

Abstract
The topical toxicities of five commercial grade pesticides commonly sprayed in apple orchards were estimated on adult worker honey bees, Apis mellifera (L.) (Hymenoptera: Apidae) and Japanese orchard bees, Osmia cornifrons (Radoszkowski) (Hymenoptera: Megachilidae). The pesticides were acetamiprid (Assail 30SG), λ-cyhalothrin (Warrior II), dimethoate (Dimethoate 4EC), phosmet (Imidan 70W), and imidacloprid (Provado 1.6F). At least 5 doses of each chemical, diluted in distilled water, were applied to freshly-eclosed adult bees. Mortality was assessed after 48 hr. Dose-mortality regressions were analyzed by probit analysis to test the hypotheses of parallelism and equality by likelihood ratio tests. For A. mellifera, the decreasing order of toxicity at LD50 was imidacloprid, λ-cyhalothrin, dimethoate, phosmet, and acetamiprid. For O. cornifrons, the decreasing order of toxicity at LD50 was dimethoate, λ-cyhalothrin, imidacloprid, acetamiprid, and phosmet. Interaction of imidacloprid or acetamiprid with the fungicide fenbuconazole (Indar 2F) was also tested in a 1∶1 proportion for each species. Estimates of response parameters for each mixture component applied to each species were compared with dose-response data for each mixture in statistical tests of the hypothesis of independent joint action. For each mixture, the interaction of fenbuconazole (a material non-toxic to both species) was significant and positive along the entire line for the pesticide. Our results clearly show that responses of A. mellifera cannot be extrapolated to responses of O.cornifrons, and that synergism of neonicotinoid insecticides and fungicides occurs using formulated product in mixtures as they are commonly applied in apple orchards.
Introduction
Pollinator species such as honey bee [Apis mellifera (L.) (Hymenoptera: Apidae)] and the Japanese orchard bee [Osmia cornifrons (Radoszkowski) (Hymenoptera: Megachilidae)] provide important services in orchard and other agricultural ecosystems [1]–[6]. Members of the genus Osmia are important and efficient pollinators of tree fruit crops such as apples, plums and cherries, among other economically important fruit and nut crops [7]–[9]. The Japanese orchard bee (also known as the Japanese horn-faced bee) is commercially used for pollination of pears and apples in Japan [9]. This species was introduced into the mid-Atlantic region of the United States by USDA scientists in 1977 [10], [11], has since become commercially available, and is now an important (and efficient) wild pollinator in tree fruit orchards in Pennsylvania [12], [13]. One O. cornifrons can set up to 80% more apple flowers per day compared with flowers set by one honey bee, A. mellifera (L.), worker [14]. These two species are complementary pollinators of apple, peach and pear orchards in the Northeastern United States [12], [13], [15]–[19].
A recent significant decline in bee populations in the United States [20]–[23] and elsewhere [24] has led researchers to investigate the effects of possible factors such as pesticide residues [25]–[33], different types of pathogens [34]–[36], adjuvants [37], various parasites [38], floral resource availability and diversity [3], and pest management and agricultural practices [39] on the health, abundance and diversity of different species of bees and pollinators. Although most past research has emphasized the effects of these factors on either A. mellifera or on bumble bee species, the relative effects of pesticide mixtures (e.g., insecticides and fungicides) on two different species of pollinators (for instance, O. cornifrons versus A. mellifera) have not been investigated.
In agricultural production systems, various classes of chemicals are used for management of various pests and diseases. Commercial and farmer tank-mixes of insecticide and fungicide are used to reduce operational or production costs (or both). In tree fruit orchards in Pennsylvania, pesticides are applied for management of a complex of diverse pest species. These include fruit-feeding Lepidoptera, leaf rollers, mites, aphids, plum curculio and stink bugs [40]–[42]. The most critical period to reduce pollinators' exposure to pesticide application in apple orchards is bloom. Different pesticides are applied for control of pests such as rosy apple aphid, European apple sawfly, and plum curculio just before, during, and immediately after bloom. This timing is also critical for control of fungal diseases such as apple scab, Venturia inequalis (Cooke) and apple powdery mildew, Podosphaera leucotricha (Ell. & Evherh.) [40]. However, information about the toxicity of these toxicant mixtures to beneficial invertebrates including pollinators such as O. cornifrons and A. mellifera must be obtained if rational conservation plans for pollinators are to be implemented. Well-designed laboratory bioassays provide the scientific basis for such decisions.
