

Abstract
Renal transplantation reliably evokes allo-specific B cell and T cell responses in mice. Yet, human recipients of kidney transplants with normal function usually exhibit little or no antibody specific for the transplant donor during the early weeks and months after transplantation. Indeed, the absence of anti-donor antibodies is taken to reflect effective immunosuppressive therapy and to predict a favorable outcome. Whether the absence of donor-specific antibodies reflects absence of a B-cell response to the donor, tolerance to the donor or immunity masked by binding of donor-specific antibodies to the graft is not known. To distinguish between these possibilities, we devised a novel ELISPOT, using cultured donor, recipient and third-party fibroblasts as targets. We enumerated donor-specific antibody-secreting cells in the blood of nine renal allograft recipients with normal kidney function before and after transplantation. Although none of the nine subjects had detectable donor-specific antibodies before or after transplantation, all exhibited increases in the frequency of donor-specific antibody-secreting cells eight weeks after transplantation. The responses were directed against the donor HLA-class I antigens. The increase in frequency of donor-specific antibody-secreting cells after renal transplantation indicates that B cells respond specifically to the transplant donor more often than previously thought.
Keywords: accommodation, rejection, tolerance, renal transplant
Introduction
Allogeneic transplantation reliably evokes humoral immune responses against histocompatibility antigens in animals. In the seminal instance in 1938, Gorer (1) reported that 21 of 22 naïve black mice engrafted with allogeneic sarcoma cells had detectable allo-specific antibodies. A similar if less intense response was observed after skin allografting (2). So sure was Gorer of this antibody response, he concluded its absence must reflect a limitation in the method used (3).
The antibodies produced in response to allogeneic transplantation recognize products of the major histocompatibility locus and formation of those antibodies is said to be “the invariable consequence of single or repeated antigen stimulation by transplantation of the skin or other normal tissues” (4). Consistent with this concept, Auchincloss et al. (5) detected cytotoxic antibodies in each of 14 C57BL/6 mice engrafted with skin from BALB/c mice. Klein et al. (6) detected cytotoxic allo-specific antibodies in 12 of 14 mice transplanted with congenic fetal hearts. Not only do allo-specific antibodies mark allo-immunity, they also have been used to map H-2 (6–9), and as key reagents for recognizing histocompatibility antigens (10, 11).
However, while allo-specific antibodies provide a sensitive index of allo-immunity in mice, these antibodies are usually detected in only scant quantities or not at all during the early months after clinical transplantation and are far from universal thereafter. Although kidney transplant recipients with late graft dysfunction often have donor specific antibodies in their blood (12), recipients with no evidence of graft dysfunction during the first year usually do not. Screening sera by microcytotoxicity, Martin et al. (13) detected newly produced anti-donor HLA antibodies in only 23 (9%) of 266 renal transplant recipients. Using this method, Halloran et al. (14) detected anti-donor HLA class I antibodies in the blood of 13 (20%) of 64 renal transplant recipients and all subjects with detectable anti-donor antibodies had episodes of rejection. Using a sensitive flow cytometry method, Scornik et al. (15) detected anti-donor IgG in 19 (40%) of 48 renal transplant recipients who had rejection but in only 2 (9%) of 22 who did not. Using a similar method, Christiaans et al. (16) detected antibodies against donor lymphocytes in the blood of 17 (12%) of 143 renal transplant recipients. Using a sensitive ELISA, Varnavidou-Nicolaidou et al. (17) detected anti-donor HLA antibodies in 45 (17%) of 264 renal transplant recipients and Cardarelli et al. (18) in only 10 (4%) of 251 renal transplantation recipients. Using an ELISA to test subjects thought to be at high risk of rejection, Zhang et al. (19) detected anti-donor HLA antibodies in only 11 (22%) of 49 renal transplant recipients. Ho et al. (20) detected anti-HLA antibodies in the blood of 221 (23%) of 950 cardiac allograft recipients during the first year after transplantation; however, antibodies specific for the donor were for the most part limited to the 23 subjects who experienced antibody-mediated rejection. Li et al. (21) detected antibodies against donor HLA in only 5 (6%) of 87 recipients of living-related kidney transplants. Using single antigen beads, Smith et al. (22) detected antibodies against donor HLA in the blood of 57 (25%) of 224 cardiac transplant recipients. These and other reports clearly show that human organ transplant recipients more often than not have little or no donor-specific antibody. Detection of anti-donor antibodies even in the absence of T cell or B cell complement-dependent cytotoxicity cross-matches, is associated with increased incidence of acute and/or chronic antibody-mediated rejection (23). On the other hand, induction therapies consisting of antibody and B cell depletion treatments clearly decrease incidence of antibody mediated rejection (24) suggesting that preventing anti-donor antibodies from forming may be necessary to reduce transplant rejection. However, the appearance anti-donor antibodies could instead be a consequence of kidney dysfunction, as a failing kidney may no longer absorb the antibodies which accumulate in the blood.
Absence of donor-specific antibodies in the blood of renal transplant recipients might be explained in several ways. Absence of donor-specific antibodies might reflect the effective immunosuppression of the clinical transplant recipients preventing T-cell-dependent B-cell responses, which presumably would otherwise reliably generate these antibodies in humans as they do in mice. Absence of donor-specific antibodies might reflect B-cell tolerance, as described in mice by Nemazee and Burki (25), and by Goodnow et al. (26) for proteins in mice, and by Fan et al. (27) in human recipients of ABO incompatible heart transplants. Absence of donor-specific antibodies might also give a false impression of absence of a B cell response if the antibodies produced in the response were absorbed to the graft. Consistent with the latter possibility, others (4, 28–32) and we (33, 34) have shown that large amounts of donor-specific antibody can be absorbed by organ transplants and cleared partially or fully from the blood.
Distinguishing B-cell responses characteristic of tolerance, immunosuppression or immunity is relatively simple in some settings. If the stimulating antigen is known, homogeneous and available in large amounts, the antigen can be used as a target for determining the number of antibody-secreting cells by ELISPOT (35, 36). In the conventional ELISPOT, the number of antigen-specific antibody-secreting cells remains small and stable if the response is absent, suppressed or circumscribed by anergy. The number of antigen-specific antibody-secreting cells decreases markedly if deletional tolerance occurs (27) and increases markedly if immunity occurs (37). Evaluating humoral immunity to HLA by this approach is not easily accomplished, however, because the immunizing antigens are highly polymorphic and not available in sufficient abundance to allow screening of B cells for the broad range of specificities represented in the HLA system; although testing using one or a limited number of specificities has been described(38, 39). And, even if the antigens were to be available, the representation of those antigens as isolated proteins could differ profoundly from the representation of the antigens on a cell surface, leading to over- or under-estimation of responses.
