

Abstract
Taurine included abundantly in skeletal muscle, particularly in the slow-twitch fibers, enhances exercise performance. However, the exact mechanisms for this effect have been unclear. The present study investigated the influence of taurine supplementation on amino acids profile in skeletal muscles as one of mechanisms in the enhancement of exercise performance induced by taurine. In the rats that received taurine solution, amino acids concentrations were comprehensively quantified in two portions with different fiber compositions in the fast-twitch fiber dominant (FFD) gastrocnemius muscle after 2 weeks, and in the gastrocnemius and additional other FFD muscles, liver, and plasma with exhausted exercise after 3 weeks. In the FFD muscles after 2 weeks, a common phenomenon that decreased concentrations of threonine (-16%), serine (-15~-16%), and glycine (-6~-16%) were observed, and they are categorized in the pyruvate precursors for hepatic gluconeogenesis rather than biosynthesis, polar, and side-chain structures. The decreases in the three amino acids were significantly emphasized after an additional week of taurine supplementation in the FFD muscles (p values in three amino acids in these tissues were less than 0.001-0.05), but not in the liver and plasma, accompanied with significantly increase of running time to exhaustion (p <0.05). In contrast, the three amino acids (threonine and serine; p < 0.05, glycine; p < 0.01) and alanine (p < 0.01) in the liver were significantly decreased and increased, respectively, following the exhaustive exercise. In conclusion, the taurine-induced reductions of these amino acids in skeletal muscle might be one of the mechanisms which underpin the enhancement of exercise performance by taurine.
Key points.
Taurine ingestion significantly decreased certain amino acids in skeletal muscles accompanied with enhanced exercise performance.
The decreased amino acids in common were threonine, serine, and glycine, but not alanine; pyruvate precursor for gluconeogenesis.
The alteration of three amino acids in muscles was maintained after exhausted exercise.
The muscular alterations of them might be one of taurine-induced roles on exercise performance.
Key words: Serine, glycine, threonine, gluconeogenic precursor, treadmill
Introduction
Taurine, 2-aminoethanesulfonic acid, is the most abundant amino acid-like compound in the body, and is found in most tissues including skeletal muscle, heart, nerve, brain, and liver in vertebrates (Jacobsen and Smith, 1968). The high tissue content of taurine is mainly derived from food intake, although the endogenous biosynthesis from the sulfur-containing amino acids methionine and cysteine in the liver is limited (Hosokawa et al., 1990). Previous studies have described many and various biological, physiological, and pharmacological roles of taurine including its conjugation to bile acids (Sjovall, 1959), membrane stabilization (Pasantes-Morales et al., 1985), detoxification (Huxtable, 1992), antioxidation (Gordon and Heller, 1992), osmoregulation (Thurston et al., 1980), modulation of ion flux (De Luca et al., 1996; Pierno et al., 1994), control of Ca2+ homeostasis (Bakker and Berg, 2002; Huxtable and Bressler, 1973), and its role as a neurotransmitter (Davison and Kaczmarek, 1971) or neuromodulator (Huxtable, 1992; Kuriyama, 1980).
In the skeletal muscle, taurine concentration is maintained at a high level by a specific transporter (TAUT) due to the absence of endogenous synthesis (Ramamoorthy et al., 1994). A close relationship between tissue taurine content and exercise performance is suggested by studies using TAUT knockout mice demonstrating a depletion of taurine in the skeletal muscle as well as other organs. This was likely due to the deficiency of exogenous taurine transport, which caused a strong reduction in overall exercise performance (Ito et al., 2008; Warskulat et al., 2004). In the skeletal muscle, the steady state level of taurine differs with muscle fiber type; higher in slow-twitch fiber than fast-twitch (FT) fiber (Iwata et al., 1986; Turinsky and Long, 1990). The difference in taurine content, depending on muscle fiber type, may have physiological significance for skeletal muscle function. We have demonstrated that the skeletal muscle taurine concentration was decreased after exhaustive exercise in rat, and the decreased concentration was significant in the fast-twitch fiber dominant (FFD) when compared to the slow-twitch fiber dominant muscles (Matsuzaki et al., 2002). Furthermore, our study showed in rat that taurine supplementation dose-dependently prevented the exercise-induced decreased tissue taurine concentration in the gastrocnemius (GC) muscle that is the FFD and protagonistic muscle in running (Sullivan and Armstrong, 1978), and consequently, significantly enhanced exercise performance (Miyazaki et al., 2004). Therefore, the FFD muscles with lower taurine content might be antecedently influenced by exercise, and profit from the taurine supplementation.
Previous studies have suggested that the mechanisms for the taurine-mediated enhancement in exercise performance might involve the reduction of exercise-induced oxidative stress (Dawson et al., 2002; Miyazaki et al., 2004; Zhang et al., 2004), increased cardiac contractility during exercise (Baum and Weiss, 2001), inhibition of exercise-induced blood lactate acid production (Imagawa et al., 2009; Manabe et al., 2003), or decreased exercise-induced muscular damage (Manabe et al., 2003). In addition, a recent study demonstrated that acute taurine supplementation in humans just before exercise produced significant increases in total whole-body fat oxidation, suggesting the possible mechanism of taurine to shift relative fuel utilization (Rutherford et al., 2010). Indeed, our previous report in humans demonstrated that taurine supplementation could maintain blood glucose concentrations during prolonged exercise (Ishikura et al., 2008). Harada et al. also showed that taurine suppmelentation improved hyperglycemia and insulin resistance with increased musclular glycogen conetent in the diabetic model rat (Harada et al., 2004). Therefore, it is suggested that there might be other possible roles of taurine on metabolism associated with carbohydrate, lipid, and amino acids for energy production. During endurance exercise, certain amino acids themselves may be utilized as fuel sources through the gluconeogenesis pathway. The best-characterized system in gluconeogenesis is the glucose-alanine cycle in which muscle alanine is catabolized in the liver for glucose homeostasis. In addition, branched-chain amino acids (BCAA); leucine, isoleucine, and valine, can be oxidized as fuels in skeletal muscle during prolonged exercise (Rennie et al., 2006). Furthermore, glutamine has been shown to enhance hepatic alanine and glutamine uptake and stimulate glucose production during exercise in dog (Iwashita et al., 2005). Thus, the amino acid metabolisms are important systems for fuel source during endurance exercise. Similarly, Korang et al., 1996 showed that the intraperitoneal injection of taurine caused significant changes in the concentrations of certain amino acids in cardiovascular tissues in rat. Therefore, exogenous administration of taurine could affect amino acid composition in some tissues.
