

Abstract
Systemic inflammation has been found in association with vascular endothelial function for clinical implications including exercise-induced pathology. However, information on the relationship between the exercise-related inflammatory responses and endothelial function is limited. This study aimed to investigate the effects of prolonged endurance exercise on the expression of selected soluble adhesion molecules and inflammatory markers. Twenty-four middle-aged males participating in a 308 km ultra-marathon were recruited in this study. Venous blood was collected at baseline, 100 km, 200 km, and 308 km for the analysis of sVCAM-1, sE-selectin, leukocytes, hs-CRP, CK, and TNF-α. Significant increases of sVCAM-1, sE-selectin, and leukocytes were observed at 100 km. sVCAM-1 had the greatest significant increase at 100 km. In addition, sVCAM-1 was significantly associated with the running speed and leukocytes. sE-selectin was significantly associated with leukocytes, hs-CRP, TNF-α, and CK. Delayed rises in hs-CRP and CK were observed at 200 km. TNF-α fluctuated throughout the race with a significant increase at 308 km. Delayed onset of hs-CRP and continuously increased sE-selectin suggest anti-inflammatory responses to suppress pro-inflammatory markers such as TNF-α. Prolonged repetition of muscle contraction may have released delayed CK and significant rise in TNF-α toward the end of the race. The present study demonstrated an activation of the surrogate markers of endothelial dysfunction in relationship to exercise intensity and leukocyte trafficking without a significant activation of the inflammatory responses. Thus, alteration of the endothelium may be related to increased blood flow and shear stress put upon the endothelium in response to increased oxygen demand on the heart.
Key points
Systemic inflammation is associated with vascular endothelial function for clinical implications including exercise-induced pathology.
Inflammatory process involves a cascade of inflammatory and endothelial markers.
A prolonged endurance ultra-marathon induced significant increases in systemic inflammation and vascular endothelial markers at different checkpoints.
sVCAM-1, a surrogate marker of endothelial dysfunction, significantly increased in response to increased exercise intensity and leukocyte trafficking without significant changes in the inflammatory markers.
Prolonged repetition of muscle contraction may have delayed released of pro-inflammatory markers, CK, hs-CRP, and TNF-α.
Key words: Endothelial function, inflammation, leukocytes, vascular adhesion molecules, ultra-marathon
Introduction
Physical activity is known for various beneficial effects on human body. Regularly performed moderate physical activity is known to reduce cardiovascular risk factors (Mora et al., 2007) and improve blood and lipid profiles (Dunn et al., 1997). However, studies on extreme physical activity involving long duration and heavy exertion have shown to increase the risk for cardiovascular events, (Maron et al., 1996; Siegel, 1997), myocardial infarction, sudden death, (Burke et al., 1999) and abnormal ranges of cardiac markers (Fortescue et al., 2007; La Gerche et al., 2004; Scott et al., 2009).
However, the negative aspects of excessive exercise on the cardiovascular system are not well understood. It is especially true for the exercise-related vascular endothelial dysfunction. Recent studies have started to show the crucial role of the endothelium in cardiovascular health. The recent observational studies closely associate the involvement of vascular endothelial and inflammatory factors to cardiovascular complications (Tousoulis et al., 2008; van Bussel et al., 2011). Inflammatory mediators are known to play an essential role in the pathogenesis of atherosclerosis and development of atheroma (Blake and Ridker, 2001; Bolad and Delafontaine, 2005). Abnormal endothelial function has been observed in various cardiovascular related pathogenesis such as congestive heart failure, coronary heart disease, and hypertension (Balciunas et al., 2009; Bolad and Delafontaine, 2005; Goel et al., 2007; van Bussel et al., 2011). In support of such reports, improvements in the endothelial function and inflammation were associated with reduction in cardiovascular events (Tousoulis et al., 2008).
Recently published reports have been associating physical activity with the activation of endothelium and inflammation (Marsh and Coombes, 2005; Suzuki et al., 2003). Moderately performed exercise is known to improve endothelium function and inflammation responses (Di Francescomarino et al., 2009; Petersen and Pedersen, 2005). However, previous reports showed that exhaustive or vigorous exercise increased circulating inflammatory markers and altered the state of endothelium for further induction into the pathogenic state (Bartzeliotou et al., 2007; Bernecker et al., 2011; Goel et al., 2007; Scherr et al., 2011). In addition, chronic inflammation and endothelial dysfunction may lead to impaired exercise capacity (Bartzeliotou et al., 2007; Marsh and Coombes, 2005; Stewart et al., 2004).
Inflammatory processes involve several markers of inflammatory cascade (Bartzeliotou et al. , 2007; Blake and Ridker, 2001; Suzuki et al., 2003). In the early phase of the endothelial damage, leukocytes migrate to the injury site for adhesion and transendothelial migration into the tissue. Selectin family of adhesion molecules such as E-, L- and P-selectin move to the endothelial surface for tethering and rolling of leukocytes on to the vascular wall (Balciunas et al., 2009; Marsh and Coombes, 2005). Immunoglobulin-like molecules, sVCAM-1 and ICAM-1, are involved in firm adherence of leukocytes onto the surface of endothelium to further promote inflammatory responses (Jaakkola et al., 2000; Porreca et al., 2002).
Pro-inflammatory markers such as TNF-α, IL-6 and hs-CRP are known to be involved in the classical symptoms of inflammation (Bernecker et al., 2011). Vigorous exercise is also known to substantially increase the pro-inflammatory markers (Bernecker et al., 2011; Ostrowski et al., 1999; Scherr et al., 2011). However, although the association between the endothelial and inflammatory markers has been suggested in previous studies (Marsh and Coombes, 2005; Pedersen et al., 2007; Smith, 2000), the relationship is still in debate (Bernecker et al., 2011).
