

Abstract
Localized translation of axonal mRNAs contributes to developmental and regenerative axon growth. Although untranslated regions (UTRs) of many different axonal mRNAs appear to drive their localization, there has been no consensus RNA structure responsible for this localization. We recently showed that limited expression of ZBP1 protein restricts axonal localization of both β-actin and GAP-43 mRNAs. β-actin 3′UTR has a defined element for interaction with ZBP1, but GAP-43 mRNA shows no homology to this RNA sequence. Here, we show that an AU-rich element (ARE) in GAP-43’s 3′UTR is necessary and sufficient for its axonal localization. Axonal GAP-43 mRNA levels increase after in vivo injury, and GAP-43 mRNA shows an increased half-life in regenerating axons. GAP-43 mRNA interacts with both HuD and ZBP1, and HuD and ZBP1 coimmunoprecipitate in an RNA-dependent fashion. Reporter mRNA with the GAP-43 ARE competes with endogenous β-actin mRNA for axonal localization and decreases axon length and branching similar to the β-actin 3′UTR competing with endogenous GAP-43 mRNA. Conversely, overexpressing GAP-43 coding sequence with it’s 3′UTR ARE increases axonal elongation and this effect is lost when just the ARE is deleted from GAP-43’s 3′UTR.
Keywords: mRNA transport, axon regeneration, GAP-43, HuD, ZBP1, RNA-immunoprecipitation
INTRODUCTION
Protein localization helps to establish the subcellular domains of polarized cells like neurons. The segregation of proteins into particular subcellular regions occurs in part by targeting mRNAs for localization and translation in these regions (Jung et al. 2012). Such spatial control of gene expression enables the long axonal processes of neurons to rapidly and autonomously respond to stimuli at distances 100 to 10,000 fold more than the cell body diameter (Donnelly et al. 2010). For example, locally synthesized β-actin protein contributes to axonal pathfinding in developing neurons (Zhang et al. 2001) and regeneration in adult neurons (Donnelly et al. 2011). Axonally synthesized Importin-β1, RanBP1 and Stat3α are needed for retrogradely signaling nerve injury and triggering transcriptional responses in neuronal cell bodies (Ben-Yaakov et al. 2012, Perry et al. 2012, Yudin et al. 2008). These and other studies indicate that mRNA localization into axons is used as a mechanism to spatially and temporally introduce new proteins at sites distant from the cell body.
Hundreds of different mRNAs have now been shown to localize into axons of different neuron types (Gumy et al. 2011, Taylor et al. 2009, Willis et al. 2007, Zivraj et al. 2010). However, the mechanism(s) driving transport of these mRNAs has only been defined for a few transcripts (Donnelly et al. 2011). A 54 nucleotide (nt) sequence in β-actin mRNA’s 3′UTR, a ‘zip-code’, is necessary for it’s subcellular localization (Kislauskis et al. 1994). Localization elements have been established for some axonal mRNAs, but the RNA-binding proteins recognizing these elements have not been identified (Ben-Yaakov et al. 2012, Vuppalanchi et al. 2010, Yudin et al. 2008, Merianda et al. 2012). Moreover, there have not been any consensus sequences for the axonally localizing mRNAs. Several lines of evidence point to secondary or higher order structures as comprising the functional elements within RNAs (Andreassi and Riccio 2009). Consistent with this, we recently showed that ZBP1, a KH domain RNA-binding protein well characterized for its role in transport of βactin mRNA (Doyle and Kiebler 2012), is also needed for axonal localization of GAP-43 mRNA, despite that the mRNA shows no clear homology to β-actin’s zip code element (Donnelly et al. 2011). Here, we show that an element in GAP-43 mRNA that was previously shown to bind to the ELAV-like RNA-binding protein HuD is necessary and sufficient for axonal localization of GAP-43 mRNA. HuD binds to an AU-rich regulatory element (ARE) in the 3′UTR of GAP-43 mRNA and stabilizes this transcript (Beckel-Mitchener et al. 2002, Mobarak et al. 2000). However, these stability studies were limited to whole cell lysates and the possibility that the localized mRNA can be stabilized has not been tested. Our data show that GAP-43 mRNA survival in axons is increased during regeneration and this correlates with an increase in HuD levels in the sciatic nerve after injury. ZBP1 and HuD associate in an RNA-dependent fashion and GAP-43 mRNA coimmunoprecipitates with both ZBP1 and HuD, suggesting that these RNA-binding proteins form a complex on GAP-43 mRNA. A reporter mRNA with the ARE of GAP-43 mRNA decreases axon outgrowth and branching and competes with endogenous β-actin mRNA for axonal localization, consistent with a ‘squelching’ mechanism by which the mRNA ligands compete for the binding to RNA-binding protein. Further, overexpressing GAP-43 fusion proteins increased axonal outgrowth but only when it’s 3′UTR ARE is included for axonal mRNA localization.
METHODS
Animal surgeries
150–175 g male Sprague Dawley rats (Harlan) were used. All animal protocols were approved by the Institutional Animal Care and Use Committees of Alfred I. duPont Hospital for Children and Drexel University. The contralateral nerve served as a “sham” control where the nerve is exposed but not injured. Animals were euthanized 7 days after nerve crush.
DNA expression constructs
All fluorescent reporter constructs for analyses of RNA localization/translation were based on eGFP with myristoylation element (GFPmyr) originally provided by Erin Schuman (Aakalu et al. 2001) or mCherry plasmid (mCherrymyr) (Vuppalanchi et al. 2012). Schematic of the constructs is included in Supplemental Information (Fig. S1). To isolate the 3′UTR of rat GAP-43 RNA (GenBank ID: NM_017195), total RNA from rat DRG was reverse-transcribed with iScript (BioRad) and then amplified by PCR using Pfu polymerase (Stratagene). Primers for amplifying the full GAP-43 3′UTR and subsequent deletion mutants were engineered to contain terminal Notl and Xhol restriction sites. All PCR products were initially cloned into pCR 2.1 TOPO (Invitrogen) for sequencing and sequenced. Inserts were subcloned into pGFPmyr or pmCherrymyr. Primer extension PCR was used to generate the 40 nt ARE sequence of GAP-43 3′UTR (nt 1211–1250) with the following primers; sense, ATTTGCGGCCGCGTGGGAGTCCACTTTCCTCTCTATTTCTCTCTGTTCCAAG; antisense, CCGCTCGAGCTTGGAACAGAGAGAAATAGAGAGGAAAGTGGACTCCCAC. This segment was cloned directly into the pGFPmyr plasmid.
For generating myc-tagged HuD (HuDMyc), the open reading frame (ORF) of HuD (GenBank accession NM_001077651) with terminal BglII and BamHI restriction sites was PCR-amplified and cloned into pAcGFP1-N3 (Clontech) replacing AcGFP (pHuD-N3). Myc tag was then N-terminally fused to HuD by adaptor ligation into AgeI and BglII sites to generate Myc-HuD fusion protein (HuDMyc). mCherry-ZBP1 with FLAG tag (mCh-ZBP1FLAG) was provided by Dr. Gary J. Bassell (Welshhans and Bassell 2011). mCherry-GAP-43 fusion protein constructs with 3′UTR of GAP-43 or γ-actin (Donnelly et al. 2013) were generated by replacing the full length 3′UTR with the modified 3′UTRs outlined above (Fig. S1).
Cell culture and transfections
L4-6 DRGs were dissociated with 50 U collagenase type XI (Sigma) for 25 min at 37°C and then triturated with a fire polished Pasteur pipette (Twiss et al. 2000). After washing by centrifugation, cells were resuspended in DMEM:F12 (Life Tech.), 1x N1 supplement (Sigma), 5% horse serum, 5% fetal bovine serum (Hyclone), and 10 μM cytosine arabinoside (Sigma). DRG neurons were plated on coverslips coated with poly-L-lysine (Sigma) and laminin (Millipore) at 37°C, 5% CO2. F11 cells, a hybridoma of embryonic rat DRG neurons and mouse neuroblastoma cell line N18TG (Platika et al. 1985), were cultured in DMEM:F12, 10% fetal bovine serum, 100 U/ml penicillin, and 0.1 mg/ml streptomycin (Sigma) on poly-L-lysine coated tissue culture plates.
For transfection, dissociated DRGs were pelleted by centrifugation at 100 × g for 5 min and resuspended in ‘nucleofector solution’ (Rat Neuron Nucleofector kit; Lonza). 5–7 μg plasmid was electroporated using an AMAXA Nucleofector apparatus (program G-13; Lonza). DRGs were immediately plated as above and medium was replaced 4 h later. F11 cells were transfected using Lipofectamine™2000 (Life Tech.) in DMEM:F12 plus 0.5% fetal bovine serum (8,000 cells/cm2).