Our studies were done to provide basic information about the comparative toxicities of commonly-used orchard pesticides and pesticide-fungicide mixtures to O. cornifrons and A. mellifera. Results of our experiments were used to test the general premise that one species such as A. mellifera can serve as a surrogate species for a larger taxonomic group such as several families of bees, or as a surrogate for an arbitrary group such as all terrestrial arthropods [43]. Our study expands a previous investigation of differential toxicity of imidacloprid to A. mellifera versus Bombyx mori [44] and suggests the direction in which the study of ecotoxiciology of pollinators can progress [45]. Of most importance, our study provides an example of the type of information necessary to improve the sensitivity of testing pesticides on diverse species in the superfamily Apoidea [46] in the future.
Materials and Methods
Insects
O.cornifrons were purchased from a single source in Wisconsin where they had been reared in an organic apple orchard. Because fruit from this orchard were used only for cider production, pesticide use was minimal. Larvae were reared in natural Phragmites reed bundles and in wooden blocks lined with paper straws. Cocoons containing the overwintering adults were removed and refrigerated at 3°C until 1 April to ensure that their chilling requirements had been met. Loose cocoons were then held inside an incubator (25°C, constant darkness) until adults emerged. Adults were held in darkness until treated 24–72 hr after emergence (24 hr for the males; 24–72 hr for the females). Emergence occurred from April through May. A. mellifera used in this study were purchased as new packages from Gardner Apiaries, Spell Bee (Baxley, GA). Each package included a queen and workers. The packages were exposed to the miticides fluvalinate and coumaphos for bee mite control. They were then put into hives pre-sterilized by irradiation. Colonies were established in the spring and kept in an isolated area at least 6 km from any pesticide applications.
Treatments
For treatment, cages were made of a Petri dish (100×20 mm) encasing a 100 mm-long wire mesh cylinder constructed of hardware cloth (with 3×3 mm openings). One side of the Petri dish had two holes made with a heated cork borer. The larger hole was used to put treated bees into the dish. Once the cage was full, the hole was sealed with tape. The other (smaller) hole held a glass vial for ad lib feeding of a 50% sucrose solution.
For each replication (per chemical or combination) with a species, 10 bees were placed in a 50 ml centrifuge tube with a pair of forceps. The tube was placed in an ice bath for 1–2 min. to immobilize the bees, which were then poured onto a paper towel. If a bee did not move after the cold anesthesia, it was discarded from the bioassay. Each bee was picked up at the base of the wings, treated on the thorax, and placed in the cage. One µl/bee was applied with a Hamilton repeating dispenser (Hamilton Company, Reno, NV) that held a 50 µl syringe. After each group of 10 bees was treated, the cage was set aside and checked after ∼3 min. to ensure that all bees appeared to be healthy. Six cages were placed on their sides inside a plastic container, which also contained a moist paper towel and a jar of a saturated NaCl solution to maintain ∼75% RH. Six caps from 20 ml scintillation vials were used to separate and hold the cages in place.
Water was used as the solvent to mimic what growers use in the field to apply the pesticides. No problems with formulation solubility were observed. The commercial formulations (AI% ; manufacturer) were Assail 30SG (acetamiprid 30%; United Phosphorous Inc., King of Prussia, PA), Dimethoate 4EC (dimethoate 43.5%; Drexel Chemical Company, Memphis, TN), Imidan 70W (phosmet 70%; Gowan Company, Yuma, AZ), Provado 1.6F (imidacloprid 17.4%; Bayer CropScience, Research Triangle Park, NC) and Warrior II (λ - cyhalothrin 22.8%; Syngenta, Wilmington, DE). Interactions of the fungicide Indar 2F (fenbuconazole 22.86%; Dow AgroSciences LLD, Indianapolis, IN) in a 1∶1 proportion with Assail 30SG and in a 1∶2 proportion with Provado 1.6F were also tested.
