbon dioxide production (VCO2; mL·min-1) were measured every 10s by Cortex II Metalyser gas analyser II (Cortex Biophysik, Leipzig, Germany). REE was calculated using the Weir equation: REE (kcal·day-1) = [3.94 (VO2) + 1.11 (VCO2)] 1.44 (Weir, 1990).
Anthropometric measurements
Height was measured to the nearest 0.1 cm using a Holtain fixed wall stadiometer. Body mass was measured to the nearest 0.01 kg using a beam balance. BMI was calculated as weight in kilograms divided by the square of height in meters. Waist circumference was measured with a soft tape, midway between lowest rib and iliac crest. Hip circumference was measured over the widest part of the gluteal region, and waist-to-hip ratio was calculated accordingly. Skinfold thickness was measured at four sites (triceps, biceps, subscapular and suprailiac) on the right-hand side of the body using calipers (Skinfold Caliper Baseline MM, Fabrication Enterprise Incorporated, New York, US). Several trials were repeated until three tests yielded a difference of less than 0.5%. Body density was calculated using a four -site formula (Durnin and Womersley, 1974) and percent body fat was then estimated using the Siri equation (Siri, 1961). All body weight measurements were obtained with subjects wearing shorts and a tee-shirt, without shoes.
Maximal aerobic capacity
Maximal oxygen uptake test was performed one week before the main trials. The subject’s VO2max was assessed during a graded exercise treadmill (HP Cosmos Mercury Med 4.0) test using standard Bruce protocol (Bruce et al., 1973) in the morning hours. The test was terminated when subjects stated they could no longer continue with the maximum workload. At the terminal workload, all subjects had to meet at least two of the following criteria for a valid test: (1) a final respiratory exchange ratio (RER) > 1.0, (2) O2 consumption increased by < 2 ml·kg-1 with an increase in exercise intensity, (3) attainment of >85% of age-predicted maximal heart rate. Respiratory gases were collected and analyzed throughout the entire exercise session with the use of a computer controlled breath-by- breath analyzer Cortex II Metalyser (Cortex Biophysik, Leipzig, Germany). The highest achieved value for oxygen consumption was considered the subject’s VO2max. Heart rate (HR) was measured continuously throughout the test using a commercially available HR monitor (Polar S725X, Polar Electro, Finland).
Main trials
Participants were given at least one week to recover from the preliminary exercise tests before performing two main trials (exercise and control) in a random, crossover design with an interval of at least one week between trials. After fasting overnight, the participants came to the laboratory at 08:00 am and an intravenous catheter was inserted into an antecubital vein (08:15 am). At 08:50 am, 10 min prior to exercise, resting blood samples were collected from the catheter. In the exercise trial, subjects completed the treadmill exercise protocol at two speeds predicted to elicit a specific VO2: 50% VO2max for 105 min and 70% VO2max for 15 min. Adjustments were made to the treadmill speed if it was necessary. For the control trial, participants rested for the entire duration of the trial.
Blood samples were collected at baseline and at 60, 120, 180 and 240 min after baseline for the determination of leptin, acylated ghrelin, insulin and glucose. All blood samples were collected while subjects lay in a semisupine position with the exception of the 60 min sample during the exercise trial; this sample was collected while subjects straddled the treadmill. Heart rate was recorded every 5 s during the treadmill exercise. Oxygen consumption and carbon dioxide production were measured every 10 s during the 2 h exercise using a Cortex II Metalyser and the mean respiratory exchange ratio (RER) was calculated from the recorded measurements.
The energy expended during the exercise and resting sessions (t = 0-120 min) was calculated using the Weir equation (Weir, 1990). The analyzer was calibrated before the test with gases of known concentration according to manufacturer’s guidelines. After the resting/exercise intervention (t = 120-240 min), participants stayed in the clinical investigation unit, but were free to write/read quietly. One hour after the end of the exercise/resting intervention (t = 180 min), participants were placed in individual booths, and presented with a buffet test meal. A diagram detailing the experimental session is shown in Figure 1.
Figure 1.
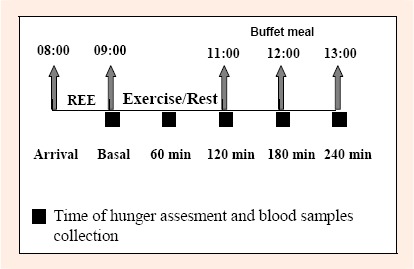
Experimental design
Subjective measurements of hunger
Ratings of subjective feelings of hunger were reported on 100 mm visual analogue scales (VAS) at baseline and at 60, 120, 180, 240 min after baseline, as described previously (Flint et al., 2000). For hunger assessment, on one end (0 mm) of the scale was the descriptor “not at all hungry ”and on the other extreme (100 mm) was the descriptor “totally hungry”.
Relative energy intake
Relative energy intake (REI) was calculated as follows: REI = EI - [ECE - (exercise time x REE)] and for control trial: REI = EI - (exercise time x REE) where EI is the energy intake during the buffet-type meal, ECE is the measured energy cost of exercise, and REE is the measured pre-exercise resting energy expenditure (Pomerleau et al., 2004).
Buffet test meal
After 180 min (3 hours after the start of each trial) at 12.00 pm, all subjects were given a buffet test meal that they had to eat until reaching satiety. The buffet test meal consisted of a standardised buffet available ad libitum, in excess of expected consumption (4200 kcal, 53% carbohydrates, 12% protein and 35% fat), with a variety of lunch-type foods (sandwiches, yogurt, fruit, salad, cake, cookies, milk, and chocolate bar). The buffet foods were weighed/counted before participants sat down to the meal and reweighed/recounted after each subject had finished eating, and energy intake calculated. The energy content of the items consumed was ascertained using manufacturer values and was analyzed by using diet analysis software (Diet Analysis Plus©, Version 4.0, ESHA Research, Salem, OR). Participants were encouraged to consume the meal within 15 min and kept to the same start and finish times on both trials. Water was available ad libitum during trials, and the volume and time of ingestion were recorded.