Fluorescence in situ hybridization and immunofluorescence (FISH/IF)
Oligonucleotide probes were used to detect endogenous mRNAs and cRNA probes were used to detect reporter mRNAs. Antisense probes for rat GAP-43 have been described (Willis et al. 2007); these span nucleotides 158–207, 300–349, 799–848 and 1157–1206. Scrambled oligonucleotide probes were used for control. Oligonucleotides were synthesized with 5′-amino modifier C6 at four thymidines per oligonucleotide; probes were chemically labeled with digoxigenin succinamide ester (Roche). To generate sense and antisense cRNA probes, linearized pcDNA3-mCherry was in vitro transcribed using SP6 or T7 RNA polymerase (Promega) and digoxigen-labeled nucleotide mixture (Roche). cRNAs were then alkaline hydrolyzed to 100–150 nt length. All probes were stored at −80°C until used.
FISH for DRG cultures was similar to previously described (Willis et al. 2007). All steps were carried out at room temperature unless otherwise indicated. Coverslips were rinsed in phosphate buffered saline (PBS), fixed in buffered 4% paraformaldehyde for 20 min, and then permeabilized in 0.3% Triton X-100 for 5 min. The coverslips were then rinsed in PBS and equilibrated in 1X SSC with 50% formamide. Hybridization was performed at 55°C for 2 h in 5X SSC, 50% formamide, 20% dextran sulphate, 10 μg/ml salmon sperm DNA, 10 μg/ml E. coli tRNA and 0.5X Denhardt’s solution containing 1.66 ng/μl oligonucleotides or 5 ng/μl cRNA. Coverslips were then washed twice for 20 min in 50% formamide/2X SSC at 37°C, followed by three washes in 1X SSC for 10 min each on a rotary shaker.
FISH for tissue sections was performed as previously described (Vuppalanchi et al. 2010). Sciatic nerves were fixed in 2% paraformaldehyde for 2 h, cryoprotected in 30% sucrose at 4°C overnight. Cryosections (10 μm) were mounted on Superfrostplus glass slides (Fisher), and stored at −20°C until used. Sections were warmed to room temperature, rinsed in PBS for 20 min and then soaked in 0.25% acetic anhydride, 0.1 M triethanolamine hydrochloride and 0.9% NaCl at pH 8.0 for 10 min. Sections were then rinsed in 2X SSC, dehydrated through a graded series of ethanol (70%, 5 min; 95%, 5 min; and 100%, 5 min), delipidated in chloroform for 5 min and rinsed with 100% ethanol. Sections were washed in 1X SSC and then hybridized overnight at 42°C with 1.2 ng/μl oligonucleotide probes in 2X SSC plus 40% formamide, 250 μg/ml tRNA, 100 μg/ml sheared salmon sperm DNA and 10% dextran sulfate. Sections were washed in 1X SSC three times for 15 min each at 50°C, PBS for 5 min, and PBS with 0.3% Triton X-100 for another 5 min.
For subsequent IF, samples were equilibrated in 50 mM Tris, 150 mM NaCl,1% heat-shock BSA, and1% protease-free BSA (Roche) (‘IF buffer’) and then blocked in IF buffer plus 2% heat-shock BSA, 2% fetal bovine serum for 1 h. After blocking, samples were incubated for 1 h in the following primary antibodies in IF buffer: chicken anti-neurofilament (NF) H (1:1,000; Chemicon), rabbit anti-peripherin (1:1,500; Millipore), rabbit anti-S100 (1:500; Dako) and sheep anti-digoxigenin (1:1,000; Roche). Samples were washed in IF buffer three times and then incubated in Cy5-conjugated anti-rabbit or anti-chicken in IF buffer for 1 h. To increase sensitivity of detecting probes, an additional Cy3-conjugated anti-mouse Fab fragments (1:200; Jackson ImmunoRes.) for tissues or Alexa488-conjugated anti-sheep IgG antibody (1:1,000, Invitrogen) for cultures was included with secondary antibodies. Coverslips were mounted using PVA-DABCO (Sigma) anti-fading mounting medium. Epifluorescent microscopy with Leica DMRXA2 was used to capture signals for cultured neurons and laser scanning confocal microscopy with Leica TCS/SP2 was used for capturing signals in tissue sections.
Analysis of RNA levels in sciatic nerve explants
Sciatic nerve explants (~ 20 mm) were incubated at 37°C and 5% CO2 for up to 12 h in DMEM:F12, 10% fetal bovine serum, 100 U/ml penicillin, 0.1 mg/ml streptomycin and 20 μM cyclosporin A (CsA; LC Laboratories). RNA was isolated from the explants using RNAqueous (Ambion). RNA samples were treated with DNase I (Promega) for 30 min at 37°C. RNA was quantified using a VersaFluor fluorometer (Biorad) with Quant-iT RiboGreen RNA (Invitrogen); 100 ng RNA was reverse transcribed (RT) using iScript (Biorad). The cDNA samples were processed for quantitative PCR (qPCR) using Prism 7900HT (Applied Biosystems) with 2x SybrGreen Master Mix (Quiagen). Nerve GAP-43 mRNA signals were normalized to GAPDH mRNA using the comparative threshold (Ct) method and expressed as fold change relative to t = 0 h from ΔΔCt calculations. The primer used were: GAP-43 sense, 5′-CGACAGGATGAGGGTAAAGAA-3′ and antisense, 5′-GACAGGAGAGGAAACTTCAGAG-3′; GAPDH (GenBank ID: NM_017008) sense, 5′-GACAACTTTGTGAAGCTCATT-3′; antisense, 5′-GGGCCTCTCTCTTGCTCTCA-3′. Quadruplicate reactions were analyzed from at least 4 separate experiments.
Fluorescence recovery after photobleaching (FRAP)
FRAP was performed as previously described (Yudin et al. 2008). Briefly, GFPmyr transfected cultures were maintained at 37°C in an environmental chamber fitted to a Leica TCS/SP2 confocal microscope. Axons were imaged every 30 sec for 2 min with 488 nm laser (15% power) to establish a baseline fluorescence. For photobleaching, a region of interest (ROI) was exposed to 90% power 488 nm laser for 40 frames at 1.6 sec intervals. ROIs consisted of the distal 50–150 μm of the axon (≥ 400 μm from the cell body). Recovery of GFP fluorescence was monitored every 60 sec over 20 min using 488 nm laser (15% power). GFP emission was collected with a band filter of 498–530 nm for each sequence. To test for translation dependence, cultures were pre-treated with 50 μM anisomycin for 15 min prior to FRAP experiments.
Recovery from photobleaching was quantified from > 7 neurons over at least three separate transfection experiments. ImageJ was used to calculate average pixels/μm2 in the ROIs of the raw confocal images. The fluorescence signal at t=0 min (post-bleach) was subtracted from intensity values; this corrected intensity was then normalized to the pre-bleach (set to 100%). Original WMV files for time-lapse sequences were converted to AVI format using Power Video Converter (v1.5.43, Apus software).
Immunoprecipitation and RNA co-immunoprecipitation
For analysis of interactions of HuD and ZBP1, transiently transfected F11 cells were lysed in ‘polysome buffer’ (100 mM KCl, 5 mM MgCl2, 0.5% NP40, 1 mM DTT, 100 U/ml RNase Out, 400 μM vanadyl ribonucleoside complexes, protease inhibitor cocktail, 10 mM HEPES, pH 7.0). Primary antibodies were absorbed to protein G Dynabeads (Life Technologies). Lysates were pre-cleared with dynabeads, and then HuDMyc or mCh-ZBP1FLAG was precipitated using anti-Myc- or anti-FLAG coated protein G Dynabeads in NT2 buffer (150 mM NaCl, 1 mM MgCl2, 0.05% NP40, 50 mM Tris-HCl, pH 7.4). 9B11 mouse anti-Myc (1:50; Cell Signaling) and M1 mouse anti-FLAG antibody (10 μg/ml; Sigma). Dynabeads coated with anti-mouse IgG were used as a negative control. Normalized lysates were incubated with Dynabeads for 1 h at 4 °C, then vigorously washed with polysome buffer and processed for SDS/PAGE or RNA isolation. The precipitates resolved by SDS/PAGE were analyzed by immunoblotting using standard methods (Towbin et al. 1979). For isolation of RNA, beads were incubated in TRIzol (Life Tech.) and extracted by phenol-chloroform. RNA was precipitated with isopropanol using a glycogen carrier. RT-PCR was used to detect bound mRNAs in quadruplicate over at least 3 separate experiments. Parallel qPCR with serial dilutions of plasmid DNA as template was used to calculate absolute yield of each mRNA.
Immunoblotting was performed as previously described (Merianda et al. 2012). The following antibodies were used: anti-HuD (1:1,000; Clonegene), mouse anti-Myc (1:1,000; Roche) and HRP-conjugated anti-rabbit or anti-mouse (1:3,000; Abcam). Signals were detected by ECLplus per manufacturer’s protocol (GE Biosciences).