Experimental Designs and Data Analyses
In all experiments, mortality was tallied after 48 hr. Control mortality was <5%. Average control mortality was 2.7%. At least 6 doses of each pesticide plus a control (water only) were tested in each replication per pesticide per bee species. Each replication included a total of 60–135 bees of each species depending on species' availability. Dose-mortality regressions were estimated assuming the normal distribution (i.e., probit model) with the computer program PoloPlus [47] as described by Robertson et al. [48]. We used a two-step procedure to analyze data for each chemical. In the first step, we examined plots of standardized residuals for outliers, which were then eliminated from the data sets. The second and final probit analysis was done to test hypotheses of parallelism (slopes not significantly different) and equality (slopes and intercepts not significantly different) with likelihood ratio tests [48]. PoloPlus also calculated lethal dose ratios (LDR's) of the most toxic chemical compared with all other chemicals for each species. An LDR provides a means to test whether two LD's are significantly different (i.e., when the 95% CI for the LDR did not include the value 1.0 [47], [48]).
For tests with a mixture, at least 5 doses of the mixture that bracketed 5–95% mortality were tested concurrently with experiments with at least 5 doses of individual mixture components. As before, 60–135 bees of each species were tested depending on species' availability. To test the hypothesis of independent joint action of fenbuconazole with acetamiprid or imidacloprid, we used the computer program PoloMix [49]. Assuming independent joint action of two mixture chemicals, test subjects can die of three possible causes. The first cause is natural mortality, with a probability p
o (a constant). The other two causes of mortality are the probabilities of mortalities for chemical 1 or chemical 2. For the first chemical, the probability of response (p1) is a function of dose D1. Usually, the probit or logit of dose X of chemical 1 is log(D1) (i.e., X1 = log[D1]). For the second chemical, the probability of response (p2) is a function of dose D2. If these three causes of mortality are independent, the probability of death (p) is p = p0+(1−p0)p1+(1−p0)(1−p1) p2. When each “+” sign means “or” and each product means “and,” this equation means that the total probability of death equals death from natural causes (p0), or no death from natural causes (1−p0) and death from the first chemical [e.g., (1−p0)p1], or no death from natural causes or from the first chemical [i.e., (1−p0)(1−p1)], but death from the second chemical [i.e., (1−p0)(1−p1) p2]. The χ2 statistic produced by PoloMix [49] was used to test the hypothesis of independent joint action. This test statistic is calculated by obtaining an estimate for the probability of mortality (p) for several dose levels of the two components and then comparing 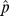 (the estimate of p) with the observed proportion killed at the corresponding dose levels. The three contributions to p are estimated separately. First, p0 is calculated as the proportional mortality observed in the control group. Next, p1 and p2 are estimated from bioassays of chemical 1 and chemical 2, with test statistics estimated from PoloPlus [47].
(the estimate of p) with the observed proportion killed at the corresponding dose levels. The three contributions to p are estimated separately. First, p0 is calculated as the proportional mortality observed in the control group. Next, p1 and p2 are estimated from bioassays of chemical 1 and chemical 2, with test statistics estimated from PoloPlus [47].
Results and Discussion
Responses of O. cornifrons (Table S1)
The hypotheses of parallelism and equality of response of O. cornifrons to the five pesticides were both rejected (P = 0.05). At LD50 (the most reliable point of comparison for dose-response regressions [48]), the decreasing order of toxicity was dimethoate>λ-cyhalothrin>imidacloprid>acetamiprid>phosmet. LDR50's indicated three groups of significantly increasing toxicity. The least toxic group consisted of phosmet, acetamiprid and imidacloprid. λ-cyhalothrin was significantly more toxic than the first group, and the most toxic pesticide, dimethoate, was ∼70 times more toxic than phosmet (the least toxic pesticide). At LD90, the increasing order of toxicity was dimethoate>λ-cyhalothrin>phosmet>imidacloprid>acetamiprid. Responses to acetamiprid and imidacloprid were not significantly different at LD90. Significantly increased susceptibilities to phosmet and λ- cyhalothrin were indicated by their LDR90's. Dimethoate was ∼60 times more toxic than acetamiprid.
Responses of A.mellifera (Table S1)
The pattern of responses for A.mellifera differed considerably from that of O. cornifrons. For A.mellifera, responses were also neither parallel nor equal (P = 0.05). The decreasing order of toxicity at LD50 was imidacloprid>λ-cyhalothrin = dimethoate>phosmet>acetamiprid. LDR50's indicated two levels of decreasing toxicity relative to imidacloprid: λ-cyhalothrin or dimethoate <phosmet. At LD90, the increasing order of toxicity was dimethoate>imidacloprid>λ- cyhalothrin>phosmet>acetamiprid. Dimethoate was ∼1200 times more toxic than acetamiprid as this response level. LDR90's suggested three groups of response: dimethoate>λ- cyhalothrin or imidacloprid>phosmet>acetamiprid.