Most of the exercise-related studies observed the inflammatory and endothelial markers before and after the exercise performance for pre-post comparison. However, the concentration of the inflammatory and endothelial markers are known to bioactively change in response to exercise intensity, duration, recruited muscle mass, and endurance capacity (Bernecker et al., 2011; Febbraio and Pedersen, 2002; Pedersen and Hoffman-Goetz, 2000). Information on the behavior of the markers can be limited by observing and comparing pre and post results. Observing the transient changes of the markers may further elucidate the influencing factors on the endothelial dysfunction.
Therefore, this study aimed to explore the releasing points and interactions of the parameters, using the consecutive measurements of the surrogate parameters of endothelial dysfunction and inflammatory markers during a prolonged endurance exercise. To accomplish this, representative circulating endothelial adhesion molecules, and inflammatory markers of sE-selectin, sVCAM-1, TNF-α, Creatine kinase (CK), hs-CRP, and leukocytes were measured at baseline, 100 km, 200 km, and at the end of the 308 km ultra-marathon race.
Methods
Subjects and study protocol
Marathon runners with previous completion records of 42.195 km marathons within five hours, minimum of one prior completion history of ultra-marathon with distance equal to or greater than 100 km, and 5 years of prior marathon experience were recruited voluntarily. The study protocol was approved and performed according to the guidelines set by the Ethics Committee of Inha University Hospital in Incheon, Korea. Informed consent was obtained from all subjects after a detailed explanation of the meaning and procedure of the study.
The personal history of the participants was collected by a questionnaire prior to the race. The participants were tested for the cardiovascular health 2 weeks prior to the ultra-marathon via graded exercise testing (GXT) on a treadmill. The participants were screened for cardiovascular disease, diabetes, renal disease, and hepatic disease, and blood pressure medications for the study criteria. The meaning, purpose, experimental protocol, and any possible risks related to this study were fully explained to all participants prior to obtaining written consent forms.
Out of 174 marathoners, 62 participants gave their consents to participate in the study. 38 participants were excluded from the study due to exclusion criteria (n = 26) or incompletion or voluntary withdrawal from the race (n = 12). The remaining 24 volunteers were included in the study. The subjects included male volunteers with median age and BMI of 49.5 (47 - 54) years and 23.7 (23.7 - 25.0) kg·m-2, respectively. The participants had marathon experience of 120.0 (78.8 - 141.8) months, participated in marathons 37.5 (20.0 - 60.8) times, and completed 42.195 km marathon in 218 (202.75 - 230) minutes with the best completion time of 198.0 (190.0 - 210.0) minutes. The general characteristics of the participants are summarized in Table 1.
Table 1.
Baseline characteristics, responses to graded exercise testing (GXT), and ultra-marathon records. Data are in median (interquartile ranges).
| Characteristics | All subjects (n=24) |
|---|---|
| Age (years) | 49.5 (47 - 54) |
| Height (m) | 168.3 (164.2 - 171.8) |
| Weight (kg) | 65.5 (64.2 - 71.4) |
| BMI (kg·m-2) | 23.7 (23.7 - 25.0) |
| Marathon experience (month) | 120.0 (78.8 - 141.8) |
| Number of participated marathons | 37.5 (20.0 - 60.8) |
| Average completion time (min) | 218 (202.75 - 230) |
| Record time (min) (n =20) | 198.0 (190.0 - 210.0) |
| Graded exercise testing (GXT) | |
| Exercise intensity (RPE) | 13 (11 - 13) |
| VO2max (ml·kg-1·min-1) | 47.0 (44.6 - 49.0) |
| At rest | |
| HR (bpm) | 60.0 (53.0 - 67.8) |
| SBP (mmHg) | 117 (108.0 - 128.0) |
| DBP (mmHg) | 77 (70.5 - 81.5) |
| At maximum | |
| HR (bpm) | 171.5 (164.5 - 181.5) |
| SBP (mmHg) | 215.0 (200.0 - 237.5) |
| DBP (mmHg) | 76.0 (68.0 - 82.8) |
| Total completion time (min) | 3614.5 (3393.0 - 3722.0) |
| Mean speed (km/hr) | 5.1 (5.0 - 5.5) |
| From 0 to 100 km | 7.6 (7.2 - 8.4) |
| From 100 to 200 km | 4.74 (4.6 - 5.1) * |
| From 200 to 308 km | 4.5 (4.3 - 4.6) *† |
Treadmill graded exercise testing (GXT) was conducted 4 weeks prior to the ultramarathon race on all volunteers. The symbols next to the values indicate following meanings.
* The difference is significant from 100 km, p < 0.001
† The difference is significant from 200 km, p < 0.001.
The race started from Changhoo-ri dock of Ganghwa- do at 10 a.m. The finishing point was in Kyungpo Beach of Gangneung on the east coast of South Korea. The 100 km checkpoint was at Hanam city hall in Gyeonggi-do, and the 200 km checkpoint was at Dunnae rest area in Gangwon-do. The runners were required to arrive at 100 km, 200 km, and 308 km checkpoints within 16, 39, and 64 hours, respectively. Mineral water and meals arranged in a Korean style with mostly of carbohydrates and proteins were offered at every 50 km. The runners were allowed to arrange extra food and water for additional supplementation. The ambient temperature at the starting point was 16.3°C with relative humidity of 40%. The temperature fluctuated between 13°C and 26.5°C from Ganghwa to Gangneung. The procedure of the study is summarized in Figure 1.
Figure 1.
