

Abstract
Chemoprevention represents a strategy designed to protect cells or tissues against various carcinogens and carcinogenic metabolites derived from exogenous or endogenous sources. Recent studies indicate that plant-derived triterpenoids, like oleanolic acid, may exert cytoprotective functions via regulation of the activity of different transcription factors. The chemopreventive effects may be mediated through induction of the nuclear factor erythroid 2-related factor 2 (Nrf2) transcription factor. Activation of Nrf2 by triterpenoids induces the expression of phase 2 detoxifying and antioxidant enzymes such as NAD(P)H quinone oxidoreductase 1 (NQO1) and heme oxygenase-1 (HO-1) - proteins which can protect cells or tissues against various toxic metabolites. On the other hand, inhibition of other transcription factors, like NF-κB leads to the decrease in the pro-inflammatory gene expression. Moreover, the modulation of microRNAs activity may constitute a new mechanism responsible for valuable effects of triterpenoids. Recently, based on the structure of naturally occurring triterpenoids and with involvement of bioinformatics and computational chemistry, many synthetic analogs with improved biological properties have been obtained. Data from in vitro and in vivo experiments strongly suggest synthetic derivatives as promising candidates in the chemopreventive and chemotherapeutic strategies.
Keywords: Anti-oxidant response, Betulin, Chemoprevention, Oleanolic acid, Triterpenoids
CANCER CHEMOPREVENTION
The main goal of chemoprevention is to delay the development of cancer or even to revert the carcinogenesis process. The appropriate use of a chemopreventive agent depends on the understanding of its mechanism of action at different stages, including molecular, cellular, tissue and organ level. Although various compounds are nowadays tested both in vitro and in vivo, still there is a need for the development of new classes of chemopreventive agents as none of the existing ones is ideal. Triterpenoids have gained much attention as potential chemopreventive agents and their synthetic analogs characterized by lower toxicity, better availability, as well as increased protective effects are nowadays thoroughly examined, some of them are even in the phase I/II clinical trials for cancer treatment.
OLEANOLIC ACID AS AN EXAMPLE OF THERAPEUTIC TRITERPENOID
Terpenoids form a large and structurally diverse family of natural compounds, structure of which is based on five-carbon 2-methyl-1,3-butadiene units (called isoprene units). Dependently on the number of isoprene units forming structural skeleton of the molecule and on the form of connection of these units, this group is divided into several classes. One of them is represented by triterpenoids, among which the most important are pentacyclic compounds containing six five-carbon isoprene units. Triterpenoids are biosynthesized in plants by the cyclization of squalene, a chain triterpene hydrocarbon and precursor of all steroids (Phillips et al., 2006) and are widely present in various plants, mostly in wax-like coatings of fruits and medicinal herbs, including apples, cranberries, figs, olives, oregano, rosemary and thyme (Petronelli et al., 2009).
Triterpenoids have been used for medicinal purposes in many Asian countries for ages. During recent 20 years mechanisms responsible for their anti-inflammatory, cytoprotective, antitumorigenic and other effects have been discovered (Fig. 1)
Fig. 1. Schematic representation of a broad action of triterpenoids.
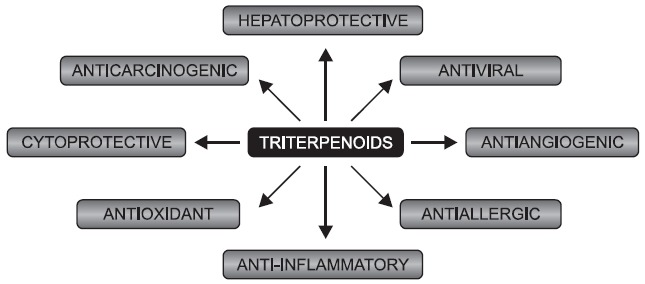
(reviewed in: Liby et al., 2007b). Moreover, a variety of additional biological activities including antiviral (anti-HIV) (Mengoni et al., 2002), antiallergic (Yuk et al., 2011) and spasmolytic activity (Begum et al., 2002) have been reported.
One of the best known and characterized triterpenoid is 3β-hydroxyolean-12-en-28-oic acid, named oleanolic acid (OA), produced by wide range of medicinal herbs and other plants, for example Olea europaea, Calendula officinalis, and Viscum album L. The anti-inflammatory effect of OA was first reported in 1969, when Gupta and coworkers showed its inhibitory effect in carrageenan-induced rat paw edema and formaldehyde-induced arthritis (Gupta et al., 1969). Later, in the late 80’s and early 90’s, these anti-inflammatory properties were additionally confirmed. OA was reported to inhibit dextran-induced rat paw edema and adjuvant-induced polyarthritis in mice and rats (Singh et al., 1992). It was noted that oral administration of OA was not as effective as intraperitoneal or subcutaneous injection in inhibiting inflammatory reactions (Takagi et al., 1980; Singh et al., 1992). Simultaneously, the experiments were aimed at elucidating the mechanism responsible for anti-inflammatory effect of triterpenoids. The inhibition of lipoxygenase and cyclooxygenase (COX-2) activity was firstly reported for other compound - ursolic acid (Najid et al., 1992) but then also for OA (Suh et al., 1998). Additionally, suppression of inducible nitric oxide synthase (iNOS) activity and the production of NO was demonstrated (Suh et al., 1998). Interestingly, anticancer activities of OA have been reported already more than 25 years ago. It was shown that topical application of OA to mice pretreated by 2-O-tetradecanoylphorbol-13-acetate (TPA) resulted in significantly lower number of papilloma-bearing animals (Tokuda et al., 1986). Later, OA was shown to be effective in treatment of other types of cancer. Three-week long oral treatment of rats with OA resulted in decreased incidence and multiplicity of azoxymethane-induced intestinal tumors (Yoshimi et al., 1992). Fujiwara et al. (2011) have recently discovered that OA inhibited the proliferation of human glioblastoma cells via inhibition of signal transducer and activator of transcription-3 (STAT3).
Apart from anti-tumorigenic and anti-inflammatory activity widely described in the literature, OA has also documented hepatoprotective properties. In animal models, it protects against variety of hepatotoxicants, for example carbon tetrachloride, cadmium and bromobenzene (references in: Liu et al., 2008). Moreover, hypoglycemic activity of OA was described. In alloxan-induced diabetic rats, OA caused amelioration of blood glucose elevation and increase of hepatic glycogen and insulin levels (Liu et al., 1994). Recently, the detailed molecular mechanism responsible for the hypoglycemic 2012.20.6.499activity of OA was reported (Zeng et al., 2012). In the model of streptozotocin-induced type II diabetes in mice, OA administration reversed the hyperglycemia by ~60%. OA was able to modulate the insulin signaling transduction, namely the phosphorylation of Akt and FoxO1 regulating gluconeogenesis was significantly increased in OA-treated mice (Zeng et al., 2012).