Comparative Responses of the Pollinator Species (Table S1)
Neither species was consistently more susceptible than the other. Their responses to the two neonicotinoid pesticides were parallel, but not equal. O. cornifrons was significantly more susceptible than A.mellifera to acetamiprid at LD50 (i.e., the LDR50 did not bracket the value 1.0 [47], [48]). At LD50, O. cornifrons ∼12 times more susceptible than A.mellifera. In contrast, A.mellifera was significantly more susceptible to imidacloprid at the 50% response level (LDR50 did not bracket the value 1.0). At LD50, A.mellifera were ∼26 times more susceptible than O. cornifrons. Responses to the two organophosporous pesticides — dimethoate and phosmet — were also not consistent by species. The hypotheses of parallelism and equality were each rejected in tests with these chemicals. For dimethoate, O. cornifrons was 3.7 times more susceptible than honey bees at LD50, but the regression slope (7.62) was so steep for A.mellifera that relative toxicities were reversed at the LD90 (at 90% response, honey bees were 3.2 times more susceptible that O. cornifrons). With phosmet, the slope of the regression line for A.mellifera was very shallow and its line crossed the dose-response line for O. cornifrons at the upper end of response. At LD50, A.mellifera workers were ∼3 times more susceptible than O. cornifrons, but at LD90, O. cornifrons was ∼2 times more susceptible than A.mellifera.
Responses to the only pyrethroid tested, λ- cyhalothrin, were neither parallel nor equal. At LD50, A. mellifera was 3 times more susceptible than O. cornifrons. LD90's for the two species were not significantly different.
Responses to Mixtures (Table 1, Figure 1)
Table 1. Responses of Osmia cornifrons and A. mellifera to to acetamiprid mixed with indar.
| Chemical | Mixture | Speciesa | n | Slope ± S.E. | LD50 b | 95% CL | LDRc | 95% CI* |
| Acetamiprid | only | A | 245 | 1.39±0.41 | 64.6 | 38.1–252 | 1.0 | – |
| Indar | only | A | 330 | Not toxic | ||||
| Acetamiprid | Indar (1∶1) | A | 360 | 3.13±0.53 | 14.3 | 8.5–30.8 | 4.5 | 2.5–8.2 |
| Acetamiprid | only | O | 272 | 0.97±0.18 | 4.0 | 1.1–7.1 | 1.0 | – |
| Indar | only | O | 60 | Not toxic | ||||
| Acetamiprid | Indar (1∶1) | O | 99 | 1.94±0.42 | 2.1 | 1.1–3.2 | 1.9 | 0.95–3.9 |
| Imidicloprid | only | A | 310 | 1.26±0.15 | 0.2 | 0.1–0.3 | 1.7 | 0.92–3.1 |
| Indar | only | A | 330 | Not toxic | ||||
| Imidicloprid | Indar (2∶1) | A | 300 | 1.84±0.33 | 0.3 | 0.1–0.4 | 1.0 | – |
| imidacloprid | only | O | 310 | 1.26±0.15 | 3.8 | 1.7–12.6 | 1.7 | 0.8–3.7 |
| Indar | only | O | 90 | Not toxic | ||||
| Imidicloprid | Indar (2∶1) | O | 522 | 3.11±1.09 | 6.6 | 1.4–9.6 | 1.0 | – |
A is A.mellifera, O is Osmia cornifrons.
LD is expressed as µg/bee.
LDR is higher LD÷lower LD.
If the 95% CI of the LDR includes the value 1.0, the LD's are not significantly different.
Figure 1. (a–d). Effects of mixtures on O. cornifrons and A. mellifera.

Boxes are responses observed and triangles are responses predicted assuming the model of independent joint action (see text). Dose is in units of µg/bee. Figure 1a is for A. mellifera treated with acetamiprid∶indar; Figure 1b is for O. cornifrons treated with acetamiprid∶indar; Figure 1c is for A. mellifera treated with imidacloprid∶indar; Fig. 1d is for O. cornifrons treated with imidacloprid∶indar.