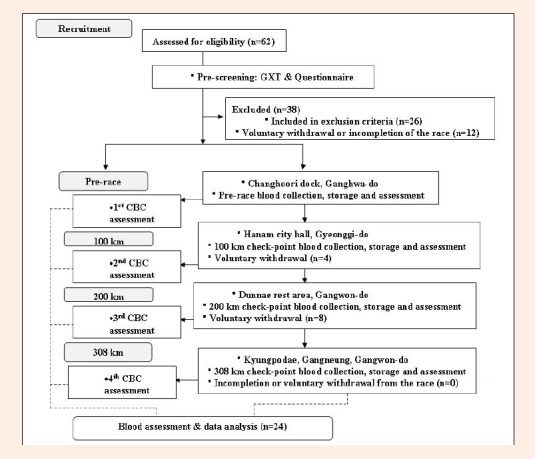
Flow chart of the study procedure.
Graded exercise testing (GXT)
GXT was performed on the participants two weeks prior to the marathon using the Bruce protocol. Twelve-lead electrocardiogram (ECG) was monitored and recorded continuously during GXT. The maximal oxygen uptake (VO2max) was defined as the highest value or the plateau of directly measured oxygen consumption using a respiratory gas analyzer (QMC; Quinton metabolic cart, Quinton Instrument CO., Boston, USA). Exercise blood pressure (BP) was measured during the last minute of each 3-min stage and at the moment of the maximum effort. BP was measured with the cuffed arm relaxed and without holding onto the side bar of the treadmill on an automatic BP monitor designed for GXT (Model 412, Quinton Instrument CO, Boston, USA). Furthermore, the use of an integrated head-set was employed by the assessors to ensure correct identification of the Korotkoff sounds for measurement of systolic and diastolic blood pressures. The maximal SBP was defined as the highest value achieved during the test. The test was terminated if the participants showed excessive fatigue to continue walking safely on a treadmill, an increase in SBP greater than 250 mmHg, chest discomfort, severe dizziness, leg fatigue, exhaustion, dyspnea, or more than 1 mm of horizontal or downslope ST segment depression. The participants with ST segment depressions were excluded from the study.
Blood sampling
Blood samples were collected from the antecubital vein of the participants according to the guidelines of the Clinical and Laboratory Standards Institute (CLSI) to identify the changes in the serum markers two hours before, 100 km, 200 km, and immediately after the ultra-marathon. The subjects consumed water, electrolytes, and carbohydrate ad libitum before, during and after the race. The blood samples were collected and immediately transferred to sterile EDTA (anticoagulant containing tube) and SST (serum separator tube) blood collection tubes (BD Vacutainer® SST, USA). EDTA blood collection tubes were immediately stored on ice and transferred to a laboratory for CBC (complete blood count) analysis. SST blood collection tubes were centrifuged at 3,400 rpm at 4 C° for 10 minutes for serum separation. Serum was separated in collection tubes and stored frozen at -70 C° for further analysis of CK, CRP, TNF-α, sVCAM-1, and sE-Selectin.
Blood analysis
The concentrations of plasma cytokines were measured using the commercial ELISA kits according to the manufacturer’s instructions. Plasma TNF-α, soluble VCAM-1, and soluble E-selectin concentrations were measured with the Quantikine Human TNF-α, Soluble VCAM-1, and sE-Selectin/CD62E kits from R&D Systems (R&D, USA). The ELISAs were performed using a microplate reader (VERSAmax, Molecular Devices, USA). The intraassay precisions of the measurement for TNF-α, sVCAM-1, and sE-Selectin were provided by the manufacturer and was reported to have 4.3%, 3.5%, and 5.7% coefficient of variations (CV) with reference values of 0.5 ~ 32 pg·mL-1, 125 ~ 4000 ng·mL-1, and 1.25 ~ 80 ng·mL-1, respectively. CK was assayed by PE, Modular Analytics using an enzymatic kinetic assay (Roche Diagnostics, Mannheim, Germany). The intra-assay precision presented by manufacturers had a coefficient of variation of 6.7 % and a reference value less than 190 U/L. HS-CRP (high sensitivity C-reactive protein) was determined by immunoturbidimetric analysis using the hs-CRP Roche Diagnostic Cobas Integra C-reactive Protein (Latex) and Modular analytics for analysis (Roche Diagnostics, Mannheim, Germany). The assay was sensitive to the value of 0.085 mg·L-1. The reference values were as followed: Low: < 1.0 mg·L-1, Average: 1.0~3.0 mg·L-1, High: > 3.0 mg·L-1. A clinical personnel performed on-site measurements for complete blood counts (CBC) for leukocytes, hematocrit, and hemoglobin. Analysis was performed with a hematology analyzer (Advia 120, Bayer Diagnostics, Berkshire, UK).
Statistical analysis
Statistical analysis of the data was performed by the Statistical Package for Social Sciences (SPSS) version 15 software (SPSS Inc., Chicago, IL, USA). Baseline characteristics and responses to GXT are expressed in median (interquartile range) for all subjects. To determine the significant changes in the endothelial dysfunction and inflammation markers for each checkpoints of baseline, 100 km, 200 km, 308 km, repeated measures analysis of variance test (ANOVA) was performed. The results are expressed in mean ± SED (Standard error of mean). The Bonferroni’s test was performed for multiple comparisons. Pearson correlation analysis was performed to examine the relationship between the markers and mean speed. The results were considered significant when the probability was less than 0.05.
Results
In order to observe the changes of the vascular endothelial and inflammation markers during prolonged strenuous and competitive physical activity, sVCAM-1, sE-Selectin, TNF-α, CK, hs-CRP, and leukocyte counts were analyzed from the blood collected at baseline, 100 km, 200 km, and 308 km. Hb and Hct were also analyzed to adjust possible hemoconcentration or hemodilution during the race according to previous studies (Dill and Costill, 1974; Maughan et al., 1985). Hb and Hct increases were measured at 100 km indicating hemoconcentration during strenuous physical activity. However, significant reductions in Hb and Hct concentrations at 200 km and 308 km showed hemodilution during prolonged physical activity. During prolonged submaximal exercise, restoration of plasma volume after the initial hemoconcentration was reported (van Beaumont et al., 1981).