The complete list of OA activities is still increasing and probably will be even longer in the future. The results obtained so far indicate that OA interacts with important signaling pathways modulating cell physiology, which has a reflection in its anti-inflammatory and cytoprotective functions.
SYNTHETIC OLEANOLIC TRITERPENOIDS AND THEIR PROTECTIVE EFFECTS
Over the past years, a large number of triterpenoids have been synthetically modified in order to increase their bioactivity and protective effects. Based on structure of OA, several hundreds of derivatives called synthetic oleanane triterpenoids (SO) have been obtained. In many cases, it was reported that they possess better anti-inflammatory and cytoprotective properties than OA (reviewed in: Liby et al., 2007b). Three of the most potent known SO are 2-cyano-3,12-dioxooleana-1,9(11)-dien-28-oic acid (CDDO), its methyl ester (CDDO-Me) and its imidazolide (CDDO-Im).
As OA, above mentioned compounds when used at high concentrations, have ability to inhibit cell proliferation and induce apoptosis in ovarian (Gao et al., 2011), prostate (Deeb et al., 2011) and other cancer cells, including leukemia and myeloma cells (reviewed in: Liby et al., 2007b). Another therapeutic agent, CDDO-ethylamide (CDDO-Ea), was shown to extend survival in mouse model of pancreatic cancer (Liby et al., 2010) as well as in lung cancer (Liby et al., 2007a). Some of such derivatives are in clinical trials for treatment of leukemia and solid tumors (Petronelli et al., 2009; Speranza et al., 2012).
Another important mechanism of SO action is the inhibition of blood vessel formation, the critical process for tumor growth and metastasis. CDDO-Me inhibited angiogenesis in matrigel sponge assays in vivo and caused significant inhibition of endothelial cell growth in vitro (Vannini et al., 2007). The same compound was able to prevent estrogen receptor (ER)-negative mammary tumorigenesis in mice and the inhibition of angiogenesis was suggested to be at least partially responsible for such effect (Liby et al., 2008). Moreover, CDDO-Me blocked angiogenesis and the growth of Kaposi's sarcoma cells in nude mice (Vannini et al., 2007). Recent work by Mu et al. (2012) indicates that proangiogenic vascular endothelial growth factor (VEGF) signaling is affected by triterpenoids. The observed anticancer effect of pristimerin, a triterpenoid isolated from Celastrus and Maytenus spp. was mediated by the suppression of VEGF receptor 2 (VEGFR-2) kinase activity (Mu et al., 2012).
SO are multifunctional drugs reacting with cellular nucleophiles such as –SH groups of cysteines on multiple target proteins. From different possible interaction, the most important in the view of anti-cancer therapy is the regulation of STAT3, retinoid X receptor alpha (RXRα), epidermal growth factor receptor (EGFR), COX-2 and iNOS (reviewed in: Liby et al., 2007b). Moreover, SO may directly bind and inhibit IκB kinase enzyme complex (IKK) responsible for nuclear translocation of NF-κB leading to the silencing of NF-κB signaling (Ahmad et al., 2006; Yore et al., 2006). CDDO reduced NF-κB activity in human bronchial epithelial cells and decreased release of NF-κB dependent gene products like IL-8 (Nichols et al., 2009). In other studies, CDDO-Me was shown to inhibit NF-κB expression in prostate (Deeb et al., 2010), colorectal (Gao et al., 2010) as well as in ovarian (Gao et al., 2011) cancer cells. The growth-inhibitory and apoptosis-inducing activity of CDDO-Me relies on the inhibition of pro-survival kinase Akt what in turn leads to the inhibition of NF-κB and phospho-mammalian target of rapamycin (p-mTOR) signaling proteins (Gao et al., 2011).
The blockade of the tumor growth and induction of apoptosis are achieved when high concentrations of triterpenoids are used. On the other hand, at low concentrations, SO are rather cytoprotective and anti-inflammatory. The cytoprotective effects are mediated by the induction of the entire set of antioxidative proteins called phase 2 enzymes that protect cells against electrophiles and oxidants. Their upregulation provides a diverse and highly reliable response that enables cells to survive in stressful conditions and it is mediated by the modulation of the activity of nuclear factor erythroid 2-related factor 2 (Nrf2) (Li and Kong, 2009).
Nrf2 AND ITS TRANSCRIPTIONAL ACTIVATION
Nuclear factor erythroid 2-related factor 2 (Nrf2) is a transcription factor that regulates expression of many detoxification and antioxidant enzymes, molecular chaperones, stress response proteins, as well as proteasome subunits (Li and Kong, 2009). Nrf2 belongs to basic leucine zipper (bZIP) transcription factors featuring a cap “n” collar (CNC) structure defined by the presence of a conserved 43-amino acid CNC domain located N-terminally to the DNA binding domain (Sykiotis and Bohmann, 2010).
Like other transcription factors, Nrf2 signaling is regulated by the compartmental segregation (Fig. 2). Activated Nrf2 quickly translocates from the cytoplasm into the nucleus to regulate gene expression. Under normal conditions Nrf2 is anchored within the cytoplasm by Kelch-like-ECH-associated protein 1 (Keap1) (Li and Kong, 2009). When cells are exposed to stressing conditions like chemopreventive agents or oxidative stress, critical cysteine residues (-SH groups) in Keap1 are modified by redox modification. As a result, enzymatic activity of the Keap1-Cullin3-Rbx1 E3 ubiquitin ligase complex (known also as Cullin3-based E3 ligase ubiquitination complex) is inhibited which causes decreased Nrf2 ubiquitination and degradation. Thus, Keap1 functions as a kind of bridge between Nrf2 and above-mentioned ubiquitination complex preventing nuclear accumulation of Nrf2 under normal condition.
Fig. 2. Nrf2 transcriptional regulation. Under normal conditions, Nrf2 is sequestered in cytoplasm by Keap1. Triterpenoids via oxidative modification of –SH groups in Keap1 or via phosphorylation of Nrf2 by various protein kinase signaling pathways including MAPK, PI3K, PKC and CK-2 facilitate the liberation process of Nrf2 from Keap1 as well as translocation of Nrf2 into the nucleus. After binding Maf proteins and antioxidant response element (ARE) activation, the increased transcription of Nrf2-regulated genes (e.g. HO-1, GST, NQO-1) can be achieved.
