

Abstract
Previous studies have shown changes in brain activity as a result of exercise; however, few studies have examined changes during exercise. The purpose of this study was to examine brain activity during a graded exercise test. Twenty male participants performed a graded exercise test on a recumbent cycle ergometer. Exercise intensity was set initially at 50W and was increased by 50W every 2 minutes until volitional fatigue was reached. Electroencephalography (EEG) was measured prior to the onset of exercise, during the last minute of each stage of exercise, immediately post-exercise, and 10 minutes into recovery. EEG was recorded from 8 scalp sites leading to analysis of alpha 1, alpha 2, beta 1, beta 2, and theta activities. Expired air was collected and analyzed for ventilation rate (VE), VO2, % of peak VO2, and Respiratory Exchange Ratio (RER). No differences were seen in EEG between the hemispheres of the brain. There was, however, a significant increase in brain activity across the spectrum occurring at 200 W through immediately post-exercise. Brain activity returned to pre- exercise levels by 10 minutes post. VO2, % of peak VO2 and RER increased linearly with exercise intensity. VE increased linearly through 200 W; however, a disproportionate increase was seen in VE from 200 W to peak exercise. The results of this investigation demonstrate that brain activity may be related to exercise intensity. Future research will want to examine how these changes in brain activity influence affective, perceptual and cognitive changes often associated with exercise. Efforts will also need to be made to determine if changes in brain activity during exercise are mediated by central (within the brain) or peripheral mechanisms.
Key points.
EEG can be recorded during exercise.
Brain EEG activity increases during exercise and may be related to exercise intensity.
Brain EEG activity returns to resting levels quickly after the cessation of exercise.
Key words: Exercise, EEG, brain activity
Introduction
Brain activity has been proposed to be important in examining affective and perceptual responses to acute bouts of exercise (Nybo and Nielsen, 2001; Petruzzello et al., 2006). For example, changes in brain activity may occur secondary to the metabolic changes associated with “central fatigue” during prolonged exercise (Dalsgaard and Secher, 2007; Davis and Bailey, 1997; Newsholme and Blomstand, 1995). Furthermore, activity in the frontal regions of the brain has been related to affective (Petruzzello et al., 2006) and perceptual (Nybo and Nielsen, 2001) responses to acute bouts of exercise.
Before the relationship between brain activity and affective and perceptual responses to exercise can be understood, normal responses in brain wave activity to various types of exercise must first be determined. Electroencepahlography (EEG) currently is the most pragmatic way to monitor changes in brain activity in humans while exercising. Although several studies have described EEG changes immediately after exercise, very few studies have examined EEG during exercise. Furthermore, the few studies that have examined EEG changes during exercise have typically focused on a limited range of exercise intensities and examined only a few electrode sites.
For example, Kakizaki, 1988 found an increase in the amplitude of the Beta2 frequency range of EEG during cycling at 90 W at a single electrode site (Oz), but found no changes at lower workloads (50 - 80 W). Kubitz and Mott, 1996 described changes in EEG during 15 minutes (three, 5 minutes stages) of cycling at progressively greater workloads (ranging from 50 to approximately 150 W). During this investigation, electrodes were placed at frontal (F3 & F4) and temporal (T3 & T4) sites and EEG changes were described globally (i.e. not specific to the electrode sites). Kubitz and Mott, 1996 found no change in either alpha or beta activity from pre- to post-exercise, but did observe reductions in alpha activity and increases in beta activity during exercise.
Nybo and Nielsen (2001) described changes in EEG during 1 hour of exercise at 60% VO2max in normal (18°C) and hot (40°C) environments. These authors reported no changes in EEG as represented by percentage change from rest in the normal environment (18°C); however, when exercising in the hot environment increases in the ratio of alpha to beta frequency were seen. During this investigation, observations of EEG were limited to three electrode sites: one in the frontal region (F3), one at the midline for the central and one at the midline for occipital cortices. Previous work examining the relationship between affective responses to exercise and EEG following exercise, have suggested that different affective responses to exercise are associated with EEG responses which are specific to either the right or left side of the frontal cortex (Petruzzello et al., 2006). Unfortunately, the experimental protocol used by Nybo and Nielsen (2001) did not allow these authors to address potential EEG differences between the left and right sides of the brain.