We hypothesized that taurine supplementation might influence the alterations in amino acid concentrations in the FFD muscles, and that this may be a factor that enhances exercise performance, because taurine is the end product of the sulfur-containing amino acid metabolic pathway (Hosokawa et al., 1990). In this study, taurine supplementation for 2 weeks resulted in a common phenomenon to decrease the pyruvate precursor amino acids; threonine, serine, and glycine in the two portions with different FT fiber composition in the GC muscle in rat. Therefore, we focused on the alterations in these amino acids in the FFD muscles as well as liver and plasma after an additional week of taurine supplementation prior to exhaustive exercise.
Methods
Animals, taurine supplementation, and exercise
Male Fischer 344 rats, eight weeks of age, were purchased from Japan SLC (Shizuoka, Japan). The rats were randomly assigned to two groups; taurine-supplemented (Tau) and non-supplemented control (Con; Figure 1). The Tau group received 3% taurine solution in drinking water, while the Con group received non-supplemented water as vehicle. The dose of taurine was previously confirmed to certainly enhance the concentrations in blood and tissues with ad libitum ingestion (Harada et al., 2004). After 2 weeks, the rats in each group were divided into two- and three-week sub-groups. The two- week groups; 2wTau (n = 8) and 2wCon (n = 7), were sacrificed, and the GC muscles were removed after euthanasia. After an additional week, the three-week groups were further divided into sedentary and exercised sub-groups; non-supplemented and sedentary (3wCon/Sed; n = 7), taurine-supplemented and sedentary (3wTau/Sed; n = 7), non-supplemented and exercised (3wCon/Exe; n = 6), and taurine- supplemented and exercised (3wTau/Exe; n = 7) groups. The rats in the exercised groups were forced to perform transient treadmill running (4E9S-6; Fuji Ika Sangyo, Chiba, Japan) at 21.7 m/min until exhaustion using a blind method. Exhaustion was defined as the point at which the rats were not able to continue to run despite contact with an electric grid attached to the treadmill, and were not able to right themselves when placed on their backs (Miyazaki et al., 2004; Yatabe et al., 2003). All rats were fed a standard diet (MF; Oriental Yeast, Tokyo, Japan) and allowed access to drinking water ad libitum, but were fasted overnight before sacrifice. One week prior to exercise, all rats were loaded on the treadmill, running (10~25 m/min for 30min) once a day for 5 days to habituate to the treadmill apparatus (Saito and Soya, 2004). Body weight was monitored daily. The rats were singly housed at a constant room temperature (20- 26°C) with a 12-hour dark/light cycle, and received human care in accordance with The Guideline of the University of Tsukuba for the Care of Laboratory Animals.
Figure 1.
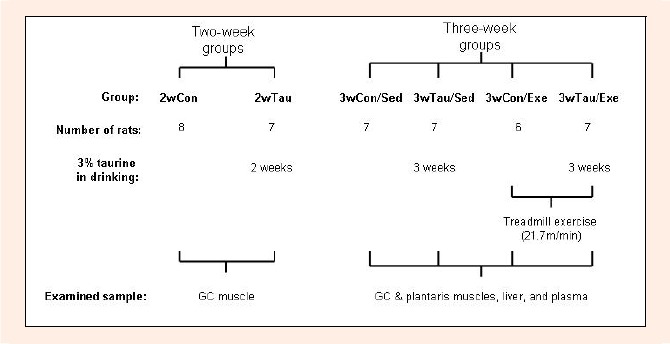
The experimental design. See the text in the methods section for a full description. Abbreviations: 2wCon; two-week non-supplemented, 2wTau; two-week taurine-supplemented, 3wCon/Sed; three-week non-supplemented and sedentary, 3wTau/Sed; three-week taurine-supplemented and sedentary, 3wCon/Exe; three-week non-supplemented and exercised, 3wTau/Exe; three-week taurine-supplemented and exercised, GC; gastrocnemius muscle.
Sample preparation
At the end of the experiment or exercise, all rats were euthanatized with sodium pentobarbital overdose, and blood, liver, and skeletal muscle (GC and plantaris) were immediately collected. In the collected GC muscle, the medial head was separated and was further divided into two portions because there were different compositions of fiber type; the red (GC-R) and white (GC-W) portions are comprised approximately 70% and 100% of FT fibers, respectively, while the plantaris muscle is almost 90% FT fiber (Sullivan and Armstrong, 1978). All samples were immediately frozen in liquid nitrogen and kept at -80 °C until assay.
Amino acids and glucose analysis
Taurine and other amino acid concentrations in samples were comprehensively determined by an automatic amino acid analyzer (JLC-300V; JEOL, Tokyo, Japan). Plasma was mixed with two-fold volumes of 3% sulphosalicylic acid, and centrifuged at 800 g for 15 min at 4 °C. Tissue was homogenized with 20 volumes of 5% TCA (Homogenizer T25 Basic Ultra Turrux; Ika Japan K.K., Nara, Japan), and centrifuged at 6,200 g for 30 min at 4 °C. The supernatants of plasma and tissue samples was passed through a 0.45 μl pore filter and stored at -20 °C until assay (Miyazaki et al., 2004; Yatabe et al., 2003).
Blood was also collected from the tail vein before and immediately after exercise, and blood glucose concentration was measured by the glucose oxidase method (Rivers et al., 2006).
Statistical analysis
All data are presented as the mean ± SD, and amino acid concentrations in the tissues are expressed as per mg wet weight. Differences between two groups and among multiple groups were analyzed by unpaired Student’s t-test and two-way ANOVA followed by Bonferroni’s post hoc analysis, respectively. Correlation relationships among data are expressed as Pearson’s correlation coefficients. All statistical analyses were performed using SPSS 11.0 (SPSS Japan Inc., Tokyo, Japan).