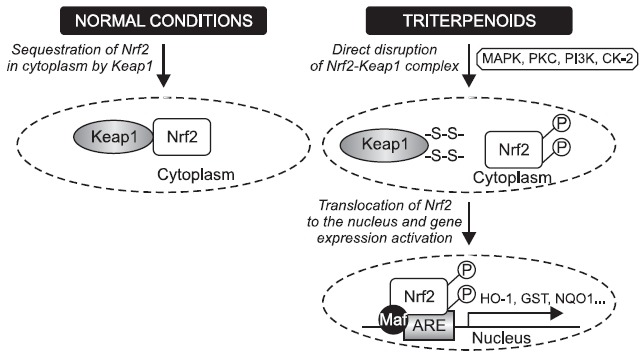
Moreover, the activation of Nrf2/ARE signaling may also rely on Nrf2 phosphorylation by multiple cellular kinase pathways: mitogen-activated protein kinases (MAPKs), protein kinase C (PKC), and phosphatidylinositol 3-kinase (PI3K) (reviewed in:Kwak and Kensler, 2010) . It was shown that phosphorylation of Nrf2 at Ser 40 facilitated its dissociation from Keap1 (Huang et al., 2002). Bloom and Jaiswal (2003) confirmed this result, however, they demonstrated that Nrf2 phosphorylation is not necessary for Nrf2 stabilization/accumulation in the nucleus. On the other hand, phosphorylation of Nrf2 by the casein kinase-2
(CK-2) was shown to play an important role not only in the activation and nuclear translocation of Nrf2 but also in the subsequent hyper-phosphorylation of Nrf2 (Pi et al., 2007).
After translocation into the nucleus, Nrf2 forms heterodimer with a group of nuclear bZIP proteins called small Maf proteins like MafF, G and K. Their heterodimerization with Nrf2 enhances the specificity to bind antioxidant response element (ARE) which represents a common cis-acting regulatory element in the 5’flanking region of promoters of many cytoprotective genes. Binding of Nrf2-Maf dimers to ARE subsequently initiates the transcription of target genes (Li and Kong, 2009). Proteins encoded by genes under control of ARE promoter can be classified into several categories: phase II xenobiotic-metabolizing enzymes, antioxidants and their modulating enzymes, molecular chaperones/proteasome systems, DNA repair enzymes and anti-inflammatory response proteins. Heme oxygenase-1 (HO-1), NAD(P)H quinone oxidoreductase 1 (NQO-1), glutamate cysteine ligase (Gcl) or glutathione-S-transferase (GST) are the examples of Nrf2 target genes. After recovery of cellular redox homeostasis, Keap1 moves into the nucleus and dissociates Nrf2 from ARE. Consequently, cytoprotective genes are no longer transcribed, whereas Nrf2-Keap1 complex is exported out of the nucleus. It associates in the cytoplasm with Cullin3-based E3 ligase ubiquitination complex resulting in degradation of Nrf2 and termination of the Nrf2/ARE signaling pathway (Li and Kong, 2009).
TRITERPENOIDS AS MODULATORS OF Nrf2/Keap1 PATHWAY
Both naturally occurring triterpenoids, like OA and its synthetic analogs (SO), affect Nrf2/ARE pathway (Yu and Kensler, 2005; Giudice et al., 2010). However, Nrf2 activation and subsequent regulation of the transcription of the plethora of protective genes may be increased thanks to the chemical modifications of the triterpenoid structure.
To show the important role of triterpenoids and their derivatives in the Nrf2/ARE signaling pathway regulation, multiple experiments using wild-type (Nrf2+/+) and knock-out (Nrf2−/−) mice/cells have been performed. CDDO-Im activated HO-1 in murine fibroblasts isolated from Nrf2 wild-type cells, but this activation was markedly lower in case of cells from Nrf2−/− mice even if higher concentrations of compound were applied (Liby et al., 2005). In another study, CDDO-Im, CDDO-trifluoroethylamide (CDDO-Tfea) and CDDO-Ea were used to examine SO ability to activate Nrf2 signaling in ARPE-19 retinal epithelial cells (Pitha-Rowe et al., 2009). All compounds activated Nrf2/ARE pathway in a dose-dependent manner, but CDDO-Im was the most potent. In vivo, oral treatment of mice by CDDO-Tfea increased neural retinal levels of the Nrf2-regulated transcripts in comparison to control animals. Also in other tissues, like liver, small intestine and cerebral cortex, these transcripts were induced (Pitha-Rowe et al., 2009).
The induction of Nrf2 and its downstream targets by triterpenoids was shown to be protective in different diseases. For example, intraperitoneal injection of OA increased protein expression and activity in livers of wild-type mice, but not in the Nrf2−/− mice and was able to protect against acetaminophen hepatotoxicity (Reisman et al., 2009). CDDO-Me treatment abrogated retinal capillary degeneration induced by ischemia-reperfusion injury in wild-type but not Nrf2−/− mice (Wei et al., 2011). Additionally, CDDO-Im increased in Nrf2 signaling in kidneys from Nrf2+/+ mice and protected them from cisplatin nephrotoxicity (Aleksunes et al., 2010). Interestingly, CDDO-Im was suggested to prevent high-fat diet-induced obesity and in fact it was able to decrease the body weight, adipose mass and hepatic lipid accumulation in Nrf2-dependent manner (Shin et al., 2009). Moreover, neuroprotective activities of OA derivatives dependent on Nrf2 have been demonstrated. In mouse model of amyotrophic lateral sclerosis (ALS), treatment with triterpenoids improved motor performance and slowed disease progression (Neymotin et al., 2011). Kaidery et al. (2012) investigated the effect of synthetic triterpenoids on Parkinson’s disease and showed that in a MPTP-induced preclinical mouse model of this disease the activation of Nrf2 by triterpenoids is very effective treatment strategy. Similarly, such compounds were shown to be promising in the treatment of the Huntington's disease. In the brain, CDDO-Ea and CDDO-Tfea upregulated Nrf2/ARE-induced genes, reduced oxidative stress, improved motor impairment and increased longevity (Stack et al., 2010).