Crabbe and Dishman, 2004 conducted a meta-analysis examining EEG responses during and after exercise and concluded that when compared to pre-exercise, alpha activity (absolute power) was greater both during and following exercise, but there was no differences in relative alpha activity. They also found that delta, theta and beta activity increased during and following exercise. Some of the limitations of previous research as pointed out in the review by Crabbe and Dishman, 2004 were that previous studies have examined narrow ranges of the EEG frequency spectrum and only a few electrodes were monitored. In order for EEG to become a useful tool in understanding the impact of various experimental manipulations (e.g. distraction, fatigue, nutritional supplementation) on brain activity during exercise, the EEG responses to exercise under normal controlled conditions must be better understood.
The purpose of this investigation was to examine a more comprehensive range of the EEG spectrum which included theta, alpha 1, alpha 2, beta 1 and beta 2, and to examine multiple sites, specifically lateral frontal (F7 & F8), mid-frontal (F3 & F4), central (C3 & C4) and parietal (P3 & P4) sites during varying intensities of graded aerobic exercise to volitional fatigue. It is hypothesized that EEG activity will increase across the EEG spectrum as workload increases in most but not all of the examined electrode sites.
Methods
Participants
Twenty healthy males with the following characteristics (mean age ± SE): age 24.0 ± 1.5 years, height 1.80 ± 0.02 m, weight 82.3 ± 3.6 kg, and VO2max 40. 4 ± 2.2 ml·kg-1·min-1 participated in this study. Prior to participating in this investigation all participants read and signed an informed consent approved by the university's Institutional Review Board for human subjects committee. Participants also completed the Physical Activity Readiness Questionnaire (PAR-Q) (Thomas et al., 1992) and were deemed “low risk” according to American College of Sports Medicine risk stratification criteria (ACSM, 2000).
Experimental protocol
Each participant performed a graded exercise test while electroencephalography (EEG; see below), heart rate (Polar Vantage Model XL monitor) and indirect calorimetry (Max 1, Physiodyne Inc., Quogue, NY) data were collected. After preparing the participant for EEG acquisition, two 1-minute baseline recordings were made while they rested, with eyes open, on the recumbent cycle ergometer. Before exercise, 2 - 3 minutes of expired gases were collected to acclimate the participant to the mask and nose clip. Throughout the tests expired gases were sampled over 15 seconds and then averaged over 60 seconds. Following the baseline measurement of expired gases, the participant started cycling at 50W for the first stage. The intensity of the graded exercise test was increased 50W every 2 minutes until the participant reached volitional exhaustion. The graded exercise test was conducted on a recumbent cycle, and not on a treadmill or upright cycle, because it was believed that the recumbent position would limit movement artifact allowing for cleaner EEG recordings.
EEG was recorded (eyes open) during the second minute of each 2-miunte stage of the exercise test. At the point of volitional exhaustion, the participant was asked to stop pedaling and EEG was collected for 1-minute immediate post exercise EEG recording. Participants were then asked to rest (non-active recovery) for 10 minutes on the recumbent cycle ergometer before a final 1-minute EEG recording.
EEG recordings
In accordance with the International 10-20 system, 21 electrodes were placed on the surface of subject's scale using a stretchable lycra electrode cap (Electro-Cap, Inc.). Ocular artifact was assessed by electro-oculogram (EOG) recordings from electrodes placed above and below the participant's right eye. All leads were referenced to linked earlobes and impedances were below 5 KΩ. All procedures were consistent with those proposed by the American Electroencephalographic Society (Allen et al., 2004). While EEG was recorded at all 21 electrode sites, this investigation focused on the data produced at 8 sites (F3, F4, F7, F8, C3, C4, P3, P4) associated with 3 different cortical areas. The data that was statistically analyzed in this investigation was limited to these 8 sites in order to not overwhelm the reader. These sites were selected based on work done by previous investigators and in an attempt to represent multiple brain areas. Topographical maps showing the relative change in EEG activity in all 21 electrode sites are presented in Figure 1.