Fenbuconazole was minimally toxic to both species. In combination with acetamiprid, the 1∶1 mixture was ∼5 times more toxic than acetamiprid alone to A. mellifera at LD50 (Fig. 1a). The toxicity of the mixture was 2 times greater than acetamiprid to O. cornifrons at LD50 (Fig. 1b). Fenbuconazole enhanced responses of imidacloprid slightly, but significantly, for both species. Although responses at the LD50 were not significantly different, a greater effect was apparent at higher levels of response (Fig. 1c, d). χ2 values of observed vs. expected values were small but still significant. Rejection of the null hypothesis of independent joint action of the fenbuconazole with either of the two pesticides indicated that significant synergism occurred in both bee species. In contrast, another neonicotinoid – thiacloprid – applied as technical material in 100% ethanol, was synergized 1141- and 559-fold by the addition, respectively, of the fungicides triflumizole and propiconazole, which are in the same class as fenbuconazole [49].
Conclusions
Because of their highly controlled conditions and rigorous experimental design, laboratory bioassays provide the ideal means to estimate comparative responses of A. mellifera and wild bees to pesticides. Our results clearly show that the response of A. mellifera cannot be extrapolated to the response of O.cornifrons. Such results might be expected given the extensive body of information from analogous (and as rigorously designed and statistically analyzed) experiments showing that responses of the Lepidopteran families Tortricidae and Lymantriidae vary significantly among genera, within a single genus, among populations or even among sibling groups of the same genetic strain [47], [50], [51] tested with the same pesticide. Equally rigorously designed bioassays among Apoidea populations of the same species, species of the same genus, and among genera done to evaluate variation at all levels of response are clearly needed. Natural variation in response for a populations of single species to single pesticides, whether applied as pure active ingredient or formulated material, also needs to be estimated [52]. This systematic approach should be part of the overhaul of the pesticide registration process as suggested by Decourtye et al. [46]. Finally, without direct access to raw data, we have no way to compare our results statistically with previously published information from previous experiments with A. mellifera tested with neonicotinoid insecticides [53], let alone explain why these differences occurred. Until such basic comparisons are made possible even for one species such as A. mellifera, bee toxicologists and ecologists will continue to debate possible explanations for different contact and oral LD50's. One solution to this situation would be establishment of a free-access network for raw data from all experiments that have been used in applications for pesticide registrations by the US EPA (US EPA) and the European Union (EU).
Our results further suggest that the current practice of using A. mellifera as a surrogate species for >90,000 other species of non-target insects, including O. cornifrons, in the pesticide registration process as required by the United States EPA is scientifically flawed, and needs significant modifications. Inherent in this registration process is the assumption that the response of A. mellifera can be extrapolated to responses of the many different species of predatory and parasitic arthropods that are important to successful Integrated Pest Management (IPM) programs, but which belong to several distantly related orders. This assumption has been shown to be untenable in other studies: many pesticides labeled as “reduced risk” or “organophosphate replacements” by the United States EPA and that have used toxicity tests with A. mellifera as part of the criteria for these classifications are in fact toxic to non-target insects and disruptive to IPM programs in tree fruit crops [42], [52], [54], [55]. Use of A. mellifera as a surrogate species has also been questioned by members of the European
Union [56], which also currently requires toxicity data for multiple insect species [43]. According to Decourtye et al. [46], 42 studies report deleterious side effects on other bee species despite the fact they passed risk assessment on honey bees. Considering the fact that pollinator species significantly differ in their relative toxicity to pesticides, the United States EPA needs to review its current policy on pollinator toxicity that requires data from contact toxicity (from pesticide residues) studies designed to investigate acute effects of a pesticide chemical (under the registration process) on individual bees [57], in addition to adding studies of sublethal effects to these toxicants [58], [59].
Supporting Information
Responses of O. cornifrons and A. mellifera to pesticides commonly used in apple orchards (complete regression results).
(DOC)
Acknowledgments
We thank Eric Mader and Scott Famous for providing some of the O. cornifrons for experiments.