Although all above mentioned SO are shown to possess protective effects, still new derivatives of OA are generated. Such derivatives are synthesized by the cyclization of squalene and thanks to sophisticated bioinformatics and structure–activity analysis, the specific carbonyl groups, suggested to be essential for the striking anti-inflammatory potency, are modified (Sporn et al., 2011). Here we investigated the effect of different synthetic analogs of OA, like 28-amino- and 28-hydroxyoleanane derivatives. In our studies we have found that especially three of them, namely ethyl oleanolate A-homo lactam (A-Lac-O-Me), erythrodiol (O-diol) and (succinyloleanolic)succinimide (Sc-O-ScIm) increase Nrf2 activity. After transfection of NIH3T3 murine fibroblasts with a construct containing the antioxidant response element sequence driving the expression of reporter gene, luciferase (ARE-luc plasmid), we were able to assess Nrf2 binding to ARE using chemiluminescent measurement (Fig. 3). After 24h treatment with both OA and its derivatives we found that all compounds increase Nrf2 activity comparable to prostaglandin J2 (PGJ2), a known Nrf2 inducer (Fig. 4A). Additionally, Nrf2 protein was nicely activated by synthetic analog of OA as shown by immunostaining (Fig. 4B). The induction of HO-1 mRNA, the Nrf2 target gene, after 6h stimulation with the compounds was greater than after PGJ2 (Fig. 4C). All those results suggest that OA derivatives have the ability to induce Nrf2/ARE pathway. However, we are still looking for better activators of antioxidant response.
Fig. 3. Schematic representation of the procedure of Nrf2 activity assessment. 24h after transfection with a reporter construct containing the antioxidant response element (ARE) sequence driving the expression of reporter gene, luciferase (ARE-luc plasmid), cells are stimulated by Nrf2 inducers (e.g. triterpenoids). Nrf2 translocates to nucleus and binds to ARE sequence what leads to increased chemiluminescence.
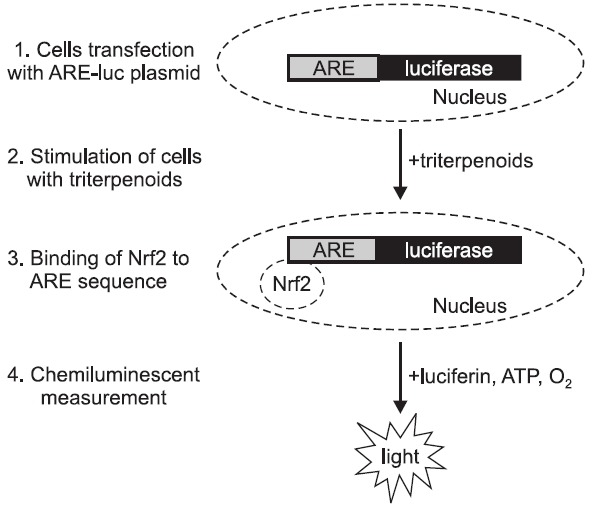
Fig. 4. OA derivatives increase Nrf2 activity and its target gene expression. 24h after transfection with a reporter construct regulated by the antioxidant response element (ARE-luc), NIH3T3 cells were stimulated by PGJ2 (positive control), OA and its derivatives (A-Lac-O-Me, O-diol, Sc-O-ScIm). After next 24h luciferase was measured showing the increased activity of Nrf2 after stimulation by all compounds (A). Moreover, the effect of selected compound (A-Lac-O-Me) on Nrf2 protein level was checked using immunostaining analysis (B). 6h treatment with the OA derivatives led to the increased expression of HO-1 (gene regulated by Nrf2) as shown by real-time PCR (C). Nrf2 activation by triterpenoids might be cell-type dependent. After transfection of human endothelial cells (HMEC-1) and human keratinocytes (HaCaT) with ARE-luc plasmid and stimulation with three different concentrations of betulin, the increase in luciferase activity was observed only in HaCaT cells (D). (A, C, D) - mean of 3 independent experiments, *p<0.05, B - representative immunostaining for Nrf2 (green). DAPI staining was used to visualize nuclei (blue).
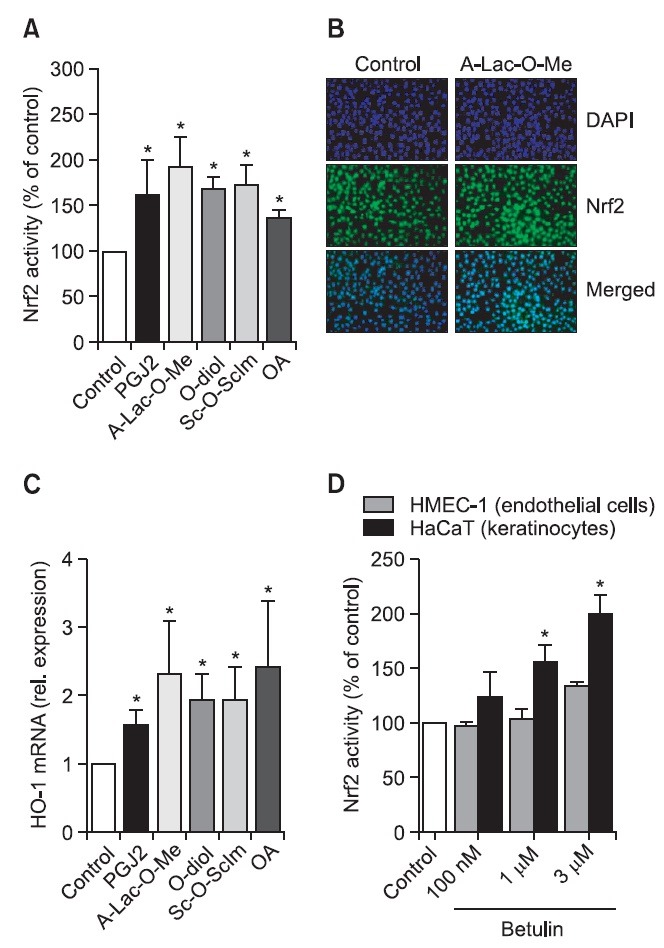
Importantly, the effect of triterpenoids on Nrf2 activation might be cell-type dependent. We compared the effect of other triterpenoid, betulin on Nrf2 activation in keratinocytes and endothelial cells. We observed greater response of human keratinocytes (HaCaT) than human microvascular endothelial cells (HMEC-1) when the effect of betulin on the Nrf2 binding to ARE sequence was tested in the transfection assay (Fig. 4D). Previously, it was shown that in retinal epithelial cells even 10-fold activation was obtained 24h after stimulation with OA derivatives (Pitha-Rowe et al., 2009). However, when human endothelial cells were treated with diterpene lactone called andrographlide, only 2-3 times upregulation of Nrf2 activity was observed (Yu et al., 2010) similarly to our results. It again suggests that OA derivatives have ability to increase Nrf2 activity in various cell types, but the power of this induction is not only dependent on compound tested, but also on cell type.
