

We thank Campioni et al. for their response to our recent publication1 in which they focus on the relevance of other molecules besides IL-6 and IL-10 for the immunomodulatory functions of multipotent stromal cells (MSC).
In our publication, we had focused on the mechanistic roles for IL-6 and IL-10 in the immunosuppressive effects of MSC. However, we agree that other factors may be important as well. We discuss here how the various, sometimes paradoxical, observations can be combined into an integrated view on the mechanism of immunodulation by MSC.
Although IL-6 had previously been proposed to be involved in the inhibitory effect of MSC on the differentiation of monocytes to dendritic cells (DC),2,3 the cell source had not been assigned. IL-10 was also previously indicated to be a key factor in monocyte-to-DC differentiation4 and to be related to MSC-modulated immune suppression.5 In our study, we showed that IL-6 is produced by unstimulated MSC and that it initiates an effect on monocytes resulting in their differentiation towards a monocyte-derived cell population (MDC), similar to type-II activated macrophages with concomitant enhanced production of IL-10. Also, IL-6 expression by MDC was significantly enhanced upon co-culture with MSC.
In their letter, Campioni et al. correctly point out that other factors, especially prostaglandin E2 (PGE2) and HLA-G, have previously been implicated in the immunomodulatory functions of MSCs. In fact, using a similar test system, it has been reported that PGE2 and not IL-6 represents the key inhibitory mediator.6 We agree that the data from that study strongly suggest a major role for PGE2 in the MSC-mediated inhibition of monocyte-to-DC differentiation. It is assumed, however, that PGE2 is MSC-derived. Since the presented data are based on the measurement of PGE2 concentrations in monocyte/MSC co-culture systems, it cannot be excluded that the PGE2 is produced by monocytes or MDC. While Spaggiari et al. have shown that IL-6 is not induced by PGE2, the possibility that monocyte-derived IL-6 is involved in PGE2 production by MSC cannot be excluded. This would agree with their observation that PGE2 production by MSC is up-regulated in monocyte/MSC co-cultures.
HLA-G is also a major player in immune regulation by MSC.7–9 It has been shown that HLA-G is directly regulated by IL-10.9,10 Since MSC skew monocytes to MDC11,12 which display prominent IL-10 expression, this seems a logical link to the HLA-G related immunosuppressive action of MSC. This is in perfect agreement with the observation that monocytes/MDC are indispensable for MSC to exert many of their immunosuppressive functions.13–15
We have combined the various observations into our current working model for immune regulation by MSC, as shown in Figure 1.
Figure 1.
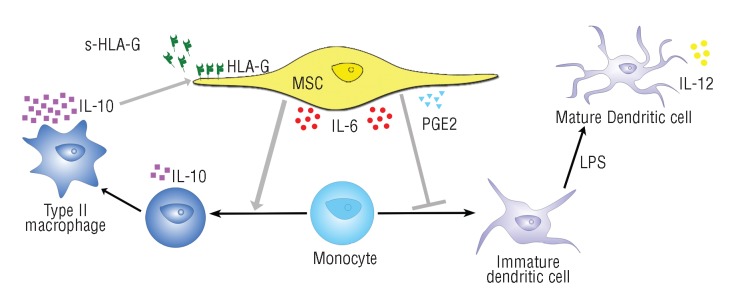
Immunomodulation by MSC. MSC express IL-6 and PGE2 that skew monocyte differentiation toward the formation of IL-10-expressing MDC. This IL-10 activates MSC to up-regulate (soluble and membrane-bound) HLA-G expression, which is linked to immunomodulatory effects on NK and adaptive immune cells. Through cell-cell contact, IL-10, IL-6 or another factor secreted after monocyte-MSC interaction, MSC are activated to up-regulate their PGE2 expression, further enforcing the monocyte-to-MDC skewing.
Footnotes
Information on authorship, contributions, and financial & other disclosures was provided by the authors and is available with the online version of this article at www.haematologica.org.