Baseline characteristics and responses to graded exercise testing (GXT) are shown in Table 1. The ultra-marathons records such as total completion time and mean speed were measured at each checkpoint also shown between the checkpoints of 100 km, 200 km, and 308 km (Table 1). The total completion time and speed expressed in median and range are 3614.5 (3393.0 - 3722.0) minutes and 5.1 (5.0 - 5.5) km·hr-1, respectively. The median speeds significantly decreased from 100 km to 200 km, from 100 km to 308 km, and 200 km to 308 km (Table 1).
sVCAM-1, sE- Selectin, TNF-α, CK, hs-CRP, and leukocytes counts were analyzed for each checkpoint (Table 2). sVCAM-1 significantly increased from baseline to 100 km and 308 km. sVCAM-1 significantly decreased from 100 km to 200 km and 308 km. sE-selectin significantly increased from baseline to 100 km, 200 km, and 308 km. sE-selectin also significantly increased from 100 km to 308 km.
Table 2.
Changes in endothelial dysfunction and inflammation markers in all subjects (n=24). Data are means (±Standard error of mean).
| Markers | baseline | 100 km | 200 km | 308 km |
|---|---|---|---|---|
| sVCAM-1 (ng·mL-1) | 870.51 (37.08) | 1233.17 (78.11) * | 921.80 (46.26) † | 953.65 (41.96) *† |
| sE-Selectin (ng·mL-1) | 30.21 (2.09) | 40.14 (2.75) * | 44.55 (2.94) * | 48.08 (4.04) *† |
| TNF-α (pg·mL-1) | 3.68 (.15) | 4.00 (.20) | 3.37 (.18) † | 4.50 (.36) *‡ |
| CK (U·L-1) | 113.92 (8.65) | 1311.35 (246.02) * | 8528.82 (1409.54) *† | 9368.97 (1652.07) *† |
| hs-CRP (mg·L-1) | .40 (.10) | 5.06 (1.46) * | 25.56 (3.82) *† | 21.87 (3.49) *† |
| Leukocyte (x106/μL) | 6.20 (.32) | 13.10 (.61) * | 13.10 (.53) * | 12.86 (.93) *‡ |
| Hb (gm/dL) | 14.28 (.28) | 14.76 (.27) * | 13.91 (.26) † | 13.15 (.24) *†‡ |
| Hct (%) | 41.89 (.75) | 42.87 (.66) | 40.24 (.68) *† | 38.49 (.65) *†‡ |
All changes were compared to previous values. The symbols next to the values indicate following meanings.
*The mean difference is significant from baseline, p < 0.05,
† The mean difference is significant from 100 km, p < 0.05,
‡ The mean difference is significant from 200 km, p < 0.05, Hb: hemoglobin, Hct: hematocrit, sE-Selectin: serum E-selectin, TNF-α: tumor necrosis factor-alpha, CK: creatine kinase, hs-CRP: high sensitivity C-reactive protein, sVCAM-1: soluble vascular cell adhesion molecule-1.
TNF-α fluctuated in concentration by significantly decreasing from 100 km to 200 km , and significantly increasing from 200 km to 308 km. Overall, TNF-α increased significantly from baseline to 308 km. CK significantly increased from baseline to all checkpoints of 100 km, 200 km, and 308 km. CK also significantly increased from 100 km to 200 km and 308 km. Hs-CRP also significantly increased from baseline to all checkpoints of 100 km, 200 km, and 308 km. Leukocytes significantly increased from baseline to 100 km and 308 km. However, a significant reduction was shown at 308 km from 200 km. The correlation between sVCAM-1 and inflammatory variables and running speed as well as sE-selectin and inflammatory variables and running speed were calculated for all values from baseline to 308 km. All values were calculated after adjusting for the cardiovascular fitness or VO2max values measured during GXT. Significant correlating relationships were observed between sVCAM-1 and running speed, and leukocytes. sE-selectin showed significant correlating relationship with leukocytes, TNF-α, hs-CRP, and CK (Table 3).
Table 3.
Correlation (r) between soluble VCAM-1, e-Selectin and running speed (n=20).
| sVCAM-1 | sE-selectin | |
|---|---|---|
| Running speed (km/hr) | .365 ** | -.242 |
| Leukocytes (x106/μL) | .407 ** | .537 ** |
| hs-CRP | -.194 | .476 ** |
| TNF-α | .081 | .347 ** |
| CK | -.075 | .300 * |
All values adjusted for VO2max. Significance level:
* p < 0.01,
** p < 0.001.
Discussion
This study observed the changes in the surrogate parameters of endothelial dysfunction in relationship with the inflammatory markers by observing sVCAM-1, sE-selectin, TNF-α, CK (creatine kinase), hs-CRP (high-sensitivity CRP), and leukocytes during a prolonged and strenuous 308 km ultra-marathon race. 24 marathoners completed the 308 km ultra-marathon with blood samplings at 4 different checkpoints of baseline, 100 km, 200 km, and 308 km.
The markers observed during the race showed different responses at each checkpoint. sVCAM-1 and sE-selectin, measured for indication of vascular endothelial dysfunction, and leukocytes, measured for inflammation, showed the greatest increases at 100 km. On the other hand, TNF-α, measured for pro-inflammatory activity, significantly decreased at 200 km and continued to increase at 308 km. Furthermore, CK and hs-CRP showed the greatest increases at 200 km (Table 2).