The protective response of Nrf2 induction relies on the effects exerted by the plethora of genes regulated in Nrf2 dependent manner. One of the well characterized gene upregulated after Nrf2 induction is HO-1, which catalyzes conversion of heme into biologically active products like carbon monoxide (CO), biliverdin and free iron. Biliverdin is quickly reduced to bilirubin by biliverdin reductase, whereas iron is sequestered by ferritin. Bilirubin inhibits NADPH oxidase and is a potent scavenger of free radicals. Decrease of heme content also reduces cellular reactive oxygen species (ROS) generation and CO has antiproliferative and anti-inflammatory properties. Thus, end-products of HO-1 activity demonstrate cytoprotective effects – they attenuate inflammatory response and lower
rate of apoptosis (reviewed in: Loboda et al., 2008). However, it has to be remembered that HO-1 increases neovascularization and growing body of evidence suggests that it may play a role in tumor induction and accelerate the growth and spread of tumors (reviewed in: Was et al., 2010).
MAY TRITERPENOIDS REGULATE miRNAs EXPRESSION?
Emerging evidence suggest that microRNAs (miRNAs), small non-coding RNAs regulating gene expression by multiple
mechanisms, play important roles in the development and progression of cancer and may be even used as a specific cancer biomarkers (reviewed in: Ciesla et al., 2011). Therefore, targeting specific miRNAs could be a novel therapeutic approach for the treatment of cancers and may be the mechanism of action of chemopreventive agents. In fact, it has been reported that miRNAs could be regulated by different natural chemopreventive compounds, like curcumin, resveratrol or isoflavones, leading to the inhibition of cancer cell growth and metastasis (reviewed in: Li et al., 2010; Karius et al., 2012). However, still little is known about the potency of triterpenoids on miRNAs expression, although such regulation is very probable.
Chintharlapalli et al. (2009) showed that methyl 2-cyano-3,11-dioxo-18β-olean-1,12-dien-30-oate (CDODA-Me), a synthetic derivative of glycyrrhetinic acid, a triterpenoid phytochemical found in licorice extracts, decreased expression of miR-27a leading to increased expression of genes responsible for inhibition of the progression of colon cancer cells through G2/M. The downregulation of miR-27a was also observed after stimulation of pancreatic cancer cells with CDDO or CDDO-Me (Jutooru et al., 2010) as well as after betulinic acid treatment of colon cancer cells (Chintharlapalli et al., 2011) or breast cancer cells (Liu et al., 2012). In the mentioned publications, different cell lines have been used and various downstream mechanisms activated by miR-27a inhibition were shown to be responsible for the anticancer activity of triterpenoids, although they were not correlated to Nrf2 activity. miR-27a is oncogenic and its expression is upregulated in different cancers, so it may be a general target of both natural and synthetic triterpenoids. However, there is no data on the effect of triterpenoids on other miRNAs.
A lot of questions still remain to be answered. Is it possible that the synthetic analog which acts more potently than its original compound regulates the specific miRNAs? Is the expression of miRNAs, which binding sites are present within 3’UTR sequence of Nrf2 (e.g. miR-17, miR-129-5p and miR-132, based on miRanda analysis), regulated by triterpenoids? It is reasonable to believe that not only miR-27a can be a target of triterpenoids action. Nevertheless, experiments proving such regulation have to be performed.
CONCLUSIONS
Triterpenoids are multitarget compounds exhibiting concentration-dependent functions: at lower doses they act as an anti-inflammatory and cytoprotective compounds whereas at higher concentrations they possess mostly anticancer activities. The induction of Nrf2 by natural and synthetic triterpenoids has become commonly accepted mechanism responsible for cytoprotection and chemoprevention. Still many questions remain unanswered and the possible involvement of miRNAs in triterpenoids-mediated effects is an intriguing aspect.
Acknowledgments
This work was supported by the grants No N N301 314837 and IP2011 031071 from the National Science Center and from the Polish Ministry of Science and Higher Education, respectively. AL is supported by the Foundation for Polish Science - PARENT-BRIDGE Programme co-financed by the European Union within European Regional Development Fund (POMOST/2010-2/8). The Faculty of Biochemistry, Biophysics and Biotechnology of the Jagiellonian University is a beneficiary of the structural funds from the European Union and the Polish Ministry of Science and Higher Education (grants No: POIG.02.01.00-12-064/08 02.02.00-00-014/08, 01.01.02-00-069/09 and 01.01.02-00-109/09). Dr. Jacek Achrem-Achremowicz is acknowledged for betulin delivery.
References
- 1.Ahmad R., Raina D., Meyer C., Kharbanda S., Kufe D. Triterpenoid CDDO-Me blocks the NF-kappaB pathway by direct inhibition of IKKbeta on Cys-179. J. Biol. Chem. (2006);281:35764–35769. doi: 10.1074/jbc.M607160200. [DOI] [PubMed] [Google Scholar]
- 2.Aleksunes L. M., Goedken M. J., Rockwell C. E., Thomale J., Manautou J. E., Klaassen C. D. Transcriptional regulation of renal cytoprotective genes by Nrf2 and its potential use as a therapeutic target to mitigate cisplatin-induced nephrotoxicity. J. Pharmacol. Exp. Ther. (2010);335:2–12. doi: 10.1124/jpet.110.170084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Begum S., Sultana I., Siddiqui B. S., Shaheen F., Gilani A. H. Structure and spasmolytic activity of eucalyptanoic acid from Eucalyptus camaldulensis var. obtusa and synthesis of its active derivative from oleanolic acid. J. Nat. Prod. (2002);65:1939–1941. doi: 10.1021/np020127x. [DOI] [PubMed] [Google Scholar]