Figure 1.
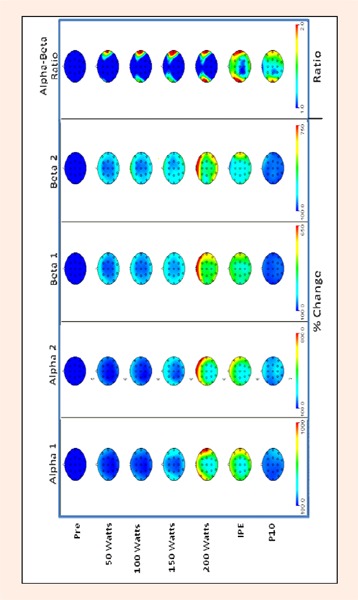
Topographical depiction of relative changes in EEG activity in 21 electrode sites during graded exercise to fatigue.
The EEG data was amplified using the Psylab Stand Alone Monitor (SAM; Contact Precision Instruments Inc.) system and sampled at 512 Hz. The gain was set at 50,000 and the data was high and low pass filtered at 3 Hz and 40 Hz, respectively (24 dB/octave).
EEG was processed and analyzed using Brain Vision Analyzer (Version 1.04, Brain Products GmH). First, the 1-minute recordings were blindly segmented into 30, 2-second epochs and visually inspected for artifact. The epochs were individually transformed to power spectra using a Hanning window, subject to Fast Fourier Transform and then averaged. Second, the total power in the theta (4.50 - 7.99), alpha 1 (8.00 - 10.49 Hz), alpha 2 (10.50-12.99), beta 1 (13.00-17.99 Hz) and beta 2 (18.00-30.00) frequency bands were measured from the average power spectrum and normalized to the average power in the same frequency bands during the two baseline recordings.
Statistical analysis
A 7 x 4 x 2 (Time x Region x Hemisphere) multivariate analysis of variance was conducted to determine if there were any differences in EEG spectral power across the different exercise intensities and sites. Significance was set a priori at the p ¼ 0.05 level. When significant main effects were observed a Newman-Keuls Multiple-Comparison Test was performed. The results of these comparisons will be shown in the tables and figures. For all values, means ± SE are reported in the text or presented in a table or figure. Cardiorespiratory data is presented in Table 1. In Figures 2, 3, 4, 5, 6 and 7, values for each frequency are expressed as a percentage of the average pre-exercise recordings to reduce inter-individual variability as suggested by Nybo and Nielsen (2001). The topographical maps presented in Figure 1 were from relative changes in EEG activity for Alpha, 1, Alpha 2, Beta 1, Beta 2, and Alpha/Beta Ratio using a spherical spline interpolation provided via Brain Vision Analyzer (Version 1.04, Brain Products GmH).
Table 1.
Cardiorespiratory responses to graded exercise test to fatigue (M+SE) (n=20) on a recumbent cycle ergometer. Data are means (±SD).
| 50 Watts | 100 Watts | 150 Watts | 200 Watts | Peak exercise | |
|---|---|---|---|---|---|
| Ventilation (L.min-1) | 16.5 (.9) | 23.2 (.8)* | 35.9 (1.1)* | 52.3 (1.8)* | 88.6 (4.0)* |
| VO2 (mL.min-1) | 852 (52) | 1323 (40)* | 1996 (41)* | 2541 (36)* | 3285 (107)* |
| % of Peak VO2 | 26.2 (1.1) | 40.9 (1.5)* | 61.3 (1.8)* | 78.6 (2.3)* | NA |
| RER | .82 (.02) | .84 (.02) | .95 (.03)* | 1.08 (.02)* | 1.18 (.03)* |
| Heart Rate (b.min-1) | 93 (3) | 108 (3)* | 133 (3)* | 157 (4)* | 179 (3)* |
* indicates difference from previous time point.