To overcome the limitations of detecting and measuring humoral immunity to organ transplants, we devised a way to enumerate antibody-secreting cells in the blood that recognize donor-specific antigens, as expressed on the surface of donor cells. We show here that the number of cells secreting donor-specific antibodies directed against HLA class I antigen(s) increases in kidney transplant recipients who have no evidence of rejection and we consider the implications for the prevalence of allo-immunity and the biology of the graft.
Materials and Methods
Human Subjects
Nine adults (8 males and 1 female; ages 34 to 60) who were to be first-time recipients of living-donor renal transplants at the University of Michigan were entered into this study. Eight recipients received kidneys from living donors (2 from related donors, 6 from unrelated donors) and one from a deceased donor. Involvement of the subjects was approved by the Institutional Review Board (IRB) of the University of Michigan and informed consent was obtained. The recipients had no history of immunological disease or treatment with immune-modulating agents and had no antibodies against the donor HLA detectable by Luminex single antigen assay (One Lambda, CA) measured at the University of Michigan histocompatibility laboratory. Following renal transplantation, the subjects received maintenance immunosuppression at the time post-transplant samples were collected. Five subjects were treated with cyclosporine, mycofenolate mofetil and prednisone in a tapering dose, and four were treated with tacrolimus, mycophenolate mofetil, alafacept and prednisone in a tapering dose. Thymoglobulin was administered before transplantation to four recipients of unrelated living donor kidneys. No subject experienced rejection during the course of this study. Sera from adults with or without antibodies directed against a broad range of HLA, as determined by the level of “panel reactive antibody” (PRA) served as positive or negative controls for ELISA.
Collection and storage of blood and tissue samples
Heparinized peripheral blood was obtained immediately prior to transplantation and 6–8 weeks afterwards. Blood was layered on Ficoll-Hypaque (GE Healthcare, Piscataway, NJ) and the peripheral blood mononuclear cells were isolated, frozen and stored in liquid nitrogen. Plasma fractions were stored at −80 °C for later use.
At the time of renal transplantation, a 10 x 10 mm segment of perinephric soft tissue was removed from the donor kidney and a 10 x 2 mm segment of Scarpa’s fascia from the wound edge of the recipient. These samples were diced with scissors into 1 x 1 mm fragments and cultured in Dulbecco’s Modified Eagle’s Medium (DMEM) supplemented with 20% fetal bovine serum (FBS), 2.5 mM L-glutamine, with penicillin and streptomycin.
Stimulation of peripheral blood mononuclear cells
Aliquots of peripheral blood mononuclear cells were thawed and cultured in RPMI supplemented with 10% FBS, 2.5mM L-glutamine, with penicillin and streptomycin. The cells were stimulated with 5 mM CpG oligonucleotide (ODN 2006, Sigma-Aldrich, St. Louis, MO), 2 ng/ml IL-15 (BD Pharmingen, San Jose, CA), 10 ng/ml CD40L (Insight Genomics, Falls Church, VA), and with 50ng/ml IL2(Insight Genomics, Falls Church, VA) in complete RPMI medium with 10% FBS, as modified from methods previously reported (40, 41) for 5 days, with fresh medium provided at day 2.5. The number of B cells in each sample of PBMC was determined by FACS using FITC-labeled monoclonal anti-CD79 (Southern Biotech, Birmingham, AL). PBMCs had between 9% and 11% B cells. Transplantation did not change the fraction of B cells in PBMCs.
Measuring the frequency of donor-specific antibody-secreting cells
The frequency of antibody-secreting cells was determined using a novel cellular ELISPOT in which fibroblasts, rather than purified antigen, served as the target. To establish the target layer, aliquots of frozen fibroblasts were thawed and cultured in DMEM with 20% FBS, penicillin and streptomycin in 96-well plates to approximately 90% confluency. To prevent ingestion of bound antibody, fibroblasts were lightly fixed by incubation with 4% paraformaldehyde, pH7.4 for 15 minutes at 25°C and washed three times with phosphate buffered saline (PBS), pH7.4, containing 1% FBS. To enumerate donor-specific antibody secreting cells, 2.5 x 105 stimulated PBMCs were re-suspended in fresh medium containing 5 mM CpG oligonucleotide (ODN 2006, Sigma-Aldrich, St. Louis, MO), 10 ng/ml IL-15 (BD Pharmingen, San Jose, CA), 10 ng/ml CD40L (Insight Genomics, Falls Church, VA), and 50ng/ml IL2 (Insight Genomics, Falls Church, VA) in complete RPMI medium with 10% FBS, added to each well in 96-well plates containing the fixed, confluent donor, recipient or third party fibroblasts. The plates were maintained at 37°C and 5% CO2 for 24 hours, and then washed three times with PBS containing 1% FBS. Secreted antibody bound to fibroblasts was detected using goat anti-human IgM- or anti-human IgG-antibodies conjugated with alkaline phosphatase (AP).
To enumerate total IgM- or IgG-secreting cells a conventional ELISPOT was performed. Each well of Immuno Spot™ M200 filter plates (Millipore, Billerica, MA) was moistened with 30 μl/well of 35% ethanol in water for 30 seconds, washed three times with sterile PBS, and coated with 100 μl of 5μg/ml of unconjugated goat anti-human IgG- or goat anti-human IgM-antibodies (SouthernBiotech, Birmingham, AL) in PBS. The wells were washed with sterile PBS, blocked with 10% FBS in PBS at room temperature for 1 hour, and then stored at room temperature with PBS for up to 2 hours before use. To each well 1 x 105 stimulated PBMCs re-suspended in fresh medium containing 10% FBS were added. The plates were maintained at 37°C and 5% CO2 for 24 hours, and then washed three times with PBS containing 1% FBS. Secreted antibody bound to the fibroblasts was detected using goat anti-human IgM or anti-human IgG, conjugated to horseradish peroxidase (HRP) (SouthernBiotech, Birmingham, AL). Bound antibody was developed according to the manufacturers’ instructions. The number of positive counts was recorded by ImmunoSpot S5 UV analyzer (CTL Analyzers, LLC)and with software version ImmunoSpot 5.0.9, set to count the center 70% of each well. Counts were corrected for the surface fraction assayed. The number of specific spots was determined by subtracting the number of spots measured in plates incubated with developing antibody without PBMCs (background). Assays were performed in triplicate.
HLA class I blockade was performed by adding 100 μg/ml mouse monoclonal antibodies that recognize HLA-A, B, and C (W6/32) (Santa Cruz Biotechnology) to monolayers of fibroblasts, for 60 minutes at 4°C, prior to incubation with PBMCs and developed as described above.
HLA typing
HLA typing was performed at the University Michigan histocompatibility laboratory for both donor and recipient by reverse Sequence Specific Oligonucleotide, according to standard procedures.
HLA specific antibody detection in blood
The presence of anti-HLA antibodies in blood was determined at the University Michigan histocompatibility laboratory by Luminex Single Antigen Assay (One Lambda, CA). The laboratory reports median fluorescence intensities (MFIs). MFIs above 700 were considered positive.