Axonal growth analyses
Axon morphology was visualized using GFP, mCherry and/or immunofluorescence as described (Donnelly et al. 2013). Mean axon length was obtained by measuring the longest axon per neuron using ImageJ (NIH). The number of branches was obtained by manually scoring processes ≥ 25 μm in length visualized by GFP or mCherry signals.
Statistical Analyses
GraphPad Prism 5 software package (GraphPad) was used for all statistical analyses. For FRAP, repeated measures ANOVA with Bonferroni post-hoc comparisons was performed to test the differences of means between groups and differences within group mean over time. For RTqPCR, axon growth and FISH quantification, results were analyzed ANOVA (one-way or two-way as indicated) with Bonferroni post-hoc comparisons. To compare regression lines of mRNA survival, results from decay experiment of naïve and injury conditioned sciatic nerve were tested using nonlinear regression analysis with a straight-line model.
RESULTS
In vivo localization of GAP-43 mRNA in axons
GAP-43 is a neuronal-specific phosphoprotein that localizes to axonal growth cones during both developmental and regenerative growth (Benowitz and Routtenberg 1997). GAP-43 mRNA has been detected in axons of cultured neurons in RNA profiling studies (Taylor et al. 2009, Willis et al. 2007) and mice with depleted ZBP1 show decreased levels of GAP-43 mRNA in peripheral nerve by RT-PCR (Donnelly et al. 2011). Since peripheral nerves contain both neuronal and non-neuronal components, we used FISH/IF to determine if the GAP-43 mRNA detected in sciatic nerve by RT-PCR is within axons. Granular FISH signals for GAP-43 mRNA were seen in the adult rat sciatic nerve that overlapped with axonal but not glial proteins, consistent with a restricted axonal localization for GAP-43 mRNA in the nerves (Fig. 1A). Similar granular GAP-43 mRNA signals were seen in the axons of dissociated DRG neurons in culture, both along the axon shaft and extending into growth cones (Fig. 1B). No signals were seen with the scrambled probes (Figs. 1A,B, insets). The cell bodies of these cultured DRG neurons showed a more robust signal for GAP-43 mRNA (Fig. 1A), suggesting that only a fraction of the GAP-43 mRNA localizes into the axonal compartment as previously shown for β-actin mRNA (Donnelly et al. 2011).
Figure 1. GAP-43 mRNA localizes to peripheral axons.
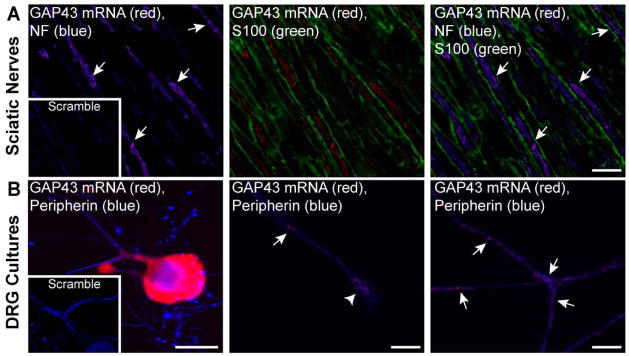
A, Representative images of single optical planes from adult rat sciatic nerve are shown for GAP-43 mRNA and indicated proteins; exposure matched image for scrambled probe (red) is shown in the inset panels. Images were taken at the center of the axoplasm for axons (arrows) to completely distinguish axonal and Schwann cell signals. Granular signals for GAP-43 mRNA overlap with only the NF positive axons [Scale bars = 5 μm].
B, Representative images of cultured DRG neurons are shown for GAP-43 mRNA and peripherin protein; exposure matched image for scrambled probe (red) is shown in the inset panels. The left panel shows a low magnification of neuron cell body; the middle and the right panels show higher power images of axon shaft (arrows) and growth cone (arrowhead) with GAP-43 mRNA extending into each [Scale bars: left panel = 50 μm, middle and right panels = 5 μm].
Increased survival of GAP-43 mRNA in injured sciatic nerve
GAP-43 mRNA and protein have been shown to increase in the DRG during PNS nerve regeneration, with GAP-43 protein levels rising in the nerve (Schreyer and Skene 1993, Van der Zee et al. 1989). FISH/IF analyses showed a clear increase in GAP-43 mRNA in the proximal segments of crushed sciatic nerve compared to the contralateral (uninjured) sciatic nerve (Fig. 2A). In XYZ, XZ and YZ projections, the GAP-43 mRNA signals in both the crushed and naïve nerve sections completely overlapped with the axonal markers (Fig. 2A). Quantification of axonal FISH signals showed a significant increase in axonal GAP-43 mRNA signals in 7 day injured sciatic nerve (Fig. S2A). Moreover, neurons cultured 7 days after an in vivo conditioning crush injury showed similarly increased GAP-43 mRNA levels in axons (Fig. S2B). The L4-5 DRGs showed an approximately five-fold increase in GAP-43 mRNA levels at 7 days after crush injury compared to naïve (Fig. S2C). This increase is much more than seen in the nerve, suggesting again that only a fraction of total GAP-43 mRNA localizes into axons. The DRG levels of amphoterin and β-actin mRNAs did not significantly change after injury, but the axonal levels of β-actin mRNA were decreased after injury both in vitro and in vivo (Figs. S2A,B). Since axonal β-actin and GAP-43 mRNAs appear to be cotransported by ZBP1 (Donnelly et al. 2011), these data suggest that the overall increase in GAP-43 mRNA after injury could deplete β-actin mRNA from the axonal transport machinery by competing for limited levels of ZBP1.
Figure 2. GAP-43 mRNA is stabilized in regenerating sciatic nerve.
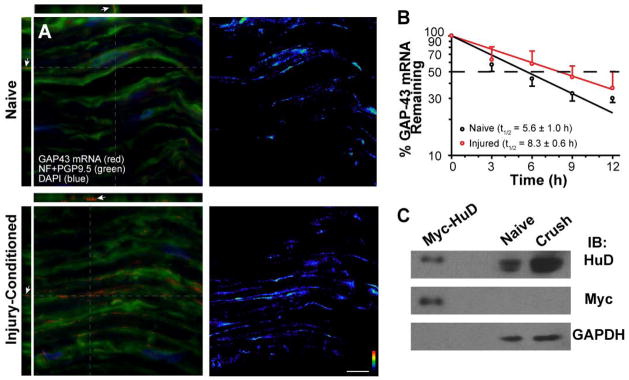
A, Matched exposure/post-processing confocal images of FISH/IF for 7 day crushed and naïve sciatic nerve are shown. Merged channels of confocal maximum projections for XYZ are shown in left column and RNA channel is shown in right column (RNA is shown as indicated intensity spectrum). Orthogonal XY and YZ projections at the levels indicated by grey dotted lines show that the GAP-43 mRNA signals completely overlap with the axons stained for NF and PGP 9.5 proteins (arrows) [Scale bars = 10 μm].
B, Relative stability of sciatic nerve GAP-43 mRNA was tested using naïve and 7 day injured nerve explants treated with 20 μM cyclosporin A to delay axonal degeneration. GAP-43 mRNA signals from RTqPCR were normalized to GAPDH mRNA and are expressed relative to time 0 to estimate of half-life of the nerve GAP-43 mRNA. Error bars represent standard deviation (n = 4). The slopes of the decay curves for GAP-43 mRNA were analyzed using nonlinear regression curves with a straight-line model (R2 =0.8855 for naïve and R2 =0.9837 for crush; p=0.0492).
C, Representative immunoblot for axoplasm extracts prepared from the naïve and 7 day crushed sciatic nerves is shown. GAPDH signals show relatively equivalent loading between crush and naïve nerves. However, HuD signals were consistently higher in the crush nerve (1.98 ± 0.28 fold crush vs. naïve axoplasm; n = 3). To test HuD antibody specificity, lysates from F11 cells transfected with HuDMyc construct were included in these gels. Probing with anti-Myc antibody shows comparably migrating band as the anti-HuD antibody.