Significantly greater increases in sVCAM-1, sE-selectin, and leukocytes at 100 km suggest a relationship between exercise intensity and endothelial function. The correlating relationship between the endothelial adhesion markers and leukocytes supports such suggestion (Table 3). Furthermore, the significant relationship between the running speed and sVCAM-1 may support a positive relationship between the vascular function and exercise intensity (Table 3). sVCAM-1, a surrogate parameter of endothelial dysfunction measured at various clinical fields, (Bolad and Delafontaine, 2005; Silvestro et al., 2005; Stewart et al., 2004) had the greatest significant increase at 100 km. The median running speed of the marathoners from the starting point to the 100 km checkpoint was comparatively vigorous in terms of exercise intensity (ACSM, 2009). Exercise intensity significantly decreased to a moderate intensity at 200 km and 308 km (Table 1).
The reasons for such correlating relationship between the endothelial adhesion molecules, leukocytes and exercise intensity could be multifactorial. Although exercise-related stress confers protection against diseases such as atherosclerosis, exercise-induced stresses may also lead to negative consequences (Marsh and Coombes, 2005). Exhaustive endurance exercise induces various significant physiological changes such as increased heat production, heart rate, leukocyte count, leftward shift of neutrophil nucleus, shear stress along with stress hormones for leukocyte release and systemic inflammation (Suzuki et al., 1999). Upon inflammatory stimulation, the endothelial barrier function is rapidly lost and adhesion molecules are expressed on the luminal surface of endothelium. Leukocytes migrate from blood into the inflammatory sites by tethering, rolling, activation, and firm adhesion to adhesion molecules expressed on endothelium (Bartzeliotou et al., 2007; Nielsen and Lyberg, 2004).
Increased leukocyte trafficking and blood velocity are characteristics of increased physical activity (Bartzeliotou et al., 2007). Shear stress increased during exercise results from oxygen demand in working muscle and a subsequent increase in cardiac output. Increased exercise intensity increases resistance or laminar shear force exerted by circulating blood upon the blood vessel walls (Marsh and Coombes, 2005; Stewart et al., 2004). Such increased shear force may increase the activation of sVCAM-1 to the risk level of endothelial dysfunction. In addition, increased systemic release of leukocytes has been related to increased stress hormones, catecholamine, cortisol, and GH, intermediated by exercise-intensity (Suzuki et al. , 1999).
The counteractive relationship should also be considered in explaining the changes in the markers during a prolong endurance race. Circulating selectin including sE-selectin is known to inhibit lymphocyte adhesion to the endothelium at a high enough concentration (Nielsen and Lyberg, 2004). Anti-inflammatory cytokines are also known to release upon increased pro-inflammatory cytokines for suppression and prevention of inflammatory tissue damage (Suzuki et al., 2003). sE-selectin also showed a relationship with the inflammatory markers such as leukocytes, hs-CRP, CK, and TNF-α (Table 3). Such results suggest a complex bioactive mechanism of sE-selectin through various influencing factors.
The inflammatory markers express different responses to exercise (Bernecker et al., 2011). CK and hs-CRP showed delayed significant increases at 200 km. TNF-α significantly decreased at 200 km and increased at 308 km. Possible reasons for such changes in the inflammatory markers could be also multifactorial. Delayed release of hs-CRP was shown at 200 km along with CK. Hs-CRP released from hepatocytes is induced by IL-6, cytokine involved in glucose homeostasis and inhibition of TNF-α production during exercise (Pue et al., 1996; Vermeire et al., 2004). Hs-CRP is also known to suppress synthesis of pro-inflammatory cytokines with the greatest plasma level shown at a day (24 hours) into the race (Pedersen and Hoffman-Goetz, 2000).
CK, a marker for muscle damage during exercise, was affected by exercise duration and intensity (Kim et al., 2007). The relationship between CK, delayed onset of muscle soreness (DOMS), and cytokines was previously reported (Nieman et al., 2005). Muscle damage depends on the rise in inflammation, induced by excessive repetition of muscle contraction. Exercise resulting in DOMS and injury induce neutrophil infiltration into the damaged tissue and delayed cytokine efflux into the circulation (Suzuki et al., 1999).
TNF-α is a known pro-inflammatory cytokine involved in pathogenesis of cardiovascular complications (Meldrum et al., 1998; Pedersen et al., 2001). Increase in TNF-α after physical activity, such as a marathon, has been associated with a higher risk of acute cardiac dysfunction (Ostrowski et al., 1999). Despite the reports of the involvement of TNF-α to the endothelial pathology, a correlation relationship between sVCAM-1 and TNF-α was not observed in this study (Dschietzig et al., 2012; Picchi et al., 2006). While sVCAM-1 significantly increased at the early phase of the race, TNF-α significantly increased toward the late phase of the race as in CK and hs-CRP. Different releasing points were observed for both markers.
TNF-α is mainly produced in macrophages and expressed by circulating monocytes or fatigued contracting muscle (Bernecker et al., 2011; Meldrum et al., 1998; Petersen and Pedersen, 2005). It was assumed that irritated exercising muscles or muscle damage caused by extended muscle contract may have elevated TNF-α at 308 km (Bernecker et al., 2011). Another reason for such delayed response may be due to different response of TNF-α to exercise. Ordinarily, in sepsis, TNF-α is the initial cytokine released followed by IL-1ॆ, IL-6, IL-1ra, sTNF- R, and IL-10 in the cytokine cascade (Pedersen and Febbraio, 2008). However, it was reported that exercise provoked primary increase in IL-6, both pro- and anti-inflammatory cytokine, followed by cytokine inhibitors such as IL-1ra and sTNF-R to inhibit TNF-α (Petersen and Pedersen, 2005).