Assay for donor-specific antibodies
Levels of antibodies in the blood specific for donor or third-party cells was determined by ELISA as described previously (42), using cultured fibroblasts as targets. Monolayers of confluent fibroblasts were prepared as above and fixed with paraformaldehyde. Samples of heparinized plasma were heat inactivated at 55°C for 30 minutes, diluted in PBS containing 1% FBS, applied to wells and incubated for 60 minutes at 4°C. The wells were washed with PBS containing 1% FBS and then incubated with goat anti-human IgM and goat anti-human IgG antibodies conjugated to alkaline phosphatase(Santa Cruz Biotechnology, Santa Cruz, CA) and developed according to the manufacturers’ instructions. Absorbance was read at 405 nm on an ELISA plate reader Biotek Synergy 2 equipped with Gen5 v.1.04.4 software (Biotek Instruments, Inc, Winooski, VT 05404). Absorbance in wells incubated only with secondary antibody and in wells with autologous fibroblasts were subtracted from readings obtained with donor fibroblast wells. The assays were performed in triplicate.
Statistical analysis
Statistical analysis was performed using Prism 6 software (Graph pad, La Jolla, CA). The results of paired samples (before and after transplantation) were compared using paired T test analysis. Comparison of averages between groups was performed using unpaired two tailed T tests.
Results
To evaluate the B cell response to transplantation, we devised an approach to enumerating donor-specific antibody-secreting cells. When an antigen recognized by B cells is known and available in quantity, antibody-secreting cells can be assayed by a conventional ELISPOT. Perry et al. (38) and Heidt et al. (39) used this approach enumerate antibody secreting cells specific for common HLA class I antigens. Fan et al. (27) used the approach to enumerate antibody secreting cells specific for blood-group antigens. However, in many circumstances the histocompatibility antigens recognized by B cells are incompletely known or defined or may not be available in quantity on a surface that models the surface of a cell. Accordingly, we adapted an approach originally used by Larson et al. (43) to measure the B-cell response to epithelial tumor antigens, but using donor fibroblasts instead of tumor cells as targets. Using fibroblasts rather than B cells of the donor allowed us to focus on HLA-class I encoded antigens and to avoid the possibility that polyreactive B cells of the donor, potentially comprising half of the B cells isolated from the blood, might recognize and stimulate B cells of the recipient.
At the time of organ harvesting and transplantation, fibroblasts from nine living donors and recipients were explanted, expanded in culture and cryopreserved. A sample of these cultured cells is shown in Figure 1. To avoid as much as possible testing pre-existing or acquired responses to HLA class I, we investigated only those subjects with no donor-specific antibodies (DSA), detected by Luminex Single Antigen Assay (One Lambda) using a secondary antibody against IgG, before and after transplantation. One subject had pre-transplant reactivity against one HLA- class I haplotype and another subject had reactivity against two HLA-class I antigens not present in the donor, after transplantation. No recipient developed anti-donor antibodies after transplantation during the time of the study. To determine whether fibroblasts cultivated from tissues of kidney donors and recipients express HLA on cell surfaces and hence could serve as cellular targets for the ELISPOT, we tested the cultured cells for binding of murine monoclonal antibodies specific for a non-polymorphic domain of HLA–A, B and C. Figures 2A and 2B show that fibroblasts explanted from transplant donors and recipients expressed appreciable levels of HLA-A, B and C and did not express HLA-class II.
Figure 1. Cellular ELISPOT diagram.

Samples of perinephric fascia from donors and Scarpa’s fascia from recipients of renal transplants were explanted and used to establish primary cultures of fibroblasts. The fibroblasts generally reached confluence at day 30. PBMCs collected at the time of transplantation and 6 to 8 weeks later were cultured for 5 days prior to overlaying onto the fixed fibroblast layer and incubated for 24 hours. Antibodies bound to the fibroblasts were detected with goat anti-human IgM or IgG, AP conjugated, and developed as with a regular ELISPOT. A typical cellular ELISPOT well is shown.
Figure 2. Use of cultured human fibroblasts as a target to assay for donor-specific antibodies.


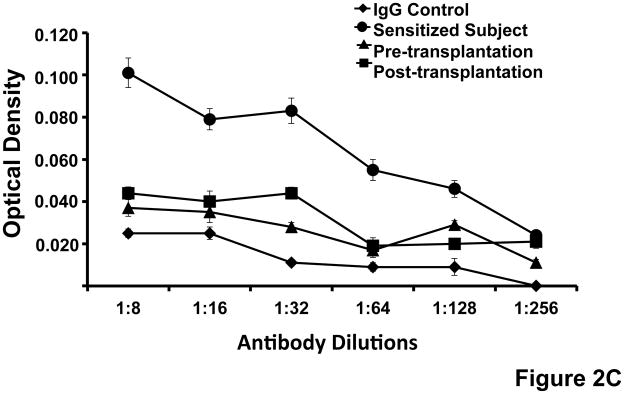
(A)Expression of HLA-class I on cultured human fibroblasts. The figure shows that cultured fibroblasts express HLA-class I detected by binding of a murine monoclonal antibody specific for a non-polymorphic domain of HLA–A, B and C (W6/32) and measured measured by ELISA. Standard errors of triplicate measurements are indicated. (B) Flow cytometry analysis of HLA-class I and class II expression by cultured fibroblasts. Shown is the light scatter plot with the lymphocyte gate depicted in red. Dot plots show staining with antibody isotype control, anti-HLA class I and anti-HLA class II antibodies. The fractions of positively stained cells are noted. (C)Use of cultured fibroblasts for assay of donor-specific antibodies in the serum of renal transplant recipients. Cultured donor fibroblasts were incubated sequentially with serial dilutions of serum from a highly sensitized subject and with serum obtained before and two months after renal transplantation and with AP labeled anti-human IgG. Donor-specific IgG was measured by detecting the optical density at 405 nm (Y-axis). The assay detects binding of IgG from a highly sensitized subject but minimal or no binding of IgG from the renal transplant recipient or pooled IgG used a control. Standard errors of triplicate measurements are indicated.
We next asked whether the recipients of renal transplants with normal graft function and lacking evidence of donor specific antibodies, detected by with Luminex Single Antigen beads, and no evidence of rejection have nonetheless antibodies specific for a donor cell surface antigen, perhaps an HLA not represented by the beads, that might be present in their transplants. To address this question we used the cultured fibroblasts as targets for ELISA. Figures 2C, 3A and 3B show that the ninerenal transplantation recipients studied had little or no antibody specific for fibroblasts of the transplant donor before or after transplantation. This result is consistent with the observations of many cited above (13–22) and others not cited in this paper, that most recipients of functioning kidney transplants have little or no detectable donor-specific antibodies in their blood during the early months after transplantation. In contrast, sera obtained from subjects sensitized to a broad range of HLA antigens in the population, as indicated by a high level of PRA, reacted strongly in the fibroblast ELISA (Figure 2C). The absence of allo-reactive antibody in the circulation of recipients could reflect failure to respond to antigen owing to immunosuppression, immunological ignorance, anergy, or tolerance, on the one hand, or a response masked by absorption of allo-specific antibody to the graft, on the other.