- 4.Bloom D. A., Jaiswal A. K. Phosphorylation of Nrf2 at Ser40 by protein kinase C in response to antioxidants leads to the release of Nrf2 from INrf2, but is not required for Nrf2 stabilization/accumulation in the nucleus and transcriptional activation of antioxidant response element-mediated NAD(P)H:quinone oxidoreductase-1 gene expression. J. Biol. Chem. (2003);278:44675–44682. doi: 10.1074/jbc.M307633200. [DOI] [PubMed] [Google Scholar]
- 5.Chintharlapalli S., Papineni S., Abdelrahim M., Abudayyeh A., Jutooru I., Chadalapaka G., Wu F., Mertens-Talcott S., Vanderlaag K., Cho S. D., Smith R. 3rd, Safe S. Oncogenic microRNA-27a is a target for anticancer agent methyl 2-cyano-3,11-dioxo-18beta-olean-1,12-dien-30-oate in colon cancer cells. Int. J. Cancer. . (2009);125:1965–1974. doi: 10.1002/ijc.24530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Chintharlapalli S., Papineni S., Lei P., Pathi S., Safe S. Betulinic acid inhibits colon cancer cell and tumor growth and induces proteasome-dependent and -independent downregulation of specificity proteins (Sp) transcription factors. BMC Cancer. (2011);11:371. doi: 10.1186/1471-2407-11-371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Ciesla M., Skrzypek K., Kozakowska M., Loboda A., Dulak J. MicroRNAs as biomarkers of disease onset. Anal. Bioanal. Chem. (2011);401:2051–2061. doi: 10.1007/s00216-011-5001-8. [DOI] [PubMed] [Google Scholar]
- 8.Deeb D., Gao X., Jiang H., Janic B., Arbab A. S., Rojanasakul Y., Dulchavsky S. A., Gautam S. C. Oleanane triterpenoid CDDO-Me inhibits growth and induces apoptosis in prostate cancer cells through a ROS-dependent mechanism. Biochem. Pharmacol. (2010);79:350–360. doi: 10.1016/j.bcp.2009.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Deeb D., Gao X, Liu Y., Jiang D., Divine G. W., Arbab A. S., Dulchavsky S. A., Gautam S. C. Synthetic triterpenoid CDDO prevents the progression and metastasis of prostate cancer in TRAMP mice by inhibiting survival signaling. Carcinogenesis. (2011);32:757–764. doi: 10.1093/carcin/bgr030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Fujiwara Y., Komohara Y., Kudo R., Tsurushima K., Ohnishi K., Ikeda T., Takeya M. Oleanolic acid inhibits macrophage differentiation into the M2 phenotype and glioblastoma cell proliferation by suppressing the activation of STAT3. Oncol. Rep. (2011);26:1533–1537. doi: 10.3892/or.2011.1454. [DOI] [PubMed] [Google Scholar]
- 11.Gao X., Deeb D., Hao J., Liu Y., Arbab A. S., Dulchavsky S. A., Gautam S. C. Synthetic triterpenoids inhibit growth, induce apoptosis and suppress pro-survival Akt, mTOR and NF-{kappa}B signaling proteins in colorectal cancer cells. Anticancer Res. (2010);30:785–792. [PMC free article] [PubMed] [Google Scholar]
- 12.Gao X., Liu Y., Deeb D., Arbab A. S., Guo A. M., Dulchavsky S. A., Gautam S. C. Synthetic oleanane triterpenoid, CDDO-Me, induces apoptosis in ovarian cancer cells by inhibiting prosurvival AKT/NF-kappaB/mTOR signaling. Anticancer Res. (2011);31:3673–3681. [PMC free article] [PubMed] [Google Scholar]
- 13.Giudice A., Arra C., Turco M. C. Review of molecular mechanisms involved in the activation of the Nrf2-ARE signaling pathway by chemopreventive agents. Methods Mol. Biol. (2010);647:37–74. doi: 10.1007/978-1-60761-738-9_3. [DOI] [PubMed] [Google Scholar]
- 14.Gupta M. B., Bhalla T. N., Gupta G. P., Mitra C. R., Bhargava K. P. Anti-inflammatory activity of natural products. I. Triterpenoids. Eur. J. Pharmacol. (1969);6:67–70. doi: 10.1016/0014-2999(69)90067-3. [DOI] [PubMed] [Google Scholar]
- 15.Huang H. C., Nguyen T., Pickett C. B. Phosphorylation of Nrf2 at Ser-40 by protein kinase C regulates antioxidant response element-mediated transcription. J. Biol. Chem. (2002);277:42769–42774. doi: 10.1074/jbc.M206911200. [DOI] [PubMed] [Google Scholar]
- 16.Jutooru I., Chadalapaka G., Abdelrahim M., Basha M. R., Samudio I., Konopleva M., Andreeff M., Safe S. Methyl 2-cyano-3,12-dioxooleana-1,9-dien-28-oate decreases specificity protein transcription factors and inhibits pancreatic tumor growth: role of microRNA-27a. Mol. Pharmacol. (2010);78:226–236. doi: 10.1124/mol.110.064451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kaidery N. A., Banerjee, R., Yang L., Smirnova N. A., Hushpulian D. M., Liby K. T., Williams C. R., Yamamoto M., Kensler T. W., Ratan R. R., Sporn M. B., Beal M. F., Gazaryan I. G., Thomas B. Targeting Nrf2-Mediated Gene Transcription by Extremely Potent Synthetic Triterpenoids Attenuate Dopaminergic Neurotoxicity in the MPTP Mouse Model of Parkinson's Disease. Antioxid. Redox Signal. (2012) doi: 10.1089/ars.2011.4491. Epub ahead of print. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Karius T., Schnekenburger M., Dicato M., Diederich M. MicroRNAs in cancer management and their modulation by dietary agents. Biochem. Pharmacol. (2012);83:1591–1601. doi: 10.1016/j.bcp.2012.02.004. [DOI] [PubMed] [Google Scholar]
- 19.Kwak M. K., Kensler T. W. Targeting NRF2 signaling for cancer chemoprevention. Toxicol. Appl. Pharmacol. (2010);244:66–76. doi: 10.1016/j.taap.2009.08.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Li W., Kong A. N. Molecular mechanisms of Nrf2-mediated antioxidant response. Mol. Carcinog. . (2009);48:91–104. doi: 10.1002/mc.20465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Li Y., Kong D., Wang Z., Sarkar F. H. Regulation of microRNAs by natural agents: an emerging field in chemoprevention and chemotherapy research. Pharm. Res. (2010);27:1027–1041. doi: 10.1007/s11095-010-0105-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Liby K., Hock T., Yore M. M., Suh N., Place A. E., Risingsong R., Williams C. R., Royce D. B., Honda T., Honda Y., Gribble G. W., Hill-Kapturczak N., Agarwal A., Sporn M. B. The synthetic triterpenoids, CDDO and CDDO-imidazolide, are potent inducers of heme oxygenase-1 and Nrf2/ARE signaling. Cancer Res. (2005);65:4789–4798. doi: 10.1158/0008-5472.CAN-04-4539. [DOI] [PubMed] [Google Scholar]
