

Abstract
There is increasing evidence that infection with the Epstein-Barr virus (EBV) plays a role in the development of multiple sclerosis (MS), a chronic inflammatory demyelinating disease of the CNS. This article provides a four-tier hypothesis proposing (1) EBV infection is essential for the development of MS; (2) EBV causes MS in genetically susceptible individuals by infecting autoreactive B cells, which seed the CNS where they produce pathogenic autoantibodies and provide costimulatory survival signals to autoreactive T cells that would otherwise die in the CNS by apoptosis; (3) the susceptibility to develop MS after EBV infection is dependent on a genetically determined quantitative deficiency of the cytotoxic CD8+ T cells that normally keep EBV infection under tight control; and (4) sunlight and vitamin D protect against MS by increasing the number of CD8+ T cells available to control EBV infection. The hypothesis makes predictions that can be tested, including the prevention and successful treatment of MS by controlling EBV infection.
Keywords: multiple sclerosis, Epstein-Barr virus, CD8 T cell, vitamin D, sunlight, treatment, autoimmune
Multiple sclerosis (MS) is a common chronic inflammatory demyelinating disease of the CNS, causing severe progressive disability, particularly in young people, and affecting more than 2.5 million people worldwide. A large body of evidence indicates that MS is an autoimmune disease (Sospedra and Martin 2005; Pender and Greer 2007), but the primary cause of MS and the other human chronic autoimmune diseases is yet unknown. There is now substantial evidence that infection with the Epstein-Barr virus (EBV) plays a major role in the development of MS. This article provides a four-tier hypothesis proposing (1) that EBV has an essential role in the development of MS, (2) how EBV infection causes MS, (3) why EBV infection causes MS in some people but not others, and (4) how sunlight protects against MS (see Box 2). The hypothesis is presented after a general introduction to EBV and a review of the evidence implicating EBV in the pathogenesis of MS.
Box 2. Four-Tier Hypothesis for the Role of Epstein-Barr Virus in the Pathogenesis of Multiple Sclerosis.
Infection with the Epstein-Barr virus (EBV) is essential for the development of multiple sclerosis (MS).
EBV causes MS in genetically susceptible individuals by infecting autoreactive B cells, which seed the CNS, where they produce pathogenic autoantibodies and provide costimulatory survival signals to autoreactive T cells that would otherwise die in the CNS by apoptosis.
The susceptibility to develop MS after EBV infection is dependent on a genetically determined quantitative deficiency of the cytotoxic CD8+ T cells that normally keep EBV infection under tight control.
Sunlight and vitamin D protect against MS by increasing the number of CD8+ T cells available to control EBV infection.
Epstein-Barr Virus
EBV is a ubiquitous human herpesvirus that has the unique ability to infect, activate, and latently persist in B lymphocytes for the lifetime of the infected individual. During primary infection, EBV transmitted via saliva infects naive B cells in the tonsil through binding of the viral surface glycoprotein gp350 to complement receptor 2 (CR2; also known as CD21), which is expressed by mature B cells and follicular dendritic cells (Nemerow and others 1987). EBV drives the infected B cell out of the resting state to become an activated B blast and then exploits the normal pathways of B cell differentiation so that the B blast can become a latently infected resting memory B cell (Thorley-Lawson and Gross 2004; Fig. 1). To achieve this, the virus deploys a series of different latency transcription programs (Thorley-Lawson and Gross 2004). On initial infection of naive B cells, EBV uses the latency III or “growth” program in which all viral latent proteins are expressed—namely, the Epstein-Barr nuclear antigens (EBNA) 1, 2, 3A, 3B, 3C, and LP and the latent membrane proteins (LMP) 1, 2A, and 2B. This activates the B cell to become a B blast, which then enters a germinal center in the tonsil where it down-regulates the expression of the EBNA proteins 2, 3A, 3B, 3C, and LP. The continuing expression of EBNA1, LMP1, and LMP2 (latency II or “default” program) allows the infected B cell to progress through a germinal center reaction to become a memory B cell, which exits from the germinal center and circulates in the blood. The EBV-infected memory B cell expresses no viral proteins except during cell division, when it expresses only EBNA1 (latency I), which engages the host cell DNA polymerase and ensures transmission of the viral genome to the daughter cells. The lack of viral protein expression allows the virus to persist in memory B cells despite a healthy immune response. Latently infected memory B cells returning to the tonsil can terminally differentiate into plasma cells, which initiates the lytic (replicative) transcription program with the production of infectious virus (Laichalk and Thorley-Lawson 2005). The resulting free virions infect tonsil epithelial cells, where the virus replicates at a high rate and is continuously shed into saliva for transmission to new hosts (Hadinoto and others 2009). Newly formed virus can also infect additional naive B cells in the same host.
Figure 1.

Normal sequence of events during infection of the tonsil by Epstein-Barr virus (EBV). During primary infection, EBV entering the tonsil from the saliva infects naive B cells, driving them out of the resting state to become activated B blasts, which then enter germinal centers where they proliferate intensely and differentiate into latently infected memory B cells, which then exit from the tonsil and circulate in the blood. The infected memory B cells do not express any viral proteins except during cell division when they express Epstein-Barr nuclear antigen 1 (EBNA1). Latently infected memory B cells returning to the tonsil can differentiate into plasma cells, which initiates the lytic phase of infection with the production of free virus particles (virions). The virions infect tonsil epithelial cells, where the virus replicates at a high rate and is shed into saliva for transmission to new hosts. Newly formed virus can also infect additional naive B cells in the same host. During primary infection, this cycle initially proceeds unchecked by the immune system. However, the infected host soon mounts an immune response against the virus. EBV-specific cytotoxic CD8+ T cells kill infected B cells expressing viral proteins, and anti-EBV antibodies neutralize viral infectivity by binding to free virus. Red lines with perpendicular bars indicate inhibition. This model is based on work published by Thorley-Lawson and colleagues (Thorley-Lawson and Gross 2004; Laichalk and Thorley-Lawson 2005; Hadinoto and others 2009; Roughan and others 2010).
Box 1. Glossary.
B cell receptor (BCR): The cell surface receptor of B cells for specific antigen. Its binding moiety is composed of a transmembrane immunoglobulin (Ig) (antibody) molecule.
Class-switch recombination: The process by which a B cell’s production of antibody changes from the IgM class of antibody, which every B cell expresses initially, to the IgG, IgA, or IgE class of antibody. This takes place in germinal centers and is mediated by recombination of C (constant) region Ig genes, which determine the class of the antibody and thus its effector function. Because the variable regions of the Ig genes do not change, class switching does not affect antigen specificity.
Germinal center: Site within lymphoid follicle where B cells primed with their specific antigen undergo intense proliferation, somatic hypermutation of their Ig genes, and class-switch recombination. Mutant clones with high affinity for antigen are positively selected (affinity maturation) and differentiate into the long-lived antibody-secreting cells and the memory B cells that sustain serological immunity after infection, whereas B cells with low affinity for antigen die by apoptosis. The complete process is known as the germinal center reaction.
HLA (Human leukocyte antigen): The human major histocompatibility complex, a cluster of genes encoding a set of membrane glycoproteins that present antigenic peptides to T cells. The HLA class I molecules present peptides generated in the cytosol to CD8+ T cells, and HLA class II molecules present peptides degraded in intracellular vesicles to CD4+ T cells.
Latent infection: A stage of viral infection in which the virus does not replicate within the infected host cell.
Lymphoblastoid cell line (LCL): A line of B cells transformed by in vitro infection with Epstein-Barr virus (EBV) and capable of indefinite growth through the effect of EBV-encoded proteins.
Lymphoid follicle: Localized collection of B cells within secondary (peripheral) lymphoid tissue (lymph nodes, spleen, and mucosa-associated lymphoid tissue such as the tonsils).
Lytic infection: A stage of viral infection in which the virus replicates within the infected host cell, leading to the destruction (lysis) of the cell and release of complete virus particles (virions), which can infect other cells.
Naive B cell: B cell that has not been exposed to its specific antigen.
Seropositivity: The presence of antibodies in the serum indicating prior exposure to a particular organism or antigen.
Somatic hypermutation: The germinal center process of introducing point mutations into the rearranged V (variable) regions of Ig genes at a very high rate, giving rise to mutant BCRs on the surface of the B cells, some of which bind antigen better than the original BCRs, with resultant increased affinity for antigen. B cells expressing high affinity receptors are preferentially selected to mature into antibody-secreting cells. These mutations affect only somatic cells and are not inherited through germline transmission.
Latently infected memory B cells display the molecular hallmarks of classical antigen-selected memory B cells—namely, somatic hypermutation and class-switch recombination of their immunoglobulin (Ig) genes (Souza and others 2005). In normal B cell differentiation, naive B cells are activated by antigen through the B cell receptor (BCR) and by T cell help through the CD40 receptor so that they proliferate and progress through a germinal center reaction. Remarkably, LMP2A and LMP1, which are expressed by EBV during latency II and latency III, mimic the antigen-activated BCR and the activated CD40 receptor, respectively. In vitro LMP2A can mimic and replace constitutive BCR signaling and thereby support an activated, proliferative state in B cells, which it renders resistant to apoptosis (Mancao and Hammerschmidt 2007). In transgenic mice, LMP1 can act as a constitutively active CD40 receptor that completely substitutes for CD40 signaling in B cells, leading to normal B cell development, activation, and immune responses, including class-switch recombination, germinal center formation, and somatic hypermutation (Rastelli and others 2008). It was initially thought that in the human tonsil, LMP2A and LMP1 drive infected B cells through a germinal center reaction independently of antigen and T cell help (Thorley-Lawson and Gross 2004), but recent evidence suggests that they work synergistically with BCR signaling and CD40 signaling, respectively (Roughan and Thorley-Lawson 2009). Another difference between the in vitro and in vivo behavior of EBV-infected B cells is that when EBV infects B cells in vitro, it activates them to become lymphoblasts, which proliferate indefinitely to form a B cell lymphoblastoid cell line (LCL), whereas in vivo, it appears that newly infected lymphoblasts in the tonsil of healthy EBV carriers undergo very limited proliferation prior to entering the germinal center, where they proliferate extensively and differentiate into memory B cells (Roughan and others 2010). The continuous proliferation of B lymphoblasts in LCL in vitro may be a consequence of their not having access to a germinal center environment to down-regulate expression of the EBNA proteins 2, 3A, 3B, 3C, and LP (Roughan and others 2010).