Considering that GAP-43 mRNA is known to be stabilized by the ELAV-like RNA-binding protein HuD (Beckel-Mitchener et al. 2002), the increase in axonal GAP-43 mRNA levels in regenerating axons could be related to stabilization. To directly test the survival of GAP-43 mRNA in axons, we turned to a nerve explant preparation. Axons of severed nerve segments normally undergo Wallerian degeneration over a few hours in vitro; however, CsA treatment delays this degeneration by preventing activation of the mitochondrial permeability transition pore and extends axon survival for several days (Barrientos et al. 2011). We used this CsA-treated explant model to compare survival of GAP-43 mRNA in isolated segments of naïve vs. crushed sciatic nerve. RTqPCR values for GAP-43 mRNA were normalized to GAPDH mRNA, which is expressed by the Schwann cells and fibroblasts included in these explants. For half-life determination, all time points were compared to t=0 h. The axonal GAP-43 mRNA showed a half-life of 5.6 ± 1.0 h in naïve nerve (Fig. 2B), which is comparable to ~5 h GAP-43 mRNA half-life measurements reported for PC12 cells (Mobarak et al. 2000). In contrast, explants of proximal crushed sciatic nerve showed GAP-43 mRNA half-life of 8.3 ± 0.6 h. The slopes of the decay curves were also significantly different comparing naïve and crushed nerve explants, confirming that GAP-43 mRNA has a longer survival in the explants of injured axons. The nerve levels of HuD protein were also consistently higher in the injured than in the naïve nerve (Fig. 2C). Taken together, these studies suggest that GAP-43 mRNA is stabilized in regenerating axons, with a corresponding increase in HuD in the regenerating nerve that could modulate survival of the axonal GAP-43 mRNA.
Axonal localization of GAP-43 mRNA requires the HuD binding site
The 3′UTRs of localizing mRNAs have most often been linked to their subcellular localization (Andreassi and Riccio 2009). To determine if GAP-43 mRNA’s 3′UTR confers axonal localization, we generated reporter constructs with diffusion-limited GFPmyr (Aakalu et al. 2001) carrying sequences from the rat GAP-43 3′UTR (Fig. S1). This approach with diffusion-limited reporters through myristoylation has previously been used by several groups to test for RNA localization in axons and dendrites by translation-dependent recovery of reporter fluorescence (see (Merianda et al.2012) and reference within). Adult rat DRG neurons were transfected with the GFPmyr3′GAP43 constructs and FRAP was used to test for reporter mRNA translation in axons. By bleaching ROIs ≥ 400 μm away from the cell body and limiting recovery to 20 min, the possibility for transport or diffusion of reporter protein from the soma is dramatically decreased (Yudin et al. 2008), allowing us to visualize axonal translation events.
Reporter constructs with the full 3′UTR of GAP-43 (nt 896-1483; GFPmyr3′GAP43896-1483; Fig. S1) showed recovery from photobleaching in distal axons (Figs. 3A,E). Analysis of multiple FRAP sequences of GFPmyr3′GAP43896-1483 transfected DRGs showed that axonal GFP signal was significantly increased compared to post-bleach (t=0 min) levels at 4 min after photobleaching; however, anisomycin-treated cultures showed no significant recovery (Figs. 3B,E). DRGs transfected with GFPmyr with nt 896-1053 of GAP-43 (GFPmyr3′GAP43896-1053; Fig. S1) showed low axonal fluorescence that did not recover from photobleaching (Figs. S3A,D). In contrast, neurons expressing GFPmyr with nt 1067-1281 of GAP-43 (GFPmyr3′GAP431067-1281; Fig. S1) showed significant recovery from photobleaching that was attenuated by anisomycin (Figs. S3B,C,E). It should be noted that although the GFPmyrGAP43896-1053 and GFPmyrGAP431067-1281 constructs do not have the GAP-43′s polyadenylation (pA) element, each would be polyadenylated through downstream bovine growth hormone (BGH) pA element in the GFPmyr construct. Consistent with this, all of the above constructs showed robust GFP mRNA signals in the cell body (Fig. S4).
Figure 3. GAP-43 3′UTR is sufficient for translation of a reporter mRNA in axons.

A–D, Representative time-lapse images for FRAP analyses of DRG neurons transfected with the indicated constructs are shown. Laser intensity, gain and offset are matched for each image sequence. The boxed regions represent the ROIs that were photobleached. Arrows indicate growth cones [Scale bars = 50 μm].
E–G, Quantifications for multiple time-lapse sequences as in A–D are shown as normalized average signal intensity relative to pre- and post-bleach ± SEM (n ≥ 7 neurons over > 3 separate transfection experiments for each construct; * = p ≤ 0.05, ** = p ≤ 0.01 and *** = p ≤ 0.001 for indicated points vs. t=0 min by repeated measures ANOVA with Bonferroni post-hoc comparisons).
These data suggest that an axonal localization signal(s) lies within nt 1067-1281 of rat GAP-43′s 3′UTR. This region contains the ARE that has been implicated in stabilization of GAP-43 mRNA through HuD binding (Mobarak et al. 2000). Thus, we asked if just the ARE could be sufficient for localizing GFPmyr mRNA into axons by generating a GFPmyr construct with GAP-43 nt 1211-1250 that comprise the ARE (GFPmyr3′GAP43ARE; Fig. S1). Axons of DRGs transfected with GFPmyr3′GAP43ARE showed significant recovery by FRAP, but the difference between basal and protein-synthesis inhibited cultures was less than we saw with GFPmyr3′GAP43896-1483 and GFPmyr3′GAP431067-1281 reporters (Figs. 3C,F). Similar to the other deletion constructs, nt 1211-1250 does not contain GAP-43’s pA element; however, the BGH pA element downstream of GFPmyr would generate a 3′UTR of at least 315 nt in the GFPmyr3′GAP43ARE mRNA. As a direct test for the ability of nt 1211-1250 to localize reporter mRNA into axons, we generated a GFPmyr reporter with full length GAP-43 3′UTR where nt 1211-1250 were deleted (GFPmyr3′GAP43896-1483ΔARE; Fig. S1). DRGs transfected with GFPmyr3′GAP43896-1483ΔARE showed only weak axonal fluorescence with no recovery by FRAP (Figs. 3D,G).
The above data point to GAP-43′s ARE as necessary and sufficient for GAP-43 mRNA targeting into axons. However, the FRAP data did show more variability in recovery than we have seen in previous studies (Akten et al. 2011, Ben-Yaakov et al. 2012, Vuppalanchi et al. 2010, Vuppalanchi et al. 2012, Yudin et al. 2008, Merianda et al. 2012, Pacheco and Twiss 2012, Perry et al. 2012). Thus, it was possible that the GFPmyr3′GAP43896-1053 and GFPmyr3′GAP43896-1483ΔARE mRNA could localize into but not be efficiently translated within axons. To test this possibility, we generated a myristoylated mCherry (mChmyr) constructs with the above GAP-43 3′UTR sequences. This allowed us to directly visualize the reporter protein and reporter mRNA by FISH without loss of fluorescence from our hybridization protocol that plagues the GFP signal in our hands. mCherry mRNA was detected in axons and growth cones of neurons transfected with the mChmyr3′GAP43896-1483, mChmyr3′GAP431067-1281 and mChmyr3′GAP43ARE constructs (Fig. 4A-C). There was no axonal mCherry mRNA signal in neurons transfected with mChmyr3′GAP43896-1053 or mChmyr3′GAP43896-1483ΔARE (Figs. 4D,E). mCherry mRNA and protein were detected in cell bodies for all of these mChmyr3′GAP43 constructs (Fig. S4), indicating that each reporter mRNA was clearly expressed. Thus, consistent with the FRAP data, reporter mRNA only localized into axons of neurons expressing mRNAs with nt 1211-1250 of GAP-43 mRNA (i.e., the ARE). Taken together, these data indicate that the GAP-43’s ARE confers axonal mRNA localization.
Figure 4. TheARE in GAP-43’s 3′UTR is necessary and sufficient for axonal localization of a reporter mRNA.

Representative exposure matched images of mCherry mRNA and NF protein are shown for DRG neurons expressing indicated constructs. Both the axons (arrowheads) and growth cones (arrows) of DRG neurons transfected with reporter constructs containing the ARE from GAP-43’s 3′UTR (nt 1211-1250) show mCherry mRNA signals (A, B and C). However, those lacking the ARE show mCherry mRNA in the cell body only (D and E, asterisks indicates cell body). Sense cRNA riboprobe for mCherry shows no signal in exposure-matched images of mChmyr3′GAP43896-1483 transfected DRGs (F) [Scale bars: A = 10 μm; B and C = 20 μm; D–F = 50 μm].