Results
Body weight progression and consumption of water, food, and taurine
Over the duration of the experiments, there was no significant difference in body weight (BW) progression between the Con and Tau groups. BW gain was 30 ± 6 g and 31 ± 4 g in the 2wCon and 2wTau groups, respectively, and 48 ± 10 g and 45 ± 10 g in the 3wCon and 3wTau groups, respectively. Dietary water intakes were 22.9 ± 1.9 and 22.5 ± 3.1 mL·day-1, and food intakes were 14.5 ± 1.2 and 14.1 ± 1.0 g·day-1 in the Con and Tau groups, respectively, and there were no significant differences in the intakes between the groups. All rats were in good health, with no pathological signs observed throughout the period of study.
Two-week taurine supplementation
At first, we attempted to categorize taurine affected amino acids by some nutritional, polar, structural, or metabolic factors; i.e., essential/non-essential, acidic/alkaline, side-chain radical, or gluconeogenesis/ketogenesis metabolism. As a result, the category of amino acids by the precursors of gluconeogenesis/ketogenesis pathways (Scheme) led to the common phenomenon of the effect of taurine supplementation on the amino acid alteration in the GC muscle, while there was no commonness in other categories.
Scheme.
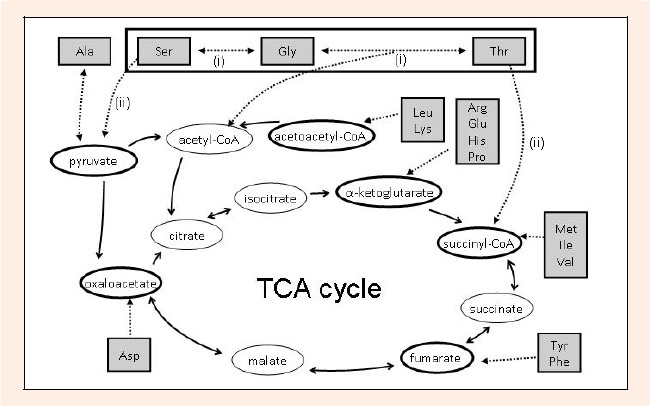
Amino acid catabolism in the gluconeogenesis pathway in rat. Serine, glycine, and threonine are reversibly and enzymatically interconverted via serine hydroxymethyltransferase (i). Serine is catabolized to pyruvate through a deamination reaction by serine/threonine dehydratase (ii). Threonine is also converted to acetyl-CoA and succinyl-CoA via these enzymes. The amino acids are categorized by the respective compounds in gluconeogenesis/ketogenesis to which they are precursors: Ser, Gly, Thr, and Ala; pyruvate, Leu and Lys; acetoacetyl-CoA, Arg, Glu, His, and Pro; -ketoglutarate, Met, Ile, and Val; succinyl-CoA, Tyr and Phe; fumarate, Asp; oxaloacetate precursors.
Table 1 show the amino acid concentrations categorized by the gluconeogenesis/ketogenesis precursors in two portions of GC muscle; GC-W and GC-R muscles, respectively, with or without taurine supplementation for 2 weeks. In both portions, the taurine concentration was significantly increased by the taurine supplementation for 2 weeks; particularly in the GC-W muscle was greater (47%; p < 0.001, and 15%; p < 0.05, increase in GC-W and GC-R, respectively). In the GC-W muscle, the concentrations of two pyruvate precursor amino acids, threonine (p < 0.01) and serine (p < 0.01), were significantly lower in the 2wTau group than in the 2wCon group. In addition, methionine (p < 0.01), histidine (p < 0.05), and lysine (p < 0.01) concentrations were significantly increased by the taurine supplementation. Similar to those in the GC-W muscle, threonine (p < 0.05) and glycine (p < 0.01) in the GC-R muscle were also significantly decreased by the taurine supplementation. There were no significant differences in other amino acids between the 2wCon and 2wTau groups in the GC-R muscle.
Table 1.
Categorized amino acid concentrations in the two portions of GC muscle following taurine supplementation for 2 weeks. Values are means (±SD).
| Amino acids | GC-W muscle | GC-R muscle | Category | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 2wCon | 2wTau | RC (±%) | 2wCon | 2wTau | RC (±%) | G/K Precursor | Essential | Polar & Side-Chain | |
| Tau | 14.81 (1.59) | 21.77 (.77) ‡ | (+47%) | 26.35 (1.61) | 30.34±3.27) † | (+15%) | |||
| Thr | .49 (.04) | .42 (.03) ‡ | (-16%) | .51 (.06) | .43 (.06)† | (-16%) | Pyr | Ess | Hydroxy |
| Ser | .66 (.07) | .56 (.04) ‡ | (-16%) | .80 (.11) | .68 (.12) | (-15%) | Pyr | Non | Hydroxy |
| Gly | 4.20 (.04) | 3.94 (.47) | (-6%) | 2.19 (.17) | 1.84 (.16) ‡ | (-16%) | Pyr | Non | Alkyl |
| Ala | 3.24 (.32) | 3.00 (.34) | (-7%) | 4.03 (.50) | 3.76 (.38) | (-7%) | Pyr | Non | Alkyl |
| Val | .26 (.05) | .27 (.06) | (+2%) | .26 (.06) | .26 (.04) | (-2%) | Suc-C | Ess | Alkyl |
| Met | .06 (.01) | .08 (.01) ‡ | (+31%) | .06 (.01) | .07 (.08) | (+14%) | Suc-C | Ess | Sulfur |
| Ile | .16 (.04) | .16 (.04) | (+3%) | .16 (.04) | .15 (.03) | (-6%) | Suc-C | Ess | Alkyl |
| Asp | .39(.07) | .41 (.06) | (+5%) | .59 (.08) | .54 (.11) | (-10%) | Oxa | Non | Acidic |
| Glu | .70 (.08) | .63 (.12) | (-10%) | 1.43 (.30) | 1.15 (.31) | (-20%) | a-keto | Non | Acidic |
| His | .18±0.01) | .20 (.01) † | (+6%) | .24 (.01) | .24 (.01) | (+1%) | a-keto | Ess | Alkaline |
| Arg | .17 (.02) | .20 (.04) | (+19%) | .20 (.04) | .22 (.04) | (+13%) | a-keto | Non | Alkaline |
| Pro | .32 (.04) | .31 (.01) | (-5%) | .32 (.02) | .31 (.05) | (-1%) | a-keto | Non | Imide |
| Tyr | .16 (.02) | .18 (.01) | (+9%) | .16 (.01) | .15 (.05) | (+6%) | Fum | Non | Aromatic |
| Phe | .09 (.02) | .10 (.02) | (+14%) | .09 (.01) | .10 (.01) | (+2%) | Fum | Ess | Aromatic |
| Lys | .40 (.05) | .58 (.01) ‡ | (+45%) | .50 (.11) | .62 (.13) | (+23%) | Ace-C | Ess | Alkaline |
| Leu | .20 (.05) | .21 (.02) | (+7%) | .21 (.05) | .20 (.04) | (-2%) | Ace-C | Ess | Alkyl |
Footnote: Amino acids concentrations are shorted by category of G/K precursor (mean ± SD). RC shows the rate of change in the 2wTau group compared to that in the 2wCon group. Category of the G/K precursor is the glucogenitic or ketogenitic precursors. These amino acids are also categorized by the essential amino acids, the polar characteristics (acidic, alkaline), and the side-chain radical (hydroxy-, alkyl-, sulfur-, imide-, and aromatic- radicals).