EBV infection is normally kept under tight control by EBV-specific immune responses, especially by cytotoxic CD8+ T cells, which eliminate proliferating and lytically infected B cells (Hislop and others 2007; Fig. 1). In the developing world, most children become infected within the first three years of life, and EBV seropositivity reaches 100% within the first decade (Rickinson and Kieff 2001). These early primary infections are almost always asymptomatic. In contrast, in the developed world, up to half the children are still EBV seronegative at the end of their first decade and subsequently become infected through intimate oral contact in adolescence or young adulthood (Rickinson and Kieff 2001). As many as 50% of these delayed primary infections are symptomatic and manifest after an incubation period of four to seven weeks as acute infectious mononucleosis (glandular fever), an illness characterized by fever, malaise, pharyngitis, and lymphadenopathy (Vetsika and Callan 2004). During the incubation period, the cycle of infection, B cell activation, germinal center reaction, lytic replication, and reinfection initially goes unchecked by cytotoxic CD8+ T cells because of the delay in mounting an adaptive immune response. As a result, the number of latently infected memory B cells during infectious mononucleosis can rise to half and perhaps even higher of the peripheral memory B cell compartment (Hochberg and others 2004). Eventually, the infection evokes a massive expansion of activated EBV-specific CD8+ T cells, which rapidly control the infection by killing a high proportion of the EBV-infected B cells (Vetsika and Callan 2004). With the rapid decline in the EBV viral load, the number of EBV-specific CD8+ T cells also rapidly declines toward the levels found in persistently infected healthy virus carriers (Hoshino and others 1999; Hadinoto and others 2008). It has been suggested that the difference between asymptomatic primary EBV infection and infectious mononucleosis is the higher number of EBV-infected B cells in the latter, with the symptoms being due to the massive destruction of virus-infected B cells by cytotoxic CD8+ T cells (Hadinoto and others 2008). Why a higher proportion of B cells should be infected when primary infection is delayed beyond childhood to adolescence or later is unclear. Possible explanations include a higher dose of viral inoculum acquired by intimate oral contact and a reduced capacity to mount a rapid effective CD8+ T cell response in adolescents/adults compared with young children. The importance of CD8+ T cells in controlling primary EBV infection is demonstrated by the development of potentially fatal B cell lymphoproliferative disease in immunosuppressed transplant recipients with low numbers of EBV-specific T cells and high viral loads (Smets and others 2002).
Evidence Implicating EBV in the Pathogenesis of MS
The first evidence for a role of EBV in the pathogenesis of MS came in 1979 when Fraser and others (1979) reported that peripheral blood lymphocytes from patients with clinically active MS have an increased tendency toward spontaneous in vitro EBV-induced B lymphocyte transformation. In the following year, Sumaya and others (1980) reported a higher frequency of EBV seropositivity and higher serum anti-EBV antibody titers in MS patients compared with controls. Subsequent studies have shown that MS patients are almost universally seropositive for EBV but not for other viruses (Bray and others 1983; Wandinger and others 2000). In a meta-analysis of 13 case control studies comparing EBV serology in MS patients and controls, 99.5% of MS patients were EBV seropositive compared with 94.0% of controls, with EBV seronegativity having an ORMH odds ratio of MS of 0.06 (exact 95% confidence interval [CI], 0.03, 0.13; P < .000000001) (Ascherio and Munger 2007). Serial studies have shown that the risk of developing MS is extremely low among individuals not infected with EBV but increases sharply in the same individuals following EBV infection, with an estimated mean interval of 5.6 years between primary EBV infection and onset of MS (Levin and others 2010). These studies suggest that EBV infection is a prerequisite for the development of MS but, by itself, is not sufficient to cause MS because the vast majority of people infected with EBV do not develop the disease. The fact that EBV infection is associated with a dramatic increase in MS risk is obscured by the fact that EBV infects such a large proportion (~95%) of the general adult population. If EBV does have an essential role in the development of MS, studies in children should show a larger numerical difference in the frequency of EBV seropositivity between MS patients and controls because the prevalence of EBV infection in the general population is considerably lower in children than in adults. Indeed, children with MS have an EBV seropositivity rate of 86% to 99% compared with 64% to 72% in age-matched controls (Pohl and others 2006; Banwell and others 2007; Lünemann and others 2008a). One of these studies concluded that EBV infection is not essential for the development of MS because 14% of the children diagnosed as having MS were classified as EBV seronegative (Banwell and others 2007). It is possible that the EBV-seronegative cases in that study either were actually infected with EBV but falsely seronegative or did not have MS but recurrent acute disseminated encephalomyelitis, which can be difficult to distinguish clinically from MS in children. A clinical history of infectious mononucleosis increases the risk of MS, with a relative risk of 2.3 (Thacker and others 2006).
A study of blood samples collected from US military personnel before the onset of MS showed that the presence of high titers of serum IgG antibodies to the EBNA complex increases the risk 36-fold for developing MS (Levin and others 2005). This risk was associated with IgG antibodies to EBNA1 but not EBNA2. Elevated serum IgG reactivity to EBNA1 was also found to increase the risk of MS in two other prospective studies (Ascherio and others 2001; Sundström and others 2004).
When measured after the onset of MS, antibody reactivity to EBV correlates with disease activity and progression. Wandinger and others (2000) found increased IgM and IgA responses to EBV early antigens and detected EBV DNA in the sera of patients with clinically active MS but not in clinically stable patients. Subsequent studies have shown that the serum level of anti-EBNA1 IgG correlates with the number of active inflammatory demyelinating lesions in the brain, as indicated by gadolinium enhancement on MRI (Farrell and others 2009), and with the total number of T2 MRI brain lesions (Lünemann and others 2010); it also correlates with future disease progression measured clinically and by MRI (Farrell and others 2009; Lünemann and others 2010). Serum levels of IgG antibodies to EBV viral capsid antigen are also associated with T2 lesion load and gray matter atrophy (Zivadinov and others 2009; Lünemann and others 2010).
In addition to anti-EBV antibodies being increased in the serum in MS, they are also elevated in the CSF (Sumaya and others 1985; Rand and others 2000; Cepok and others 2005; Jaquiéry and others 2010), which could indicate specific synthesis of antibody against EBV or simply reflect polyspecific antibody synthesis. The corrected Antibody Index makes this distinction (Reiber and Lange 1991) and reveals specific anti-EBV antibody synthesis in the CNS (Rand and others 2000; Jaquiéry and others 2010), which is also demonstrated by the reactivity of some of the characteristic CSF oligoclonal IgG bands to EBV proteins (Rand and others 2000; Cepok and others 2005). Indeed, the anti-EBV response is the dominant antibody response in the CSF in MS, as revealed by Cepok and others (2005), who screened CSF IgG for reactivity against 37,000 proteins and found the two most frequent reactivities to be against the EBV proteins, EBNA1 and BRRF2. However, to complicate matters, there is also specific synthesis of antibodies to other viruses such as measles and rubella (Reiber and Lange 1991; Rand and others 2000). This differs from the situation in CNS diseases caused by a single infectious agent, such as subacute sclerosing panencephalitis, where there is specific synthesis in the CNS of antibody against only the causative virus, in this case the measles virus (Reiber and Lange 1991). A possible explanation for the findings in MS is that EBV is the causative infectious agent in the CNS and thus elicits a dominant anti-EBV antibody response as well as promoting the synthesis of other antibodies by infecting B cells of other specificities (Pender 2003).
Further indirect evidence that EBV is present in the CNS in MS is provided by the finding that EBV-specific CD8+ T cells are enriched in the CSF, compared with the blood, in early MS but not in patients with other inflammatory neurological diseases (Jaquiéry and others 2010). The recruitment of CD8+ T cells was selective for EBV-specific T cells because T cells reactive to cytomegalovirus, another herpesvirus, were not enriched in the CSF. Given that antigen-specific CD8+ T cells infiltrate the brain only when their cognate antigen is present (Galea and others 2007), these findings suggest that EBV is present in the CNS in MS. An alternative but less likely explanation is that EBV-specific CD8+ T cells in the CSF are recognizing not EBV but a CNS autoantigen with which they cross-react.
Recently, Serafini and others (2007) reported that a substantial proportion of the B cells and plasma cells in 21 of 22 brains of MS patients were infected with EBV. They identified EBV-infected cells using in situ hybridization for EBV-encoded small RNA (EBER-ISH), the gold standard for detection of EBV-infected B cells in histological material (Gulley and Tang 2008), and by immunohistochemistry with antibodies specific for EBV proteins. EBV-infected B cells were not detected in the brains of patients with other inflammatory CNS diseases. The major sites of EBV persistence in the MS brain were meningeal structures resembling B cell lymphoid follicles with germinal centers, which the authors had previously demonstrated to be present in patients with secondary progressive MS (Magliozzi and others 2007; Fig. 2). B cells expressing EBV latent proteins were regularly found in the brains of patients with MS, whereas B cells expressing the EBV early lytic cycle protein, BFRF1, were restricted to acute lesions and meningeal B cell follicles. Their finding that EBV-infected B cells in meningeal follicles express LMP1 and LMP2A but not EBNA2 indicates that here EBV deploys the latency II (“default”) transcription program (Serafini and others 2007; Serafini and others 2010), which allows infected B cells to progress through a germinal center reaction to become latently infected memory B cells (Thorley-Lawson and Gross 2004). It also accords with the observation that, in the tonsils of healthy EBV carriers, EBV-infected B cells expressing this viral transcription program not only are functionally indistinguishable from classical germinal center B cells but also are physically located in germinal centers (Roughan and Thorley-Lawson 2009).