Funding Statement
The authors thank the USDA NIFA for a SCRI grant (# PEN04398) on sustainable fruit pollination and the State Horticultural Association of Pennsylvania for their financial support. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Calderone NW (2012) Insect Pollinated Crops, Insect Pollinators and US Agriculture: Trend Analysis of Aggregate Data for the Period 1992–2009. PLoS ONE 7 5: e37235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Losey JE, Vaughan M (2006) The economic value of ecological services provided by insects. Bioscience 56: 311–323. [Google Scholar]
- 3. Klein A-M, Vaissière BE, Cane JH, Steffan-Dewenter I, Cunningham SA, et al. (2007) Importance of pollinators in changing landscapes for world crops. Proc R Soc Lond B 274: 303–313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Bosch J, Kemp WP (2002) Developing and establishing bee species as crop pollinators: the example of Osmia spp. (Hymenoptera: Megachilidae) and fruit trees. Bull Entomol Research 92: 3–16. [DOI] [PubMed] [Google Scholar]
- 5. Gallai N, Salles J-M, Settele J, VaissiËre BE (2009) Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol Econ 68: 810–821. [Google Scholar]
- 6. Bosch J, Kemp W, Trostle G (2006) Bee population returns and cherry yields in an orchard pollinated with Osmia lignaria (Hymenoptera: Megachilidae). J Econ Entomol 99: 408–413. [DOI] [PubMed] [Google Scholar]
- 7. Bosch J, Kemp W (2001) How to manage the blue orchard bee. USDA SARE Handbook 5. [Google Scholar]
- 8.Mader E, Spivak M, Evans E (2010) Managing alternative pollinators. USDA SARE Handbook. Available: http://palspublishing.cals.cornell.edu/nra_map.html. Accessed 2013 July 31.
- 9. Maeta V, Kitamura T (1984) How to manage the mame-kobachi, Osmia cornifrons for pollination of the fruit and crops. Ask Co Ltd 1974: 1–16. [Google Scholar]
- 10. Batra SWT (1979) Osmia cornifrons and Pithitis smaragdula, two Asian bees introduced into the United States for crop pollination. Maryland Agricultural Experiment Station, Special Miscellaneous Publication 1: 307–312. [Google Scholar]
- 11. Batra SWT (1998) Hornfaced bees for apple pollination. Am Bee J 138: 364–365. [Google Scholar]
- 12. Biddinger D, Frazier J, Frazier M, Rajotte E, Donovall L, et al. (2009) Solitary bees as alternative pollinators in Pennsylvania fruit crops. Penn Fruit News 89: 84–94. [Google Scholar]
- 13. Biddinger D, Rajotte E, Joshi NK, Ritz A (2011) Wild bees as alternative pollinators. Fruit Times 30: 1–4. [Google Scholar]
- 14. Maeta Y, Kitamura T (1981) Pollinating efficiency by Osmia cornifrons (Radoszkowski) in relation to required number of nesting bees for economic fruit production. Honeybee Science 2: 65–72. [Google Scholar]
- 15. Park MG, Orr MC, Danforth BN (2010) The role of native bees in apple pollination. NY Fruit Quart 18: 21–25. [Google Scholar]
- 16. Ritz A, Biddinger D, Rajotte E, Sahli H, Joshi N (2012) Quantifying the efficacy of native bees for orchard pollination in Pennsylvania to offset the increased cost and decreased reliability of honey bees. Penn Fruit News 92: 6–66. [Google Scholar]
- 17.Joshi NK, Biddinger D, Rajotte EG (2011) A survey of apple pollination practices, knowledge and attitudes of fruit growers in Pennsylvania. 10th International Pollination Symposium, Puebla, Mexico.
- 18. Biddinger DJ, Ngugi H, Frazier J, Frazier M, Leslie T, Donovall LR (2010) Development of the mason bee, Osmia cornifrons, as an alternative pollinator to honey bees and as a targeted delivery system for biological control agents in the management of fire blight. Penn Fruit News 90: 35–44. [Google Scholar]
- 19.Biddinger D, Rajotte E, Joshi NK (2011) Wild bees as alternative pollinators in Pennsylvania apple orchards. Proceedings, the Great Lakes Fruit, Vegetable and Farm Market EXPO and the Michigan Greenhouse Growers Expo, Michigan. Available: http://www.glexpo.com/summaries/2011summaries/webCropPollination.pdf. Accessed 2013 July 31.
- 20.vanEngelsdorp D, Pettis J, Rennich K, Rose R, Caron D, et al. (2012) Preliminary results: honey bee colonies losses in the U.S., Winter 2011–2012. Available: http://beeinformed.org/2012/05/winter2012. Accessed 2013 July 31.
- 21. vanEngelsdorp D, Evans JD, Saegerman C, Mullin C, Haubruge E, et al. (2009) Colony Collapse Disorder: A descriptive study. PLoS ONE 4: e6481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. vanEngelsdorp D, Hayes J Jr, Underwood RM, Pettis J (2008) A survey of honey bee colony losses in the U.S., fall 2007 to spring 2008. PLoS ONE 3: e4071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.National Research Council (2006) Status of pollinators in North America. Washington, DC: National Academies Press. 322 p.