Blood biochemistry
Blood samples were drawn into chilled tubes containing Na2EDTA (1.25 mg/ml) and aprotinin (500 TIU/ml (Kallikrein Ýnhibitor Unit), Phoenix, Burlingame, USA). Immediately after collecting blood samples, ghrelin tubes were centrifuged at 1500g for 15 min at 4°C. The obtained plasma samples were mixed with 1 mol/L HCl (Hydrochloric acid) at a ratio of 1/10 and were stored at -70°C until assayed. Samples for glucose were immediately centrifuged, and the plasma was separated and kept on ice for processing that day. Plasma glucose levels were measured using a Abbott-Architect C8000 (Abbott Laboratories, Illinois, USA) automated analyser by an enzymatic assay involving hexokinase and glucose-6- phosphate dehydrogenase. Blood samples for leptin and insulin hormone were drawn into red cap tubes were centrifuged after completion of clot formation and the serum samples were stored at -70°C until the day of the hormone measurements. Serum leptin concentration was measured using a commercial sandwich enzyme-linked immunosorbent assay (ELISA; DRG Leptin EIA-2395; DRG International Inc., Marburg, Germany) with a limit of detection of 1.0 ng/ml. The intra- and inter-assay coefficients of variation were 6.91% and 8.66%, respectively. Serum insulin concentration was measured using a ultrasensitive enzyme immunoassay (ELISA; DRG Insulin EIA-2337; DRG International Inc., Marburg, Germany) with a limit of detection of < 0.07 mU/l. The intra- and inter-assay coefficients of variation were 5.1% and 1.8%, respectively. Plasma acylated ghrelin assay was performed using a commercially available ELISA (SCETI Acylated Ghrelin EIA-97751; SCETI Co., Ltd., Tokyo, Japan) with a detection limit of 2.5 fmol/ml. The intra- and inter-assay coefficients of variation were 5% and 10%, respectively. At each blood sampling point, duplicate 20 μL blood samples were collected into micropipettes for the measurement of hemoglobin concentration and triplicate 20 μL blood samples were collected into heparinised microhaematocrit tubes for the determination of hematocrit. Hemoglobin and hematocrit values were used to assess plasma volume changes (Dill and Costill, 1974).
Statistical analysis
Statistical analysis was carried out using SPSS version 15. 0 (SPSS, Inc., Chicago, IL, USA). All the variables were checked regarding their normal distribution using the Shapiro-Wilk test and data are presented as means ±SD. Area under the curve (AUC) values for acylated ghrelin, glucose, insulin, leptin and hunger were calculated using the trapezoidal rule. Paired sample t-tests were used to assess differences between baseline and area under the curve values for each of these variables on the control and exercise trials. A repeated-measures, two factor ANOVA was used to examine differences between the two trials over time for acylated ghrelin, glucose, insulin, leptin, hunger and plasma volume change. Between-trial differences at each time point were examined using one-way ANOVA and Bonferroni post hoc tests when significant interactions were found. Mauchley’s test was conducted to examine sphericity for the repeated measures analyses. If the assumption of sphericity was violated, the Greenhouse-Geisser adjustment was used to protect against type I error. Differences in absolute and relative energy intake at the buffet lunch between the two experimental conditions were assessed using paired sample t-tests. For correlation analysis, the Pearson coefficient was calculated. Plasma volume changes did not differ significantly between trials, and the unadjusted values are presented. Statistical significance was accepted at the 5% level.
Results
No significant differences were observed between trials in energy or macronutrient intake in the 24 h prior to each trial.
Resting energy expenditure
Resting energy expenditure averaged 2713 ± 52 kcal·d-1.
Responses to treadmill running
During the first 105 min of 120 min of exercise, average heart rate was 145 ± 9 beats·min-1, mean % VO2max was 52.34 ± 3.63 % ml·kg-1·min-1, mean respiratory exchange ratio was 0.76 ± 0.05 and gross energy expenditure was 1320 ± 116 kcal. During the last 15 min of the exercise, average heart rate was 181 ± 6 beats·min-1, mean % VO2max was 70.10 ± 2.93 % ml·kg-1·min-1, mean respiratory exchange ratio was 0.90 ± 0.06 and gross energy expenditure was 230 ± 20 kcal.
Hunger and energy intake
Baseline fasting hunger did not differ significantly (Paired sample t-test, p = 0.611) between trials (control 74.28 ± 6.65 mm, exercise 71.57 ± 0.80 mm). Two-factor ANOVA revealed a main effect of trial (p = 0.013), a main effect of time (p < 0.05), and a trial and time interaction (p = 0.005) for hunger, indicating that responses differed over time between trials. Post hoc analysis indicated significant differences between exercise and control trial at 120 and 180 min (p < 0.05; Figure 2). The area under the curve for hunger was significantly lower during exercise (15159 ± 1693) trial compared to control (18236 ± 1922; p < 0.05) trial.
Figure 2.