- 23.Liby K., Risingsong R., Royce D. B., Williams C. R., Yore M. M., Honda T., Gribble G. W., Lamph W. W., Vannini N., Sogno I., Albini A., Sporn M. B. Prevention and treatment of experimental estrogen receptor-negative mammary carcinogenesis by the synthetic triterpenoid CDDO-methyl Ester and the rexinoid LG100268. Clin. Cancer Res. (2008);14:4556–4563. doi: 10.1158/1078-0432.CCR-08-0040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Liby K., Royce D. B., Williams C. R., Risingsong R., Yore M. M., Honda T., Gribble G. W., Dmitrovsky E., Sporn T. A., Sporn M. B. The synthetic triterpenoids CDDO-methyl ester and CDDO-ethyl amide prevent lung cancer induced by vinyl carbamate in A/J mice. Cancer Res. (2007a);67:2414–2419. doi: 10.1158/0008-5472.CAN-06-4534. [DOI] [PubMed] [Google Scholar]
- 25.Liby K. T., Royce D. B., Risingsong R., Williams C. R., Maitra A., Hruban R. H., Sporn M. B. Synthetic triterpenoids prolong survival in a transgenic mouse model of pancreatic cancer. Cancer Prev. Res. (Phila) (2010);3:1427–1434. doi: 10.1158/1940-6207.CAPR-10-0197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Liby K. T., Yore M. M., Sporn M. B. Triterpenoids and rexinoids as multifunctional agents for the prevention and treatment of cancer. Nat. Rev. Cancer. (2007b);7:357–369. doi: 10.1038/nrc2129. [DOI] [PubMed] [Google Scholar]
- 27.Liu J., Mao Q., Klaassen C. D. The effects of 10 triterpenoid compounds on experimental liver injury in mice. Fundam. Appl. Toxicol. (1994);22:34–40. doi: 10.1006/faat.1994.1005. [DOI] [PubMed] [Google Scholar]
- 28.Liu J., Wu Q., Lu Y. F., Pi J. New insights into generalized hepatoprotective effects of oleanolic acid: key roles of metallothionein and Nrf2 induction. Biochem. Pharmacol. (2008);76:922–928. doi: 10.1016/j.bcp.2008.07.021. [DOI] [PubMed] [Google Scholar]
- 29.Liu X., Jutooru I., Lei P., Kim K., Lee S. O., Brents L. K., Prather P. L., Safe S. Betulinic acid targets YY1 and ErbB2 through cannabinoid receptor-dependent disruption of microRNA-27a:ZBTB10 in breast cancer. Mol. Cancer Ther. (2012);11:1421–1431. doi: 10.1158/1535-7163.MCT-12-0026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Loboda A., Jazwa A., Grochot-Przeczek A., Rutkowski A. J., Cisowski J., Agarwal A., Jozkowicz A., Dulak J. Heme oxygenase-1 and the vascular bed: from molecular mechanisms to therapeutic opportunities. Antioxid. Redox Signal. (2008);10:1767–1812. doi: 10.1089/ars.2008.2043. [DOI] [PubMed] [Google Scholar]
- 31.Mengoni F., Lichtner M., Battinelli L., Marzi M., Mastroianni C. M., Vullo V., Mazzanti G. In vitro anti-HIV activity of oleanolic acid on infected human mononuclear cells. Planta Med. (2002);68:111–114. doi: 10.1055/s-2002-20256. [DOI] [PubMed] [Google Scholar]
- 32.Mu X., Shi W., Sun L., Li H., Jiang Z., Zhang L. Pristimerin, a triterpenoid, inhibits tumor angiogenesis by targeting VEGFR2 activation. Molecules. (2012);17:6854–6868. doi: 10.3390/molecules17066854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Najid A., Simon A., Cook J., Chable-Rabinovitch H., Delage C., Chulia A. J., Rigaud M. Characterization of ursolic acid as a lipoxygenase and cyclooxygenase inhibitor using macrophages, platelets and differentiated HL60 leukemic cells. FEBS Lett. (1992);299:213–217. doi: 10.1016/0014-5793(92)80117-y. [DOI] [PubMed] [Google Scholar]
- 34.Neymotin A., Calingasan N. Y., Wille E., Naseri N., Petri S., Damiano M., Liby K. T., Risingsong R., Sporn M., Beal M. F., Kiaei M. Neuroprotective effect of Nrf2/ARE activators, CDDO ethylamide and CDDO trifluoroethylamide, in a mouse model of amyotrophic lateral sclerosis. Free Radic. Biol. Med. (2011);51:88–96. doi: 10.1016/j.freeradbiomed.2011.03.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Nichols D. P., Ziady A. G., Shank S. L., Eastman J. F., Davis P. B. The triterpenoid CDDO limits inflammation in preclinical models of cystic fibrosis lung disease. J. Physiol. Lung Cell Mol. Physiol. (2009);297:L828–836. doi: 10.1152/ajplung.00171.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Petronelli A., Pannitteri G., Testa U. Triterpenoids as new promising anticancer drugs. Anticancer Drugs . (2009);20:880–892. doi: 10.1097/CAD.0b013e328330fd90. [DOI] [PubMed] [Google Scholar]
- 37.Phillips D. R., Rasbery J. M., Bartel B., Matsuda S. P. Biosynthetic diversity in plant triterpene cyclization. Curr. Opin. Plant Biol. (2006);9:305–314. doi: 10.1016/j.pbi.2006.03.004. [DOI] [PubMed] [Google Scholar]
- 38.Pi J., Bai Y., Reece J. M., Williams J., Liu D., Freeman M. L., Fahl W. E., Shugar D., Liu J., Qu W., Collins S., Waalkes M. P. Molecular mechanism of human Nrf2 activation and degradation: role of sequential phosphorylation by protein kinase CK2. Free Radic. Biol. Med. (2007);42:1797–1806. doi: 10.1016/j.freeradbiomed.2007.03.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Pitha-Rowe I., Liby K., Royce D., Sporn M. Synthetic triterpenoids attenuate cytotoxic retinal injury: cross-talk between Nrf2 and PI3K/AKT signaling through inhibition of the lipid phosphatase PTEN. Invest. Ophthalmol. Vis. Sci. (2009);50:5339–5347. doi: 10.1167/iovs.09-3648. [DOI] [PubMed] [Google Scholar]