Figure 2.
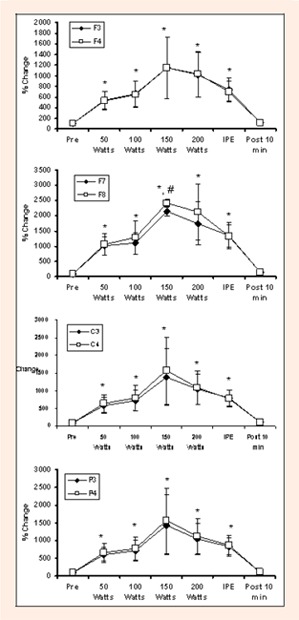
Changes in Theta activity during graded exercise to fatigue at several EEG sites. * indicates difference from Pre. # indicates difference from 50 Watts.
Figure 3.
Changes in Alpha 1 activity during graded exercise to fatigue at several EEG sites. * indicates difference from Pre, 50 Watts, 100 Watts, 150 Watts and Post 10 min. # indicates difference from Pre, 50 Watts and 100 Watts.
Figure 4.
Changes in Alpha 2 activity during graded exercise to fatigue at several EEG sites. * indicates difference from Pre, 50 Watts, 100 Watts, 150 Watts and Post 10 min. # indicates difference from Pre, 50 Watts and 100 Watts.
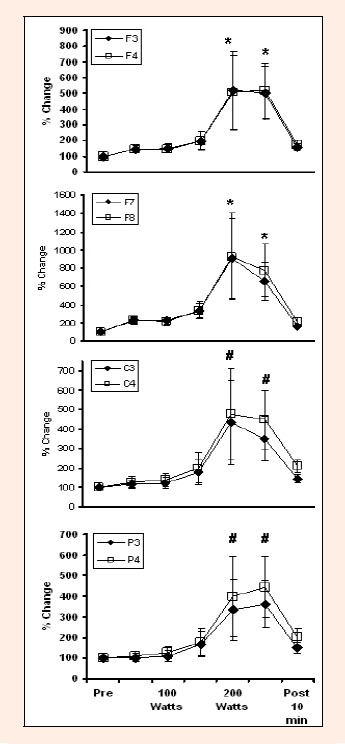
Figure 5.
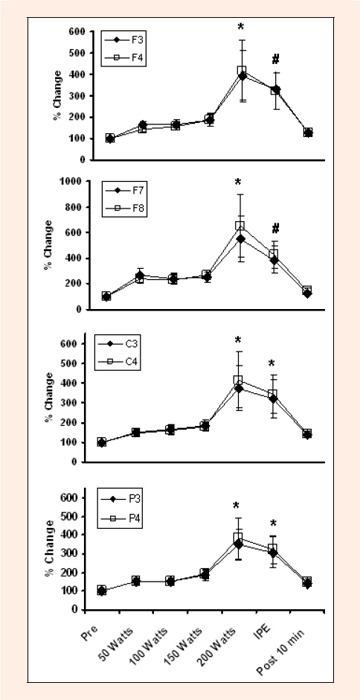
Changes in Beta 1 activity during graded exercise to fatigue at several EEG sites. * indicates difference from Pre, 50 Watts, 100 Watts, 150 Watts and Post 10 min. # indicates difference from Pre and Post 10 min.