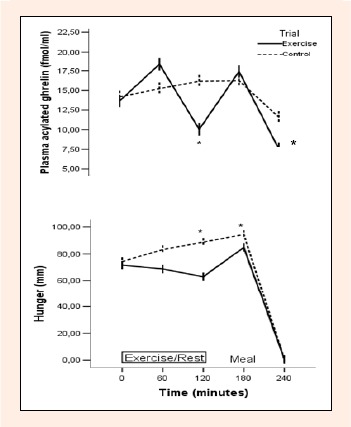
Acylated ghrelin concentrations and hunger over 240 min during the exercise and control trials.
There was an effect of trial (p = 0.032), an effect of time (p < 0.05), and a trial x time interaction (p = 0.005) for acylated ghrelin. Subject hunger ratings (mean ± SEM) response to the question “How hungry do you feel right now? ”on a VAS from 0 to 100 (0 = not hungry at all; 100 = extremely hungry). Two-factor ANOVA revealed a main effect of trial (P = 0.013), a main effect of time (p < 0.05), and a trial and time interac-tion (p = 0.005) for hunger *Significantly different from the control trial (p < 0.05) after Bonferroni adjustment.
Absolute and relative energy intake
Absolute energy intake after the exercise trial (2051 ± 690 kcal) was not significantly (Paired sample t-test, p = 0.777) different from the control trial (1957 ± 518 kcal). To further investigate the effects of exercise on energy intake, we calculated the REI. Statistical analyses showed a significantly lower REI (Paired sample t-test, p = 0.018) during the exercise trials than during the control trials (736 ± 701 and 1722 ± 520 kcal, respectively).
Plasma glucose, acylated ghrelin and serum leptin and insulin, changes over time: Acylated Ghrelin
Baseline fasting plasma acylated ghrelin concentrations did not differ significantly (Paired sample t-test, p = 0.586) between trials (control 14.18 ± 3.82 fmol/ml, exercise 13.23 ± 3.69 fmol/ml). A two-factor ANOVA revealed a main effect of trial (p = 0.032), a main effect of time (p < 0.05) and a trial and time interaction (p = 0.005) for acylated ghrelin concentrations. Post hoc analysis indicated that there were significant differences between exercise and control trial at 120 and 240 min (p < 0.05) (Figure 2). The plasma concentration of acylated ghrelin was suppressed after consumption of the meal in both trials (Figure 2). The area under the curve (AUC) for acylated ghrelin was significantly lower during exercise (3383 ± 309 fmol/mlx240 min) trial compared to control (3631 ± 213 fmol/mlx240 min; p = 0.046) trial.
Leptin
Baseline fasting leptin concentrations did not differ significantly (Paired sample t-test, p = 0.577) between trials (control 4.34 ± 0.69 ng·ml-1, exercise 4.57 ± 0.80 ng·ml-1). Two-factor ANOVA revealed a main effect of time (p < 0.05) for serum leptin, but there was no main effect of trial and no interaction effect (Figure 3), indicating that leptin changed significantly during the trials but were not influenced by long duration exercise with mixed intensity. The serum concentration of leptin was stimulated after consumption of the meal in both trials (Figure 3). The AUC for leptin was not different between exercise (893 ± 155 ng/mlx240 min) and control (928 ± 117 ng/mlx240 min; p = 0.623) trials.
Figure 3.
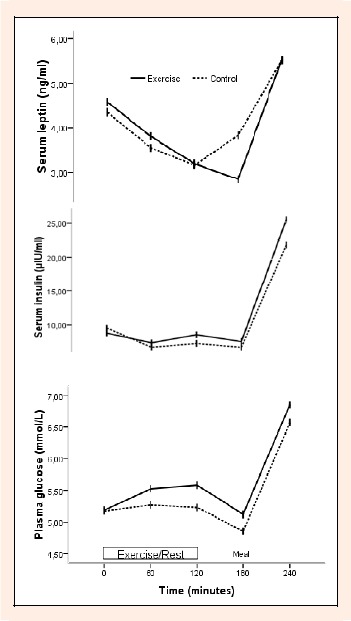
Serum leptin, insulin and plasma glucose concentrations over 240 min during the exercise and control trials. Two-factor ANOVA revealed a main effect of time (p < 0.05) for leptin, insulin and glucose. The interaction between time and trials was not significant for leptin, insulin and glucose.
Glucose and insulin
Baseline fasting plasma glucose concentrations did not differ significantly (Paired sample t-test, p = 0.902) between trials (control 5.17 ± 0.22 mmol·L-1, exercise 5.18 ± 0.20 mmol·L-1). Two-factor ANOVA revealed a main effect of time (p < 0.05), but there was no main effect of trial for plasma glucose (Figure 3). Baseline fasting serum insulin concentrations did not differ significantly (Paired sample t-test, p = 0.191) between trials (control 9.53 ± 0.54 μIU·ml-1, exercise 8.77 ± 1.29 μIU·ml-1). Two-factor ANOVA revealed a main effect of time (p < 0.05) for serum insulin, but there was no main effect of trial and no interaction effect (Figure 3). These results indicate that glucose and insulin changed significantly during the trials but were not influenced by long duration exercise with mixed intensity. The AUC for glucose (exercise: 24094 ± 943 mmol/Lx240 min, control: 22929 ± 944 mmol/Lx240 min; p = 0.066) and insulin (exercise: 2423 ± 484 μIU/mlx240 min, control: 2316 ± 302 μIU/mlx240 min; p = 0.529) were not different between exercise and control trials.