Figure 2.
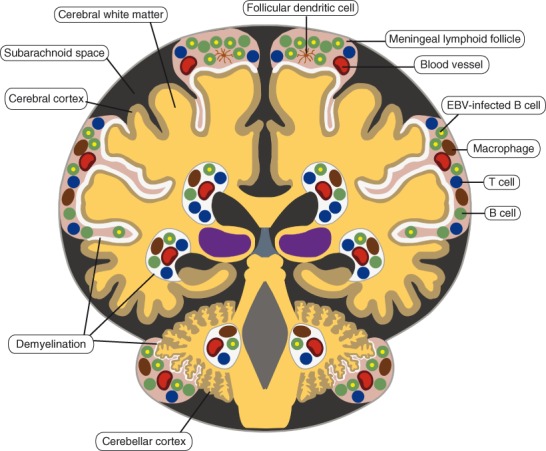
Epstein-Barr virus (EBV)–infected B cells in the brain in secondary progressive multiple sclerosis (MS). EBV-infected B cells are localized in perivascular inflammatory infiltrates in the white matter and in meningeal inflammatory infiltrates overlying the cortex, with the highest concentration of EBV-infected B cells in meningeal infiltrates that resemble lymphoid follicles with germinal centers. The white areas in the cortex and white matter represent demyelinated regions. This distribution of EBV-infected B cells in the MS brain is based on the study of Serafini and others (2007).
In contrast to Serafini and others (2007), Willis and others (2009) found EBV in only 2 of 24 brains of MS patients. They did not detect EBV-infected B cells by EBER-ISH or immunohistochemistry using antibodies against LMP1 or EBNA2 in any of the specimens but did detect low levels of EBV DNA in two samples by real-time polymerase chain reaction. They concluded that EBV infection is rare in MS brain tissue and unlikely to contribute directly to the pathogenesis. A similar conclusion was reached by Peferoen and others (2010), who detected nuclear EBER by ISH in only one specimen from 16 MS brains exhibiting prominent B cell infiltrates. Possible explanations for the discrepancy between these findings and those of Serafini and others (2007) are differences in experimental design and interpretation, tissue selection and processing, the degree of CNS B cell infiltration, and the sensitivities and specificities of the techniques used. It has been suggested that the inability of Willis and others (2009) to detect any EBV-infected B cells by EBER-ISH in their MS brain specimens might have been due to degradation of EBER RNA by long tissue fixation times in formalin (Pender 2009b). Another important difference from the work of Serafini and others (2007) is that meningeal B cell follicles, which were found by Serafini and others to be major sites of EBV persistence, were not found by Willis and others (2009) or Peferoen and others (2010) to analyze.
Lünemann and others (2010) found the EBV DNA load in peripheral blood mononuclear cells (PBMCs) to be moderately increased in patients with clinically isolated syndromes (first episodes of the type seen in MS) compared with healthy subjects. Using the same assay in an earlier study, Lünemann and others (2006) concluded that the EBV viral load is not increased in the PBMCs of patients with relapsing–remitting MS; however, the viral load was substantially higher in the MS patients than in the healthy controls, although the difference was not statistically significant, possibly because of the small numbers of subjects.
It has been noted since 1981 that the epidemiological features of MS can be explained by a requirement for a late primary EBV infection in adolescence or young adulthood (Warner and Carp 1981). Indeed, delayed primary infection with EBV can account for each of the following epidemiological characteristics of MS: the association of MS with higher socioeconomic status; the latitudinal variation in MS prevalence, with prevalence increasing with distance from the equator; the effects of migration on the risk of developing MS; and the occurrence of clusters and epidemics of MS (Haahr and Höllsberg 2006).
Hypothesis Tier 1: EBV Infection Is Essential for MS
It is important to distinguish the role of EBV from that of other infectious agents in MS. It is well established that clinical exacerbations of MS are three times more likely to occur at the time of acute systemic infection with a wide variety of viruses and bacteria than at other times (Sibley and others 1985; Correale and others 2006). EBV differs from other infectious agents in the following ways: (1) EBV infection precedes the initial onset of MS by years in contrast to other infections, which precede exacerbations of already established MS by only one to two weeks; (2) EBV persists for the lifetime of the host, whereas most other infections are eradicated after the acute phase; (3) EBV is the only agent infecting virtually all patients with MS; and (4) EBV is the only agent that infects and modulates cells (B cells) involved in the disease process. These features of EBV infection strongly suggest that it is essential for the development of MS. Possible mechanisms for the triggering of attacks by other infections include cross-reactivity between microbial antigens and CNS antigens and a general up-regulation of the immune system.
Hypothesis Tier 2: How EBV Infection Causes MS
Several hypotheses have been proposed to explain the role of EBV in the development of MS: (1) the EBV cross-reactivity hypothesis, (2) the EBV bystander damage hypothesis, (3) the αB-crystallin (mistaken self) hypothesis, and (4) the EBV-infected autoreactive B cell hypothesis, which is the hypothesis proposed in this article. For many years, the favored hypothesis was the EBV cross-reactivity hypothesis, which postulates that T cells primed by exposure to EBV antigens cross-react with and attack CNS antigens (Lang and others 2002). In support of this, 3% to 4% of EBNA1-specific CD4+ T cells in healthy subjects and MS patients react with peptides derived from myelin proteins (Lünemann and others 2008b). However, the EBV cross-reactivity hypothesis does not explain the unique and obligatory role of EBV in the development of MS. EBV is the only infectious agent known to be implicated in virtually all (>99%) cases of MS, yet all infectious agents have the potential to induce cross-reactivity with CNS antigens (Pender 2003). Nor does this hypothesis require or explain the presence of EBV-infected B cells in the brain (Serafini and others 2007) because cross-reactivity is initiated by exposure of T cells to EBV in lymphoid tissue outside the CNS. CD4+ T cell cross-reactivity between EBV and CNS antigens might contribute to the disease process in MS but is unlikely to represent the main role of EBV in causing MS (Pender 2003).
The EBV bystander damage hypothesis proposes that the immune attack on the CNS in MS is primarily directed against EBV and that this results in bystander damage to the CNS (Serafini and others 2007). In this scenario, MS would not be an autoimmune disease, although secondary autoimmune responses could develop as a result of sensitization to CNS antigens released after virus-targeted bystander damage. Thus, this hypothesis does not explain the evidence for a primary role of autoimmunity in the development of MS (Sospedra and Martin 2005; Pender and Greer 2007). It also questions why an EBV-directed immune response sufficient to cause CNS bystander damage does not eliminate EBV-infected B cells from the CNS. EBV-targeted bystander damage might contribute to the disease process in MS, but it is unlikely to constitute the main role of EBV in causing MS (Pender 2003).
The αB-crystallin or “mistaken self” hypothesis proposes that exposure to infectious agents induces the expression of αB-crystallin, a small heat-shock protein, in lymphoid cells and that the immune system mistaking self αB-crystallin for a microbial antigen generates a CD4+ T cell response, which attacks αB-crystallin derived from oligodendrocytes, with resultant inflammatory demyelination (Van Noort and others 2000). αB-crystallin is reported to be an immunodominant antigen of CNS myelin from MS patients that is expressed in oligodendrocytes and myelin in early MS lesions but not in normal white matter (Van Noort and others 2000). A key requirement of the hypothesis is that infection of the CNS by a microbial agent, not necessarily the same as that inducing αB-crystallin in lymphoid cells, up-regulates the expression of αB-crystallin in oligodendrocytes and provides other “danger” signals in the CNS, allowing inflammation to develop. Although the hypothesis is not specific for EBV, EBV is a candidate because it induces the expression of αB-crystallin in B cells, which present the protein to CD4+ T cells in a HLA-DR-restricted manner (Van Sechel and others 1999). This hypothesis by itself cannot account for the initial development and subsequent persistence of inflammation in the CNS but might explain how CD4+ T cells target oligodendrocytes and myelin after inflammation has been initiated in the CNS.
The EBV-infected autoreactive B cell hypothesis of autoimmunity published in 2003 proposes that, in genetically susceptible individuals, EBV-infected autoreactive B cells seed the target organ where they produce pathogenic autoantibodies and provide costimulatory survival signals to autoreactive T cells, which would otherwise die in the target organ by activation-induced apoptosis (Pender 2003). The probability of EBV infecting naive autoreactive B cells is not low because at least 20% of human naive B cells are autoreactive (Wardemann and others 2003). The hypothesis makes several predictions, some of which have been verified in the case of MS—namely, the presence of EBV-infected B cells in the CNS (Serafini and others 2007), although this was not confirmed by Willis and others (2009); a beneficial response to B cell depletion with rituximab (Hauser and others 2008); and decreased CD8+ T cell immunity to EBV-infected B cells (Pender and others 2009). It also explains why irradiation of the CNS fails to eradicate the production of oligoclonal IgG (Tourtellotte and others 1980) because the EBV protein BHRF1, a homologue of the antiapoptotic protein bcl2 and produced by both latently and lytically infected cells (Kelly and others 2009), inhibits radiation-induced B cell apoptosis (McCarthy and others 1996).
Based on the EBV-infected autoreactive B cell hypothesis, the following scenario is proposed to lead to the development of MS (Pender 2003; Pender and Greer 2007; Fig. 3). EBV infection in individuals genetically susceptible to autoimmunity results in a high frequency of EBV-infected memory B cells, including autoreactive B cells, which seed the CNS and produce oligoclonal IgG in the CSF. In those individuals with class II HLA types (such as DRB1*1501), which predispose to MS, common systemic infections lead to the activation of CD4+ T cells, which cross-react with CNS antigens and traffic into the CNS, where they are reactivated by EBV-infected B cells presenting CNS antigens. These EBV-infected B cells provide costimulatory survival signals to the T cells and thereby inhibit the activation-induced T cell apoptosis, which normally occurs when autoreactive T cells enter the CNS (Tabi and others 1994; Pender 1998). The autoreactive T cells orchestrate an immune attack on the CNS through the recruitment of macrophages and B cells. CNS antigens released by this attack lead to spreading of the immune response to other CNS antigens. Repeated T cell attacks on the CNS supported by local EBV-infected B cells lead to the development of meningeal B cell follicles with germinal centers, which generate CNS-reactive B cells producing autoantibodies that cause demyelination and neuronal damage in the underlying cerebral and cerebellar cortex.