Previous studies reported that prolonged strenuous exercise induces inflammatory and endothelial markers reaching levels seen in major trauma, septic shock, systemic inflammation, or a near-death state (Goussetis et al., 2009; Margeli et al., 2005). However, most of the studies reported of a reversible or transient increase and recovery within hours to days after the termination of exercise. The balance between endothelial injury and recovery is crucial in reducing cardiovascular events (Endemann and Schiffrin, 2004). Endothelial progenitor cells (EPCs) binds to the activated dysfunctional endothelium and reconstitute the endothelial cell layer by proliferating or survival mediating factors (Rehman et al., 2003). After a bout of strenuous exercise, hematopoietic and endothelial progenitor cells significantly rise followed by a significant elevation of mature endothelial cells with circulating VEGF levels as a physiological counteraction to maintain an intact endothelial cell layer (Mobius-Winkler et al. , 2009).
However, the elevation of EPCs varied. For example, Goussetis et al. reported of a 10-fold increase in colony forming unit of EPCs after a 246-km spartathlon, suggesting exercise related EPC release (Goussetis et al., 2009). On the other hand, other studies reported of either insignificant or downregulated hematopoietic and endothelial progenitor cells. (Endemann and Schiffrin, 2004). Based on the most recent studies, no conclusion can be drawn on the availability of exercise-related progenitor cells. Nonetheless, it would be critical to know the significant factors that trigger the changes in endothelial function as in this study.
There are many bioactive mediators involved in endothelial dysfunction. This study observed the acute changes in some of the mediators during a prolonged endurance marathon running. This study demonstrated that the surrogate markers of endothelial dysfunction were significantly influenced by exercise intensity and leukocyte concentration. The inflammatory markers, also involved in vascular pathology, responded in delayed fashion.
There are several limitations to this study. First of all, the anti-inflammatory cytokines should have been analyzed to further elucidate the counteractive relationship between the surrogate parameters of endothelial dysfunction and inflammatory cytokines. In addition, since cytokine changes are related to glycogen depletion of the musculature, (Petersen and Pedersen, 2005) the participants’ glycogen levels along with hydration level should have been closely monitored and controlled throughout the race. Increased standard deviation of the measurements toward the end of the race may be related to the exercise capacity and completion time of the participants. Finally, continuous measurements of hemodynamic responses and exercise intensities during the race may have further elucidated the responses of endothelium to increased oxygen demand and repetitive muscle contractions.
Conclusion
There are many undetermined factors that may alter the state of endothelium to the point of dysfunction. This study observed the changes in inflammatory and endothelial markers during a prolonged endurance marathon for the first time to our knowledge. The results of the study indicated that the surrogate marker of endothelial dysfunction, sVCAM-1, was closely related to leukocyte responses and exercise intensity. Increased shear stress and leukocyte trafficking were suggested to release sVCAM-1. The inflammatory markers showed delayed responses. Delayed hs-CRP release and continuously increased sE-selectin suggest anti-inflammatory responses to suppress pro-inflammatory cytokines such as TNF-α.
Biographies
Haemi Jee
Employment
Researcher at Asan Institute for Life Science
Degree
MS
Research interest
Sports medicine and exercise physiology
E-mail: amyjeez@gmail.com
Youngsoo Jin
Employment
Asan Medical Center and University of Ulsan College of Medicine, Seoul, Korea
Degree
MD, PhD
Research interest
Sports Medicine
E-mail: ysjin@amc.seoul.kr
References
- American College of Sports Medicine (2009) ACSM’s Guidelines for Exercise Testing and Prescription. 8th edition New York: Lippincott Williams & Wilkin; [Google Scholar]
- Balciunas M., Bagdonaite L., Samalavicius R., Baublys A.(2009) Markers of endothelial dysfunction after cardiac surgery: soluble forms of vascular-1 and intercellular-1 adhesion molecules. Medicina (Kaunas), 445(6), 434-439 [PubMed] [Google Scholar]
- Bartzeliotou A.I., Margeli A.P., Tsironi M., Skenderi K., Bacoula C., Chrousos G.P., Papassotiriou I.(2007) Circulating levels of adhesion molecules and markers of endothelial activation in acute inflammation induced by prolonged brisk exercise. Clinical Biochemistry 440(11), 765-770 [DOI] [PubMed] [Google Scholar]
- Bernecker C., Scherr J., Schinner S., Braun S., Scherbaum W.A., Halle M.(2011). Evidence for an exercise induced increase of TNF-alpha and IL-6 in marathon runners. Scandinavian Journal of Medicine & Science in Sports. doi:10.1111/j.1600-0838.2011.01372.x [DOI] [PubMed] [Google Scholar]
- Blake G.J., Ridker P.M.(2001). Novel clinical markers of vascular wall inflammation. Circulation Research 889(9), 763-771 [DOI] [PubMed] [Google Scholar]