† p < 0.05,
‡ p < 0.01 for the Control vs. Taurine groups by student’s t-test.
Abbreviations: 2wCon; two-week non-supplemented group, 2wTau; two-week taurine-supplemented, GC-W; white portion of gastrocnemius muscle, GC-R; red portion of gastrocnemius muscle, Pyr: pyruvate, Suc-C: succinyl-CoA, Oxa; oxaloacetate; a-keto: a-ketoglutarate; Fum; fumarate, Ace-C: acetyl-CoA, Ess; essential amino acids, Non; non-essential amino acids
Three-week taurine supplementation: Exercise performance after taurine treatment
With taurine supplementation for 3 weeks, the duration of running to exhaustion was significantly increased compared to that in the non-supplemented group (76 ± 8 vs. 59 ± 11 min; p < 0.05). In the exercised rats, blood glucose levels in both the 3wCon and 3wTau groups were significantly lower at exhaustion than before exercise (before vs. after exercise; 79 ± 2 vs. 52 ± 6 mg·dl-1 in 3wCon; p < 0.05, 83 ± 4 vs. 45 ± 6 mg·dl-1 in 3wTau; p < 0.05). There were no significant differences in the blood glucose levels at exhaustion between the 3wCon and 3wTau groups.
Taurine concentration in tissues and plasma after taurine treatment with exercise
Table 2 shows taurine concentration in both portions of GC muscle, plantaris muscle, liver, and plasma after a transient exercise to exhaustion with or without taurine supplementation for 3 weeks. In the non-exercised rats, taurine concentration in all examined tissues and plasma was significantly increased by the three-week taurine supplementation, similar to those after the two-week supplementation (p < 0.001 in all samples). Likewise, in the exercised groups, a significant increase in taurine concentration in the 3wTau group was observed in all samples (p < 0.001 in all samples). This elevated concentration was maintained after the exercise. However, the taurine concentration in the GC-W muscle was significantly decreased in the 3wTau/Exe group compared to that in the 3wTau/Sed group (p < 0.05).
Table 2.
Taurine concentration in tissues and plasma following taurine supplementation for 3 weeks. Values are mean ± SD.
| Tissue | 3wCon/Sed | 3wTau/Sed | 3wCon/Exe | 3wTau/Exe | |
|---|---|---|---|---|---|
| GC-W | (nmol·mg-1) | 13.8 (.3) | 21.0 (.3) ‡ | 14.1 (.3) | 20.1 (.3) ‡* |
| GC-R | (nmol·mg-1) | 21.4 (.7) | 29.0 (.7) ‡ | 21.7 (.7) | 27.8 (.7) ‡ |
| Plantaris | (nmol·mg-1) | 18.3 (.6) | 24.0 (.6) ‡ | 17.8 (0.6) | 23.3 (.6) ‡ |
| Liver | (nmol·mg-1) | 2.1 (1.8) | 17.9 (1.8) ‡ | 5.0 (2.0) | 13.2 (1.8) ‡ |
| Plasma | (nmol·mg-1) | 326 (288) | 876 (561) ‡ | 481 (396) | 1201 (430) ‡ |
Footnote:
‡, p < 0.01 for the significant difference to the respective non-supplemented control group by two-way ANOVA.
*, p < 0.05 for the significant difference to the 3wTau/Sed group by two-way ANOVA followed by Bonferroni’s post hoc analysis.
Abbreviations: 3wCon/Sed; three-week non-supplemented and sedentary group, 3wTau/Sed; three-week taurine-supplemented and sedentary group, 3wCon/Exe; three-week non-supplemented and exercised group, 3wCon/Exe; three-week taurine-supplemented and exercised group, and see the footnote of Table 1 for other abbreviations.
Amino acid concentrations in tissues after taurine treatment
Following taurine supplementation for 2 weeks, the pyruvate precursor amino acids; threonine (p < 0.01) and serine (p < 0.01) in the GC-W muscle, and threonine (p < 0.05) and glycine (p <0.01) in the GC-R muscle, were significantly decreased. Therefore, we focused on the effects of an additional week of taurine supplementation and exercise to exhaustion on the concentrations of pyruvate precursor amino acids. Figure 2 shows the threonine (A), serine (B), glycine (C), and alanine (D) concentrations in the GC-W and GC-R muscles, plantaris muscle, liver, and plasma in the four groups. In Figure 2(A), threonine concentration was significantly decreased in the 3wTau/Sed and 3wTau/Exe groups compared to that in the respective control in all examined muscles; GC-W (p < 0.05 in both groups), GC-R (p < 0.001 in both groups), and plantaris (p < 0.05 in both groups). Similarly, serine (p <0.05, p < 0.01 and p < 0.001 in GC-W, GC-R, and plantaris, respectively in both groups) and glycine (p < 0.05, p < 0.001, and p < 0.001, respectively in both groups) concentrations were significantly decreased in the skeletal muscles by taurine supplementation in both the sedentary and exercised groups (Figure 2B and 2C). However, alanine concentration in the skeletal muscles was not affected by taurine supplementation in either the sedentary or exercised groups, although there was a significant difference between the 3wCon/Exe and 3wTau/Exe groups in the GC-R muscle (p < 0.05) (Figure 2D). In the liver and plasma, none of the four pyruvate precursor amino acids were influenced by taurine supplementation in either the sedentary or exercised groups.