Correlations
Baseline acylated ghrelin concentration was significantly correlated with fasting glucose concentration (r = 0.560, p = 0.037) and hunger (r = 0.595, p = 0.032) and also a significant correlation (r = 0.698, p = 0.041) was observed between serum leptin and %BF. As we observe at the values during the exercise; immediately after 60 minutes of exercise, a significant negative correlation (r = 0.855, p = 0.014) was found between hunger and glucose concentration. Immediately after 120 minutes of exercise, there were significant correlations between acylated ghrelin and hunger (r = 0.753, p = 0.041), insulin and glucose (r = 0.795, p = 0.033), leptin and glucose (r = - 0.850, p = 0.015) during exercise trial. There were no significant correlations between other variables over other periods either for the exercise trial or the control trial.
Discussion
The objective of this study was to investigate the effect of an extreme energy deficit created by the combination of long duration moderate intensity exercise and short duration high intensity exercise on the hunger sensations, energy intake and acylated ghrelin concentration. The main findings in the present study are that hunger and acylated ghrelin concentrations were briefly suppressed after long duration mixed intensity treadmill running whereas energy intake was unaffected.
To the authors’ knowledge, the present study is the first study to examine the acylated ghrelin response to large negative energy balance stimulated by 120 min. of prolonged exercise. Previous studies showed that acylated ghrelin is suppressed immediately after a short bout (≤ 60 min.) of strenuous (≥ 70% VO2max; RER: ≥ 0.90) aerobic exercise (Broom et al., 2007; 2009; Marzullo et al., 2008). Although the phenomenon of ‘exercise-induced anorexia is traditionally associated with intense exercise (King et al., 1994), the current study demonstrated that short duration intense exercise performed after acute, long duration moderate intensity exercise also suppresses acylated ghrelin and hunger. This is a novel and important finding because low to moderate intensity of exercise is considered the preferred exercise intensity for inducing weight loss (Carey, 2009). In addition to the suppression in plasma acylated ghrelin concentrations immediately after exercise, acylated ghrelin concentration declined after buffet meal in both trials. This result is consistent with the report from studies examining the response of plasma acylated ghrelin to feeding (Blom et al., 2005; El Khoury et al., 2006; Lucidi et al., 2004).
The effects of exercise on hunger sensations and food intake are fairly controversial and depend on the intensity and duration of exercise (Blundell and King, 1999; Blundell et al., 2003; King et al., 1994; Maraki et al., 2005; Martins et al., 2008). In the present study, acute aerobic exercise of moderate intensity (50% VO2max) at 60 min of duration equivalent to an energy expenditure of 690 kcal had no effect on hunger sensation. On the other hand, exercise performed at moderate intensity (50% VO2max) for a period of 105 min (1319 kcal) and at a higher intensity (70% VO2max) for the last 15 min (230 kcal) for a total period of 120 min led to a decrease of hunger without having an effect on energy intake. This finding is concordant with many previous studies showing that low to moderate intensity exercise did not have an affect on hunger sensation, but high intensity exercise suppressed hunger (Erdmann et al., 2007; King et al., 1994; Martins et al., 2007; Thompson et al., 1988). This impact on hunger was explained by decreases in acylated ghrelin after exercise. Reduced hunger and acylated ghrelin concentration after high intensity exercise might be due to an increased sympathetic nervous system activity and redistribution of blood flow from the splanchnic circulation, towards the skeletal muscles during exercise (Dickhuth et al., 2004; Ikeda et al., 2010; Perini et al., 2003; Powers and Howley, 2002). This response may well be implicated in the phenomenon of intense exercise-induced anorexia.
However, despite the fact that hunger sensation increased in the recovery period immediately before consumption of a buffet meal, this increase was significantly lower in the exercise trial than in the control trial. Energy intake at the buffet meal was not different between trials. This result was unexpected given the extreme energy deficit induced during exercise. It was thought that energy intake would be higher at some point within the hours after exercise in an effort to the replenish energy stores that are depleted during exercise. The reason that the hunger sensation was significantly reduced after food intake in exercise trial compared to the control trial could be due to increased sympathetic nervous system activity but needs to be investigated in relation to ghrelin. It would be of interest to examine food intake in the immediate hours after exercise to assess whether food intake is elevated in response to the replenished energy stores.
Several studies showed that des-acylated ghrelin induces a state of negative energy balance. It reduces body weight by decreasing the food intake and delaying the gastric emptying in mice (Perboni and Inui, 2010). Mackelvie et al., 2007. Therefore the des-acyl ghrelin/acyl ghrelin ratio should also be evaluated during food intake. Because decreasing of acyl ghrelin and/or GHS-R1a, and overexpression of des-acyl ghrelin show benefits in the therapy of obesity and metabolic syndrome (Chen et al., 2009)
The reason that there was no increase in food intake after exercise in the present study could be due to a possible increase in des-acyl ghrelin. However des-acylated ghrelin measurement was not determined in this study. Mackelvie and et al (2007) reported a decrease in des-acylated ghrelin in normal male children and an increase in obese male children after aerobic exercise. However, des-acylated ghrelin was not measured directly in their study but it was measured by subtracting acylated ghrelin from total ghrelin and therefore it may not reflect the real values.
The finding that hunger is suppressed immediately after a large energy deficit induced by treadmill running and that this suppression of hunger tends not to affect energy intake is consistent with previous studies showing that post-exercise food intake may be independent of energy balance (King et al., 1994: 2010b; Westerterp et al., 1997). This apparent uncoupling between hunger and energy intake have been reported previously (Blundell et al., 1999; Flint et al., 2000; Martins et al., 2007; Mattes, 1990). Evidence shows that only 19% of the intervention studies report an increase in energy intake after exercise, 16% show a decrease and 65% show no change in appetite, (Blundell and King, 1999). Despite the extreme energy expenditure induced by the acute exercise of 120 min (1549 kcal), energy intake was not statistically different between trials. However, when accounting for the excess energy expended during exercise, relative energy intake during the exercise trial was lower than the control trial. This finding has positive implications for the use of physical activity in weight loss or weight maintenance.