- 40.Reisman S. A., Aleksunes L. M., Klaassen C. D. Oleanolic acid activates Nrf2 and protects from acetaminophen hepatotoxicity via Nrf2-dependent and Nrf2-independent processes. Biochem. Pharmacol. . (2009);77(1282):1273–1282. doi: 10.1016/j.bcp.2008.12.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Shin S., Wakabayashi J., Yates M. S., Wakabayashi N., Dolan P. M., Aja S., Liby K. T., Sporn M. B., Yamamoto M., Kensler T. W. Role of Nrf2 in prevention of high-fat diet-induced obesity by synthetic triterpenoid CDDO-imidazolide. Eur. J. Pharmacol. (2009);620:138–144. doi: 10.1016/j.ejphar.2009.08.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Singh G. B., Singh S., Bani S., Gupta B. D., Banerjee S. K. Anti-inflammatory activity of oleanolic acid in rats and mice. J. Pharm. Pharmacol. (1992);44:456–458. doi: 10.1111/j.2042-7158.1992.tb03646.x. [DOI] [PubMed] [Google Scholar]
- 43.Speranza G., Gutierrez M. E., Kummar S., Strong J. M., Parker R. J., Collins J., Yu Y., Cao L., Murgo A. J., Doroshow J. H., Chen A. Phase I study of the synthetic triterpenoid, 2-cyano-3, 12-dioxoolean-1, 9-dien-28-oic acid (CDDO), in advanced solid tumors. Cancer Chemother. Pharmacol. (2012);69:431–438. doi: 10.1007/s00280-011-1712-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Sporn M. B., Liby K. T., Yore M. M., Fu L., Lopchuk J. M., Gribble G. W. New synthetic triterpenoids: potent agents for prevention and treatment of tissue injury caused by inflammatory and oxidative stress. J. Nat. Prod. (2011);74:537–545. doi: 10.1021/np100826q. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Stack C., Ho D., Wille E., Calingasan N. Y., Williams C., Liby K., Sporn M., Dumont M., Beal M. F. Triterpenoids CDDO-ethyl amide and CDDO-trifluoroethyl amide improve the behavioral phenotype and brain pathology in a transgenic mouse model of Huntington's disease. Free Radic. Biol. Med. (2010);49:147–158. doi: 10.1016/j.freeradbiomed.2010.03.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Suh N., Honda T., Finlay H. J., Barchowsky A., Williams C., Benoit N. E., Xie Q. W., Nathan C., Gribble G. W., Sporn M. B. Novel triterpenoids suppress inducible nitric oxide synthase (iNOS) and inducible cyclooxygenase (COX-2) in mouse macrophages. Cancer Res. . (1998);58:717–723. [PubMed] [Google Scholar]
- 47.Sykiotis G. P., Bohmann D. Stress-activated cap' n' collar transcription factors in aging and human disease. Sci. Signal. (2010);3:re3. doi: 10.1126/scisignal.3112re3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Takagi K., Park E. H., Kato H. Anti-inflammatory activities of hederagenin and crude saponin isolated from Sapindus mukorossi Gaertn. Chem. Pharm. Bull. (Tokyo) . (1980);28:1183–1188. doi: 10.1248/cpb.28.1183. [DOI] [PubMed] [Google Scholar]
- 49.Tokuda H., Ohigashi H., Koshimizu K., Ito Y. Inhibitory effects of ursolic and oleanolic acid on skin tumor promotion by 12-O-tetradecanoylphorbol-13-acetate. Cancer Lett. . (1986);33:279–285. doi: 10.1016/0304-3835(86)90067-4. [DOI] [PubMed] [Google Scholar]
- 50.Vannini N., Lorusso G., Cammarota R., Barberis M., Noonan D. M., Sporn M. B., Albini A. The synthetic oleanane triterpenoid, CDDO-methyl ester, is a potent antiangiogenic agent. Mol. Cancer Ther. (2007);6:3139–3146. doi: 10.1158/1535-7163.MCT-07-0451. [DOI] [PubMed] [Google Scholar]
- 51.Was H., Dulak J., Jozkowicz A. Heme oxygenase-1 in tumor biology and therapy. Curr. Drug Targets. (2010);11:1551–1570. doi: 10.2174/1389450111009011551. [DOI] [PubMed] [Google Scholar]
- 52.Wei Y., Gong J., Yoshida T., Eberhart C. G., Xu Z., Kombairaju P., Sporn M. B., Handa J. T., Duh E. J. Nrf2 has a protective role against neuronal and capillary degeneration in retinal ischemia-reperfusion injury. Free Radic. Biol. Med. (2011);51:216–224. doi: 10.1016/j.freeradbiomed.2011.04.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Yore M. M., Liby K. T., Honda T., Gribble G. W., Sporn M. B. The synthetic triterpenoid 1-[2-cyano-3,12-dioxooleana-1,9(11)-dien-28-oyl]imidazole blocks nuclear factor-kappaB activation through direct inhibition of IkappaB kinase beta. Mol. Cancer. Ther. (2006);5:3232–3239. doi: 10.1158/1535-7163.MCT-06-0444. [DOI] [PubMed] [Google Scholar]
- 54.Yoshimi N., Wang A., Morishita Y., Tanaka T., Sugie S., Kawai K., Yamahara J., Mori H. Modifying effects of fungal and herb metabolites on azoxymethane-induced intestinal carcinogenesis in rats. Jpn. J. Cancer Res. (1992);83:1273–1278. doi: 10.1111/j.1349-7006.1992.tb02758.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Yu A. L., Lu C. Y., Wang T. S., Tsai C. W., Liu K. L., Cheng Y. P., Chang H. C., Lii C. K., Chen H. W. Induction of heme oxygenase 1 and inhibition of tumor necrosis factor alpha-induced intercellular adhesion molecule expression by andrographolide in EA.hy926 cells. J. Agric. Food Chem. . (2010);58:7641–7648. doi: 10.1021/jf101353c. [DOI] [PubMed] [Google Scholar]
- 56.Yu X., Kensler T. Nrf2 as a target for cancer chemoprevention. Mutat. Res. (2005);591:93–102. doi: 10.1016/j.mrfmmm.2005.04.017. [DOI] [PubMed] [Google Scholar]
- 57.Yuk J. E., Lee M. Y., Kwon O. K., Cai X. F., Jang H. Y., Oh S. R., Lee H. K., Ahn K. S. Effects of astilbic acid on airway hyperresponsiveness and inflammation in a mouse model of allergic asthma. Int. Immunopharmacol. (2011);11:266–273. doi: 10.1016/j.intimp.2010.12.002. [DOI] [PubMed] [Google Scholar]
- 58.Zeng X. Y., Wang Y. P., Cantley J., Iseli T. J., Molero J. C., Hegarty B. D., Kraegen E. W., Ye Y., Ye J. M. Oleanolic acid reduces hyperglycemia beyond treatment period with Akt/FoxO1-induced suppression of hepatic gluconeogenesis in type-2 diabetic mice. PLoS One . (2012);7:e42115. doi: 10.1371/journal.pone.0042115. [DOI] [PMC free article] [PubMed] [Google Scholar]