Figure 2.
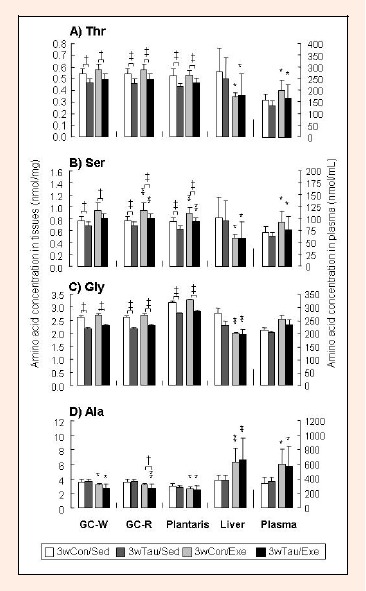
Pyruvate precursor amino acid concentrations in the tissues and plasma with or without taurine supplementation for 3 weeks or a transient exercise. Amino acid concentrations are shown for A) Threonine, B) Serine, C) Glycine, D) Alanine. Abbreviations: GC-W; white portion of gastrocnemius muscle, GC-R, red portion of gastrocnemius muscle. See the legend of Figure 1 for other abbreviations. Values are shown as the mean ± SD. Differences were analyzed by two-way ANOVA followed by Bonferroni’s post-hoc analysis. † with bar, p < 0.05; ‡ with bar, p < 0.01 for the non-supplemented vs. taurine-supplemented groups in the respective conditions. * without bar, p < 0.05; ** without bar, p <0 .01, for the exercised groups versus the respective sedentary group.
Amino acid concentrations in tissues with exercise
In both the 3wCon and 3wTau groups, neither threonine nor glycine concentrations in the skeletal muscles were changed by the exercise (Figure 2A and 2C). Serine and alanine concentrations were increased in the GC-R (p < 0.001 in both groups) and plantaris (p < 0.001 in both groups) muscles and decreased in the GC-W (p < 0.05 in both groups), GC-R (p < 0.001), and plantaris (p < 0.05 in both groups) muscles, respectively, in the exercised groups compared to that in the respective sedentary group, but this did not reach significance for the serine concentrations in the GC-W muscle (Figure 2B and 2D). In the liver, threonine (p < 0.05 in both groups), serine (p < 0.05 in both groups), and glycine (p < 0.01 in both groups) concentrations were significantly decreased by the exercise in both the 3wCon and 3wTau groups, while alanine (p < 0.01 in both groups) was significantly increased by the exercise. In plasma, the concentrations of all four amino acids were increased by the exercise in both the 3wCon and 3wTau groups, but this was not statistically significant for glycine (p < 0.05 in threonine and serine, and p < 0.01 in alanine in both groups).
Relationship between taurine and pyruvate precursor amino acids
Table 3 shows the correlation coefficients between the concentrations of taurine and pyruvate precursor amino acids in the tissues and plasma. In the skeletal muscles, there were significant inverse correlations between taurine and each pyruvate precursor amino acid, except for serine in the GC-R muscle. However, no correlation between taurine and alanine was found in any of the skeletal muscle types tested. In the liver and plasma, there were no significant correlations between the concentration of taurine and any of these amino acids.
Table 3.
Correlation coefficients between taurine and each pyruvate precursor amino acid following taurine supplementation for 3 weeks.
| Thr | Ser | Gly | Ala | |
|---|---|---|---|---|
| GC-W | -.476 * | -.477 * | -.482 * | -.144 |
| GC-R | -.530 * | -.254 | -.847 ** | -.033 |
| Plantaris | -.791 ** | -.612 ** | -.882 ** | -.090 |
| Liver | -.168 | -.151 | -.300 | -.070 |
| Plasma | .171 | .211 | .212 | .348 |
Footnote:
*, p < 0.05;
**, p < 0.01 for correlations with taurine concentration analyzed by Pearson’s correlation coefficient.
See footnote in Table 2 for abbreviations.
Discussion
In the present study, we investigated the effects of taurine supplementation on amino acid concentrations in the skeletal muscles, liver, and plasma, and the influence of a transient exercise-to-exhaustion regime on the amino acid levels in these tissues. First, in the FFD muscles following taurine supplementation for 2 weeks, there was a common phenomenon to decreased concentrations of the pyruvate precursor amino acids; threonine, serine, and glycine, rather than amino acids categorized into other gluconeogenesis pathways. Therefore, we focused on the alterations in the pyruvate precursor amino acids after an additional week of taurine supplementation in the FFD muscles, liver, and plasma. We found that taurine supplementation for 3 weeks significantly decreased the concentrations of threonine, serine, and glycine, but not alanine, in all examined FFD muscles. To our knowledge, this is the first report demonstrating that taurine supplementation caused a decrease in these pyruvate precursor amino acids in the skeletal muscle despite the contradictory increase of taurine concentration.