Figure 3. Donor-specific IgM and IgG in renal transplant recipients.


Serum was obtained from nine subjects before and two months after renal transplantation. The serum diluted 1:16 was assayed for binding of IgM (A) or IgG (B) to donor fibroblasts and expressed as absorbance at 405nm (Y-axis). The results show that renal transplant recipients had no detectable donor-specific antibodies two months after transplantation. The average ODs and standard deviations for each population were 0.04±0.003, IgM pre-TX; 0.04±0.004, IgM post-TX; 0.03±0.003, IgG pre-TX; 0.03±0.004, IgG post-TX.
Detecting such cryptic B cell responses can be challenging because normal cells can, over a period of hours, absorb enough antibody specific for plasma membrane antigens to constitute 1% of total cellular protein (44) and the activation of complement that may ensue promotes uptake and degradation (45). Thus, the absorption of antibodies and activation of complement in an organ transplant can leave scant amounts of cell-specific antibody for detection by ELISA or immunopathology (46). Still, any B cell response should be ascertainable by enumerating antibody-secreting cells in the blood. Thus, to determine whether the absence of allo-reactive antibodies in the circulation of renal transplant recipients reflects failure to respond to the antigen or a response belied by binding of antibodies to the graft, we measured the number of circulating allo-specific antibody-secreting B cells before, and two months after, transplantation by ELISPOT assay using lightly fixed monolayers of cultured cells as a target. Because some fraction of B cells in normal individuals might recognize auto- as well as donor-antigens (47, 48), we subtracted the number of B cells responding to recipient cells from the number responding to donor cells in our determination of the donor-specific response.
This approach undoubtedly under-estimates the B cell response to the transplant, but it does lend greater confidence to the responses we report.
Figure 4 shows the average number of anti-specific antibody-secreting cells measured by ELISPOT using donor fibroblasts as targets for nine renal transplant recipients with normal graft function. At the time of transplantation, these subjects had a discernable fraction of B cells (average of 86 antibody-secreting cells per 5 x 104 B cells, (Figures 4 A–C)specific for the transplant donor, as indicated by the production of IgM antibodies that defined the sites where the B cells bound to allogeneic fibroblasts. Also, as expected, a smaller but still discernable fraction of B cells recognized autologous fibroblasts (Figure 4A). Considering the difference between donor-reactive and auto-reactive B cells to represent the donor-specific response, Figures 4B and 4C show that donor-specific IgM-secreting cells increased from an average of 86 to an average of 144 antibody-secreting cells per 5 x 104 B cells, in the nine renal transplant recipients during the eight weeks after transplantation. The increase in the number of donor-specific IgM-secreting B cells suggests that, following transplantation, some fraction of the B cells committed to recognizing the donor expanded in all subjects studied. In contrast, in a typical recipient the total number of IgM- secreting cells in blood, enumerated using conventional ELISPOT, decreases slightly after transplantation (from an average of 1417 antibody-secreting cells per 5 x 104 B cells before to an average of 1003 antibody-secreting cells after transplantation) (Figure 4F).
Figure 4. Donor-specific antibody-secreting cells in renal transplant recipients.






The number and specificity of donor-specific antibody-secreting cells was determined using a novel ELISPOT in which cultured fibroblasts were used as targets and following incubation with 5 x 104 stimulated B cells in unfractionated PBMC for 24 hours. Antibody bound to the fibroblasts was detected by ELISPOT using murine monoclonal anti-human-IgM or –IgG. (A) Typical cellular ELISPOT wells. Cellular ELISPOT wells revealing anti-donor or anti-recipient (autologous) IgM or IgG antibodies. Numbers indicate the ELISPOT readings. (B) Frequency of B cells producing donor-specific IgM detected by cellular ELISPOT before and 2 months after renal transplantation. The number of antibody-secreting cells producing donor-specific IgM increased significantly after renal transplantation (p=0.0001, paired T test). (C) Frequency of B cells producing donor-specific IgM detected by cellular ELISPOT before and 2 months after renal transplantation in individual recipients. Averages of ASC before and after transplantation were compared by paired T test (p=0.0001). (D) Frequency of B cells producing donor-specific IgG detected by cellular ELISPOT before and 2 months after renal transplantation. The number of antibody-secreting cells producing donor-specific IgG increased (indicated in red) in four and decreased in two (indicated in green) of the nine subjects after renal transplantation. Pre- and post transplant values did not significantly differ by paired T test analysis (p=0.0574). (E) Frequency of B cells producing donor-specific IgG detected by cellular ELISPOT before and 2 months after renal transplantation in three typical patients. Individual experimental values and standard errors are shown. Averages of ASC before and after transplantation were compared by paired T test (p=0.0574). (F) Frequency of total IgM and IgG producing cells before and 2 months after transplantation for a typical recipient. Recipients had 1417 IgM and 411.7 IgG secreting B cells per 5x104 B cells pre-transplantation, and 1003 IgM and 265.7 IgG secreting B cells per 5x104 B cells post-transplantation, respectively. The number of IgM or IgG secreting B cells before and after transplantation did significantly differ by T test analysis (for IgM p=0.0256, for IgG p=0.0337).
As might be expected of recipients with normal graft function and no prior episodes of rejection, the number of allo-specific IgG-secreting cells was much lower than the number of IgM-secreting cells and the mean changed minimally, from 16 per 5 x 104 B cells to 28 per 5 x 104 B cells, after transplantation (Figure 4D). In contrast, the total number of IgG-secreting cells decreased after transplantation (from an average of 411.7 per 5 x 104 B cells pre-transplantation, to an average of 265.7 per 5 x 104 B cells, post-transplantation), (Figure 4F). However, in five subjects, the number of IgG secreting cells increased markedly (from 0 to 29 per 5 x 104 B cells, from 0 to 19 per 5 x 104 B cells, from 19 to 41 per 5 x 104 B cells, from 16 to 42 and from 9 to 32 per 5 x 104 B cells, Figure 4E and marked in red in Figure 4D). The subjects showing the greatest incremental increase in the number of IgG donor-specific B cells were well at the time of this analysis. Figure 4D also shows that the number of donor-specific IgG-secreting cells decreased significantly in two recipients (from 29 to 14 per 5 x 104 B cells in one, and from 18 to 8 per 5 x 104 B cells in another, Figure 4E and marked in green in Figure 4D). Whether these changes reflect sampling, clonal deletion or anergy remains to be established in longer follow up studies.