Figure 3.

Proposed role of Epstein-Barr virus (EBV) infection in the development of multiple sclerosis (MS). During primary infection, EBV infects autoreactive naive B cells in the tonsil, driving them to enter germinal centers, where they proliferate intensely and differentiate into latently infected autoreactive memory B cells (step 1), which then exit from the tonsil and circulate in the blood (step 2). The number of EBV-infected B cells is normally controlled by EBV-specific cytotoxic CD8+ T cells, which kill the infected B cells, but not if there is a defect in this defense mechanism. Surviving EBV-infected autoreactive memory B cells enter the CNS, where they take up residence and produce oligoclonal IgG and pathogenic autoantibodies, which attack myelin and other components of the CNS (step 3). Autoreactive T cells that have been activated in peripheral lymphoid organs by cross-reacting infectious antigens circulate in the blood and enter the CNS, where they are reactivated by EBV-infected autoreactive B cells presenting CNS peptides (Cp) bound to major histocompatibility complex (MHC) molecules (step 4). These EBV-infected B cells provide costimulatory survival signals (B7) to the CD28 receptor on the autoreactive T cells and thereby inhibit the activation-induced T cell apoptosis that normally occurs when autoreactive T cells enter the CNS and interact with nonprofessional antigen-presenting cells such as astrocytes and microglia, which do not express B7 costimulatory molecules (Tabi and others 1994; Pender 1998) (step 6). After the autoreactive T cells have been reactivated by EBV-infected autoreactive B cells, they produce cytokines such as interleukin 2 (IL2), interferon γ (IFNγ) and tumor necrosis factor β (TNFβ) and orchestrate an autoimmune attack on the CNS with resultant oligodendrocyte and myelin destruction (step 5). TCR, T cell receptor.
Hypothesis Tier 3: Why EBV Infection Causes MS
Although virtually all MS patients are infected with EBV, only a minority of people infected with EBV develop MS. Why is this so? The EBV-infected autoreactive B cell hypothesis of autoimmunity (Pender 2003) proposes that following EBV infection, the HLA class II type will determine the organ in which autoimmune disease develops by determining which self antigens cross-react with CD4+ T cells activated by infectious agents. Thus, for MS to develop after EBV infection, MS susceptibility HLA class II alleles such as DRB1*1501 are required for infectious agents to activate CD4+ T cells that cross-react with CNS antigens and enter the CNS, where they interact with EBV-infected B cells and trigger an attack. However, there must also be another factor conferring susceptibility to MS after EBV infection because it is still only a minority of people carrying HLA-DRB1*1501 who develop MS after EBV infection. Possible candidates for this additional factor include (1) EBV strain variation, (2) a B cell abnormality increasing the uptake of EBV or enhancing the proliferative capacity of infected cells, (3) resistance of infected B cells to killing by cytotoxic CD8+ T cells, (4) an impairment in the cytotoxic function of CD8+ T cells, and (5) a quantitative deficiency of CD8+ T cells, which is the hypothesis proposed in this article.
EBV strain variation could influence susceptibility to MS in two ways. First, different strains of EBV may differ in their intrinsic biological activity, causing alterations in infectivity, B cell activation, or lytic potential, which could affect B cell homeostasis and the development of autoimmunity. Second, variant EBV strains could elicit variant antibody and T cell immune responses, leading to less protective immunity or more pathogenic immunity through cross-reactivity with CNS antigens. A recent study found that several single-nucleotide polymorphisms (SNPs) within the EBNA1 gene and one within the BRRF2 gene occurred at marginally different frequencies in EBV strains infecting MS patients versus controls (Brennan and others 2010). This suggests that EBV strain variation might influence susceptibility to MS in some patients but is unlikely to be the dominant factor influencing susceptibility in the majority of patients.
Susceptibility to MS following EBV infection could also be due to a B cell abnormality increasing the uptake of EBV or enhancing the proliferative capacity of infected cells. At present, there is no evidence for such abnormalities; however, this area has received little attention except for one study that found no differences between MS patients and controls in the frequencies of SNPs within the gene for CR2, the primary receptor for EBV on B cells (Simon and others 2007).
Resistance of EBV-infected B cells to killing by cytotoxic CD8+ T cells is unlikely to underlie the susceptibility to MS because EBV-infected LCL from MS patients can be killed normally by HLA-matched EBV-specific cytotoxic CD8+ T cell clones from healthy subjects (Pender and others 2009). An impairment in the cytotoxic function of CD8+ T cells is also unlikely because MS patients can generate EBV-specific cytotoxic CD8+ T cell lines in vitro, which can kill their own EBV-infected LCL (Pender and others 2009).
A quantitative deficiency of CD8+ T cells is the most likely factor underlying the susceptibility to develop MS after EBV infection. MS patients have decreased CD8+ T cell reactivity to EBV-infected B cells, as assessed by the frequency of PBMC producing interferon-g in response to autologous LCL in enzyme-linked immunospot (ELISPOT) assays (Pender and others 2009). In contrast to the use of synthetic EBV peptides as target antigens (Jilek and others 2008), this approach provides a direct measure of the aggregate T cell response to EBV-infected B cells in each subject because it uses each person’s natural antigen-processing mechanisms to present viral antigens at normal physiological concentrations on the surface of their own EBV-infected B cells, and it represents the total T cell response to all EBV antigens presented by all HLA molecules on LCL in each subject. LCL express not only the latent proteins of EBV but also the lytic proteins (Pudney and others 2005; Tynan and others 2005) because a proportion of the cells in LCL are in the lytic phase of infection. Using a pool of HLA class I–restricted EBV peptides in ELISPOT assays, Jilek and others (2008) found an increased EBV-specific CD8+ T cell response in patients with clinically isolated syndromes but a normal response in established MS. These results are difficult to interpret because it is likely that in any given subject, only a minority of the peptides were restricted by HLA class I alleles carried by the subject. Thus, the T cell response to the pool of peptides might have been composed largely of responses that have no in vivo relevance. It is noteworthy that the mean frequency of T cells responding to the pool of peptides in healthy subjects in this study (Jilek and others 2008) was only 5% of the mean frequency of LCL-specific T cells in the study analyzing the total T cell response to EBV-infected B cells (Pender and others 2009). Using a similar assay to that of Jilek and others (2008), Lünemann and others (2010) found no significant difference in the CD8+ T cell response to pooled EBV peptides between patients with clinically isolated syndromes and healthy controls.
The finding of decreased CD8+ T cell reactivity to EBV-infected B cells in patients with MS (Pender and others 2009) is consistent with earlier reports that MS patients have decreased T cell control of Ig-secreting B cells after in vitro infection with EBV (Craig and others 1988) and that peripheral blood lymphocytes from patients with MS have an increased tendency toward spontaneous in vitro EBV-induced B cell transformation (Fraser and others 1979). A key question is whether the decreased frequency of LCL-specific CD8+ T cells in the blood is the cause or the effect of the accumulation of EBV-infected B cells in the CNS (Serafini and others 2007) or a combination of both. Some of the decrease might be due to sequestration of EBV-specific CD8+ T cells in the CNS because CD8+ T cells are present in the CNS in proportion to the number of EBV-infected B cells (Serafini and others 2007). If the decreased frequency of LCL-specific CD8+ T cells in the blood were to be attributed solely to sequestration within the CNS, it would raise the question why EBV-infected B cells accumulate in the CNS in the first place and are not eliminated by EBV-specific CD8+ T cells, given that EBV-infected B cells of MS patients can be killed by their own EBV-specific CD8+ T cells in vitro (Pender and others 2009). The most likely scenario is that a deficiency of EBV-specific CD8+ T cells allows the accumulation of EBV-infected B cells in the CNS in MS and that a vicious circle then ensues whereby the inherently deficient CD8+ T cell response is further compromised by T cell exhaustion as a result of the persistent high EBV load in the CNS. Interestingly, lower LCL-specific T cell frequencies were associated with earlier age of onset of MS, suggesting that the more severe the deficiency of EBV-specific CD8+ T cells, the sooner MS will develop after primary EBV infection (Pender and others 2009).