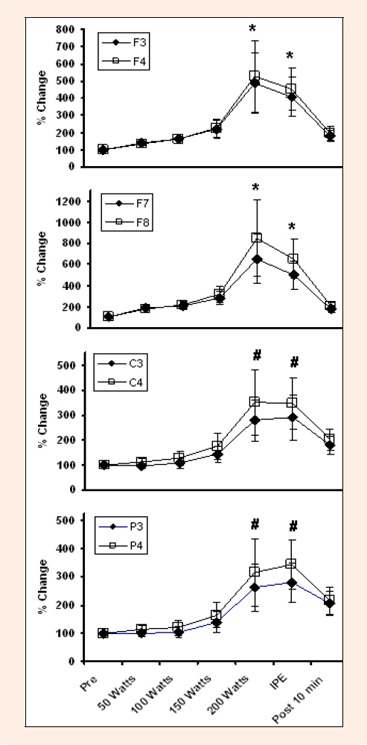
Figure 6.
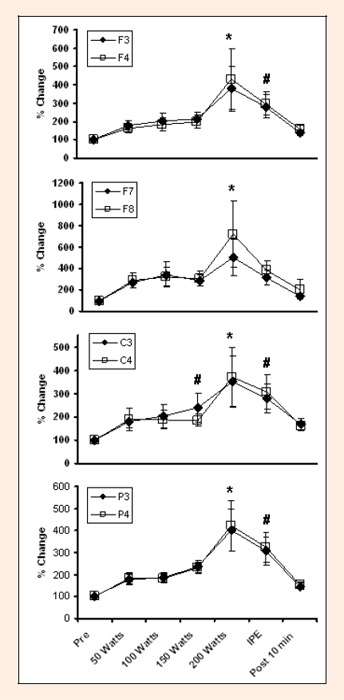
Changes in Beta 2 activity during graded exercise to fatigue at several EEG sites. * indicates difference from Pre, 50 Watts, 100 Watts, 150 Watts and Post 10 min. # indicates difference from Pre and Post 10 min.
Figure 7.
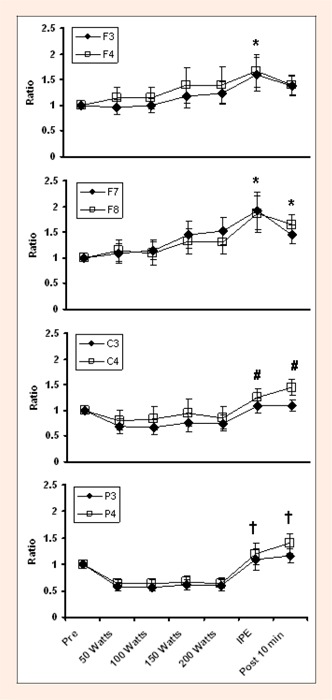
Changes in Alpha-Beta ratio during graded exercise to fatigue at several EEG sites. * indicates difference from Pre. # indicates difference from 50 Watts and 100 Watts. † indicates difference from 50 watts, 100, watts, 150 watts and 200 watts.
Results
The cardiorespiratory responses to graded exercise are presented in Table 1. Peak VO2 values (absolute Peak VO2= 3285 ± 107 ml.min-1, relative VO2= 40.4 ± 2.2 ml.kg-1.min-1) suggest that these subjects had an average to below average aerobic fitness level (ACSM, 2000).
A multivariate analysis of variance was conducted to determine if there are any differences in theta, alpha 1, alpha 2, beta 1 and beta 2 frequencies over time or by site. Below is a description of relative changes in EEG during exercise. These changes are presented in Figures 2, 3, 4, 5, 6 and 7.