- Bolad I., Delafontaine P.(2005) Endothelial dysfunction: its role in hypertensive coronary disease. Current Opinion in Cardiology 220(4), 270-274 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burke A.P., Farb A., Malcom G.T., Liang Y., Smialek J.E., Virmani R.(1999) Plaque rupture and sudden death related to exertion in men with coronary artery disease. The Journal of the American Medical Association, 2281(10), 921-926 [DOI] [PubMed] [Google Scholar]
- Di Francescomarino S., Sciartilli A., Di Valerio V., Di Baldassarre A., Gallina S.(2009) The effect of physical exercise on endothelial function. Sports Medicine 339(10), 797-812 [DOI] [PubMed] [Google Scholar]
- Dill D.B., Costill D.L.(1974) Calculation of percentage changes in volumes of blood, plasma, and red cells in dehydration. Journal of Applied Physiology 337(2), 247-248 [DOI] [PubMed] [Google Scholar]
- Dschietzig T., Brecht A., Bartsch C., Baumann G., Stangl K., Alexiou K.(2012) Relaxin improves TNF-alpha-induced endothelial dysfunction: the role of glucocorticoid receptor and phosphatidylinositol 3-kinase signalling. Cardiovascular Research 995(1), 97-107 [DOI] [PubMed] [Google Scholar]
- Dunn A.L., Marcus B.H., Kampert J.B., Garcia M.E., Kohl H.W., 3rd, Blair S.N.(1997) Reduction in cardiovascular disease risk factors: 6-month results from Project Active. Preventive Medicine 226(6), 883-892 [DOI] [PubMed] [Google Scholar]
- Endemann D.H., Schiffrin E.L.(2004) Endothelial dysfunction. Journal of the American Society of Nephrology 115(8), 1983-1992 [DOI] [PubMed] [Google Scholar]
- Febbraio M.A., Pedersen B.K.(2002) Muscle-derived interleukin-6: mechanisms for activation and possible biological roles. FASEB Journal 116(11), 1335-1347 [DOI] [PubMed] [Google Scholar]
- Fortescue E.B., Shin A.Y., Greenes D.S., Mannix R.C., Agarwal S., Feldman B.J., Shah M.I., Rifai N., Landzberg M.J., Newburger J.W., Almond C.S.(2007) Cardiac troponin increases among runners in the Boston Marathon. Annals of Emergency Medicine 449(2), 137-143 [DOI] [PubMed] [Google Scholar]
- Goel R., Majeed F., Vogel R., Corretti M.C., Weir M., Mangano C., White C., Plotnick G.D., Miller M.(2007) Exercise-induced hypertension, endothelial dysfunction, and coronary artery disease in a marathon runner. American Journal of Cardiology 999(5), 743-744 [DOI] [PubMed] [Google Scholar]
- Goussetis E., Spiropoulos A., Tsironi M., Skenderi K., Margeli A., Graphakos S., Baltopoulos P., Papassotiriou I.(2009) Spartathlon, a 246 kilometer foot race: effects of acute inflammation induced by prolonged exercise on circulating progenitor reparative cells. Blood Cells, Molecules, & Diseases 442(3), 294-299 [DOI] [PubMed] [Google Scholar]
- Jaakkola K., Jalkanen S., Kaunismaki K., Vanttinen E., Saukko P., Alanen K., Kallajoki M., Voipio-Pulkki L.M., Salmi M.(2000) Vascular adhesion protein-1, intercellular adhesion molecule-1 and P-selectin mediate leukocyte binding to ischemic heart in humans. Journal of the American College of Cardiology 336(1), 122-129 [DOI] [PubMed] [Google Scholar]
- Kim H.J., Lee Y.H., Kim C.K.(2007) Biomarkers of muscle and cartilage damage and inflammation during a 200 km run. European Journal of Applied Physiology 999(4), 443-447 [DOI] [PubMed] [Google Scholar]
- La Gerche A., Boyle A., Wilson A.M., Prior D.L.(2004) No evidence of sustained myocardial injury following an Ironman distance triathlon. International Journal of Sports Medicine 225(1), 45-49 [DOI] [PubMed] [Google Scholar]
- Margeli A., Skenderi K., Tsironi M., Hantzi E., Matalas A.L., Vrettou C., Kanavakis E., Chrousos G., Papassotiriou I.(2005) Dramatic elevations of interleukin-6 and acute-phase reactants in athletes participating in the ultradistance foot race spartathlon: severe systemic inflammation and lipid and lipoprotein changes in protracted exercise. The Journal of Clinical Endocrinology and Metabolism 990(7), 3914-3918 [DOI] [PubMed] [Google Scholar]
- Maron B.J., Poliac L.C., Roberts W.O.(1996) Risk for sudden cardiac death associated with marathon running. Journal of the American College of Cardiology 228(2), 428-431 [DOI] [PubMed] [Google Scholar]
- Marsh S.A., Coombes J.S.(2005) Exercise and the endothelial cell. Int J Cardiol 999(2), 165-169 [DOI] [PubMed] [Google Scholar]
- Maughan R.J., Whiting P.H., Davidson R.J.(1985) Estimation of plasma volume changes during marathon running. British Journal of Sports Medicine 119(3), 138-141 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meldrum D.R., Cleveland J.C., Jr., Cain B.S., Meng X., Harken A.H.(1998) Increased myocardial tumor necrosis factor-alpha in a crystalloid-perfused model of cardiac ischemia-reperfusion injury. The Annals of Thoracic Surgery 665(2), 439-443 [DOI] [PubMed] [Google Scholar]
- Mobius-Winkler S., Hilberg T., Menzel K., Golla E., Burman A., Schuler G., Adams V.(2009) Time-dependent mobilization of circulating progenitor cells during strenuous exercise in healthy individuals. Journal of Applied Physiology 1107(6), 1943-1950 [DOI] [PubMed] [Google Scholar]
- Mora S., Cook N., Buring J. E., Ridker P.M., Lee I.M.(2007) Physical activity and reduced risk of cardiovascular events: potential mediating mechanisms. Circulation 1116(19), 2110-2118 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nielsen H.G., Lyberg T.(2004). Long-distance running modulates the expression of leucocyte and endothelial adhesion molecules. Scandinavian Journal of Immunology 660(4), 356-362 [DOI] [PubMed] [Google Scholar]