If a quantitative deficiency of EBV-specific CD8+ T cells is responsible for the accumulation of EBV-infected B cells in the CNS, what causes this deficiency? Since 1980, it has been recognized that MS patients have a decreased proportion and number of CD8+ T cells and increased CD4/CD8 T cell ratio in peripheral blood (Reinherz and others 1980; Bach and others 1980; Antel and others 1984; Thompson and others 1986; Trotter and others 1989; Kreuzfelder and others 1992; Michałowska-Wender and Wender 2006). This was initially interpreted as a decrease in suppressor T cells leading to disinhibition of autoimmune responses (Reinherz and others 1980) but later attributed to sequestration of CD8+ T cells in the CNS (Kreuzfelder and others 1992) because CD8+ T cells predominate in the T cell infiltrate in the MS brain (Booss and others 1983; Hauser and others 1986). However, if CD8+ T cells are accumulating in the CNS because of the presence there of EBV, the number of CD8+ T cells in the blood should increase, not decrease, because normally the CD8+ T cell response increases with EBV load (Hoshino and others 1999; Yajima and others 2008; Strowig and others 2009). An alternative explanation, and the one proposed here, is that a genetic deficiency of CD8+ T cells results in a decreased CD8+ T cell response to EBV, which allows EBV-infected B cells to accumulate in the CNS. The CD4/CD8 T cell ratio in humans is genetically controlled (Amadori and others 1995), with at least some of the responsible genes being located in the HLA complex (Ferreira and others 2010). A genetic deficiency of CD8+ T cells would exert its effect from the time of primary EBV infection in the tonsil by prolonging the survival and proliferation of infected B lymphoblasts and germinal center cells and the survival of plasma cells replicating the virus. This would facilitate the development of infectious mononucleosis and increase the probability of clonal expansion of EBV-infected autoreactive B cells. In individuals with HLA class II backgrounds conducive to MS, continuing CD8+ T cell deficiency would allow the accumulation of EBV-infected autoreactive B cells in the CNS and ultimately the recruitment of autoreactive CD4+ T cells and autoimmune attack, as described above (Fig. 4). In individuals with HLA class II backgrounds conducive to other autoimmune diseases, EBV-infected autoreactive B cells would accumulate in the respective target organ and reactivate autoreactive T cells there with resultant autoimmune attack (Fig. 4). In MS, the autoimmune process itself could foster the survival and proliferation of EBV-infected autoreactive B cells in the CNS by releasing CNS antigens and giving CD4+ T cell help, which would complement the BCR and CD40 receptor signaling already provided by LMP2A and LMP1, respectively (Pender 2009b). This could lead to a vicious circle wherein EBV-infected autoreactive B cells promote autoimmunity, which in turn promotes EBV infection in the CNS. Because the number of CD8+ T cells normally declines with increasing age (Amadori and others 1995; Hall and others 2000), the primary CD8+ T cell deficiency will be aggravated as each patient ages (Fig. 5). This could underlie the age-dependent accumulation of disability in MS (Confavreux and Vukusic 2006). In the later stages of MS, the prolonged high EBV load could lead to T cell exhaustion, also aggravating the primary CD8+ T cell deficiency.
Figure 4.

Proposed sequence of events leading to the development of multiple sclerosis (MS) and other human autoimmune diseases, for example, autoimmune thyroid disease. In individuals with a genetic deficiency of CD8+ T cells (carried by “autoimmune genes”) and with HLA class II genes predisposing to MS (such as HLA-DR15), late Epstein-Barr virus (EBV) infection leads to the infection of autoreactive B cells, which accumulate in the CNS, where they reactivate autoreactive T cells that orchestrate an autoimmune attack on the CNS leading to the development of MS (upper panel). In individuals with a genetic deficiency of CD8+ T cells and with HLA class II genes predisposing to autoimmune thyroid disease (such as HLA-DR3), late EBV infection leads to the infection of autoreactive B cells, which accumulate in the thyroid gland, where they reactivate autoreactive T cells that orchestrate an autoimmune attack on the thyroid gland, leading to the development of autoimmune thyroid disease (lower panel).
Figure 5.

Proposed genetic deficiency of CD8+ T cells underlying the development of multiple sclerosis (MS). The upper green panel on the graph represents health, the middle orange panel the development of relapsing MS, and the lower red panel the development of progressive MS. In healthy individuals (Health), the number of CD8+ T cells declines with increasing age but still remains sufficient to control Epstein-Barr virus (EBV) infection. In individuals with a mild genetic deficiency of CD8+ T cells, the deficiency of CD8+ T cells is aggravated by increasing age, eventually leading to insufficient CD8+ T cells to control EBV infection. In individuals carrying HLA class II genes predisposing to MS, this leads to the accumulation of EBV-infected B cells in the CNS and the development of relapsing MS, which ultimately evolves into progressive MS as the CD8+ T cell count further declines with age and as the EBV load in the CNS subsequently increases. In individuals with a severe genetic deficiency of CD8+ T cells, MS develops at a younger age and progresses more rapidly. Lower exposure to sunlight at higher latitudes aggravates the genetic CD8+ T cell deficiency and increases the incidence and progression of MS.
Hypothesis Tier 4: How Sunlight Protects against MS
It has long been recognized that the prevalence of MS increases with increasing latitude (Ulett 1948). Because sunlight exposure decreases with increasing latitude, it has been proposed that sunlight protects against the development of MS (Acheson and others 1960). Subsequently, considering that the ultraviolet component of sunlight is essential for the synthesis of vitamin D in the skin, Goldberg (1974) proposed that vitamin D protects against MS. Support for this comes from prospective epidemiological studies showing that higher vitamin D intake (Munger and others 2004) and higher serum levels of 25-hydroxyvitamin D (Munger and others 2006) are associated with a reduced risk of developing MS. Because vitamin D prevents the induction of experimental autoimmune encephalomyelitis (EAE) in mice (Lemire and Archer 1991), the most favored hypothesis has been that vitamin D protects against MS by suppressing autoreactive T cell responses. However, this is unlikely because the levels of 1,25-dihydroxyvitamin D required to suppress EAE are well above those that can be produced naturally upon exposure to sunlight (Becklund and others 2010). More recently, it has been proposed that vitamin D protects against MS by modulating the immune response to EBV, either by inhibiting the activation of autoreactive T cells (Holmøy 2008) or through enhanced protective innate immunity mediated by cathelicidin (Grant 2008).
Another possible EBV-related mechanism whereby sunlight/vitamin D protects against MS, and the one proposed here, is a sunlight-induced increase in the number of CD8+ T cells available to control EBV infection (Figure 5). Exposure to natural sunlight or treatment in a solarium increases the proportion of CD8+ T cells and decreases the CD4/CD8 T cell ratio in peripheral blood (Hersey and others 1983a; Hersey and others 1983b; Falkenbach and Sedlmeyer 1997). Exactly how sunlight increases the number of CD8+ T cells is unclear, but the effect is probably mediated at least in part by vitamin D because (1) among cells of the immune system, activated CD8+ T cells express the highest concentrations of the vitamin D receptor (Veldman and others 2000); (2) vitamin D increases the mitogen-induced proliferation of CD8+ T cells and decreases the CD4/CD8 ratio in bovine PBMC in vitro (Nonnecke and others 1993); (3) vitamin D administration increases the CD8+ T cell count (Žofková and Kancheva 1997); and (4) vitamin D deficiency is associated with a decreased proportion of CD8+ T cells and increased CD4/CD8 ratio (Çakmak and others 1999). Thus, it is proposed that the increasing incidence of MS with increasing latitude is due to aggravation of genetic CD8+ T cell deficiency, with resulting poorer control of EBV infection. A higher frequency of late primary EBV infection at a higher latitude might also contribute to the latitudinal gradient (Haahr and Höllsberg 2006).
Testing the Hypothesis
Is EBV Infection Essential for the Development of MS?
If EBV infection is essential for the development of MS, it should be possible to prevent and cure MS by controlling EBV infection (Pender 2009a).
Prevention
Vaccination of healthy EBV-seronegative young adults with recombinant gp350 is effective in preventing the development of infectious mononucleosis induced by EBV infection, although it does not prevent asymptomatic EBV infection (Sokal and others 2007). The vaccinated subjects showed seroconversion to anti-gp350 antibodies, which persisted >18 months and are probably responsible for the protective effect because anti-gp350 antibody neutralizes EBV infectivity (Haque and others 2006). Given that infectious mononucleosis increases the risk of MS (Thacker and others 2006), vaccination with gp350 might decrease the incidence of MS by reducing the incidence of infectious mononucleosis. By reducing the infectivity of EBV, it might prevent MS also in individuals who would not have developed infectious mononucleosis after infection with EBV.
Treatment
There are three ways to treat MS by controlling EBV infection: (1) B cell depletion with monoclonal antibodies, (2) boosting immunity to EBV, and (3) antiviral drugs. B cell depletion with rituximab reduces inflammatory brain lesions and clinical relapses in patients with relapsing–remitting MS (Hauser and others 2008) but has the disadvantage of eliminating not only EBV-infected B cells but also uninfected B cells, which normally confer protective immunity against infectious agents. The effect of rituximab is consistent with an essential role of EBV in the development of MS but does not constitute proof because it could be beneficial by eliminating autoreactive B cells not infected by EBV. More convincing evidence for an essential role of EBV would be eradication of disease by boosting immunity to EBV or by antiviral drugs. In people who have already been infected with EBV and have subsequently developed MS, boosting immunity to EBV could be achieved by vaccination with gp350, administration of humanized or human monoclonal antibody against gp350, or by the intravenous infusion of autologous EBV-specific cytotoxic CD8+ T cells after expansion in vitro, which controls EBV-induced posttransplant lymphoproliferative disease (Savoldo and others 2006). With regard to antiviral drugs, treatment with aciclovir, which inhibits herpesvirus DNA polymerase, 2.4 g daily for two years reduced the clinical relapse rate by 34% in patients with relapsing–remitting MS (P = .08; Lycke and others 1996). In a subsequent study, treatment with valaciclovir, a prodrug of aciclovir that increases aciclovir bioavailability, 3 g daily for 24 weeks reduced the number of new active MRI brain lesions in a subgroup of patients with relapsing–remitting MS who had high levels of MRI-evident disease activity (Bech and others 2002). The limited efficacy of aciclovir and valaciclovir in the treatment of MS might be explained by the fact that these drugs act on EBV only when it is using its own DNA polymerase to replicate its DNA. This will apply only to lytically infected cells but not to latently infected cells, which replicate EBV DNA through the use of EBNA1 to engage host cell DNA polymerase. Thus, these drugs will inhibit EBV in only some of the EBV-infected B cells in the MS brain where some cells express BFRF1 (indicative of lytic infection) but others express EBNA2 and/or LMP1 (indicative of latency; Serafini and others 2007). One strategy to overcome this would be first to administer rituximab to eliminate as many EBV-infected B cells as possible and to follow this with long-term antiviral drug therapy. An alternative approach is to target LMP1 (Mei and others 2006), LMP2A, or EBNA1 (Li and others 2010) to inhibit EBV in latently infected cells. If EBV infection of B cells in the CNS underpins the development of MS, effective antiviral drugs have the potential to cure MS.
Are EBV-Infected B Cells in the CNS Autoreactive?