In the present study we measured the concentrations of leptin, glucose and insulin during and after exercise and feeding because they may interact with acylated ghrelin. Consistent with previous findings, plasma glucose and serum insulin and leptin levels remained unchanged during and after exercise in the present study (Jürimae and Jürimae, 2005; Martins et al., 2007; Zoladz et al., 2005), and are therefore unlikely to contribute to the regulation of plasma acylated ghrelin fluctuations during and after acute, long duration exercise.
This study has several limitations. First, the sample of participants was composed of a relatively homogenous population of young, well-trained, healthy males. Therefore, the findings may not be generalized to clinical populations. Second, the present study lacked an acylated ghrelin measurement at the end of 105 min of exercise. If this measurement had been taken, the effects of the intensity of exercise on acylated ghrelin could have been better understood. Finally, the plasma acylated ghrelin was measured at only five time points over the 4 h observation. More frequent measurements would provide a clearer picture of the responses to exercise. There is certainly a need for more comprehensive studies to better understand the relationship between the intensity of exercise, hunger and acylated ghrelin concentrations.
Conclusion
This study demonstrates that large energy deficit induced by a long bout of treadmill running with mixed intensity, performed in a fasted state, significantly decreases hunger, acylated ghrelin and relative energy intake but has no effect on absolute energy intake. This supports the hypothesis that acute exercise is able to produce a short-term negative energy balance. Therefore, these variations in exercise intensity should theoretically be a useful means of weight loss. Further research is required to determine the influence of other models, intensities, and duration of exercise on plasma acylated ghrelin concentration in different subject groups. Such research could have important implications regarding the role of mixed intensity exercise in weight management.
Acknowledgments
We are grateful to our study participants for their willing cooperation and considerable time spent in participating in this study.
Biographies

Serife Vatansever-Ozen
Employment
Ass. Prof., Department of Training Science, School of Physical Education and Sports, University of Abant Izzet Baysal, Turkey.
Degree
PhD.
Research interests
Exercise and obesity, exercise physiology, altitude physiology.
E-mail: serifevatansever@yahoo.com

Gul Tiryaki-Sonmez
Employment
Assoc. Prof., Department of Health Sciences, Lehman College, The City University of New York, Bronx, New York, USA.
Degree
PhD.
Research interests
Exercise and obesity, childhood obesity, performance analysis, sports physiology.
E-mail: gultiryakisonmez@gmail.com

Guler Bugdayci
Employment
Assoc. Prof., Department of Biochemistry, Faculty of Medicine, University of Abant Izzet Baysal, Bolu, Turkey.
Degree
MD.
Research interests
Exercise biochemistry.
E-mail: gbugdayci@yahoo.com

Guclu Ozen
Employment
Research Assistant, Depart. of Sports Management, School of Physical Education and Sports, Univ. of Abant Izzet Baysal, Turkey.
Degree
PhD.
Research interests
Biostatistics, high altitude, physical fitness.
E-mail: guclu1234@yahoo.com
References
- Adams K.F., Schatzkin A., Haris T.B., Kipnis V., Mouw T., Ballard-Barbash R., Hollenbeck A., Leitzmann M.F. (2006) Overweight, obesity, and mortality in a large prospective cohort of persons 50 to 71 years old. New England Journal of Medicine 355, 763-778 [DOI] [PubMed] [Google Scholar]
- Air E.L., Benoit S.C., Blake-Smith K.A., Clegg D.J., Woods S.C. (2002) Acute third ventricular administration of insulin decreases food intake in two paradigms. Pharmacology Biochemistry and Behavior 772, 423-429 [DOI] [PubMed] [Google Scholar]
- Bilski J., Teległów A., Bilska J. Z., Dembiński A., Warzecha Z. (2009) Effects of exercise on appetite and food intake regulation. Medicina Sportiva 13, 82-94 [Google Scholar]
- Blom W.A.M., Stafleu A., de Graaf C., Kok F.J., Schaafsma G., Hendriks H.F.J. (2005) Ghrelin response to carbohydrate-enriched breakfast is related to insulin. American Journal of Clinical Nutrition 881, 367-375 [DOI] [PubMed] [Google Scholar]
- Blundell J.E., King N.A. (1999) Physical activity and regulation of food intake: current evidence. Medicine and Science in Sports and Exercise 31, 573-583 [DOI] [PubMed] [Google Scholar]
- Blundell J.E., Stubbs R.J., Hughes D.A., Whybrow S., King N.A. (2003) Cross talk between physical activity and appetite control: does physical activity stimulate appetite? Proceedings of the Nutrition Society 62, 651-661 [DOI] [PubMed] [Google Scholar]
- Blundell JE., King NA. (2000) Exercise, appetite control, and energy balance. Nutrition 16, 519-522 [DOI] [PubMed] [Google Scholar]