References
- 1.Melief SM, Geutskens SB, Fibbe W, Roelofs H. Multipotent stromal cells skew monocytes towards an anti-inflammatory IL-10 producing phenotype by production of IL-6. Haematologica. 2013. (In press) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Nauta AJ, Kruisselbrink AB, Lurvink E, Willemze R, Fibbe WE. Mesenchymal Stem Cells Inhibit Generation and Function of Both CD34+-Derived and Monocyte-Derived Dendritic Cells. J Immunol. 2006;4(177):2080–7 [DOI] [PubMed] [Google Scholar]
- 3.Djouad F, Charbonnier LM, Bouffi C, Louis-Plence P, Bony C, Apparailly F, et al. Mesenchymal stem cells inhibit the differentiation of dendritic cells through an interleukin-6-dependent mechanism. Stem Cells. 2007;8(25):2025–32 [DOI] [PubMed] [Google Scholar]
- 4.Allavena P, Piemonti L, Longoni D, Bernasconi S, Stoppacciaro A, Ruco L, et al. IL-10 prevents the differentiation of monocytes to dendritic cells but promotes their maturation to macrophages. Eur J Immunol. 1998;1(28):359–69 [DOI] [PubMed] [Google Scholar]
- 5.Beyth S, Borovsky Z, Mevorach D, Liebergall M, Gazit Z, Aslan H, et al. Human mesenchymal stem cells alter antigen-presenting cell maturation and induce T-cell unresponsiveness. Blood. 2005; 5(105):2214–9 [DOI] [PubMed] [Google Scholar]
- 6.Spaggiari GM, Abdelrazik H, Becchetti F, Moretta L. MSCs inhibit monocyte-derived DC maturation and function by selectively interfering with the generation of immature DCs: central role of MSC-derived prostaglandin E2. Blood. 2009;26(113):6576–83 [DOI] [PubMed] [Google Scholar]
- 7.Selmani Z, Naji A, Zidi I, Favier B, Gaiffe E, Obert L, et al. Human leukocyte antigen-G5 secretion by human mesenchymal stem cells is required to suppress T lymphocyte and natural killer function and to induce CD4+CD25highFOXP3+ regulatory T cells. Stem Cells. 2008;1(26):212–22 [DOI] [PubMed] [Google Scholar]
- 8.Nasef A, Mathieu N, Chapel A, Frick J, Francois S, Mazurier C, et al. Immunosuppressive effects of mesenchymal stem cells: involvement of HLA-G. Transplantation. 2007;2(84):231–7 [DOI] [PubMed] [Google Scholar]
- 9.Rizzo R, Campioni D, Stignani M, Melchiorri L, Bagnara GP, Bonsi L, et al. A functional role for soluble HLA-G antigens in immune modulation mediated by mesenchymal stromal cells. Cytotherapy. 2008;4(10):364–75 [DOI] [PubMed] [Google Scholar]
- 10.Rizzo R, Lanzoni G, Stignani M, Campioni D, Alviano F, Ricci F, et al. A simple method for identifying bone marrow mesenchymal stromal cells with a high immunosuppressive potential. Cytotherapy. 2011;5(13):523–7 [DOI] [PubMed] [Google Scholar]
- 11.Kim J, Hematti P. Mesenchymal stem cell-educated macrophages: a novel type of alternatively activated macrophages. Exp Hematol. 2009;12(37):1445–53 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Zhang QZ, Su WR, Shi SH, Wilder-Smith P, Xiang AP, Wong A, et al. Human gingiva-derived mesenchymal stem cells elicit polarization of m2 macrophages and enhance cutaneous wound healing. Stem Cells. 2010;10(28):1856–68 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Francois M, Romieu-Mourez R, Li M, Galipeau J. Human MSC suppression correlates with cytokine induction of indoleamine 2,3-dioxygenase and bystander M2 macrophage differentiation. Mol Ther. 2012;1(20):187–95 [DOI] [PubMed] [Google Scholar]
- 14.Cutler AJ, Limbani V, Girdlestone J, Navarrete CV. Umbilical cord-derived mesenchymal stromal cells modulate monocyte function to suppress T cell proliferation. J Immunol. 2010;11(185):6617–23 [DOI] [PubMed] [Google Scholar]
- 15.Melief SM, Schrama CLM, Brugman MH, Tiemessen MM, Hoogduijn MJ, Fibbe WE, et al. Multipotent stromal cells induce human regulatory T cells through a novel pathway involving skewing of monocytes toward anti-inflammatory macrophages. Stem Cells. 2013. (In press). [DOI] [PubMed] [Google Scholar]