HuD and ZBP1 bind to GAP-43 mRNA
Although the data above could link HuD to GAP-43 localization into and stabilization within axons, our previous work pointed to ZBP1 as needed for axonal localization of GAP-43 mRNA (Donnelly et al. 2011). Atlas et al (2004) had shown that HuD and the human ortholog of ZBP1 (IMP-1) can bind to the 3′UTR of Tau mRNA, presumably forming a heteromeric complex. Therefore, we tested if GAP-43 mRNA may interact with both HuD and ZBP1. We first asked if ZBP1 and HuD coimmunoprecipitate in conditions that preserve RNA-protein associations. Using neuronal-like F11 cells transiently transfected with epitope-tagged HuD and ZBP1 (HuDMyc and mCh-ZBP1FLAG; Fig. S1), HuD was detected in ZBP1 immunoprecipitates and ZBP1 was detected in HuD immunoprecipitates (Fig. 5A). This interaction was lost when lysates were treated with RNase A prior to immunoprecipitation, suggesting that HuD and ZBP1 interaction is RNA-dependent. β-actin and GAP-43 mRNAs were easily amplified from these immunoprecipitates, but we were also able to detect amphoterin and histone H4 mRNAs in these immunoprecipitates (Fig. 5B). Amphoterin mRNA is not predicted to be a target for ZBP1 interaction based on previous studies (Donnelly et al. 2011, Donnelly et al. 2013); however, its interaction with HuD has not been tested. The RNA co-immunoprecipitation approach or ‘RIP assay’ has been associated with high background for other proteins and non-specific RNA-protein associations (Darnell 2010). Thus, we turned to RTqPCR to determine if these coimmunoprecipitating mRNAs were enriched in the HuD or ZBP1 immunocomplexes compared to IgG control and inputs. Both β-actin and GAP-43 mRNAs showed significant enrichment in the HuD and ZBP1 immunocomplexes compared to IgG immunocomplexes, but amphoterin and H4 mRNAs showed no enrichment (Fig. 5C). Thus, the amphoterin and H4 mRNA signals detected in the HuD and ZBP1 immunoprecipitates by standard RT-PCR represent non-specific interactions in the RIP assay. By directly comparing the amounts of GAP-43 and β-actin mRNAs coimmunoprecipitating with HuD and ZBP1, β-actin mRNA showed significantly higher coimmunoprecipitation with ZBP1 than with HuD and GAP-43 mRNA showed significantly higher coimmunoprecipitation with HuD than with ZBP1 (Fig. 5C).
Figure 5. HuD and ZBP1 interact in an RNA-dependent fashion.
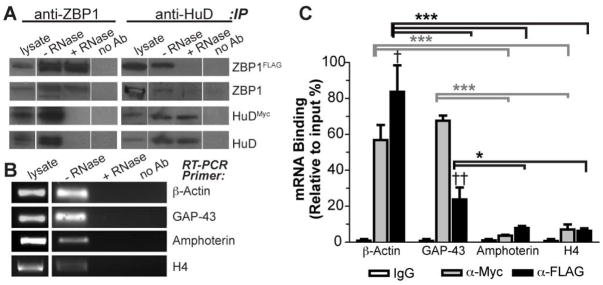
A, ZBP1 and HuD were immunoprecipitated from mCh•ZBP1FLAG and HuDMyc co-transfected F11 cells and immunoblotted for indicated proteins; immunoprecipitation with mouse IgG was used as a control for non-specific binding (no Ab). HuD coimmunoprecipitates with ZBP1 and ZBP1 coimmunoprecipates with HuD (− RNase). However, the interaction of HuD and ZBP1 is near completely lost in lysates that were pretreated with RNase A (+ RNase). Note that gels were loaded with empty lanes separating lysate and no Ab samples from the immunoprecipitates to avoid any contamination from adjacent lanes (lanes shown were cropped from the same immunoblot exposure).
B, Standard RT-PCR analysis of RNAs isolated from immunoprecipitates shows that several transcripts can be detected in ZBP1 and HuD immunoprecipitates performed as in A (− RNase). No RNAs were detected when coimmunoprecipitation was performed with mouse IgG (no Ab) or from samples pretreated with RNase (+ RNase).
C, RTqPCR analysis was used to determine if mRNAs from B are enriched in the HuD (anti-Myc, grey columns) or ZBP1 (anti-FLAG, black columns) immunoprecipitates. Average ± S.D. for each mRNA normalized to input and then to no Ab control is shown († = p<0.05 and †† = p<0.01 by Student’s T-test for HuDMyc vs. mCh•ZBP1FLAG precipitates; * = p<0.05, ** = p<0.01 and *** = p<0.001 by two-way ANOVA with Bonferroni post-hoc comparisons with grey symbols representing comparison between indicated HuD precipitates and black lines representing comparison between indicated ZBP1 precipitates).
Axonal growth modulation by locally generated GAP-43 protein requires the 3′UTR ARE of GAP-43 mRNA
We recently showed that neurons expressing the GFPmyr with the GAP-43 3′UTR show decreased axon length and branching compared to control conditions (Donnelly et al. 2013). This effect likely occurs through the exogenous mRNA competing with endogenous βactin and GAP-43 mRNAs for localization into axons, so we asked if the ARE element alone might similarly attenuate axonal growth. Identical to prior work (Donnelly et al. 2013), neurons expressing GFPmyr3′GAP43896-1483 showed decreased axon length and branching compared to neurons expressing GFPmyr plus the 3′UTR of γ-actin (GFPmyr3′γ-actin) (Figs. 6A,B and S5). However, neurons expressing GFPmyr3′GAP43896-1483ΔARE showed no significant difference in axon length and branching compared to GFPmyr3′γ-actin expressing neurons. On the other hand, neurons expressing GFPmyr3′GAP43ARE showed significantly shorter axons with fewer branches compared to neurons expressing GFPmyr3′γ-actin (Figs. 6A,B and Fig. S5). These data are consistent with the exogenous 3′UTR of GAP-43 competing with endogenous mRNAs for transport into axons through its ARE. Consistent with this, neurons expressing GFPmyr3′GAP43ARE showed 22% reduction (p<0.01, Student’s T-test) in axonal levels of endogenous β-actin mRNA compared to neurons expressing control GFP or GFP 3′γ-actin mRNAs (Fig. S2D). Interestingly, the increased DRG and axonal levels of GAP-43 mRNA after nerve injury are accompanied by a decrease in axonal levels of β-actin mRNA without any significant change in DRG levels of β-actin mRNA (Figs. S2A,B,C). This raises the possibility that endogenous GAP-43 and β-actin mRNAs compete for ZBP1 and HuD binding.
Figure 6. Introduction of GAP-43’s 3′UTR attenuates axon outgrowth, but axonal GAP-43 coding sequence mRNA increases axonal outgrowth.

A–B, Quantifications of axon length (A) and axon branching (B) in DRG neurons transfected with indicated constructs are shown. Transfection with the ARE-containing constructs (GFPmyr3′GAP43896-1483 and GFPmyr3′GAP43ARE) significantly decreased axon length and branching compared to the ARE deleted GFPmyr3′GAP43896-14833ARE and empty GFP constructs.
C,D, Quantifications of axon length (A) and axon branching (B) in DRG neurons transfected with indicated constructs are shown. The ARE containing mCh•GAP43 fusion protein constructs (mCh•GAP43-3′GAP43ARE, mCh•GAP43-3′GAP43ARE-3′γ-actin) significantly increased axon length and decreased axon branch numbers compared to those lacking the GAP-43 mRNA ARE (mCh•GAP43-3′γ-actin, GAP43•mCh-3′GAP43896-1483ΔARE) (N ≥ 30 neurons for each construct over 3 separate experiments; * = p<0.05, ** = p < 0.001 and *** = p < 0.001 by one-way ANOVA with Bonferroni post-hoc comparisions).
Overexpression of axonally targeted GAP-43 was recently shown to support elongating growth of axons (Donnelly et al. 2013). To determine whether the ARE of GAP-43’s 3′UTR is sufficient to increase axonal growth by localizing GAP-43 mRNA into axons, we generated mCherry-GAP-43 fusion protein constructs with and without the GAP-43 3′UTR ARE (mCh•GAP43-3′GAP43ARE; GAP43•mCh-3′GAP43896-1483ΔARE; Fig. S1). Neurons expressing mCh•GAP43ARE showed a significant increase in axon length compared to neurons expressing GAP43•mCh-3′GAP43896-1483ΔARE (Figs. 6C and S6). Branching of axons was also significantly decreased in the mCh•GAP43-3′GAP43ARE expressing neurons (Figs. 6D and S6). Neurons expressing mCh•GAP43 with the non-localizing 3′UTR of γ-actin (mCh•GAP43-3′γ-actin) showed no significant change in axon length or branching compared to controls (Figs. 6C,D and Fig. S6). However, neurons expressing GAP43mCh-3′γ-actin where we had inserted the GAP-43 ARE sequence (mCh•GAP43-3′GAP43ARE-3′γ-actin) showed significantly increased axon length and decreased branching similar to the mCh•GAP43-3′GAP43ARE expressing neurons (Figs. 6C,D). Taken together, these data indicate that the ARE in GAP-43 3′UTR is responsible for GAP-43 mRNA’s delivery into axons that accounts for the enhanced axonal outgrowth recently demonstrated by Donnelly et al. (2013).