- Broom D.R., Batterham R.L., King J.A., Stensel D.J. (2009) Influence of resistance and aerobic exercise on hunger, circulating levels of acylated ghrelin, and peptide YY in healthy males. American Journal of Physiology - Regulatory, Integrative and Comparative Physiology ,2961, R29-R35 [DOI] [PubMed] [Google Scholar]
- Broom D.R., Stensel D.J., Bishop N.C., Burns S.F., Miyashita M. (2007) Exercise-induced suppression of acylated ghrelin in humans. Journal of Physiology 102, 2165-2171 [DOI] [PubMed] [Google Scholar]
- Bruce R.A., Kusumi F., Hosmer D. (1973) Maximal oxygen intake and nomographic assessment of functional aerobic impairment in cardiovascular disease. American Heart Journal 85, 546-562 [DOI] [PubMed] [Google Scholar]
- Burns S.F., Broom D.R., Mundy C., Miyashita M., Stensel D.J. (2007) A single session of treadmill running has no effect on plasma total ghrelin concentrations. Journal of Sports Science 25, 635-642 [DOI] [PubMed] [Google Scholar]
- Carey D.G. (2009) Quantifying differences in the "fat burning" zone and the aerobic zone: implications for training. Journal of Strength and Conditioning Research 23, 2090-2095 [DOI] [PubMed] [Google Scholar]
- Chen C.Y., Asakawa A., Fujimiya M., Lee S.D., Inui A. (2009) Ghrelin gene products and the regulation of food intake and gut motility. Pharmacological Reviews 661, 430-481 [DOI] [PubMed] [Google Scholar]
- Christ E.R., Zehnder M., Boesch C., Trepp R., Mullis P.E., Diem P., De’combaz J. (2006) The effect of increased lipid intake on hormonal responsesduring aerobic exercise in endurance-trained men. European Journal of Endocrinology 154, 397-403 [DOI] [PubMed] [Google Scholar]
- Cummings D.E., Weigle D.S., Frayo R.S., Breen P.A., Ma M.K., Dellinger E.P., Purnell J.Q. (2002) Plasma ghrelin levels after diet-induced weight loss or gastric bypass surgery. The New England Journal of Medicine 346, 1623-1630 [DOI] [PubMed] [Google Scholar]
- Dall R., Kanaley J., Hansen T.K., Moller N., Christiansen J.S., Hosoda H., Kangawa K., Jorgensen J.O. (2002) Plasma ghrelin levels during exercise in healthy subjects and in growth hormone-deficient patients. European Journal of Endocrinology 147, 65-70 [DOI] [PubMed] [Google Scholar]
- Dickhuth H.H., Röcker K., Mayer F., König D., Korsten-Reck U. (2004) Endurance training and cardial adaptation (athlete’s heart). Herz 29, 373-380 (In German with English abstract) [DOI] [PubMed] [Google Scholar]
- Dill D.B., Costill D.L. (1974) Calculation of percentage changes in volumes of blood, plasma, and red cells in dehydration. Journal of Applied Physiology 37, 247-248 [DOI] [PubMed] [Google Scholar]
- Durnin V.G.A., Womersly J. (1974) Body fat assessed from total body density and its estimation from skinfold thickness: measurements on 481 men and women aged from 16 to 72 years. British Journal of Nutrition 32, 77-97 [DOI] [PubMed] [Google Scholar]
- El Khoury D.T., Obeid O., Azar S.T., Hwalla N. (2006) Variations in postprandial ghrelin status following ingestion of high-carbohydrate, high-fat, and high-protein meals in males. Annals of Nutrution and Metabolism 50, 260-269 [DOI] [PubMed] [Google Scholar]
- Erdmann J., Tahbaz R., Lippl F., Wagenpfeil S., Schusdziarra V. (2007) Plasma ghrelin levels during exercise-Effects of intensity and duration. Regulatory Pept 143, 3127-3135 [DOI] [PubMed] [Google Scholar]
- Fisher G., Schwartz D.D., Quindry J.C., Barberio M.D., Foster E.B., Jones K.W., Pascoe D.D. (2010) Lymphocyte enzymatic antioxidant responses to oxidative stress following high-intensity interval exercise. Journal of Applied Physiology. (In press) [DOI] [PubMed] [Google Scholar]
- Flanagan D.E., Evans M.L., Monsod T.P., Rife F., Heptulla R.A., Tamborlane W.V., Sherwin R.S. (2003) The influence of insulin on circulating ghrelin. American Journal of Physiology-Endocrinology and Metabolism 284, E313-E316 [DOI] [PubMed] [Google Scholar]
- Flint A., Raben A., Blundell J.E., Astrup A. (2000) Reproducibility, power and validity of visual analogue scales in assessment of appetite sensations in single test meal studies. International Journal of Obesity and Related Metabolic Disorders 224, 38-48 [DOI] [PubMed] [Google Scholar]
- Hensrud D.D., Klein S. (2006) Extreme obesity: a new medical crisis in the United States. Mayo Clinic Proceedings 81, 5-10 [DOI] [PubMed] [Google Scholar]
- Hopkins M., King N.A., Blundell J.E. (2010) Acute and long-term effects of exercise on appetite control: is there any benefit for weight control?. Current Opinion in Clinical Nutrition and Metabolic Care 113, 635-40 [DOI] [PubMed] [Google Scholar]
- Ikeda N., Yasu Takanori., Tsuboi K., Sugawara Y., Kubo N., Umemoto T., Arao K., Kawakami M., Momomura S. (2010) Effects of submaximal exercise on blood rheology and sympathetic nerve activity. Circulation Journal 774, 730-734 [DOI] [PubMed] [Google Scholar]
- Imbeault P., Saint-Pierre S., Alméras N., Tremblay A. (1997) Acute effects of exercise on energy intake and feeding behaviour. British Journal of Nutrition 77, 511-521 [DOI] [PubMed] [Google Scholar]