There are several reports in experimental animals that have comprehensively evaluated the relationship between taurine and the three amino acid alterations in the skeletal muscle under various conditions. In the Zucker diabetic fatty rat, significant decreases of threonine, serine, and glycine concentrations have been shown, while the contradictory increase of taurine concentration was observed in the skeletal muscle (Wijekoon et al., 2004). Conversely, the significant decrease of taurine in the skeletal muscle induced by 2-guanidinoethanesulphonate, a structural analogue of taurine, combined with a taurine-free diet caused a corresponding increase of those three amino acids (Marnela et al., 1984). Based on the previous and present findings, the three amino acids appear to be competitively altered by the presence of taurine, specifically in the skeletal muscle of animals. In human, one previous study examined the amino acids concentrations in the biopsied vastus lateralis muscle following taurine supplementation for one week (Galloway et al., 2008). Although glycine concentration was significantly decreased, serine and threonine concentrations were unchanged and significantly increased, respectively, without the effectiveness of taurine supplementation on muscular taurine level and exercise performance. Although there is the difference of species, the similar effectiveness of taurine found in the present study would be expected in human if the muscular taurine level could be enhanced by any efficient protocols of taurine supplementation.
The reasons for the taurine-induced decrements of the specific amino acids in the skeletal muscles should be considered. In general, only three amino acids are not used for biosynthesis into protein, and the amino acids metabolized in the skeletal muscle are only BCAAs, alanine, aspartic acid, and glutamic acid. Therefore, it is suggested that the significant decreases in the specific three amino acids might be a result of either lower uptake or higher release from the skeletal muscle. The tissue balances of amino acids are tightly regulated by amino acids transport systems (Palacín et al., 1998). Until now, many amino acid transporters have been cloned, and its characteristic and substrate specificity have been clarified to depend on the polar and also the structures side chain-, amino-, and carboxyl-radicals (Palacín et al., 1998). Since there is no commonness of these conditions for transport in three amino acids, it is the lower possibility that taurine might affect on these uptake into skeletal muscle via a specific transporter. However, the direct dynamics of amino acids in the skeletal muscle need to be clarified in the future study.
Several studies have demonstrated the importance of muscle taurine level on exercise performance in the taurine supplemented human and experimental animals and the taurine deficient model mice (Dawson et al., 2002; Ito et al., 2008; Miyazaki et al., 2004; Warskulat et al., 2004; Yatabe et al., 2003). The mechanisms of taurine-induced enhancement of performance have still been debated, but the taurine-induced alteration of the certain amino acids in skeletal muscles might be one of the potential mechanisms. Indeed, previous studies have suggested the possibility that taurine might affect some metabolic abilities for energy production (Harada et al., 2004; Ishikura et al., 2008; Mozaffari et al., 1986; Rutherford et al., 2010). In rat, the taurine-depleted heart, abnormal energy metabolism to stimulate significantly glycolysis and glycogenolysis was found (Mozaffari et al., 1986), while the opposite responses including the promotion of glycogen synthesis were observed in the isolated heart with perfusion of taurine (Lampson et al., 1983). In our previous study, taurine supplementation in human prevented the reduction of serum glucose level during prolonged exercise (Ishikura et al., 2008). In the present study, there was no significant difference in blood glucose level at the exhaustion points between the non-supplemented and taurine-supplemented groups, although the exercised duration was significantly prolonged by the taurine supplementation. In this point, we suggested the blood glucose level at certain points before the exhaustion might be higher in the taurine-supplemented group than in the non-supplemented group. In general, blood glucose level is maintained during exercise by the augmented release of glucose from the liver. With prolonged exercise, liver glycogen stores are depleted, and the rate of glucose production fails to keep pace with the rate of glucose use, resulting in a fall in glucose concentrations (Ahlborg and Felig, 1982). Gluconeogenesis from amino acids is one of the mechanisms of hepatic glucogenesis. In the present study, threonine, serine, and glycine concentrations in the liver were significantly decreased after exercise. Accordingly, we speculated that taurine might be indirectly involved in hepatic gluconeogenesis from amino acids, by modulating amino acid balance in the whole body.
In the present study, the significant increases of methionine, lysine, and histidine in the GC-W muscle were found after 2 weeks of taurine supplementation. To our knowledge, there is no study to observe that taurine treatment would alter the concentrations of these amino acids in the skeletal muscles. Contrasted to the GC-W muscle in the two-week group, the commonly alterations of these amino acids were not found in the GC-R muscle in the two-week group (Table 1) as well as in the both GC-W and GC-R muscles and plantaris muscle in the three-week taurine supplemented groups (no data shown). Furthermore, these amino acids do not belong to the same categories examined in the present study, although they are essential amino acids. Therefore, it is unclear why these amino acids were affected by taurine supplementation for 2 weeks only in the GC-W muscle.
Conclusion
In conclusion, taurine supplementation significantly decreased the concentrations of the pyruvate precursor amino acids; threonine, serine, and glycine, particularly in the skeletal muscles accompanied with the enhancement of exercise performance in rat. Therefore, the reduction of the three muscular amino acids utilized for gluconeogenesis induced by taurine supplementation might be one of the possible mechanisms for the enhancement of exercise performance by taurine.
Acknowledgments
This study was supported by a grant from the Chemical Analysis Center at the University of Tsukuba. We would like to thank Taisho Pharmaceutical Co., Ltd (Tokyo, Japan) for the provision of taurine.
Biographies

Keisuke Ishikura
Employment:
PhD student, Doctoral Program of Health and Sport Sciences, Graduate School of Comprehensive Human Sciences, University of Tsukuba, Japan
Degree:
MSc
Research interests:
Exercise physiology.
E-mail: storagestone@yahoo.co.jp

Teruo Miyazaki
Employment:
Assistant Research Professor, Department of development for community medicine, Tokyo Medical University, Japan
Degree:
PhD
Research interests:
Physiology, nutrition, and development of skeletal muscle, and metabolic interaction between muscle and liver.
E-mail: teruom@tokyo-med.ac.jp

Song-Gyu Ra
Employment:
PhD student, Doctoral Program of Health and Sport Sciences, Graduate School of Comprehensive Human Sciences, University of Tsukuba, Japan
Degree:
MSc
Research interests:
Exercise physiology.
E-mail: ra.song.gyu@gmail.com

Shoji Endo
Employment:
Bachelor of physical Education, School of Health and Physical Education, University of Tsukuba, Japan
Degree:
BSc
Research interests:
Exercise physiology.
E-mail: es052598@kentaku.co.jp

Yusuke Nakamura
Employment:
Under graduate school, School of Health and Physical Education, University of Tsukuba, Japan
Degree:
BSc
Research interests:
Exercise physiology.