A significant main effect was found for Time [Wilks' λ = 0.75, F (1, 36) = 8.94, p ¼ 0.0001], Region [Wilks' λ = 0.92, F (1, 18) = 5.41, p ¼ 0.0001], and a Time x Region interaction [Wilks' λ = 0. 88, F (1, 108) = 1.30, p = 0.03], but not for Hemisphere, Time x Hemisphere interaction or Time x Region x Hemisphere interaction. The significant main effect of Time was attributable to changes in Theta, F (1, 6) = 15.65, p ¼ 0.0001, Alpha 1, F (1, 6) = 5.47, p ¼ 0.0001, Alpha 2, F (1, 6) = 13.61, p ¼ 0.0001, Beta 1, F (1, 6) = 18.94, p ¼ 0.0001, and Beta 2 F (1, 6) = 17.59, p ¼ 0.0001. The significant main effect for Region was attributable to changes in Alpha 2, F (1, 3) = 6. 89, p ¼ .0001, Beta 1, F (1, 3) = 2.89, p = .034, and Beta 2, F (1, 3) = 4.03, p = .007. These results can be seen in Figures 2, 3, 4, 5, and 6. In general, EEG activity increased (p ¼ 0.05) for all waves, in all leads during maximal graded exercise. For all wave forms, EEG activity became significantly increased (p ¼ .05) at 150 or 200 watts, remained elevated immediately post exercise (IPE); p ¼ .05), and approached baseline by 10 minutes post exercise.
An analysis of variance was also conducted to determine if there are any differences in alpha-beta ratio over time or by site. A significant main effect was found for Time [F (1, 6) = 8.44, p ¼ . 0001] and Region [F (1, 3) = 18.27, p ¼ .0001], but not for Time x Region, Hemisphere, Time x Hemisphere interaction or Time x Region x Hemisphere interaction. These results can be seen in Figure 7. While trends in alpha-beta ratio differed between leads during exercise (F3/F4 & F7/F8 gradually increased, C3/C4 & P3/P4 decreased), alpha-beta ratio was significantly increased (p ¼ 0.05) at IPE in all leads and waveforms. Alpha-beta ratio remained elevated (p ¼ .05) at 10 minutes post exercise in all leads except F3/F4.
Discussion
The purpose of this investigation was to examine the changes in EEG activity during and following an acute bout of incremental graded exercise at numerous EEG sites and at multiple EEG frequencies. The results of this investigation indicate the EEG activity is increased above resting levels at higher workloads and at fatigue during graded exercise. These increases were seen across EEG frequencies (theta, alpha 1, alpha 2, beta 1, beta 2) and electrode sites (F3, F4, F7, F8, C3, C4, P3, P4). Furthermore, EEG changes were not localized to either hemisphere.
The results from the present study are consistent with research from previous studies that have shown that during and following exercise there is an increase in EEG activity in the theta, alpha and/or beta frequencies (Crabbe and Dishman, 2004; Kamijo et al., 2004; Nielsen et al., 2001; Nybo and Nielsen, 2001). Unlike previous studies, the present study showed these changes occurred at multiple electrode sites (8 different sites) and across EEG frequencies.
Interestingly, Kubitz and colleagues (1996; 1997) observed decreases in alpha activity during exercise, while the present investigation observed increases in alpha activity. The mechanism underlying these different findings is unclear; however, it is reasonable to speculate that they may be related to specific differences in the exercise protocol. Subjects in this investigation completed graded exercise to fatigue, while subjects in the work done by Kubitz and colleagues (1997) exercised for a finite period of time (15 minutes) at predetermined exercise intensities. The differences between our findings and those reported by Kubitz and colleagues (1997) underscore the need for further investigation in this area.
The present study is the first investigation to document intensity related increases in EEG activity during graded exercise to fatigue. Few studies have examined changes in EEG as a result of differing exercise intensities (Kamijo et al., 2004; Mechau et al., 1998), but none of these studies examined EEG during the exercise bout. One study examined EEG during a rest period following running five or six discontinuous stages with increasing velocity (Mechau et al., 1998). During the investigation by Mechau and colleagues, the first three stages were below the lactate threshold, while the last two or three stages resulted in lactate accumulation (Mechau et al., 1998). In the exercise conditions without blood lactate accumulation there were increases in theta, delta and alpha-1 frequency bands. At the higher intensities, with blood lactate accumulation, there were significant decreases in the beta-2, beta-1 and alpha-1 frequency bands. Upon initial comparison it appears that the results of the present investigation are contradictory to those described by Mechau and colleagues (1998) because we observed increases in these same frequency bands during exercise at higher workloads. These contrasting findings may simply be a consequence of the timing of the EEG recording (during or immediately after exercise) and strengthen the argument for monitoring EEG during exercise (rather than immediately after) when attempting to describe changes in brain activity during exercise. Furthermore, it seems appropriate to begin differentiating changes in brain activity during acute exercise from changes in brain activity subsequent to chronic exercise.