- 24. Potts SG, Biesmeijer JC, Kremen C, Neumann P, Schweiger O, et al. (2010) Global pollinator declines: trends, impacts and drivers. Trends Ecol Evol 25: 345–353. [DOI] [PubMed] [Google Scholar]
- 25. Henry M, Béguin M, Requier F, Rollin O, Odoux J, et al. (2012) A common pesticide decreases foraging success and survival in honey bees. Science 336: 348–350. [DOI] [PubMed] [Google Scholar]
- 26. Whitehorn PR, O'Connor S, Wackers FL, Goulson D (2012) Neonicotinoid pesticide reduces bumblebee colony growth and queen production. Science 336: 351–352. [DOI] [PubMed] [Google Scholar]
- 27. Krupke CH, Hunt GJ, Eitzer BD, Andino G, Given K (2012) Multiple routes of pesticide exposure for honey bees living near agricultural fields. PLoS ONE 7: e29268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Laycock I, Lenthall K, Barratt A, Cresswell J (2012) Effects of imidacloprid, a neonicotinoid pesticide, on reproduction in worker bumble bees (Bombus terrestris). Ecotoxicology Doi:10.1007/s10646-012-0927-y [DOI] [PubMed] [Google Scholar]
- 29. Cresswell JE (2011) A meta-analysis of experiments testing the effects of a neonicotinoid insecticide (imidacloprid) on honey bees. Ecotoxicology 20: 149. [DOI] [PubMed] [Google Scholar]
- 30. De la Rúa P, Jaffé R, Dall'Olio R, Muñoz I, Serrano J (2009) Biodiversity, conservation and current threats to European honeybees. Apidologie 40: 263–284. [Google Scholar]
- 31. Chauzat M-P, Faucon J-P, Martel A-C, Lachaize J, Cougoule N, et al. (2006) A survey of pesticide residues in pollen loads collected by honey bees in France. J Econ Entomol 99: 253–262. [DOI] [PubMed] [Google Scholar]
- 32. Mullin CA, Frazier M, Frazier JL, Ashcraft S, Simonds R, et al. (2010) High levels of miticides and agrochemicals in North American apiaries: Implications for honey bee health. PLoS ONE 5 3: e9754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Frazier J, Mullin C, Frazier M, Ashcraft S (2011) Managed pollinator coordinated agricultural project: Pesticides and their involvement in colony collapse disorder. Am Bee J 151 8: 779–784. [Google Scholar]
- 34. Cox-Foster DL, Conlan S, Holmes EC, Palacios G, Evans JD, et al. (2007) A metagenomic survey of microbes in honey bee colony collapse disorder. Science 318: 283–287. [DOI] [PubMed] [Google Scholar]
- 35. Singh R, Levitt AL, Rajotte EG, Holmes EC, Ostiguy N, et al. (2010) RNA Viruses in Hymenopteran Pollinators: Evidence of Inter-Taxa Virus Transmission via Pollen and Potential Impact on Non-Apis Hymenopteran Species. PLoS ONE 5 12: e14357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Bromenshenk JJ, Henderson CB, Wick CH, Stanford MF, Zulich AW, et al. (2010) Iridovirus and microsporidian linked to honey bee colony decline. PLoS ONE 5: e13181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Ciarlo TJ, Mullin CA, Frazier JL, Schmehl DR (2012) Learning Impairment in Honey Bees Caused by Agricultural Spray Adjuvants. PLoS ONE 7 7: e40848. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Evison SEF, Roberts KE, Laurenson L, Pietravalle S, Hui J, et al. (2012) Pervasiveness of Parasites in Pollinators. PLoS ONE 7 1: e30641 doi:10.1371/journal.pone.0030641 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Andersson GKS, Rundlöf M, Smith HG (2012) Organic Farming Improves Pollination Success in Strawberries. PLoS ONE 7 2: e31599 doi:10.1371/journal.pone.0031599 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Anonymous (2012) Pennsylvania Tree Fruit Production Guide, 324 p. Available: http://agsci.psu.edu/tfpg. Accessed 2013 July 31.