- Nieman D.C., Dumke C.L., Henson D.A., McAnulty S.R., Gross S.J., Lind R.H.(2005) Muscle damage is linked to cytokine changes following a 160-km race. Brain, Behavior, and Immunity 119(5), 398-403 [DOI] [PubMed] [Google Scholar]
- Ostrowski K., Rohde T., Asp S., Schjerling P., Pedersen B.K.(1999) Pro- and anti-inflammatory cytokine balance in strenuous exercise in humans. The Journal of Physiology 5515(Pt 1), 287–291 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pedersen B.K., Akerstrom T.C., Nielsen A.R., Fischer C.P.(2007) Role of myokines in exercise and metabolism. Journal of Applied Physiology 1103(3), 1093-1098 [DOI] [PubMed] [Google Scholar]
- Pedersen B. K., Febbraio M. A.(2008). Muscle as an endocrine organ: focus on muscle-derived interleukin-6. Physiological Reviews 888(4), 1379-1406 [DOI] [PubMed] [Google Scholar]
- Pedersen B.K., Hoffman-Goetz L.(2000) Exercise and the immune system: regulation, integration, and adaptation. Physiological Reviews 880(3), 1055-1081 [DOI] [PubMed] [Google Scholar]
- Pedersen B.K., Steensberg A., Schjerling P.(2001) Exercise and interleukin-6. Current Opinion in Hematology 88(3), 137-141 [DOI] [PubMed] [Google Scholar]
- Petersen A.M., Pedersen B.K.(2005) The anti-inflammatory effect of exercise. J Appl Physiol 998(4), 1154-1162 [DOI] [PubMed] [Google Scholar]
- Picchi A., Gao X., Belmadani S., Potter B.J., Focardi M., Chilian W.M., Zhang C.(2006) Tumor necrosis factor-alpha induces endothelial dysfunction in the prediabetic metabolic syndrome. Circulation Research 999(1), 69-77 [DOI] [PubMed] [Google Scholar]
- Porreca E., Di Febbo C., di Castelnuovo A., Baccante G., Amore C., Angelini A., Di Nisio M., Donati M., Cuccurullo F., Iacoviello L.(2002). Association of factor VII levels with inflammatory parameters in hypercholesterolemic patients. Atherosclerosis 1165(1), 159-166 [DOI] [PubMed] [Google Scholar]
- Pue C.A., Mortensen R.F., Marsh C.B., Pope H.A., Wewers M.D.(1996) Acute phase levels of C-reactive protein enhance IL-1 beta and IL-1ra production by human blood monocytes but inhibit IL-1 beta and IL-1ra production by alveolar macrophages. The Journal of Immunology 1156(4), 1594-1600 [PubMed] [Google Scholar]
- Rehman J., Li J., Orschell C.M., March K.L.(2003). Peripheral blood “endothelial progenitor cells” are derived from monocyte/macrophages and secrete angiogenic growth factors. Circulation 1107(8), 1164-1169 [DOI] [PubMed] [Google Scholar]
- Scherr J., Braun S., Schuster T., Hartmann C., Moehlenkamp S., Wolfarth B., Pressler A., Halle M.(2011) 72-h kinetics of high-sensitive troponin T and inflammatory markers after marathon. Medicine and Science in Sports and Exercise 443(10), 1819-1827 [DOI] [PubMed] [Google Scholar]
- Scott J.M., Esch B.T., Shave R., Warburton D.E., Gaze D., George K.(2009). Cardiovascular consequences of completing a 160-km ultramarathon. Medicine and Science in Sports and Exercise 441(1), 26-34 [DOI] [PubMed] [Google Scholar]
- Siegel A.J.(1997) Relative risk of sudden cardiac death during marathon running. Archives of Internal Medicine 1157(11), 1269-1270 [PubMed] [Google Scholar]
- Silvestro A., Brevetti G., Schiano V., Scopacasa F., Chiariello M.(2005). Adhesion molecules and cardiovascular risk in peripheral arterial disease. Soluble vascular cell adhesion molecule-1 improves risk stratification. Thrombosis and haemostasis 93(3), 559-563 [DOI] [PubMed] [Google Scholar]
- Smith L.L.(2000)Cytokine hypothesis of overtraining: a physiological adaptation to excessive stress? Medicine and Science in Sports and Exercise 32(2), 317-331 [DOI] [PubMed] [Google Scholar]
- Stewart K.J., Sung J., Silber H.A., Fleg J.L., Kelemen M.D., Turner K.L., Bacher A.C., Dobrosielski D.A., DeRegis J.R., Shapiro E.P., Ouyang P.(2004) Exaggerated exercise blood pressure is related to impaired endothelial vasodilator function. American Journal of Hypertension 117(4), 314-320 [DOI] [PubMed] [Google Scholar]
- Suzuki K., Nakaji S., Yamada M., Liu Q., Kurakake S., Okamura N., Kumae T., Umeda T., Sugawara K.(2003) Impact of a competitive marathon race on systemic cytokine and neutrophil responses. Medicine and Science in Sports and Exercise 335(2), 348-355 [DOI] [PubMed] [Google Scholar]
- Suzuki K., Totsuka M., Nakaji S., Yamada M., Kudoh S., Liu Q., Sugawara K., Yamaya K., Sato K.(1999) Endurance exercise causes interaction among stress hormones, cytokines, neutrophil dynamics, and muscle damage. Journal of Applied Physiology 887(4), 1360-1367 [DOI] [PubMed] [Google Scholar]
- Tousoulis D., Charakida M., Stefanadis C.(2008) Endothelial function and inflammation in coronary artery disease. Postgraduate Medicine Journal 884(993), 368-371 [DOI] [PubMed] [Google Scholar]
- van Beaumont W., Underkofler S., van Beaumont S.(1981) Erythrocyte volume, plasma volume, and acid-base changes in exercise and heat dehydration. Journal of Applied Physiology 50(6), 1255-1262 [DOI] [PubMed] [Google Scholar]
- van Bussel B.C., Schouten F., Henry R.M., Schalkwijk C.G., de Boer M.R., Ferreira I., Smulders Y.M., Twisk J.W., Stehouwer C.D.(2011) Endothelial dysfunction and low-grade inflammation are associated with greater arterial stiffness over a 6-year period. Hypertension 558(4), 588-595 [DOI] [PubMed] [Google Scholar]
- Vermeire S., Van Assche G., Rutgeerts P.(2004) C-reactive protein as a marker for inflammatory bowel disease. Inflammation Bowel Disease 110(5), 661-665 [DOI] [PubMed] [Google Scholar]