In order to create a better understanding of the acute effects of exercise on brain activity, future investigations should determine if EEG activity changes as a result of exercise duration (independent of exercise intensity) alone. Furthermore, the presumption that changes in EEG activity during exercise represent changes in cognitive brain activity and perception may not be appropriate. In order to address this issue, functional measures of brain activity (i.e. cognitive function), affect and perception should be simultaneously measured.
A methodological concern of measuring EEG during exercise is the potential impact of physiologic and mechanical parameters on the electrical signal. Measures that could impact the electrical signal include but are not limited to heart rate, ventilation, pCO2, sweat rate, and pedal frequency. While it is impossible to completely eliminate the impact of these factors on EEG, they were minimized by filtering out electrical signals below 3 Hz and cycling in a semi-recumbent posture. It does appear that investigations where exercise is performed at similar workloads, but varying postures (recumbent, semi-recumbent, and upright cycling) would be valuable to insure that the changes in EEG activity during exercise primarily reflect changes in brain electrical activity.
When examining EEG changes during exercise it is also important to consider the underlying mechanisms that drive these changes. In the current investigation, it appears that increases in EEG activity in the Alpha 1, Alpha 2, Beta 1, and Beta 2 bands mirror increases in VE during exercise. In comparison, increases in EEG activity in the Theta band seem to be more significant at lower workloads than increases in VE. Furthermore, increases in EEG activity during exercise occurred at all electrode sites and were not specific to one side of the brain. As a consequence, it is reasonable to conclude that the observed changes in EEG activity may have been driven by changes in peripheral physiology and not within the brain itself. This hypothesis is supported by Kraaier and colleagues (1988, 1992) who have described an increase in EEG subsequent to a decrease in pCO2 produced by hyperventilation. Future, experiments should be designed to address this potential issue.
Conclusion
In conclusion, the present study demonstrates that EEG can be recorded during exercise. The findings of the study documented increases in theta, alpha and beta frequency activity during exercise and at multiple electrode sites. Even though there was not specificity, in region or hemisphere, in these results, future studies may want to examine how these changes in EEG influence or are associated with changes in cognition, affect and/or perception during exercise. The impact of peripheral physiology on EEG during exercise also needs to be examined.
Biographies

Stephen P. Bailey
Employment
Department of Physical Therapy Education, Elon University.
Degree
PT, PhD
Research interests
Changes in brain function during exercise, Role of central fatigue during prolonged exercise.
E-mail: baileys@elon.edu

Eric E. Hall
Employment
Department of Health and Human Performance, Elon University.
Degree
PhD
Research interests
Influence of exercise intensity on affective responses and cognitive function.
E-mail: ehall@elon.edu

Stephen E. Folger
Employment
Department of Physical Therapy Education, Elon University.
Degree
PT, PhD
E-mail: folgers@elon.edu

Paul C. Miller
Employment
Department of Health and Human Performance, Elon University.
Degree
PhD
Research interests
Adaptations to exercise; The use of supplements and their impact on patterns of muscle recovery and growth.