- Inhoff T., Monnikes H., Noetzel S., Stengel A., Goebel M., Dinh Q.T., Riedl A., Bannert N., Wisser A.S., Wiedenmann B., Klapp B.F., Taché Y., Kobelt P. (2008) Desacyl ghrelin inhibits the orexigenic effect of peripherally injected ghrelin in rats. Peptides 229, 2159-2168 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jurimae J., Jurimae T. (2005) Leptin responses to short term exercise in college level male rowers. British Journal of Sports Medicine 39, 6-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jurimae J., Hofmann P., Jurimae T., Palm R., Mäestu J., Purge P., Sudi K., Rom K., von Duvillard S.P. (2007) Plasma ghrelin responses to acute sculling exercises in elite male rowers. European Journal of Applied Physiology 99, 467-474 [DOI] [PubMed] [Google Scholar]
- Kim H.J., Lee S., Kim T.W., Kim H.H., Jeon T.Y., Yoon Y.S., Oh S.W., Kwak H., Lee J.G. (2008) Effects of exercise-induced weight loss on acylated and unacylated ghrelin in overweight children. Clinical Endocrinology 68, 416-422 [DOI] [PubMed] [Google Scholar]
- King J.A., Wasse L.K., Broom D.R., Stensel D.J. (2010a) Influence Of Brisk Walking On Appetite, Energy Intake, And Plasma Acylated Ghrelin. Medicine and Science in Sports and Exercise 442 ,, 485-492 [DOI] [PubMed] [Google Scholar]
- King J.A., Miyashita M., Wasse L.K., Stensel D.J. (2010b) Influence of prolonged treadmill running on appetite, energy intake and circulating concentrations of acylated ghrelin. Appetite 54(3), 492-498 [DOI] [PubMed] [Google Scholar]
- King N.A., Burley V.J., Blundell J.E. (1994) Exercise-induced suppression of appetite: effects on food intake and implications for energy balance. European Journal of Clinical Nutrition 448, 715-724 [PubMed] [Google Scholar]
- Kirchner H., Gutierrez J.A., Solenberg P.J., Pfluger P.T., Czyzyk T.A., Willency J.A., Schürmann A., Joost H.G., Jandacek R.J., Hale J.E. (2009) GOAT links dietary lipids with the endocrine control of energy balance. Nature Medicine 115, 741-745 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kissileff H.R., Pi-Sunyer F.X., Segal K., Meltzer S., Foelsch P.A. (1990) Acute effects of exercise on food intake in obese and nonobese women. American Journal of Clinical Nutrition 552, 240-245 [DOI] [PubMed] [Google Scholar]
- Kojima M., Hosoda H., Date Y., Nakazato M., Matsuo H., Kangawa K. (1999) Ghrelin is a growth-hormone-releasing acylated peptide from stomach. Nature 402, 656-660 [DOI] [PubMed] [Google Scholar]
- Kraemer R.R., Durand R.J, Acevedo E.O., Johnson L.G., Kraemer G.R., Hebert E.P., Castracane V.D. (2004) Rigorous running increases growth hormone and insulin like growth factor-I without altering ghrelin. Experimental Biology and Medicine 229, 240-246 [DOI] [PubMed] [Google Scholar]
- Kumar R., Salehi A., Rehfeld J.F., Höglund P., Lindström E., Håkanson R. (2010) Proghrelin peptides: Desacyl ghrelin is a powerful inhibitor of acylated ghrelin, likely to impair physiological effects of acyl ghrelin but not of obestatin A study of pancreatic polypeptide secretion from mouse islets. Regulatory Peptides 164, 65-70 [DOI] [PubMed] [Google Scholar]
- Lely A.J., van der. Tschöp M., Heiman M.L., Ghigo E. (2004) Biological, physiological, pathophysiological, and pharmacological aspects of ghrelin. Endocrine Reviews 225, 426-457 [DOI] [PubMed] [Google Scholar]
- Lucidi P., Murdolo G., Di Loreto C., Parlanti N., De Cicco A., Ranchelli A., Fatone C., Taglioni C., Fanelli C., Santeusanio F., De Feo P. (2004) Meal intake similarly reduces circulating concentrations of octanoyl and total ghrelin in humans. Journal of Endocrinological Investigation 227, RC12-RC15. [PubMed] [Google Scholar]
- Mackelvie K.J., Meneilly G.S., Elahi D., Wong A.C.K., Barr S.I., Chanoine J.P. (2007) Regulation of appetite in lean and obese adolescents after exercise: role of acylated and desacyl ghrelin. Journal of Clinical Endocrinology and Metabolism 992, 648-654 [DOI] [PubMed] [Google Scholar]
- Maraki M., Tsofliou F., Pitsiladis Y.P., Malkova D., Mutrie N., Higgins S. (2005) Acute effects of a single exercise class on appetite, energy intake and mood. Is there a time of day effect? Appetite 45, 272-278 [DOI] [PubMed] [Google Scholar]
- Martins C., Morgan L.M, Bloom S.R., Robertson M.D. (2007) Effects of exercise on gut peptides, energy intake and appetite. Journal of Endocrinology 193, 251-258 [DOI] [PubMed] [Google Scholar]
- Martins C., Robertson M.D., Linda M.M. (2008) Effects of exercise and restrained eating behaviour on appetite control. Proceedings of the Nutrition Society 67, 28-41 [DOI] [PubMed] [Google Scholar]
- Marzullo P., Salvadori A., Brunani A., Verti B., Walker G.E., Fanari P., Tovaglieri I., Medici C.D., Savia G., Liuzzi A. (2008) Acylated ghrelin decreases during acute exercise in the lean and obese state. Clinical Endocrinology 69, 970-971 [DOI] [PubMed] [Google Scholar]