To assure that the “response” was truly donor-specific, we determined the number of antibody-secreting cells specific for third-party fibroblasts. Figures 5A and 5B show that the number of IgM- or IgG-secreting cells specific for third-party fibroblasts did not change after transplantation. Thus, B-cell responses after transplantation, as determined here, are donor-specific, and the relative absence of B-cell responses to third-party cells did not reflect idiosyncratic suppression of antibody secretion by cultured B cells, since samples from recipients bearing grafts from which fibroblast targets were derived exhibited strong responses.
Figure 5. (A) and (B). Specificity of donor-specific B cells for donor and not for third party fibroblasts.




Shown are results from cellular ELISPOTS of B cells obtained from two recipients with donor or third party fibroblasts. The number of antibody-secreting cells producing donor-specific IgM increased significantly (pair A, p=0.0015; pair B, p=0.0035, unpaired, T test) after renal transplantation while the number the number of antibody-secreting cells producing third party-specific IgM did not (pair A–B, p=0.6; pair B–A p=0.3). (C) and (D). Specificity of donor-specific B cells for HLA class I. To determine the extent to which donor-specific B cells recognize HLA class I versus other potential targets on fibroblast cell surfaces, cultured donor fibroblasts were blocked with murine anti-HLA class I antibodies and then the ELISPOT was performed. Figures 5C and 5D depict the number of anti-donor-specific IgM (C) or IgG (D)-secreting cells before and 2 months after transplantation with (+) or without (−) MHC class I blockade and shows that donor-specific antibody-secreting cells predominantly recognize HLA class I. HLA blockade was specific since incubating with an isotype control did not decrease anti-donor reactivity. Averages and standard errors of samples obtained from 4 distinct recipients were assayed in triplicate. Comparison of the means was by paired T test (IgM pre-transplantation p<0.0001, IgM post-transplantation, p=0.001, IgG pre-transplantation, p=0.0482; IgG post-transplantation, p=0.0468). * (P<0.05),** (P<0.01), *** (p<0.001).
Since B cell responses were observed in all subjects and were donor-specific and since fibroblasts do not express HLA class II (figure 2B), the most likely targets were HLA class I antigens. To test this concept, monolayers of fibroblasts were blocked with a saturating concentration of a murine monoclonal antibody that recognizes HLA-A, B and C and then used the blocked cells as targets for ELISPOT as described above. HLA class I blockade dramatically decreased the reactivity of pre-transplant (from 102.9 IgM ASC per 5 x 104 B cell, to 19.8 IgM ASC per 5 x 104 B cells, on average) and post-transplant (from 193.8 IgM ASC per 5 x 104 B cells to 32.8 IgM ASC per 5 x 104 B cells, on average) (Figure 5C). HLA class I blockade also decreased the reactivity of pre-transplant donor-specific IgG-secreting cells (from 12.5 IgG ASC per 5 x 104 B cells to 4.5 IgG ASC per 5 x 104 B cells, on average) and post-transplant (from 19.3 IgM ASC per 5 x 104 B cells to 5 IgG ASC per 5 x 104 B cells, on average) (Figure 5D). HLA blockade was specific since incubating with an isotype control did not decrease anti-donor reactivity.
Discussion
Here we report advances in concept and technique regarding the immunological response to clinical kidney transplantation. The advances stem from our application of a novel method to probe antibody secreting cells and our finding that the number of donor-specific antibody-secreting cells in the blood of transplant recipients increases during the two months following transplantation. This finding establishes that B cells in humans, as in experimental animals, do respond reliably to allogeneic transplantation and sets the stage for understanding the fate and impact of those B cells in disease and health of grafts.
Although B cells of mice usually respond to allogeneic transplantation (6), it is not intuitive that B cells of humans would do likewise. Clinical transplant recipients are treated aggressively with immunosuppressive agents that inhibit T cell activation, especially during the early weeks after transplantation, and the B-cell response to MHC class I antigens is T-cell dependent. Thus, one might anticipate an attenuation of the B-cell response in parallel with attenuation of the T-cell response (49, 50). Also consistent with the possibility that immunosuppressive therapy might impair B-cell responses to allo-antigens is the weak primary response to vaccines (51–54) and the absence of detectable donor-specific antibodies during the first several months after successful transplantation (22, 30, 55, 56). We also detected no donor-specific antibodies in the subjects we investigated. Of course, the trauma of surgery might activate and potentially expand populations of B cells non-specifically, however, the expanded population of B cells recognize donor HLA class I but not autologous or third party HLA class I and hence the expansion is donor-specific. The number of B cells secreting IgG specific for the donor MHC class I exceeded by 3.5 fold the number obtained by others(39). This difference probably reflects the fact that our assay detected antibodies directed against multiple MHC class I expressed by donor fibroblasts.
Use of cultured cells rather than purified antigens as a target for ELISPOT has some advantages and limitations worthy of comment. The B cells identified by this assay and hence the antibodies they produce recognize antigens as expressed in native conformation and in the context of the native constituents of the donor cell membrane. At the same time, the method excludes from detection B cells that recognize only denatured, degraded, or artificially clustered antigens and B cells producing antibodies the binding of which would be sterically blocked because of steric hindrance or blocking by the membrane associated glycocalyx. Accordingly, we have previously found that some antibodies capable of binding to antigens that can be isolated from cell membrane do not actually bind to those antigens on intact membranes (57, 58). Another potential disadvantage of the approach we took is that B cells potentially directed against MHC class II are excluded. Human endothelium commonly expresses MHC class II proteins (59), and hence the fraction of responding cells we describe may underestimate the total fraction of antibody-secreting cells responding. However, accurate detection of cells secreting antibody specific for HLA class II would be confounded by the properties of the most readily available HLA-class II targets (B cells and phagocytes). Up to one half of circulating B cells are polyreactive, bearing surface Ig that recognizes Ig and B cells and many phagocytes bear Fc receptors.
From our findings we can conclude that the recipients of clinical allografts do not exhibit tolerance or “suppressed” immunity to their grafts. However, it is possible that antibody secretion per se, as opposed to B-cell stimulation, is hindered in some way by immunosuppressive agents. Whether the antibody-secreting cells we detected represent activated B cells secreting antibodies that bind to the graft is consistent with our findings, but not proved. Others (4, 28–32)and we (33, 34) have shown that a normal organ or a normally functioning organ graft can absorb large amounts of anti-donor antibodies, leaving little or none remaining in the blood. Some antibodies bound to a graft might induce accommodation, a condition in which a graft or other tissue exhibits acquired resistance to antibody-mediated injury (60, 61), and this condition might explain ongoing absorption of anti-donor antibodies (59, 61). If the increase in the number of donor-specific B cells does reflect accommodation, then accommodation might well be a common outcome of allo-transplantation during at least the first several months and the classical way of detecting accommodation, i.e. the simultaneous presence of antibody in the blood and normal graft function (62, 63), might significantly underestimate the prevalence. In this case, our original definition of accommodation needs to be revised to encompass some organ transplant recipients who have no detectable antibody against the donor.