Whether EBV-infected B cells in the CNS are autoreactive could be addressed by determining whether they bind biotinylated CNS antigens in the same way that intrathyroidal germinal center B cells specifically bind thyroid antigens in autoimmune thyroid disease (Armengol and others 2001).
Does a Deficiency of CD8+ T Cells Underlie the Development of MS following EBV Infection?
Whether a deficiency of CD8+ T cells underlies the development of MS following EBV infection could be addressed by the following experiments: (1) prospective studies to determine whether CD8+ T cell deficiency precedes the development of MS; (2) determining whether genetic variants associated with a decreased number of CD8+ T cells, such as the A allele of rs2524054 in HLA-B (Ferreira and others 2010), predispose to MS; and (3) examining whether MS can be successfully treated by the intravenous infusion of autologous EBV-specific cytotoxic CD8+ T cells after expansion in vitro.
Does Vitamin D Deficiency Contribute to the Development of MS by Depleting CD8+ T Cells?
Whether vitamin D deficiency contributes to the development of MS by depleting CD8+ T cells could be tested by determining whether treatment of vitamin D deficiency in MS patients by dietary supplementation or exposure to sunlight increases the CD8+ T cell response to EBV, decreases the EBV load, and produces clinical improvement.
Acknowledgments
The author thanks Mr Stuart Reid for assistance in preparing the illustrations.
Footnotes
The author(s) declared no potential conflicts of interest with respect to the authorship and/or publication of this article.
The author(s) disclosed receipt of the following financial support for the research and/or authorship of this article: My research in this field has been supported by Multiple Sclerosis Research Australia.
References
- Acheson ED, Bachrach CA, Wright FM. 1960. Some comments on the relationship of the distribution of multiple sclerosis to latitude, solar radiation, and other variables. Acta Psychiatr Scand Suppl 35:132–47 [DOI] [PubMed] [Google Scholar]
- Amadori A, Zamarchi R, De Silvestro G, Forza G, Cavatton G, Danieli GA. and others. 1995. Genetic control of the CD4/CD8 T-cell ratio in humans. Nat Med 1:1279–83 [DOI] [PubMed] [Google Scholar]
- Antel JP, Peeples DM, Reder AT, Arnason BGW. 1984. Analysis of T regulator cell surface markers and functional properties in multiple sclerosis. J Neuroimmunol 6:93–103 [DOI] [PubMed] [Google Scholar]
- Armengol MP, Juan M, Lucas-Martín A, Fernández-Figueras MT, Jaraquemada D, Gallart T. and others. 2001. Thyroid autoimmune disease: demonstration of thyroid antigen-specific B cells and recombination-activating gene expression in chemokine-containing active intrathyroidal germinal centers. Am J Pathol 159:861–73 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ascherio A, Munger KL. 2007. Environmental risk factors for multiple sclerosis: Part I. The role of infection. Ann Neurol 61:288–99 [DOI] [PubMed] [Google Scholar]
- Ascherio A, Munger KL, Lennette ET, Spiegelman D, Hernán MA, Olek MJ. and others. 2001. Epstein-Barr virus antibodies and risk of multiple sclerosis: a prospective study. JAMA 286:3083–8 [DOI] [PubMed] [Google Scholar]
- Bach M-A, Phan-Dinh-Tuy F, Tournier E, Chatenoud L, Bach J-F, Martin C. and others. 1980. Deficit of suppressor T cells in active multiple sclerosis. Lancet 316:1221–3 [DOI] [PubMed] [Google Scholar]
- Banwell B, Krupp L, Kennedy J, Tellier R, Tenembaum S, Ness J. and others. 2007. Clinical features and viral serologies in children with multiple sclerosis: a multinational observational study. Lancet Neurol 6:773–81 [DOI] [PubMed] [Google Scholar]
- Bech E, Lycke J, Gadeberg P, Hansen HJ, Malmeström C, Andersen O. and others. 2002. A randomized, double-blind, placebo-controlled MRI study of anti-herpes virus therapy in MS. Neurology 58:31–6 [DOI] [PubMed] [Google Scholar]
- Becklund BR, Severson KS, Vang SV, DeLuca HF. 2010. UV radiation suppresses experimental autoimmune encephalomyelitis independent of vitamin D production. Proc Natl Acad Sci U S A 107:6418–23 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Booss J, Esiri MM, Tourtellotte WW, Mason DY. 1983. Immunohistological analysis of T lymphocyte subsets in the central nervous system in chronic progressive multiple sclerosis. J Neurol Sci 62:219–32 [DOI] [PubMed] [Google Scholar]
- Bray PF, Bloomer LC, Salmon VC, Bagley MH, Larsen PD. 1983. Epstein-Barr virus infection and antibody synthesis in patients with multiple sclerosis. Arch Neurol 40:406–8 [DOI] [PubMed] [Google Scholar]
- Brennan RM, Burrows JM, Bell MJ, Bromham L, Csurhes PA, Lenarczyk A. and others. 2010. Strains of Epstein-Barr virus infecting multiple sclerosis patients. Multiple Sclerosis 16:643–51 [DOI] [PubMed] [Google Scholar]
- Çakmak FN, Erol M, Ergül P, Yalçýner A. 1999. T lymphocytes and vitamins. J Pediatr 135:531. [DOI] [PubMed] [Google Scholar]
- Cepok S, Zhou D, Srivastava R, Nessler S, Stei S, Büssow K. and others. 2005. Identification of Epstein-Barr virus proteins as putative targets of the immune response in multiple sclerosis. J Clin Invest 115:1352–60 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Confavreux C, Vukusic S. 2006. Age at disability milestones in multiple sclerosis. Brain 129:595–605 [DOI] [PubMed] [Google Scholar]
- Correale J, Fiol M, Gilmore W. 2006. The risk of relapses in multiple sclerosis during systemic infections. Neurology 67:652–9 [DOI] [PubMed] [Google Scholar]
- Craig JC, Haire M, Merrett JD. 1988. T-cell-mediated suppression of Epstein-Barr virus-induced B lymphocyte activation in multiple sclerosis. Clin Immunol Immunopathol 48:253–60 [DOI] [PubMed] [Google Scholar]
- Falkenbach A, Sedlmeyer A. 1997. Travel to sunny countries is associated with changes in immunological parameters. Photodermatol Photoimmunol Photomed 13:139–42 [DOI] [PubMed] [Google Scholar]
- Farrell RA, Antony D, Wall GR, Clark DA, Fisniku L, Swanton J. and others. 2009. Humoral immune response to EBV in multiple sclerosis is associated with disease activity on MRI. Neurology 73:32–8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferreira MAR, Mangino M, Brumme CJ, Zhao ZZ, Medland SE, Wright MJ. and others. 2010. Quantitative trait loci for CD4:CD8 lymphocyte ratio are associated with risk of type 1 diabetes and HIV-1 immune control. Am J Hum Genet 86:88–92 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fraser KB, Millar JHD, Haire M, McCrea S. 1979. Increased tendency to spontaneous in-vitro lymphocyte transformation in clinically active multiple sclerosis. Lancet 314:715–7 [PubMed] [Google Scholar]
- Galea I, Bernardes-Silva M, Forse PA, van Rooijen N, Liblau RS, Perry VH. 2007. An antigen-specific pathway for CD8 T cells across the blood-brain barrier. J Exp Med 204:2023–30 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldberg P. 1974. Multiple sclerosis: vitamin D and calcium as environmental determinants of prevalence (a viewpoint): Part 1. Sunlight, dietary factors and epidemiology. Int J Environ Stud 6:19–27 [Google Scholar]
- Grant WB. 2008. Hypothesis—ultraviolet-B irradiance and vitamin D reduce the risk of viral infections and thus their sequelae, including autoimmune diseases and some cancers. Photochem Photobiol 84:356–65 [DOI] [PubMed] [Google Scholar]
- Gulley ML, Tang W. 2008. Laboratory assays for Epstein-Barr virus-related disease. J Mol Diagn 10:279–92 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haahr S, Höllsberg P. 2006. Multiple sclerosis is linked to Epstein-Barr virus infection. Rev Med Virol 16:297–310 [DOI] [PubMed] [Google Scholar]
- Hadinoto V, Shapiro M, Greenough TC, Sullivan JL, Luzuriaga K, Thorley-Lawson DA. 2008. On the dynamics of acute EBV infection and the pathogenesis of infectious mononucleosis. Blood 111:1420–7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hadinoto V, Shapiro M, Sun CC, Thorley-Lawson DA. 2009. The dynamics of EBV shedding implicate a central role for epithelial cells in amplifying viral output. PLoS Pathog 5:e1000496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hall MA, Ahmadi KR, Norman P, Snieder H, MacGregor AJ, Vaughan RW. and others. 2000. Genetic influence on peripheral blood T lymphocyte levels. Genes Immun 1:423–7 [DOI] [PubMed] [Google Scholar]
- Haque T, Johannessen I, Dombagoda D, Sengupta C, Burns DM, Bird P. and others. 2006. A mouse monoclonal antibody against Epstein-Barr virus envelope glycoprotein 350 prevents infection both in vitro and in vivo. J Infect Dis 194:584–7 [DOI] [PubMed] [Google Scholar]
- Hauser SL, Bhan AK, Gilles F, Kemp M, Kerr C, Weiner HL. 1986. Immunohistochemical analysis of the cellular infiltrate in multiple sclerosis lesions. Ann Neurol 19:578–87 [DOI] [PubMed] [Google Scholar]
- Hauser SL, Waubant E, Arnold DL, Vollmer T, Antel J, Fox RJ. and others. 2008. B-cell depletion with rituximab in relapsing-remitting multiple sclerosis. N Engl J Med 358:676–88 [DOI] [PubMed] [Google Scholar]