- Mattes R. (1990) Hunger ratings are not a valid proxy measure of reported food intake in humans. Appetite 115, 103-113 [DOI] [PubMed] [Google Scholar]
- Murdolo G., Lucidi P., Di Loreto C., Parlanti N., De Cicco A., Fatone C., Fanelli C.G., Bolli G.B., Santeusanio F., De Feo P. (2003) Insulin is required for prandial ghrelin suppression in humans. Diabetes 552, 2923-2927 [DOI] [PubMed] [Google Scholar]
- Muscaritoli M., Molfino A., Chiappini M.G., Laviano A., Ammann T., Spinsanti P., Melchiorri D., Inui A., Alegiani F., Rossi Fanelli F. (2007) Anorexia in hemodialysis patients: the possible role of des-acyl ghrelin. American Journal of Nephrology 227, 360-365 [DOI] [PubMed] [Google Scholar]
- Perini R., Veicsteinas A. (2003) Heart rate variability and autonomic activity at rest and during exercise in various physiological conditions. European Journal of Applied Physiology 90, 317-325 [DOI] [PubMed] [Google Scholar]
- Perboni S., Inui A. (2010) Appetite and gastrointestinal motility: Role of ghrelin-family peptides. Clinical Nutrition 29, 227-234 [DOI] [PubMed] [Google Scholar]
- Pomerleau M., Imbeault P., Parker T., Doucet E. (2004) Effects of exercise intensity on food intake and appetite in women. American Journal Clinical Nutrition 80, 1230-1236 [DOI] [PubMed] [Google Scholar]
- Powers. S.K., Howley. E.T. (2002) Exercise physiology: Theory and application to fitness and performance. 3th McGraw-Hill, New York: 167-168 [Google Scholar]
- Schmidt A., Maier C., Schaller G. (2004) Acute exercise has no effect on ghrelin plasma concentrations. Hormone and Metabolic Research 36, 174-177 [DOI] [PubMed] [Google Scholar]
- Shintani M., Ogawa Y., Ebihara K. (2001) Ghrelin, an endogenous growth hormone secretagogue is a novel orexigenic peptide that antagonizes leptin action through the activation of hypothalamic neuropeptide Y/Y1 receptor pathway. Diabetes 550, 227-232 [DOI] [PubMed] [Google Scholar]
- Shiya T., Nakazato M., Mizuta M., Date Y., Mondal M.S., Tanaka M., Nozoe S., Hosoda H., Kangawa K., Matsukura S. (2002) Plasma ghrelin levels in lean and obese humans and the effect of glucose on ghrelin secretion. Journal of Clinical Endocrinology and Metabolism 887, 240-244 [DOI] [PubMed] [Google Scholar]
- Siri W.E. (1961) Body composition from fluid spaces and density: analysis of methods. In: Techniques for measuring body composition. : Henschel A.National Academy of Sciences, Washington, National Research Council 223-244 [Google Scholar]
- Stengel A., Wisser A., Kobelt P., Goebel M., Wiedenmann B., Klapp B.F., Tache’ Y., Monnikes H. (2008) 987 Des-acyl ghrelin suppresses food intake and body weight gain in adult obese Zucker rats. Gastroenterology 134, A-148 [Google Scholar]
- Thompson D.A., Wolfe L.A., Eikelboom R. (1988) Acute effects of exercise intensity on appetite in young men. Medicine and Science in Sports and Exercise 220, 222-227 [DOI] [PubMed] [Google Scholar]
- Unick J.L., Otto A.D., Goodpaster B.H., Helsel D.L., Pellegrini C.A., Jakicic J.M. (2010) Acute effect of walking on energy intake in overweight/obese women. Appetite 55, 413-419 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vestergaard E.T., Dall R., Lange K.H.W., Kjaer M., Christiansen J.S., Jorgensen J.O.L. (2007) The ghrelin response to exercise before and after growth hormone administration. Journal of Clinical Endocrinology and Metabolism 92, 297-303 [DOI] [PubMed] [Google Scholar]
- Vriese De C., Delporte C. (2008) Ghrelin: A new peptide regulating growth hormone release and food intake. International Journal of Biochemistry and Cell Biology 40, 1420-1424 [DOI] [PubMed] [Google Scholar]
- Weir J.B. (1990) New methods for calculating metabolic rate with special reference to protein metabolism. 1949. Nutrition 6, 213-221 [PubMed] [Google Scholar]
- Westerterp-Plantenga M., Verwegen C.R, Ijedema M.J., Wijckmans N.E., Saris W.H. (1997) Acute effects of exercise or sauna on appetite in obese and non obese men. Physiological Behavior 62, 1345-1354 [DOI] [PubMed] [Google Scholar]
- Wren A.M., Seal L.J., Cohen M.A., Brynes A.E., Frost G.S., Murphy K.G., Dhillo W.S., Ghatei M.A., Bloom S.R. (2001) Ghrelin enhances appetite and increases food intake in humans. Journal of Clinical Endocrinology and Metabolism 86, 5992-5995 [DOI] [PubMed] [Google Scholar]
- Wren A.M., Small C.J., Ward H.L., Murphy K.G., Dakin C.L., Taheri S., Kennedy A.R., Roberts G.H., Morgan D.G., Ghatei M.A., Bloom S.R. (2000) The novel hypothalamic peptide ghrelin stimulates food intake and growth hormone secretion. Endocrinology 141, 4325-4328 [DOI] [PubMed] [Google Scholar]
- Zoladz J.A., Konturek S.J., Duda K., Majerczak J., Sliwowski Z., Grandys M., Bielanski W. (2005) Effect of moderate incremental exercise, performed in fed and fasted state on cardio-respiratory variables and leptin and ghrelin concentrations in young healthy men. Journal of Physiology and Pharmacology 56, 63-85 [PubMed] [Google Scholar]