DISCUSSION
Neurons transport mRNAs with subsequent translation of the localized transcripts to regulate gene expression over time and distance. For axons, distance is a daunting factor since these processes can extend for several centimeters even in the rodents used here. Several studies have shown functional roles for axonally synthesized proteins (Jung et al. 2012), including recent work from our lab pointing to locally generated GAP-43’s role in axon growth and regeneration (Donnelly et al. 2013, Donnelly et al. 2011). Transport of mRNAs into axons appears to be regulated during development, regeneration and response to extracellular stimuli that locally modulate axonal growth (Gumy et al. 2011, Taylor et al. 2009, Willis et al. 2007, Zivraj et al. 2010). The translational output from an mRNA can obviously be affected by the stability of the transcript, with longer lived transcripts being available for translation over longer durations, but the half-lives of axonal mRNAs have not been addressed. GAP-43 mRNA is known to be stabilized by HuD binding to its ARE sequence (Beckel-Mitchener et al. 2002, Mobarak et al. 2000), and both HuD and GAP-43 mRNA have been shown to localize to neuronal processes (Smith et al. 2004). In the present study, we identify the HuD binding site in GAP-43 mRNA as a cis-element that is necessary and sufficient for transporting GAP-43 mRNA into axons of cultured sensory neurons. Moreover, axonal mRNA transport through this element is needed for GAP-43’s axon growth promoting effects. Axonal GAP-43 mRNA appears to be stabilized during regeneration. Thus, axonal GAP-43 protein levels can be regulated both by targeting the mRNA for transport into axons and locally stabilizing the mRNA within axons. Putting these data together with previous studies showing that ZBP1 is needed for transport of GAP-43 mRNA into DRG axons (Donnelly et al. 2011), our present data suggest a collaborative effort of a HuD-ZBP1 complex for post-transcriptional regulation of GAP-43 mRNA.
Cells know which mRNAs to localize based on structural features of the mRNAs, consisting of either primary sequence or higher order structures (Andreassi and Riccio 2009). These structures are thought to be bound by RNA-binding proteins like ZBP1 that are needed for targeting mRNAs to subcellular domains. Despite that hundreds of mRNAs localize into axons (Gumy et al. 2011, Taylor et al. 2009, Willis et al. 2007, Zivraj et al. 2010), no consensus sequences for axonal mRNA targeting have come to light. Based on protein-RNA coimmunoprecipitation assays, RNA-binding proteins appear fairly promiscuous in their interactions with mRNAs. For example, 293 mRNAs coimmunoprecipitated with IMP1 (i.e., human ZBP1) in HEK1 cells (Jonson et al. 2007), and 670 mRNAs coimmunoprecipitated with HuD in brain lysates (Bolognani et al. 2010). Protein-RNA cross-linking assays have shown higher specificity for proteins directly interacting with mRNAs, but these have not addressed the transported RNA particle or localized mRNA. It seems unlikely that all localizing mRNAs will use a single RNA-binding protein for transport. A complex of proteins on a single mRNA may provide a mechanism for higher specificity and/or functionality than could be achieved by single proteins. Enrichment of GAP-43 mRNA in both HuD and ZBP1 immunocomplexes and the RNA-dependent coimmunoprecipitation of HuD with ZBP1 seen here are consistent with a heteromeric complex of RNA-binding proteins. The relative affinities of GAP-43 and β-actin mRNAs for HuD and ZBP1 in the RIP assays may reflect additional complexity in these mRNA:protein interactions that have not been considered in post-transcriptional mechanisms studies to date.
Similar to GAP-43 studied here, the 3′UTRs of a few other axonal mRNAs have been shown to contain elements needed to localize the transcripts (Aronov et al. 1999, Ben-Yaakov et al. 2012, Kislauskis et al. 1994, Vuppalanchi et al. 2010, Yudin et al. 2008, Merianda et al. 2012, Donnelly et al. 2013, Perry et al. 2012). Our previous studies indicated that ZBP1 is needed for axonal localization of GAP-43 mRNA in the DRG neurons studied here (Donnelly et al. 2011), despite that GAP-43 has no clear zipcode element for direct ZBP1 binding. Thus, it is surprising that GAP-43’s ARE, both by FRAP analyses for GFP translation in distal axons and FISH for axonal localization of reporter mRNA, is necessary and sufficient for localization of GAP-43 mRNA into axons. This ARE shows no homology to the zipcode of β-actin, so it is not clear how or if ZBP1 directly interacts with GAP-43 mRNA. One possibility is that a complex of ZBP1 and HuD binds to the GAP-43 ARE, with HuD providing the RNA binding capacity. The RNA dependent interaction of ZBP1 and HuD as well as the GAP-43 ARE competing with β-actin mRNA for axonal localization would be consistent with this notion.
GAP-43 is a neuron-specific phosphoprotein that contributes to axonal growth and has been suggested to play a role in presynaptic plasticity (Benowitz and Routtenberg 1997, Skene 1989). Though GAP-43 protein is known to increase axonal growth, transgenic overexpression of GAP-43 produced only modest increases in axonal growth on its own (Bomze et al. 2001). However, subcellular localization of GAP-43 mRNA has only recently been recognized and the above transgenic model did not incorporate GAP-43’s 3′UTR. Recent work indicates that the axonally generated GAP43 protein supports elongating growth of axons (Donnelly et al. 2013). The data here indicate that the ARE is responsible for delivery of GAP-43 mRNA into axons, so it is not surprising that the previously published GAP-43 transgene without the GAP-43 3′UTR sequences had little effect on axon growth (Bomze et al. 2001).
Consistent with previous studies (Schreyer and Skene 1993, Van der Zee et al. 1989), expression of GAP-43 mRNA increases in our nerve injury model. There is also a commensurate increase in GAP-43 mRNA levels in the proximal sciatic nerve after injury and in the axons of injury-conditioned DRG neurons in culture. The depletion of axonal β-actin mRNA with expression of GFPmyr3′GAP43ARE suggests that ZBP1-dependent transport of GAP-43 mRNA into axons contributes to the increased axonal levels of GAP-43 mRNA in the injured neurons. The increased expression of GAP-43 and its ARE competing with β-actin mRNA for binding to ZBP1 are also likely responsible for the decrease in axonal β-actin mRNA levels after nerve injury. Thus, transcriptional upregulation of GAP-43 mRNA has the potential to change the cohort of mRNAs that localize into axons. However, our data also indicate that GAP-43 mRNA is stabilized in axons, which is a function previously attributed to HuD rather than ZBP1 (Beckel-Mitchener et al. 2002, Mobarak et al. 2000) and could further increase GAP-43 protein levels in the regenerating axon.
Multi-functionality has increasingly been recognized for RNA-binding proteins. HuD has been linked to stabilization, transport and translation of neuronal mRNAs. HuD is a member of the ELAV-like/Hu family of RNA-binding proteins and is highly neuron-specific (Robinow et al. 1988). Colocalization of GAP-43 mRNA and HuD have previously been reported for the neurites of differentiated PC12 cells (Smith et al. 2004), but it was not clear if HuD contributed to localization of the mRNA. HuD’s interaction with Tau mRNA was defined as a protein complex of HuD, IMP1 (i.e., ZBP1) and Ras-GAP SH3 domain binding protein 1 (G3BP1) in the axonal-like processes of differentiated P19 cells, but the function of individual proteins in this complex was not tested (Aronov et al. 1999, Atlas et al. 2004). HuD was similarly isolated as a protein binding to neuritin mRNA (also called CPG15), and here HuD in complex with SMN protein was linked to axonal transport of neuritin mRNA in cultured CNS neurons (Akten et al. 2011). HuD interacting with the 3′ poly-A tails of mRNAs was shown to facilitate translational initiation, but this appeared to be a non-specific interaction through the third RNA binding domain (RBD) of HuD (Fukao et al. 2009), rather than the specific interaction of HuD with mRNAs that we have studied here. HuD RBDs 1 and 2 appear to be responsible for binding to AREs (Deschenes-Furry et al. 2006). It is tempting to speculate that HuD may transition from ARE to poly-A binding and facilitate translation of GAP-43 mRNA in axons. Alternatively, ZBP1 may play a role in translational regulation of GAP-43 just as it has been shown to do for β-actin mRNA (Huttelmaier et al. 2005, Sasaki et al. 2010).
The possibility that the half-life of a localized mRNA is altered under different growth conditions has not previously been tested. The increased survival of GAP-43 mRNA after nerve injury seen here is consistent with stabilization of the transcript in regenerating axons and is further supported by a qualitative increase in HuD levels in the regenerating nerve. Thus, HuD may contribute to both localization and stabilization of GAP-43 mRNA. Localized bicoid mRNA is similarly stabilized in Drosophila embryos, but bicoid contains distinct localizing and destabilizing elements in its 3′UTR (Gottlieb 1992, Surdej and Jacobs-Lorena 1998), suggesting that different proteins mitigate these functions. Arc mRNA stability is also regulated in dendrites, but this involves separate RNA elements for localization and stability since Arc mRNA is rapidly degraded following translation through non-sense mediated decay pathways (Bramham et al. 2008, Giorgi et al. 2007). For GAP-43, survival and transport of the mRNA appear to be linked, with GAP-43’s ARE contributing to both mechanisms.