- Hersey P, Bradley M, Hasic E, Haran G, Edwards A, McCarthy WH. 1983a. Immunological effects of solarium exposure. Lancet 321:545–8 [DOI] [PubMed] [Google Scholar]
- Hersey P, Haran G, Hasic E, Edwards A. 1983b. Alteration of T cell subsets and induction of suppressor T cell activity in normal subjects after exposure to sunlight. J Immunol 31:171–4 [PubMed] [Google Scholar]
- Hislop AD, Taylor GS, Sauce D, Rickinson AB. 2007. Cellular responses to viral infection in humans: lessons from Epstein-Barr virus. Annu Rev Immunol 25:587–617 [DOI] [PubMed] [Google Scholar]
- Hochberg D, Souza T, Catalina M, Sullivan JL, Luzuriaga K, Thorley-Lawson DA. 2004. Acute infection with Epstein-Barr virus targets and overwhelms the peripheral memory B-cell compartment with resting, latently infected cells. J Virol 78:5194–204 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holmøy T. 2008. Vitamin D status modulates the immune response to Epstein Barr virus: synergistic effect of risk factors in multiple sclerosis. Med Hypotheses 70:66–9 [DOI] [PubMed] [Google Scholar]
- Hoshino Y, Morishima T, Kimura H, Nishikawa K, Tsurumi T, Kuzushima K. 1999. Antigen-driven expansion and contraction of CD8+-activated T cells in primary EBV infection. J Immunol 163:5735–40 [PubMed] [Google Scholar]
- Jaquiéry E, Jilek S, Schluep M, Meylan P, Lysandropoulos A, Pantaleo G. and others. 2010. Intrathecal immune responses to EBV in early MS. Eur J Immunol 40:878–87 [DOI] [PubMed] [Google Scholar]
- Jilek S, Schluep M, Meylan P, Vingerhoets F, Guignard L, Monney A. and others. 2008. Strong EBV-specific CD8+ T-cell response in patients with early multiple sclerosis. Brain 131:1712–21 [DOI] [PubMed] [Google Scholar]
- Kelly GL, Long HM, Stylianou J, Thomas WA, Leese A, Bell AI. and others. 2009. An Epstein-Barr virus anti-apoptotic protein constitutively expressed in transformed cells and implicated in Burkitt lymphomagenesis: the Wp/BHRF1 link. PLoS Pathog 5:e1000341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kreuzfelder E, Shen G, Bittorf M, Scheiermann N, Thraenhart O, Seidel D. and others. 1992. Enumeration of T, B and natural killer peripheral blood cells of patients with multiple sclerosis and controls. Eur Neurol 32:190–4 [DOI] [PubMed] [Google Scholar]
- Laichalk LL, Thorley-Lawson DA. 2005. Terminal differentiation into plasma cells initiates the replicative cycle of Epstein-Barr virus in vivo. J Virol 79:1296–307 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lang HL, Jacobsen H, Ikemizu S, Andersson C, Harlos K, Madsen L. and others. 2002. A functional and structural basis for TCR cross-reactivity in multiple sclerosis. Nat Immunol 3:940–3 [DOI] [PubMed] [Google Scholar]
- Lemire JM, Archer DC. 1991. 1,25-Dihydroxyvitamin D3 prevents the in vivo induction of murine experimental autoimmune encephalomyelitis. J Clin Invest 87:1103–7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levin LI, Munger KL, O’Reilly EJ, Falk KI, Ascherio A. 2010. Primary infection with the Epstein-Barr virus and risk of multiple sclerosis. Ann Neurol 67:824–30 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levin LI, Munger KL, Rubertone MV, Peck CA, Lennette ET, Spiegelman D. and others. 2005. Temporal relationship between elevation of Epstein-Barr virus antibody titers and initial onset of neurological symptoms in multiple sclerosis. JAMA 293:2496–500 [DOI] [PubMed] [Google Scholar]
- Li N, Thompson S, Schultz DC, Zhu W, Jiang H, Luo C. and others. 2010. Discovery of selective inhibitors against EBNA1 via high throughput in silico virtual screening. PLoS One 5:e10126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lünemann JD, Edwards N, Muraro PA, Hayashi S, Cohen JI, Münz C. and others. 2006. Increased frequency and broadened specificity of latent EBV nuclear antigen-1-specific T cells in multiple sclerosis. Brain 129:1493–506 [DOI] [PubMed] [Google Scholar]
- Lünemann JD, Huppke P, Roberts S, Brück W, Gärtner J, Münz C. 2008a. Broadened and elevated humoral immune response to EBNA1 in pediatric multiple sclerosis. Neurology 71:1033–5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lünemann JD, Jelčić I, Roberts S, Lutterotti A, Tackenberg B, Martin R. and others. 2008b. EBNA1-specific T cells from patients with multiple sclerosis cross react with myelin antigens and co-produce IFN-g and IL-2. J Exp Med 205:1763–73 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lünemann JD, Tintoré M, Messmer B, Strowig T, Rovira A, Perkal H. and others. 2010. Elevated Epstein-Barr virus-encoded nuclear antigen-1 immune responses predict conversion to multiple sclerosis. Ann Neurol 67:159–69 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lycke J, Svennerholm B, Hjelmquist E, Frisen L, Badr G, Andersson M. and others. 1996. Acyclovir treatment of relapsing-remitting multiple sclerosis: a randomized, placebo-controlled, double-blind study. J Neurol 243:214–24 [DOI] [PubMed] [Google Scholar]
- Magliozzi R, Howell O, Vora A, Serafini B, Nicholas R, Puopolo M. and others. 2007. Meningeal B-cell follicles in secondary progressive multiple sclerosis associate with early onset of disease and severe cortical pathology. Brain 130:1089–104 [DOI] [PubMed] [Google Scholar]
- Mancao C, Hammerschmidt W. 2007. Epstein-Barr virus latent membrane protein 2A is a B-cell receptor mimic and essential for B-cell survival. Blood 110:3715–21 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCarthy NJ, Hazlewood SA, Huen DS, Rickinson AB, Williams GT. 1996. The Epstein-Barr virus gene BHRF1, a homologue of the cellular oncogene Bcl-2, inhibits apoptosis induced by gamma radiation and chemotherapeutic drugs. Adv Exp Med Biol 406:83–97 [DOI] [PubMed] [Google Scholar]
- Mei YP, Zhu XF, Zhou JM, Huang H, Deng R, Zeng YX. 2006. siRNA targeting LMP1-induced apoptosis in EBV-positive lymphoma cells is associated with inhibition of telomerase activity and expression. Cancer Lett 232:189–98 [DOI] [PubMed] [Google Scholar]
- Michałowska-Wender G, Wender M. 2006. Mononuclear subsets in the peripheral blood of multiple sclerosis patients in relation to results of brain gadolinium-enhancing imaging. Folia Neuropathol 44:67–71 [PubMed] [Google Scholar]
- Munger KL, Levin LI, Hollis BW, Howard NS, Ascherio A. 2006. Serum 25-hydroxyvitamin D levels and risk of multiple sclerosis. JAMA 296:2832–8 [DOI] [PubMed] [Google Scholar]
- Munger KL, Zhang SM, O’Reilly E, Hernán MA, Olek MJ, Willett WC. and others. 2004. Vitamin D intake and incidence of multiple sclerosis. Neurology 62:60–5 [DOI] [PubMed] [Google Scholar]
- Nemerow GR, Mold C, Schwend VK, Tollefson V, Cooper NR. 1987. Identification of gp350 as the viral glycoprotein mediating attachment of Epstein-Barr virus (EBV) to the EBV/C3d receptor of B cells: sequence homology of gp350 and C3 complement fragment C3d. J Virol 61:1416–20 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nonnecke BJ, Franklin ST, Reinhardt TA, Horst RL. 1993. In vitro modulation of proliferation and phenotype of resting and mitogen-stimulated bovine mononuclear leukocytes by 1,25-dihydroxyvitamin D3. Vet Immunol Immunopathol 38:75–89 [DOI] [PubMed] [Google Scholar]
- Peferoen LAN, Lamers F, Lodder LNR, Gerritsen WH, Huitinga I, Melief J. and others. 2010. Epstein Barr virus is not a characteristic feature in the central nervous system in established multiple sclerosis. Brain 133:e137. [DOI] [PubMed] [Google Scholar]
- Pender MP. 1998. Genetically determined failure of activation-induced apoptosis of autoreactive T cells as a cause of multiple sclerosis. Lancet 351:978–81 [DOI] [PubMed] [Google Scholar]
- Pender MP. 2003. Infection of autoreactive B lymphocytes with EBV, causing chronic autoimmune diseases. Trends Immunol 24:584–8 [DOI] [PubMed] [Google Scholar]
- Pender MP. 2009a. Preventing and curing multiple sclerosis by controlling Epstein-Barr virus infection. Autoimmun Rev 8:563–568 [DOI] [PubMed] [Google Scholar]
- Pender MP. 2009b. Does Epstein-Barr virus infection in the brain drive the development of multiple sclerosis? Brain 132:3196–8 [DOI] [PubMed] [Google Scholar]
- Pender MP, Csurhes PA, Lenarczyk A, Pfluger CMM, Burrows SR. 2009. Decreased T cell reactivity to Epstein-Barr virus infected lymphoblastoid cell lines in multiple sclerosis. J Neurol Neurosurg Psychiatry 80:498–505 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pender MP, Greer JM. 2007. Immunology of multiple sclerosis. Curr Allergy Asthma Rep 7:285–92 [DOI] [PubMed] [Google Scholar]
- Pohl D, Krone B, Rostasy K, Kahler E, Brunner E, Lehnert M. and others. 2006. High seroprevalence of Epstein-Barr virus in children with multiple sclerosis. Neurology 67:2063–5 [DOI] [PubMed] [Google Scholar]