- 41. Hull LA, Joshi NK, Zaman F (2009) Management of internal feeding lepidopteran pests in apple, 2008. Arthropod Mgmt Tests 34 doi:10.4182/amt.2009.A8 [Google Scholar]
- 42. Biddinger DJ, Hull LA (1995) Effects of several types of insecticides on the mite predator, Stethorus punctum (Coleoptera: Coccinellidae), including insect growth regulators and abamectin. J Econ Entomol 88: 358–366. [Google Scholar]
- 43.Summary of a SETAC Pellston Workshop. Pensacola FL (USA): Society of Environmental Toxicology and Chemistry (SETAC). 45 p.
- 44. Cresswell JE, Page CJ, Uygun MB, Holmbergh M, Li Y, et al. (2012) Differential sensitivity of honey bees and bumble bees to a dietary insecticide (imidacloprid). Zoology 115: 365–371. [DOI] [PubMed] [Google Scholar]
- 45. Cresswell JE, Laycock I (2012) Towards the comparative ecotoxicology of bees: the response-response relationship. Julius-Kühn-Archiv 437: 55–60. [Google Scholar]
- 46. Decourtye A, Henry M, Desneux N (2013) Overhaul pesticide testing on bees. Nature 497: 168. [DOI] [PubMed] [Google Scholar]
- 47.LeOra Software, PoloPlus (2005) Available: http://www.LeOraSoftware.com. Accessed 2013 July 31.
- 48.Robertson JL, Russell RM, Preisler HK, Savin NE (2007) Bioassays with Arthropods. Boca Raton: CRC Press. 199 p.
- 49.LeOra Software, PoloMix (2005) Available: http://www.LeOraSoftware.com. Accessed 2013 July 31.
- 50. Iwasa T, Motoyama N, Ambrose JT, Roe RM (2004) Mechanism for the differential toxicity of neonicotinoid insecticides in the honey bee, Apis mellifera . Crop Prot 23: 371–378. [Google Scholar]
- 51. Robertson JL, Boelter LM, Russell RM, Savin NE (1978) Variation in response to insecticides by Douglas-fir tussock moth, Orgyia pseudotsugata (Lepidoptera: Lymantriidae), populations. Can Entomol 110: 325–28. [Google Scholar]
- 52. Robertson JL, Preisler HK, Ng SS, Hickle L, Gelernter W (1995) Natural variation: A complicating factor in bioassays with chemical and microbial insecticides. J Econ Entomol 88: 1–10. [Google Scholar]
- 53.Hopwood J, Vaughan M, Shepherd M, Biddinger D, Mader E, et al.. (2012) Xerces Society for Invertebrate Conservation 32 p.
- 54. Brunner JF, Dunley JE, Doerr M, Beers EH (2001) Effects of pesticides on Colpoclypeus florus (Hymenoptera: Eulophidae) and Trichogramma platneri (Hymenoptera: Trichogrammatidae), parasitoids of leafro00llers in Washington. J Econ Entomol 94: 1075–1084. [DOI] [PubMed] [Google Scholar]
- 55.Biddinger DJ, Hull LA, LT (2010) Wooly apple aphid control in Pennsylvania apple orchards and impact of Delegate on Aphelinus mali 84th Orchard Pest & Disease Management Conference Abstract, Jan. 14–16, 2010. Portland, OR. Available: http://entomology.tfrec.wsu.edu/wopdmc. Accessed 2013 July 31.
- 56. Jones VP, Steffan SA, Hull LA, Biddinger DJ (2010) Effects of the loss of organophosphate pesticides in the U.S.: opportunities and needs to improve IPM programs. Outlooks on Pest Management 21: 161–66. [Google Scholar]
- 57.Roessink I, van der Steen JJM, Kasina M, Gikungu M, Nocelli RCF (2011) Is the European honey bee (Apis mellifera mellifera) a good representative for other pollinator species? SETAC Europe 21st Annual Meeting: Ecosystem Protection in a Sustainable World: a Challenge for Science and Regulation. Milan, Italy, 15–19 May 2011.
- 58. http://www.epa.gov/pesticides/ecosystem/pollinator/then-now.html. Accessed 2013 July 31.
- 59. Desneux N, Decourtye A, Delpuech JM (2007) The sublethal effects of pesticides on beneficial arthropods. Ann. Rev. Entomology 52: 81–106. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Responses of O. cornifrons and A. mellifera to pesticides commonly used in apple orchards (complete regression results).
(DOC)