Proteomic analyses of neuronal RNA transport particles have shown a complex population of proteins, setting the stage for multiple proteins interacting in a single RNP complex (Elvira et al. 2006, Kanai et al. 2004). Despite the high background of the ‘RIP’ assays used here (Darnell 2010), our data indicate that by using appropriate immunoprecipitation controls and further accounting for input levels of mRNAs by RTqPCR, amphoterin and H4 mRNAs were not enriched in the ZBP1 and HuD immunocomplexes and represent non-specific binding in the RIP analysis. On the other hand, GAP-43 and β-actin mRNAs clearly were enriched in the ZBP1 and HuD precipitates, and GAP-43 mRNA showed higher enrichment in the HuD vs. ZBP1 immunocomplexes and β-actin mRNA showed higher enrichment in the ZBP1 vs. HuD immunocomplexes. This may reflect different affinities of these proteins for the mRNAs or interactions through protein-protein rather than direct protein-RNA interactions. Future studies will be needed to resolve this at a molecular level. Nonetheless, our studies point to a higher level of complexity for transport of individual axonal mRNAs than has previously been uncovered. The competition for mRNAs binding to RNA interacting proteins also raises the possibility that changes in levels of individual mRNAs, through transcription or stabilization or both, can alter the axonal proteome.
Supplementary Material
Acknowledgments
This work was funded by grants from NIH (P20GM103464 to SY, R01-NS41596 to JLT and R01-NS30255 to NPB), the Craig H. Neilsen Foundation (124124 to SY), and the Dr. Miriam and Sheldon G. Adelson Medical Research Foundation. Erin Schuman provided the original GFPmyr reporter construct. ZBP1FLAG was kindly provided by Dr. Gary J. Bassell at Emory University. The authors declare that they have no conflicts of interest. S.Y. and J.L.T. designed research; S.Y., H.H.K, P.K., C.J.D., A.K., D.V., M.P., S.J.L., and T.T.M. performed research; N.PB contributed new reagents; S.Y., H.H.K. and J.L.T analyzed data. All authors edited and approved the final submitted version.
Abbreviations used
- ARE
AU-rich regulatory element
- CsA
cyclosporin A
- DRG
dorsal root ganglion
- FISH
fluorescence in situ hybridization
- IF
immunofluorescence
- FRAP
fluorescence recovery after photobleaching
- ROI
region of interest
- UTR
untranslated region
References
- Aakalu G, Smith WB, Nguyen N, Jiang C, Schuman EM. Dynamic visualization of local protein synthesis in hippocampal neurons. Neuron. 2001;30:489–502. doi: 10.1016/s0896-6273(01)00295-1. [DOI] [PubMed] [Google Scholar]
- Akten B, Kye MJ, Hao le T, Wertz MH, Singh S, Nie D, Huang J, Merianda TT, Twiss JL, Beattie CE, Steen JA, Sahin M. Interaction of survival of motor neuron (SMN) and HuD proteins with mRNA cpg15 rescues motor neuron axonal deficits. Proc Natl Acad Sci U S A. 2011;108:10337–10342. doi: 10.1073/pnas.1104928108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andreassi C, Riccio A. To localize or not to localize: mRNA fate is in 3′UTR ends. Trends Cell Biol. 2009;19:465–474. doi: 10.1016/j.tcb.2009.06.001. [DOI] [PubMed] [Google Scholar]
- Aronov S, Marx R, Ginzburg I. Identification of 3′UTR region implicated in tau mRNA stabilization in neuronal cells. J Mol Neurosci. 1999;12:131–145. doi: 10.1007/BF02736927. [DOI] [PubMed] [Google Scholar]
- Atlas R, Behar L, Elliott E, Ginzburg I. The insulin-like growth factor mRNA binding-protein IMP-1 and the Ras-regulatory protein G3BP associate with tau mRNA and HuD protein in differentiated P19 neuronal cells. J Neurochem. 2004;89:613–626. doi: 10.1111/j.1471-4159.2004.02371.x. [DOI] [PubMed] [Google Scholar]
- Barrientos SA, Martinez NW, Yoo S, Jara JS, Zamorano S, Hetz C, Twiss JL, Alvarez J, Court FA. Axonal degeneration is mediated by the mitochondrial permeability transition pore. J Neurosci. 2011;31:966–978. doi: 10.1523/JNEUROSCI.4065-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beckel-Mitchener AC, Miera A, Keller R, Perrone-Bizzozero NI. Poly(A) tail length-dependent stabilization of GAP-43 mRNA by the RNA-binding protein HuD. J Biol Chem. 2002;277:27996–28002. doi: 10.1074/jbc.M201982200. [DOI] [PubMed] [Google Scholar]
- Ben-Yaakov K, Dagan SY, Segal-Ruder Y, Shalem O, Vuppalanchi D, Willis DE, Yudin D, Rishal I, Rother F, Bader M, Blesch A, Pilpel Y, Twiss JL, Fainzilber M. Axonal transcription factors signal retrogradely in lesioned peripheral nerve. Embo J. 2012;31:1350–1363. doi: 10.1038/emboj.2011.494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benowitz LI, Routtenberg A. GAP-43: an intrinsic determinant of neuronal development and plasticity. Trends Neurosci. 1997;20:84–91. doi: 10.1016/s0166-2236(96)10072-2. [DOI] [PubMed] [Google Scholar]
- Bolognani F, Contente-Cuomo T, Perrone-Bizzozero NI. Novel recognition motifs and biological functions of the RNA-binding protein HuD revealed by genome-wide identification of its targets. Nucleic Acids Res. 2010;38:117–130. doi: 10.1093/nar/gkp863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bomze HM, Bulsara KR, Iskandar BJ, Caroni P, Skene JH. Spinal axon regeneration evoked by replacing two growth cone proteins in adult neurons. Nat Neurosci. 2001;4:38–43. doi: 10.1038/82881. [DOI] [PubMed] [Google Scholar]
- Bramham CR, Worley PF, Moore MJ, Guzowski JF. The immediate early gene arc/arg3.1: regulation, mechanisms, and function. J Neurosci. 2008;28:11760–11767. doi: 10.1523/JNEUROSCI.3864-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Darnell RB. HITS-CLIP: panoramic views of protein-RNA regulation in living cells. Wiley Interdiscip Rev RNA. 2010;1:266–286. doi: 10.1002/wrna.31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deschenes-Furry J, Perrone-Bizzozero N, Jasmin BJ. The RNA-binding protein HuD: a regulator of neuronal differentiation, maintenance and plasticity. Bioessays. 2006;28:822–833. doi: 10.1002/bies.20449. [DOI] [PubMed] [Google Scholar]
- Donnelly CJ, Fainzilber M, Twiss JL. Subcellular communication through RNA transport and localized protein synthesis. Traffic. 2010;11:1498–1505. doi: 10.1111/j.1600-0854.2010.01118.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Donnelly CJ, Park M, Spillane M, Yoo S, Pacheco A, Gomes C, Kim HH, Merianda TT, Gallo G, Twiss JL. Axonally Synthesized beta-Actin and GAP-43 Proteins Support Distinct Modes of Axonal Growth. J Neurosci. 2013;33:3311–3322. doi: 10.1523/JNEUROSCI.1722-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Donnelly CJ, Willis DE, Xu M, Tep C, Jiang C, Yoo S, Schanen NC, Kirn-Safran CB, van Minnen J, English A, Yoon SO, Bassell GJ, Twiss JL. Limited availability of ZBP1 restricts axonal mRNA localization and nerve regeneration capacity. Embo J. 2011;30:4665–4677. doi: 10.1038/emboj.2011.347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doyle M, Kiebler MA. A zipcode unzipped. Genes Dev. 2012;26:110–113. doi: 10.1101/gad.184945.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elvira G, Wasiak S, Blandford V, Tong XK, Serrano A, Fan X, del Rayo Sanchez-Carbente M, Servant F, Bell AW, Boismenu D, Lacaille JC, McPherson PS, DesGroseillers L, Sossin WS. Characterization of an RNA granule from developing brain. Mol Cell Proteomics. 2006;5:635–651. doi: 10.1074/mcp.M500255-MCP200. [DOI] [PubMed] [Google Scholar]
- Fukao A, Sasano Y, Imataka H, Inoue K, Sakamoto H, Sonenberg N, Thoma C, Fujiwara T. The ELAV protein HuD stimulates cap-dependent translation in a Poly(A)- and eIF4A-dependent manner. Mol Cell. 2009;36:1007–1017. doi: 10.1016/j.molcel.2009.11.013. [DOI] [PubMed] [Google Scholar]
- Giorgi C, Yeo GW, Stone ME, Katz DB, Burge C, Turrigiano G, Moore MJ. The EJC factor eIF4AIII modulates synaptic strength and neuronal protein expression. Cell. 2007;130:179–191. doi: 10.1016/j.cell.2007.05.028. [DOI] [PubMed] [Google Scholar]