- Pudney VA, Leese AM, Rickinson AB, Hislop AD. 2005. CD8+ immunodominance among Epstein-Barr virus lytic cycle antigens directly reflects the efficiency of antigen presentation in lytically infected cells. J Exp Med 201:349–60 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rand KH, Houck H, Denslow ND, Heilman KM. 2000. Epstein-Barr virus nuclear antigen-1 (EBNA-1) associated oligoclonal bands in patients with multiple sclerosis. J Neurol Sci 173:32–9 [DOI] [PubMed] [Google Scholar]
- Rastelli J, Hömig-Hölzel C, Seagal J, Müller W, Hermann AC, Rajewsky K. and others. 2008. LMP1 signaling can replace CD40 signaling in B cells in vivo and has unique features of inducing class-switch recombination to IgG1. Blood 111:1448–55 [DOI] [PubMed] [Google Scholar]
- Reiber H, Lange P. 1991. Quantification of virus-specific antibodies in cerebrospinal fluid and serum: sensitive and specific detection of antibody synthesis in brain. Clin Chem 37:1153–60 [PubMed] [Google Scholar]
- Reinherz EL, Weiner HL, Hauser SL, Cohen JA, Distaso JA, Schlossman SF. 1980. Loss of suppressor T cells in active multiple sclerosis: analysis with monoclonal antibodies. N Engl J Med 303:125–9 [DOI] [PubMed] [Google Scholar]
- Rickinson AB, Kieff E. 2001. Epstein-Barr virus. In: Knipe DM, Howley PM. editors. Fields Virology. Philadelphia: Lippincott Williams & Wilkins; p 2575–627 [Google Scholar]
- Roughan JE, Thorley-Lawson DA. 2009. The intersection of Epstein-Barr virus with the germinal center. J Virol 83:3968–76 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roughan JE, Torgbor C, Thorley-Lawson DA. 2010. Germinal center B cells latently infected with Epstein-Barr virus proliferate extensively but do not increase in number. J Virol 84:1158–68 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Savoldo B, Goss JA, Hammer MM, Zhang L, Lopez T, Gee AP. and others. 2006. Treatment of solid organ transplant recipients with autologous Epstein Barr virus-specific cytotoxic T lymphocytes (CTLs). Blood 108:2942–9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Serafini B, Rosicarelli B, Franciotta D, Magliozzi R, Reynolds R, Cinque P. and others. 2007. Dysregulated Epstein-Barr virus infection in the multiple sclerosis brain. J Exp Med 204:2899–912 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Serafini B, Severa M, Columba-Cabezas S, Rosicarelli B, Veroni C, Chiappetta G. and others. 2010. Epstein-Barr virus latent infection and BAFF expression in B cells in the multiple sclerosis brain: implications for viral persistence and intrathecal B-cell activation. J Neuropathol Exp Neurol 69:677–93 [DOI] [PubMed] [Google Scholar]
- Sibley WA, Bamford CR, Clark K. 1985. Clinical viral infections and multiple sclerosis. Lancet 325:1313–5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simon KC, Yang X, Munger KL, Ascherio A. 2007. Variation in the Epstein-Barr virus receptor, CR2, and risk of multiple sclerosis. Multiple Sclerosis 13:947–8 [DOI] [PubMed] [Google Scholar]
- Smets F, Latinne D, Bazin H, Reding R, Otte JB, Buts JP. and others. 2002. Ratio between Epstein-Barr viral load and anti-Epstein-Barr virus specific T-cell response as a predictive marker of posttransplant lymphoproliferative disease. Transplantation 73:1603–10 [DOI] [PubMed] [Google Scholar]
- Sokal EM, Hoppenbrouwers K, Vandermeulen C, Moutschen M, Léonard P, Moreels A. and others. 2007. Recombinant gp350 vaccine for infectious mononucleosis: a phase 2, randomized, double-blind, placebo-controlled trial to evaluate the safety, immunogenicity, and efficacy of an Epstein-Barr virus vaccine in healthy young adults. J Infect Dis 196:1749–53 [DOI] [PubMed] [Google Scholar]
- Sospedra M, Martin R. 2005. Immunology of multiple sclerosis. Annu Rev Immunol 23:683–747 [DOI] [PubMed] [Google Scholar]
- Souza TA, Stollar BD, Sullivan JL, Luzuriaga K, Thorley-Lawson DA. 2005. Peripheral B cells latently infected with Epstein-Barr virus display molecular hallmarks of classical antigen-selected memory B cells. Proc Natl Acad Sci U S A 102:18093–8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strowig T, Gurer C, Ploss A, Liu Y, Arrey F, Sashihara J. and others. 2009. Priming of protective T cell responses against virus-induced tumors in mice with human immune system components. J Exp Med 206:1423–34 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sumaya CV, Myers LW, Ellison GW. 1980. Epstein-Barr virus antibodies in multiple sclerosis. Arch Neurol 37:94–6 [DOI] [PubMed] [Google Scholar]
- Sumaya CV, Myers LW, Ellison GW, Ench Y. 1985. Increased prevalence and titer of Epstein-Barr virus antibodies in patients with multiple sclerosis. Ann Neurol 17:371–7 [DOI] [PubMed] [Google Scholar]
- Sundström P, Juto P, Wadell G, Hallmans G, Svenningsson A, Nyström L. and others. 2004. An altered immune response to Epstein-Barr virus in multiple sclerosis: a prospective study. Neurology 62:2277–82 [DOI] [PubMed] [Google Scholar]
- Tabi Z, McCombe PA, Pender MP. 1994. Apoptotic elimination of Vb8.2+ cells from the central nervous system during recovery from experimental autoimmune encephalomyelitis induced by the passive transfer of Vb8.2+ encephalitogenic T cells. Eur J Immunol 24:2609–17 [DOI] [PubMed] [Google Scholar]
- Thacker EL, Mirzaei F, Ascherio A. 2006. Infectious mononucleosis and risk for multiple sclerosis: a meta-analysis. Ann Neurol 59:499–503 [DOI] [PubMed] [Google Scholar]
- Thompson AJ, Brazil J, Whelan CA, Martin EA, Hutchinson M, Feighery C. 1986. Peripheral blood T lymphocyte changes in multiple sclerosis: a marker of disease progression rather than of relapse? J Neurol Neurosurg Psychiatry 49:905–12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thorley-Lawson DA, Gross A. 2004. Persistence of the Epstein-Barr virus and the origins of associated lymphomas. N Engl J Med 350:1328–37 [DOI] [PubMed] [Google Scholar]
- Tourtellotte WW, Potvin AR, Baumhefner RW, Potvin JH, Ma BI, Syndulko K. and others. 1980. Multiple sclerosis de novo CNS IgG synthesis: effect of CNS irradiation. Arch Neurol 37:620–4 [DOI] [PubMed] [Google Scholar]
- Trotter JL, Clifford DB, McInnis JE, Griffeth RC, Bruns KA, Perlmutter MS. and others. 1989. Correlation of immunological studies and disease progression in chronic progressive multiple sclerosis. Ann Neurol 25:172–8 [DOI] [PubMed] [Google Scholar]
- Tynan FE, Elhassen D, Purcell AW, Burrows JM, Borg NA, Miles JJ. and others. 2005. The immunogenicity of a viral cytotoxic T cell epitope is controlled by its MHC-bound conformation. J Exp Med 202:1249–60 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ulett G. 1948. Geographic distribution of multiple sclerosis. Dis Nerv Syst 9:342–6 [PubMed] [Google Scholar]
- Van Noort JM, Bajramovic JJ, Plomp AC, van Stipdonk MJB. 2000. Mistaken self, a novel model that links microbial infections with myelin-directed autoimmunity in multiple sclerosis. J Neuroimmunol 105:46–57 [DOI] [PubMed] [Google Scholar]
- Van Sechel AC, Bajramovic JJ, van Stipdonk MJB, Persoon-Deen C, Geutskens SB, van Noort JM. 1999. EBV-induced expression and HLA-DR-restricted presentation by human B cells of aB-crystallin, a candidate autoantigen in multiple sclerosis. J Immunol 162:129–35 [PubMed] [Google Scholar]
- Veldman CM, Cantorna MT, DeLuca HF. 2000. Expression of 1,25-dihydroxyvitamin D3 receptor in the immune system. Arch Biochem Biophys 374:334–8 [DOI] [PubMed] [Google Scholar]
- Vetsika EK, Callan M. 2004. Infectious mononucleosis and Epstein-Barr virus. Expert Rev Mol Med 6(23):1–16 [DOI] [PubMed] [Google Scholar]
- Wandinger K-P, Jabs W, Siekhaus A, Bubel S, Trillenberg P, Wagner H-J. and others. 2000. Association between clinical disease activity and Epstein-Barr virus reactivation in MS. Neurology 55:178–84 [DOI] [PubMed] [Google Scholar]
- Wardemann H, Yurasov S, Schaefer A, Young JW, Meffre E, Nussenzweig MC. 2003. Predominant autoantibody production by early human B cell precursors. Science 301:1374–77 [DOI] [PubMed] [Google Scholar]
- Warner HB, Carp RI. 1981. Multiple sclerosis and Epstein-Barr virus. Lancet 318:1290. [DOI] [PubMed] [Google Scholar]
- Willis SN, Stadelmann C, Rodig SJ, Caron T, Gattenloehner S, Mallozzi SS. and others. 2009. Epstein-Barr virus infection is not a characteristic feature of multiple sclerosis brain. Brain 132:3318–28 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yajima M, Imadome K, Nakagawa A, Watanabe S, Terashima K, Nakamura H. and others. 2008. A new humanized mouse model of Epstein-Barr virus infection that reproduces persistent infection, lymphoproliferative disorder, and cell-mediated and humoral immune responses. J Infect Dis 198:673–82 [DOI] [PubMed] [Google Scholar]
- Zivadinov R, Zorzon M, Weinstock-Guttman B, Serafin M, Bosco A, Bratina A. and others. 2009. Epstein-Barr virus is associated with grey matter atrophy in multiple sclerosis. J Neurol, Neurosurg Psychiatry 80:620–5 [DOI] [PubMed] [Google Scholar]
- Žofková I, Kancheva RL. 1997. The effect of 1,25(OH)2 vitamin D3 on CD4+/CD8+ subsets of T lymphocytes in postmenopausal women. Life Sci 61:147–52 [DOI] [PubMed] [Google Scholar]