E-mail: santa_pr1@yahoo.co.jp

Takashi Matsuzaka
Employment:
Assistant Professor, Department of Internal Medicine (Endocrinology and Metabolism), Graduate School of Comprehensive Human Sciences, University of Tsukuba, Japan
Degree:
PhD
Research interests:
Energy metabolism, metabolic syndrome, diabetes, atherosclerosis.
E-mail: t-matsuz@md.tsukuba.ac.jp

Shumpei Miyakawa
Employment:
Professor, Doctoral Program in Sports Medicine, Graduate School of Comprehensive Human Sciences, University of Tsukuba, Japan
Degree:
MD, PhD
Research interests:
Sports medicine, preventive medicine.
E-mail: miyakawa@taiiku.tsukuba.ac.jp

Hajime Ohmori
Employment:
Associate Professor, Doctoral Program of Health and Sport Sciences, Graduate School of Comprehensive Human Sciences, University of Tsukuba, Japan
Degree:
PhD
Research interests:
Effects of exercise on the physiological and biochemical functions of the brain, muscle and peripheral tissues.
E-mail: ohmori@taiiku.tsukuba.ac.jp
References
- Ahlborg G., Felig P. (1982) Lactate and glucose exchange across the forearm, legs, and splanchnic bed during and after prolonged leg exercise. The Journal of Clinical Investigation 69, 45-54 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bakker A., Berg H. (2002) Effect of taurine on sarcoplasmic reticulum function and force in skinned fast-twitch skeletal muscle fibres of the rat. The Journal of Physiology 538, 185-194 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baum M., Weiss M. (2001) The influence of a taurine containing drink on cardiac parameters before and after exercise measured by echocardiography. Amino Acids 20, 75-82 [DOI] [PubMed] [Google Scholar]
- Davison A., Kaczmarek L. (1971) Taurine--a possible neurotransmitter? Nature 234, 107-108 [DOI] [PubMed] [Google Scholar]
- Dawson R.J., Biasetti M., Messina S., Dominy J. (2002) The cytoprotective role of taurine in exercise-induced muscle injury. Amino Acids 22, 309-324 [DOI] [PubMed] [Google Scholar]
- De Luca A., Pierno S., Camerino D. (1996) Effect of taurine depletion on excitation-contraction coupling and Cl- conductance of rat skeletal muscle. Europian Journal of Pharmacology 296, 215-222 [DOI] [PubMed] [Google Scholar]
- Galloway S., Talanian J., Shoveller A., Heigenhauser G., Spriet L. (2008) Seven days of oral taurine supplementation does not increase muscle taurine content or alter substrate metabolism during prolonged exercise in humans. Journal of Applied Physiology 105, 643-651 [DOI] [PubMed] [Google Scholar]
- Gordon R., Heller R. (1992) Taurine protection of lungs in hamster models of oxidant injury: a morphologic time study of paraquat and bleomycin treatment. Advances in Experimental Medicine and Biology 315, 319-328 [DOI] [PubMed] [Google Scholar]
- Harada N., Ninomiya C., Osako Y., Morishima M., Mawatari K., Takahashi A., Nakaya Y. (2004) Taurine alters respiratory gas exchange and nutrient metabolism in type 2 diabetic rats. Obesity Research 12, 1077-1084 [DOI] [PubMed] [Google Scholar]
- Hosokawa Y., Matsumoto A., Oka J., Itakura H., Yamaguchi K. (1990) Isolation and characterization of a cDNA for rat liver cysteine dioxygenase. Biochemical and Biophysical Research Communucations 168, 473-478 [DOI] [PubMed] [Google Scholar]
- Huxtable R. (1992) Physiokogical action of taurine. In, the American Physiological Society, USA, Vol. 72, 101-163 [DOI] [PubMed] [Google Scholar]
- Huxtable R., Bressler R. (1973) Effect of taurine on a muscle intracellular membrane. Biochimica et Biophysica Acta 323, 573-583 [DOI] [PubMed] [Google Scholar]
- Imagawa T., Hirano I., Utsuki K., Horie M., Naka A., Matsumoto K., Imagawa S. (2009) Caffeine and taurine enhance endurance performance. International Journal of Sports Medicine 30, 485-488 [DOI] [PubMed] [Google Scholar]
- Ishikura K., Miyakawa S., Yatabe Y., Takekoshi K., Omori H. (2008) Effect of taurine supplementation in blood glucose concentration during prolonged exercise (in Japanese with Englishi abstruct). Japanese Journal of Physical Fitness Sports Medicine 57, 475-484 [Google Scholar]
- Ito T., Kimura Y., Uozumi Y., Takai M., Muraoka S., Matsuda T., Ueki K., Yoshiyama M., Ikawa M., Okabe M., Schaffer S., Fujio Y., Azuma J. (2008) Taurine depletion caused by knocking out the taurine transporter gene leads to cardiomyopathy with cardiac atrophy. Journal of Molecular and Cellular Cardiology 44, 927-937 [DOI] [PubMed] [Google Scholar]
- Iwashita S., Williams P., Jabbour K., Ueda T., Kobayashi H., Baier S., Flakoll P. (2005) Impact of glutamine supplementation on glucose homeostasis during and after exercise. Journal of Applied Physiology 99, 1858-1865 [DOI] [PubMed] [Google Scholar]
- Iwata H., Obara T., Kim B., Baba A. (1986) Regulation of taurine transport in rat skeletal muscle. Journal of Neurochemistry 47, 158-163 [DOI] [PubMed] [Google Scholar]
- Jacobsen J., Smith L. (1968) Biochemistry and physiology of taurine and taurine derivatives. Physiological Reviews 48, 424-511 [DOI] [PubMed] [Google Scholar]
- Korang K., Milakofsky L., Hare T., Hofford J., Vogel W. (1996) Levels of taurine, amino acids and related compounds in plasma, vena cava, aorta and heart of rats after taurine administration. Pharmacology 52, 263-270 [DOI] [PubMed] [Google Scholar]
- Kuriyama K. (1980) Taurine as a neuromodulator. Federation Proceedings 39, 2680-2684 [PubMed] [Google Scholar]