E-mail: millerp@elon.edu
References
- Allen J.J.B., Coan J.A., Nazarian M. (2004) Issues and assumptions on the road from raw signals to metrics of frontal EEG asymmetry in emotion. Biological Psychology 67, 183-218 [DOI] [PubMed] [Google Scholar]
- American College of Sports Medicine. (2000) ACSM's guidelines for exercise testing and prescription. 6th edition. Lippincott Williams & Wilkins, Baltimore [Google Scholar]
- Crabbe J.B., Dishman R.K. (2004) Brain electrocortical activity during and after exercise: A quantitive synthesis. Psychophysiology 441, 563-574 [DOI] [PubMed] [Google Scholar]
- Davis J.M., Bailey S.P. (1997) Possible mechanisms of central nervous system fatigue during exercise. Medicine and Science in Sport and Exercise 29(1), 45-57 [DOI] [PubMed] [Google Scholar]
- Dalsgaard M.K., Secher N.H. (2007) The brain at work: a cerebral metabolic manifestation of central fatigue?. Journal of Neuroscience Research 85, 3334-3339 [DOI] [PubMed] [Google Scholar]
- Kakizaki T. (1988) Effects of bicycle exercise on occipital EEG amplitude in male students. Industrial Health 26, 191-195 [DOI] [PubMed] [Google Scholar]
- Kamijo K., Nishihira Y., Hatta A., Kaneda T., Kida T., Higashiura T., Kuroiwa K. (2004) Changes in arousal level by differential exercise intensity. Clinical Neurophysiology 1115, 2693-2698 [DOI] [PubMed] [Google Scholar]
- Kraaier V., Van Huffelen A.C., Wieneke G.H. (1988) Changes in quantitative EEG and blood flow velocity due to standardized hyperventilation; a model of transient ischaemia in young human subjects. Electroencephalography and Clinical Neurophysiology 70, 377-387 [DOI] [PubMed] [Google Scholar]
- Kraaier V., Van Huffelen A.C., Wieneke G.H., Van der Worp H.B., Bär P.R. (1992) Quantitative EEG changes due to cerebral vasoconstriction. Indomethacin versus hyperventilation-induced reduction in cerebral blood flow in normal subjects. Electroencephalography and Clinical Neurophysiology 82, 208-212 [DOI] [PubMed] [Google Scholar]
- Kubitz K.A., Mott A.A. (1996) EEG power spectral densities during and after cycle ergometer exercise. Research Quarterly for Exercise and Sport 667(1), 91-96 [DOI] [PubMed] [Google Scholar]
- Kubitz K.A., Pothakos K. (1997) Does aerobic exercise decrease brain activation?. Journal of Sport and Exercise Psychology 119, 291-301 [Google Scholar]
- Newsholme E.A., Blomstrand E. (1995) Tryptophan, 5-hydroxytryptamine and a possible explanation for central fatigue. Advances in Experimental Medicine and Biology 384, 315-320 [DOI] [PubMed] [Google Scholar]
- Nielsen B., Hgldig T., Bidstrup F., Gonzalez-Alonso J., Christoffersen G.R.J. (2001) Brain activity and fatigue during prolonged exercise in the heat. European Journal Applied Physiology 4442, 41-48 [DOI] [PubMed] [Google Scholar]
- Nybo L., Nielsen B. (2001) Perceived exertion is associated with an altered brain activity during exercise with progressive hyperthermia. Journal of Applied Physiology 991, 2017-2023 [DOI] [PubMed] [Google Scholar]
- Mechau D., Mucke S., Weiss M., Liesen H. (1998) Effect of increasing running velocity on electroencephalogram in a field test. European Journal Applied Physiology 778(4), 340-345 [DOI] [PubMed] [Google Scholar]
- Petruzzello S.J., Ekkekakis P., Hall E.E. (2006) Physical activity and affect: EEG studies. Psychobiology of exercise and sport. Acevedo E.O., Ekkekakis P.Human Kinetics, Champaign, IL: 111-128 [Google Scholar]
- Thomas S., Reading J., Shephard R.J. (1992) Revision of the Physical Activity Readiness Questionnaire (PAR-Q). Canadian Journal of Sports Science 117(4), 338-345 [PubMed] [Google Scholar]