Acknowledgments
Supported by grants from the National Institutes of Health HL52297, HL79067 (JLP and MC) and by an ASTS-Roche Laboratories Scientist Scholarship (RL).
The authors thank Ms. Mary Maliarik for organizing the recruitment of patients for the study and Ms. Jane Bugden for editorial help.
Footnotes
Disclosure
The authors of this manuscript have no conflicts of interest to disclose as described by the American Journal of Transplantation.
References
- 1.Gorer PA. The antigenic basis of tumour transplantation. Journal of Pathology and Bacteriology. 1938;47:231–252. [Google Scholar]
- 2.Gorer PA. The antibody response to skin homografts in mice. Annals of the New York Academy of Sciences. 1955;59(3):365–373. doi: 10.1111/j.1749-6632.1955.tb45948.x. [DOI] [PubMed] [Google Scholar]
- 3.Gorer P. Some recent work on tumor immunity. Advances in Cancer Research. 1956;4:149–186. doi: 10.1016/s0065-230x(08)60724-1. [DOI] [PubMed] [Google Scholar]
- 4.Stetson CA. The role of humoral antibody in the homograft reaction. Advances in Immunology. 1963;3:97–130. [Google Scholar]
- 5.Auchincloss H, Jr, Ghobrial RR, Russell PS, Winn HJ. Prevention of alloantibody formation after skin grafting without prolongation of graft survival by anti-L3T4 in vivo. Transplantation. 1988;45(6):1118–1123. doi: 10.1097/00007890-198806000-00024. [DOI] [PubMed] [Google Scholar]
- 6.Klein J, Chiang C, Lofgreen J, Steinmuller D. Participation of H-2 regions in heart-transplant rejection. Transplantation. 1976;22(4):384–390. doi: 10.1097/00007890-197610000-00011. [DOI] [PubMed] [Google Scholar]
- 7.Amos DB, Gorer PA, Mikulska BM, Billingham RE, Sparrow EM. An antibody response to skin homografts in mice. British journal of experimental pathology. 1954;35(2):203–208. [PMC free article] [PubMed] [Google Scholar]
- 8.Klein J, Hauptfeld V, Hauptfeld M. Evidence for a Third, Ir-Associated Histocompatibility Region in the H-2 Complex of the Mouse. Immunogenetics. 1974;1:56–56. [Google Scholar]
- 9.Klein J, Geib R, Chiang C, Hauptfeld V. Histocompatibility antigens controlled by the I region of the murine H-2 complex. I. Mapping of H-2A and H-2C loci. The Journal of experimental medicine. 1976;143(6):1439–1452. doi: 10.1084/jem.143.6.1439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Snell GD. The genetics of transplantation. Journal of the National Cancer Institute. 1953;14(3):691–704. [PubMed] [Google Scholar]
- 11.Terasaki P, McClelland JP. Microdroplet assay of human serum cytotoxins. Nature. 1964;204:998–1000. doi: 10.1038/204998b0. [DOI] [PubMed] [Google Scholar]
- 12.Matas AJ, Leduc R, Rush D, Cecka JM, Connett J, Fieberg A, et al. Histopathologic clusters differentiate subgroups within the nonspecific diagnoses of CAN or CR: preliminary data from the DeKAF study. American journal of transplantation. 2010;10(2):315–323. doi: 10.1111/j.1600-6143.2009.02943.x. [DOI] [PubMed] [Google Scholar]
- 13.Martin S, Dyer PA, Mallick NP, Gokal R, Harris R, Johnson RW. Posttransplant antidonor lymphocytotoxic antibody production in relation to graft outcome. Transplantation. 1987;44(1):50–53. doi: 10.1097/00007890-198707000-00012. [DOI] [PubMed] [Google Scholar]
- 14.Halloran PF, Schlaut J, Solez K, Srinivasa NS. The significance of the anti-class I response. II. Clinical and pathologic features of renal transplants with anti-class I-like antibody. Transplantation. 1992;53(3):550–555. [PubMed] [Google Scholar]
- 15.Scornik JC, Salomon DR, Lim PB, Howard RJ, Pfaff WW. Posttransplant antidonor antibodies and graft rejection. Evaluation by two-color flow cytometry. Transplantation. 1989;47(2):287–290. doi: 10.1097/00007890-198902000-00018. [DOI] [PubMed] [Google Scholar]
- 16.Christiaans MH, Overhof-de Roos R, Nieman F, van Hooff JP, van den Berg-Loonen EM. Donor-specific antibodies after transplantation by flow cytometry: relative change in fluorescence ratio most sensitive risk factor for graft survival. Transplantation. 1998;65(3):427–433. doi: 10.1097/00007890-199802150-00024. [DOI] [PubMed] [Google Scholar]
- 17.Varnavidou-Nicolaidou A, Iniotaki-Theodoraki AG, Doxiadis II, Georgiou D, Patargias T, Stavropoulos-Giokas C, et al. Expansion of humoral donor-specific alloreactivity after renal transplantation correlates with impaired graft outcome. Human immunology. 2005;66(9):985–988. doi: 10.1016/j.humimm.2005.07.002. [DOI] [PubMed] [Google Scholar]
- 18.Cardarelli F, Pascual M, Tolkoff-Rubin N, Delmonico FL, Wong W, Schoenfeld DA, et al. Prevalence and significance of anti-HLA and donor-specific antibodies long-term after renal transplantation. Transplant international. 2005;18(5):532–540. doi: 10.1111/j.1432-2277.2005.00085.x. [DOI] [PubMed] [Google Scholar]
- 19.Zhang Q, Liang LW, Gjertson DW, Lassman C, Wilkinson AH, Kendrick E, et al. Development of posttransplant antidonor HLA antibodies is associated with acute humoral rejection and early graft dysfunction. Transplantation. 2005;79(5):591–598. doi: 10.1097/01.tp.0000155246.52249.ac. [DOI] [PubMed] [Google Scholar]
- 20.Ho EK, Vlad G, Vasilescu ER, de la Torre L, Colovai AI, Burke E, et al. Pre- and posttransplantation allosensitization in heart allograft recipients: major impact of de novo alloantibody production on allograft survival. Human immunology. 2011;72(1):5–10. doi: 10.1016/j.humimm.2010.10.013. [DOI] [PubMed] [Google Scholar]
- 21.Li X, Ishida H, Yamaguchi Y, Tanabe K. Poor graft outcome in recipients with de novo donor-specific anti-HLA antibodies after living related kidney transplantation. Transplant international. 2008;21(12):1145–1152. doi: 10.1111/j.1432-2277.2008.00755.x. [DOI] [PubMed] [Google Scholar]
- 22.Smith JD, Banner NR, Hamour IM, Ozawa M, Goh A, Robinson D, et al. De novo donor HLA-specific antibodies after heart transplantation are an independent predictor of poor patient survival. American journal of transplantation. 2011;11(2):312–319. doi: 10.1111/j.1600-6143.2010.03383.x. [DOI] [PubMed] [Google Scholar]