- Lampson W., Kramer J., Schaffer S. (1983) Potentiation of the actions of insulin by taurine. Canadian Journal of Physiology and Pharmacology 61, 457-463 [DOI] [PubMed] [Google Scholar]
- Manabe S., Kurroda I., Okada K., Morishima M., Okamoto M., Harada N., Takahashi A., Sakai K., Nakaya Y. (2003) Decreased blood levels of lactic acid and urinary excretion of 3-methylhistidine after exercise by chronic taurine treatment in rats. Journal of Nutritional Science and Vitaminology 49, 375-380 [DOI] [PubMed] [Google Scholar]
- Marnela K., Kontro P., Oja S. (1984) Effects of prolonged guanidinoethanesulphonate administration on taurine and other amino acids in rat tissues. Medical Biologyg 62, 239-244 [PubMed] [Google Scholar]
- Matsuzaki Y., Miyazaki T., Miyakawa S., Bouscarel B., Ikegami T., Tanaka N. (2002) Decreased taurine concentration in skeletal muscles after exercise for various durations. Medicine and Science in Sports and Exercise 34, 793-797 [DOI] [PubMed] [Google Scholar]
- Miyazaki T., Matsuzaki Y., Ikegami T., Miyakawa S., Doy M., Tanaka N., Bouscarel B. (2004) Optimal and effective oral dose of taurine to prolong exercise performance in rat. Amino Acids 27, 291-298 [DOI] [PubMed] [Google Scholar]
- Mozaffari M., Tan B., Lucia M., Schaffer S. (1986) Effect of drug-induced taurine depletion on cardiac contractility and metabolism. Biochemical Pharmacology 35, 985-989 [DOI] [PubMed] [Google Scholar]
- Palacín M., Estévez R., Bertran J., Zorzano A. (1998) Molecular biology of mammalian plasma membrane amino acid transporters. Physiological Reviews 78, 969-1054 [DOI] [PubMed] [Google Scholar]
- Pasantes-Morales H., Wright C., Gaull G. (1985) Taurine protection of lymphoblastoid cells from iron-ascorbate induced damage. Biochemical Pharmacology 34, 2205-2207 [DOI] [PubMed] [Google Scholar]
- Pierno S., Tricarico D., De Luca A., Campagna F., Carotti A., Casini G., Conte Camerino D. (1994) Effects of taurine analogues on chloride channel conductance of rat skeletal muscle fibers: a structure-activity relationship investigation. Naunyn- Schmiedeberg’s Archives of Pharmacology 349, 416-421 [DOI] [PubMed] [Google Scholar]
- Ramamoorthy S., Leibach F., Mahesh V., Han H., Yang-Feng T., Blakely R., Ganapathy V. (1994) Functional characterization and chromosomal localization of a cloned taurine transporter from human placenta. The Biochemical Journal 300 (Pt 3), 893-900 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rennie M., Bohé J., Smith K., Wackerhage H., Greenhaff P. (2006) Branched-chain amino acids as fuels and anabolic signals in human muscle. The Journal of Nutrition 136, 264S-268S [DOI] [PubMed] [Google Scholar]
- Rivers S., Kane M., Bakst G., Busch R., Hamilton R. (2006) Precision and accuracy of two blood glucose meters: FreeStyle Flash versus One Touch Ultra. American Journal of Health-System Pharmacy 63, 1411-1416 [DOI] [PubMed] [Google Scholar]
- Rutherford J., Spriet L., Stellingwerff T. (2010) The effect of acute taurine ingestion on endurance performance and metabolism in well-trained cyclists. International Journal of Sport Nutrition and Exercise Metab 20, 322-329 [DOI] [PubMed] [Google Scholar]
- Saito T., Soya H. (2004) Delineation of responsive AVP-containing neurons to running stress in the hypothalamus. American Journal of Physiology. Regulatory, Integrative and Comparative Physiology 286, R484-490 [DOI] [PubMed] [Google Scholar]
- Sjovall J. (1959) Dietary glycine and taurine on bile acid conjugation in man; bile acids and steroids 75. Proceedings of the Society for Experimental Biology and Medicine 100, 676-678 [DOI] [PubMed] [Google Scholar]
- Sullivan T., Armstrong R. (1978) Rat locomotory muscle fiber activity during trotting and galloping. Journal of Applied Physiology 44, 358-363 [DOI] [PubMed] [Google Scholar]
- Thurston J., Hauhart R., Dirgo J. (1980) Taurine: a role in osmotic regulation of mammalian brain and possible clinical significance. Life Sciences 26, 1561-1568 [DOI] [PubMed] [Google Scholar]
- Turinsky J., Long C. (1990) Free amino acids in muscle: effect of muscle fiber population and denervation. The American Journal of Physiology 258, E485-491 [DOI] [PubMed] [Google Scholar]
- Warskulat U., Flögel U., Jacoby C., Hartwig H., Thewissen M., Merx M., Molojavyi A., Heller-Stilb B., Schrader J., Häussinger D. (2004) Taurine transporter knockout depletes muscle taurine levels and results in severe skeletal muscle impairment but leaves cardiac function uncompromised. The FASEB journal 18, 577-579 [DOI] [PubMed] [Google Scholar]
- Wijekoon E., Skinner C., Brosnan M., Brosnan J. (2004) Amino acid metabolism in the Zucker diabetic fatty rat: effects of insulin resistance and of type 2 diabetes. Canadian Journal of Physiology and Pharmacology 82, 506-514 [DOI] [PubMed] [Google Scholar]
- Yatabe Y., Miyakawa S., Miyazaki T., Matsuzaki Y., Ochiai N. (2003) Effects of taurine administration in rat skeletal muscles on exercise. Journal of Orthopaedic Science 8, 415-419 [DOI] [PubMed] [Google Scholar]
- Zhang M., Izumi I., Kagamimori S., Sokejima S., Yamagami T., Liu Z., Qi B. (2004) Role of taurine supplementation to prevent exercise-induced oxidative stress in healthy young men. Amino Acids 26, 203-207 [DOI] [PubMed] [Google Scholar]