- Gottlieb E. The 3′ untranslated region of localized maternal messages contains a conserved motif involved in mRNA localization. Proc Natl Acad Sci U S A. 1992;89:7164–7168. doi: 10.1073/pnas.89.15.7164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gumy LF, Yeo GS, Tung YC, Zivraj KH, Willis D, Coppola G, Lam BY, Twiss JL, Holt CE, Fawcett JW. Transcriptome analysis of embryonic and adult sensory axons reveals changes in mRNA repertoire localization. RNA. 2011;17:85–98. doi: 10.1261/rna.2386111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huttelmaier S, Zenklusen D, Lederer M, Dictenberg J, Lorenz M, Meng X, Bassell GJ, Condeelis J, Singer RH. Spatial regulation of beta-actin translation by Src-dependent phosphorylation of ZBP1. Nature. 2005;438:512–515. doi: 10.1038/nature04115. [DOI] [PubMed] [Google Scholar]
- Jonson L, Vikesaa J, Krogh A, Nielsen LK, Hansen T, Borup R, Johnsen AH, Christiansen J, Nielsen FC. Molecular composition of IMP1 ribonucleoprotein granules. Mol Cell Proteomics. 2007;6:798–811. doi: 10.1074/mcp.M600346-MCP200. [DOI] [PubMed] [Google Scholar]
- Jung H, Yoon BC, Holt CE. Axonal mRNA localization and local protein synthesis in nervous system assembly, maintenance and repair. Nat Rev Neurosci. 2012;13:308–324. doi: 10.1038/nrn3210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kanai Y, Dohmae N, Hirokawa N. Kinesin transports RNA: isolation and characterization of an RNA-transporting granule. Neuron. 2004;43:513–525. doi: 10.1016/j.neuron.2004.07.022. [DOI] [PubMed] [Google Scholar]
- Kislauskis EH, Zhu X, Singer RH. Sequences responsible for intracellular localization of beta-actin messenger RNA also affect cell phenotype. J Cell Biol. 1994;127:441–451. doi: 10.1083/jcb.127.2.441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Merianda TT, Vuppalanchi D, Yoo S, Blesch A, Twiss JL. Axonal Transport of Neural Membrane Protein 35 mRNA Increases Axon Growth. J Cell Sci. 2012 doi: 10.1242/jcs.107268. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mobarak CD, Anderson KD, Morin M, Beckel-Mitchener A, Rogers SL, Furneaux H, King P, Perrone-Bizzozero NI. The RNA-binding protein HuD is required for GAP-43 mRNA stability, GAP-43 gene expression, and PKC-dependent neurite outgrowth in PC12 cells. Mol Biol Cell. 2000;11:3191–3203. doi: 10.1091/mbc.11.9.3191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pacheco A, Twiss JL. Localized IRES-dependent translation of ER chaperone protein mRNA in sensory axons. PLoS One. 2012;7:e40788. doi: 10.1371/journal.pone.0040788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perry RB, Doron-Mandel E, Iavnilovitch E, Rishal I, Dagan SY, Tsoory M, Coppola G, McDonald MK, Gomes C, Geschwind DH, Twiss JL, Yaron A, Fainzilber M. Subcellular Knockout of Importin beta1 Perturbs Axonal Retrograde Signaling. Neuron. 2012;75:294–305. doi: 10.1016/j.neuron.2012.05.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Platika D, Boulos MH, Baizer L, Fishman MC. Neuronal traits of clonal cell lines derived by fusion of dorsal root ganglia neurons with neuroblastoma cells. Proc Natl Acad Sci U S A. 1985;82:3499–3503. doi: 10.1073/pnas.82.10.3499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robinow S, Campos AR, Yao KM, White K. The elav gene product of Drosophila, required in neurons, has three RNP consensus motifs. Science. 1988;242:1570–1572. doi: 10.1126/science.3144044. [DOI] [PubMed] [Google Scholar]
- Sasaki Y, Welshhans K, Wen Z, Yao J, Xu M, Goshima Y, Zheng JQ, Bassell GJ. Phosphorylation of zipcode binding protein 1 is required for brain-derived neurotrophic factor signaling of local beta-actin synthesis and growth cone turning. J Neurosci. 2010;30:9349–9358. doi: 10.1523/JNEUROSCI.0499-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schreyer DJ, Skene JH. Injury-associated induction of GAP-43 expression displays axon branch specificity in rat dorsal root ganglion neurons. J Neurobiol. 1993;24:959–970. doi: 10.1002/neu.480240709. [DOI] [PubMed] [Google Scholar]
- Skene JH. Axonal growth-associated proteins. Annu Rev Neurosci. 1989;12:127–156. doi: 10.1146/annurev.ne.12.030189.001015. [DOI] [PubMed] [Google Scholar]
- Smith CL, Afroz R, Bassell GJ, Furneaux HM, Perrone-Bizzozero NI, Burry RW. GAP-43 mRNA in growth cones is associated with HuD and ribosomes. J Neurobiol. 2004;61:222–235. doi: 10.1002/neu.20038. [DOI] [PubMed] [Google Scholar]
- Surdej P, Jacobs-Lorena M. Developmental regulation of bicoid mRNA stability is mediated by the first 43 nucleotides of the 3′ untranslated region. Mol Cell Biol. 1998;18:2892–2900. doi: 10.1128/mcb.18.5.2892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taylor AM, Berchtold NC, Perreau VM, Tu CH, Li Jeon N, Cotman CW. Axonal mRNA in uninjured and regenerating cortical mammalian axons. J Neurosci. 2009;29:4697–4707. doi: 10.1523/JNEUROSCI.6130-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Towbin H, Staehelin T, Gordon J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc Natl Acad Sci U S A. 1979;76:4350–4354. doi: 10.1073/pnas.76.9.4350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Twiss JL, Smith DS, Chang B, Shooter EM. Translational control of ribosomal protein L4 mRNA is required for rapid neurite regeneration. Neurobiol Dis. 2000;7:416–428. doi: 10.1006/nbdi.2000.0293. [DOI] [PubMed] [Google Scholar]
- Van der Zee CE, Nielander HB, Vos JP, Lopes da Silva S, Verhaagen J, Oestreicher AB, Schrama LH, Schotman P, Gispen WH. Expression of growth-associated protein B-50 (GAP43) in dorsal root ganglia and sciatic nerve during regenerative sprouting. J Neurosci. 1989;9:3505–3512. doi: 10.1523/JNEUROSCI.09-10-03505.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vuppalanchi D, Coleman J, Yoo S, Merianda TT, Yadhati AG, Hossain J, Blesch A, Willis DE, Twiss JL. Conserved 3′-untranslated region sequences direct subcellular localization of chaperone protein mRNAs in neurons. J Biol Chem. 2010;285:18025–18038. doi: 10.1074/jbc.M109.061333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vuppalanchi D, Merianda TT, Donnelly C, Williams G, Yoo S, Ratan RR, Willis DE, Twiss JL. Lysophosphatidic acid differentially regulates axonal mRNA translation through 5′UTR elements. Mol Cell Neurosci. 2012 doi: 10.1016/j.mcn.2012.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Welshhans K, Bassell GJ. Netrin-1-induced local beta-actin synthesis and growth cone guidance requires zipcode binding protein 1. J Neurosci. 2011;31:9800–9813. doi: 10.1523/JNEUROSCI.0166-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Willis DE, van Niekerk EA, Sasaki Y, Mesngon M, Merianda TT, Williams GG, Kendall M, Smith DS, Bassell GJ, Twiss JL. Extracellular stimuli specifically regulate localized levels of individual neuronal mRNAs. J Cell Biol. 2007;178:965–980. doi: 10.1083/jcb.200703209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yudin D, Hanz S, Yoo S, Iavnilovitch E, Willis D, Gradus T, Vuppalanchi D, Segal-Ruder Y, Ben-Yaakov K, Hieda M, Yoneda Y, Twiss JL, Fainzilber M. Localized regulation of axonal RanGTPase controls retrograde injury signaling in peripheral nerve. Neuron. 2008;59:241–252. doi: 10.1016/j.neuron.2008.05.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang HL, Eom T, Oleynikov Y, Shenoy SM, Liebelt DA, Dictenberg JB, Singer RH, Bassell GJ. Neurotrophin-induced transport of a beta-actin mRNP complex increases beta-actin levels and stimulates growth cone motility. Neuron. 2001;31:261–275. doi: 10.1016/s0896-6273(01)00357-9. [DOI] [PubMed] [Google Scholar]
- Zivraj KH, Tung YC, Piper M, Gumy L, Fawcett JW, Yeo GS, Holt CE. Subcellular profiling reveals distinct and developmentally regulated repertoire of growth cone mRNAs. J Neurosci. 2010;30:15464–15478. doi: 10.1523/JNEUROSCI.1800-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.