- 23.Hirai T, Kohei N, Omoto K, Ishida H, Tanabe K. Significance of low-level DSA detected by solid-phase assay in association with acute and chronic antibody-mediated rejection. Transplant international. 2012;25(9):925–934. doi: 10.1111/j.1432-2277.2012.01518.x. [DOI] [PubMed] [Google Scholar]
- 24.Kohei N, Hirai T, Omoto K, Ishida H, Tanabe K. Chronic antibody-mediated rejection is reduced by targeting B-cell immunity during an introductory period. American journal of transplantation. 2012;12(2):469–476. doi: 10.1111/j.1600-6143.2011.03830.x. [DOI] [PubMed] [Google Scholar]
- 25.Nemazee DA, Burki K. Clonal deletion of B lymphocytes in a transgenic mouse bearing anti-MHC class I antibody genes. Nature. 1989;337(6207):562–566. doi: 10.1038/337562a0. [DOI] [PubMed] [Google Scholar]
- 26.Goodnow CC, Crosbie J, Jorgensen H, Brink RA, Basten A. Induction of self-tolerance in mature peripheral B lymphocytes. Nature. 1989;342(6248):385–391. doi: 10.1038/342385a0. [DOI] [PubMed] [Google Scholar]
- 27.Fan X, Ang A, Pollock-BarZiv SM, Dipchand AI, Ruiz P, Wilson G, et al. Donor-specific B-cell tolerance after ABO-incompatible infant heart transplantation. Nature medicine. 2004;10(11):1227–1233. doi: 10.1038/nm1126. [DOI] [PubMed] [Google Scholar]
- 28.Mohanakumar T, Waldrep JC, Phibbs M, Mendez-Picon G, Kaplan AM, Lee HM. Serological characterization of antibodies eluted from chronically rejected human renal allografts. Transplantation. 1981;32(1):61–66. doi: 10.1097/00007890-198107000-00013. [DOI] [PubMed] [Google Scholar]
- 29.Martin L, Guignier F, Mousson C, Rageot D, Justrabo E, Rifle G. Detection of donor-specific anti-HLA antibodies with flow cytometry in eluates and sera from renal transplant recipients with chronic allograft nephropathy. Transplantation. 2003;76(2):395–400. doi: 10.1097/01.TP.0000078895.24606.45. [DOI] [PubMed] [Google Scholar]
- 30.Briggs D, Zehnder D, Higgins RM. Development of non-donor-specific HLA antibodies after kidney transplantation: frequency and clinical implications. Contributions to nephrology. 2009;162:107–116. doi: 10.1159/000170843. [DOI] [PubMed] [Google Scholar]
- 31.Higgins R, Lowe D, Hathaway M, Lam F, Kashi H, Tan LC, et al. Rises and falls in donor-specific and third-party HLA antibody levels after antibody incompatible transplantation. Transplantation. 2009;87(6):882–888. doi: 10.1097/TP.0b013e31819a6788. [DOI] [PubMed] [Google Scholar]
- 32.Ishida H, Hirai T, Kohei N, Yamaguchi Y, Tanabe K. Significance of qualitative and quantitative evaluations of anti-HLA antibodies in kidney transplantation. Transplant international. 2011;24(2):150–157. doi: 10.1111/j.1432-2277.2010.01166.x. [DOI] [PubMed] [Google Scholar]
- 33.Collins BH, Cotterell AH, McCurry KR, Alvarado CG, Magee JC, Parker W, et al. Cardiac xenografts between primate species provide evidence for the importance of the alpha-galactosyl determinant in hyperacute rejection. Journal of immunology. 1995;154(10):5500–5510. [PubMed] [Google Scholar]
- 34.McCurry KR, Parker W, Cotterell AH, Weidner BC, Lin SS, Daniels LJ, et al. Humoral responses to pig-to-baboon cardiac transplantation: implications for the pathogenesis and treatment of acute vascular rejection and for accommodation. Human immunology. 1997;58(2):91–105. doi: 10.1016/s0198-8859(97)00229-2. [DOI] [PubMed] [Google Scholar]
- 35.Sedgwick JD, Holt PG. A solid-phase immunoenzymatic technique for the enumeration of specific antibody-secreting cells. Journal of Immunological Methods. 1983;57(1–3):301–309. doi: 10.1016/0022-1759(83)90091-1. [DOI] [PubMed] [Google Scholar]
- 36.Czerkinsky CC, Nilsson L, Nygren H, Ouchterlony O, Tarkowski A. A solid-phase enzyme-linked immunospot (ElISPOT) assay for enumeration of specific antibody-secreting cells. Journal of Immunological Methods. 1983;65:109–121. doi: 10.1016/0022-1759(83)90308-3. [DOI] [PubMed] [Google Scholar]
- 37.Tarkowski A, Czerkinsky C, Nilsson LA. Simultaneous induction of rheumatoid factor- and antigen-specific antibody-secreting cells during the secondary immune response in man. Clinical and experimental immunology. 1985;61(2):379–387. [PMC free article] [PubMed] [Google Scholar]
- 38.Perry DK, Pollinger HS, Burns JM, Rea D, Ramos E, Platt JL, et al. Two novel assays of alloantibody-secreting cells demonstrating resistance to desensitization with IVIG and rATG. American Journal of Transplantation. 2008;8(1):133–143. doi: 10.1111/j.1600-6143.2007.02039.x. [DOI] [PubMed] [Google Scholar]
- 39.Heidt S, Roelen DL, de Vaal YJ, Kester MG, Eijsink C, Thomas S, et al. A NOVel ELISPOT assay to quantify HLA-specific B cells in HLA-immunized individuals. American journal of transplantation. 2012;12(6):1469–1478. doi: 10.1111/j.1600-6143.2011.03982.x. [DOI] [PubMed] [Google Scholar]
- 40.Bernasconi NL, Traggiai E, Lanzavecchia A. Maintenance of serological memory by polyclonal activation of human memory B cells. Science. 2002;298(5601):2199–2202. doi: 10.1126/science.1076071. [DOI] [PubMed] [Google Scholar]
- 41.Jourdan M, Caraux A, De Vos J, Fiol G, Larroque M, Cognot C, et al. An in vitro model of differentiation of memory B cells into plasmablasts and plasma cells including detailed phenotypic and molecular characterization. Blood. 2009;114(25):5173–5181. doi: 10.1182/blood-2009-07-235960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Parker W, Bruno D, Holzknecht ZE, Platt JL. Characterization and affinity isolation of xenoreactive human natural antibodies. Journal of immunology. 1994;153(8):3791–3803. [PubMed] [Google Scholar]
- 43.Larson LN, Johansson C, Lindholm L. Localization of antibody-secreting cells against human colon adenocarcinoma cell lines using the enzyme-linked immunospot assay. International archives of allergy and applied immunology. 1988;87(4):405–408. doi: 10.1159/000234710. [DOI] [PubMed] [Google Scholar]
- 44.Schneider YJ, Tulkens P, de Duve C, Trouet A. Fate of plasma membrane during endocytosis. I. Uptake and processing of anti-plasma membrane and control immunoglobulins by cultured fibroblasts. The Journal of cell biology. 1979;82(2):449–465. doi: 10.1083/jcb.82.2.449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Mantovani B, Rabinovitch M, Nussenzweig V. Phagocytosis of immune complexes by macrophages. Different roles of the macrophage receptor sites for complement (C3) and for immunoglobulin (IgG) The Journal of experimental medicine. 1972;135(4):780–792. doi: 10.1084/jem.135.4.780. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Platt JL, Cascalho M. Donor specific antibodies after transplantation. Pediatric Transplantation. 2011;15(7):686–690. doi: 10.1111/j.1399-3046.2010.01436.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Zhou ZH, Notkins AL. Polyreactive antigen-binding B (PAB−) cells are widely distributed and the PAB population consists of both B-1+ and B-1− phenotypes. Clinical and experimental immunology. 2004;137(1):88–100. doi: 10.1111/j.1365-2249.2004.02511.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Zhou ZH, Tzioufas AG, Notkins AL. Properties and function of polyreactive antibodies and polyreactive antigen-binding B cells. J Autoimmun. 2007;29(4):219–228. doi: 10.1016/j.jaut.2007.07.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Mestre M, Massip E, Bas J, Alsina J, Romeu A, Castelao AM, et al. Longitudinal study of the frequency of cytotoxic T cell precursors in kidney allograft recipients. Clinical and experimental immunology. 1996;104(1):108–114. doi: 10.1046/j.1365-2249.1996.d01-657.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Baker RJ, Hernandez-Fuentes MP, Brookes PA, Chaudhry AN, Lechler RI. The role of the allograft in the induction of donor-specific T cell hyporesponsiveness. Transplantation. 2001;72(3):480–485. doi: 10.1097/00007890-200108150-00020. [DOI] [PubMed] [Google Scholar]
- 51.Stiver HG, Graves P, Meiklejohn G, Schroter G, Eickhoff TC. Impaired serum antibody response to inactivated influenza A and B vaccine in renal transplant recipients. Infection and immunity. 1977;16(3):738–741. doi: 10.1128/iai.16.3.738-741.1977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Admon D, Engelhard D, Strauss N, Goldman N, Zakay-Rones Z. Antibody response to influenza immunization in patients after heart transplantation. Vaccine. 1997;15(14):1518–1522. doi: 10.1016/s0264-410x(97)00193-x. [DOI] [PubMed] [Google Scholar]
- 53.Candon S, Thervet E, Lebon P, Suberbielle C, Zuber J, Lima C, et al. Humoral and cellular immune responses after influenza vaccination in kidney transplant recipients. American journal of transplantation. 2009;9(10):2346–2354. doi: 10.1111/j.1600-6143.2009.02787.x. [DOI] [PubMed] [Google Scholar]
- 54.Meyer S, Adam M, Schweiger B, Ilchmann C, Eulenburg C, Sattinger E, et al. Antibody response after a single dose of an AS03-adjuvanted split-virion influenza A (H1N1) vaccine in heart transplant recipients. Transplantation. 2011;91(9):1031–1035. doi: 10.1097/TP.0b013e3182115be0. [DOI] [PubMed] [Google Scholar]
- 55.Hidalgo LG, Campbell PM, Sis B, Einecke G, Mengel M, Chang J, et al. De novo donor-specific antibody at the time of kidney transplant biopsy associates with microvascular pathology and late graft failure. American journal of transplantation. 2009;9(11):2532–2541. doi: 10.1111/j.1600-6143.2009.02800.x. [DOI] [PubMed] [Google Scholar]
- 56.Cooper JE, Gralla J, Cagle L, Goldberg R, Chan L, Wiseman AC. Inferior kidney allograft outcomes in patients with de novo donor-specific antibodies are due to acute rejection episodes. Transplantation. 2011;91(10):1103–1109. doi: 10.1097/TP.0b013e3182139da1. [DOI] [PubMed] [Google Scholar]
- 57.Everett ML, Lin SS, Worrell SS, Platt JL, Parker W. The footprint of antibody bound to pig cells: evidence of complex surface topology. Biochemical and biophysical research communications. 2003;301(3):751–757. doi: 10.1016/s0006-291x(03)00043-3. [DOI] [PubMed] [Google Scholar]
- 58.Parker W, Stitzenberg KB, Yu PB, Pratt VS, Nakamura YC, Farel LS, et al. Biophysical characteristics of anti-Gal(alpha)1–3Gal IgM binding to cell surfaces: implications for xenotransplantation. Transplantation. 2001;71(3):440–446. doi: 10.1097/00007890-200102150-00018. [DOI] [PubMed] [Google Scholar]
- 59.Platt JL, LeBien TW, Michael AF. Stages of renal ontogenesis identified by monoclonal antibodies reactive with lymphohemopoietic differentiation antigens. The Journal of experimental medicine. 1983;157(1):155–172. doi: 10.1084/jem.157.1.155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Lynch RJ, Platt JL. Accommodation in organ transplantation. Current opinion in organ transplantation. 2008;13(2):165–170. doi: 10.1097/MOT.0b013e3282f6391e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Pelletier RP, Hennessy PK, Adams PW, VanBuskirk AM, Ferguson RM, Orosz CG. Clinical significance of MHC-reactive alloantibodies that develop after kidney or kidney-pancreas transplantation. American journal of transplantation. 2002;2(2):134–141. doi: 10.1034/j.1600-6143.2002.020204.x. [DOI] [PubMed] [Google Scholar]
- 62.Koch CA, Khalpey ZI, Platt JL. Accommodation: preventing injury in transplantation and disease. J Immunol. 2004;172(9):5143–5148. doi: 10.4049/jimmunol.172.9.5143. [DOI] [PubMed] [Google Scholar]
- 63.Platt JL, Vercellotti GM, Dalmasso AP, Matas AJ, Bolman RM, Najarian JS, et al. Transplantation of discordant xenografts: a review of progress. Immunology today. 1990;11(12):450–457. doi: 10.1016/0167-5699(90)90174-8. [DOI] [PubMed] [Google Scholar]