

Abstract
An extreme halophilic bacterium was isolated from solar saltern samples and identified based on biochemical tests and 16S r RNA sequencing as Chromohalobacter sp. strain TVSP101. The halophilic protease was purified using ultrafiltration, ethanol precipitation, hydrophobic interaction column chromatography and gel permeation chromatography to 180 fold with 22% yield. The molecular mass of the protease determined by SDS PAGE was 66 kDa. The purified enzyme was salt dependent for its activity and stability with an optimum of 4.5 M NaCl. The optimum temperature for maximum protease activity was 75ºC. The protease was optimally active at pH 8 and retained more than 80% of its activity in the range of pH 7-10. Sucrose and glycine at 10% (w/v) were the most effective osmolytes, retained 100% activity in the absence of NaCl. The activity was completely inhibited by ZnCl2 (2 mM), 0.1% SDS and PMSF (1mM). The enzyme was not inhibited by 1mM of pepstatin, EDTA and PCMB. The protease was active and retained 100% it activity in 10% (v/v) DMSO, DMF, ethanol and acetone.
Keywords: Chromohalobacter sp. TVSP101, halothermophilic protease, purification, organic solvents, osmolytes
INTRODUCTION
Halophilic microorganisms require very high salt (2 to 5 M NaCl) concentrations for growth and are found in salterns and hypersaline lakes. Many extreme and moderate halophiles have been isolated and investigated for possible biotechnological applications. These include the production of β-carotene, polyhydroxy alkanoates, enzymes and compatible solutes, enhanced oil recovery and degradation of toxic chemicals that can pollute hypersaline habitats. Additionally, halophiles produce exozymes such as amylases, proteases and nucleases of potential commercial values (1,11,12). Halophilic proteins are distinguished from their homologous proteins by exhibiting remarkable instability in solutions with low salt concentrations and by maintaining soluble and active conformations in high concentrations of salt upto 5 M NaCl (13,24). There are number of enzymes of this type produced by some halophilic microorganisms that have optimal activity at high salinities and could therefore used in many harsh industrial processes where the concentrated salt solutions used would otherwise inhibit many enzymatic conversions (7,12,23).
Halophilic microorganisms play an essential role in various fermentation processes that occur in the presence of salt. The high salt tolerance of extreme halophiles enables their cultivation under non-sterile and thus cost-reducing conditions (22). So far, several wellknown proteases such as bromelain, papain, and pepsin have been used as biocatalysts of protein hydrolysis in fish sauce fermentation (3,9). However, most of these proteases were not sufficiently stable in the presence of high salt concentration. Halophilic organisms have evolved in saline environment and are able to overcome the deterious effects of salt, and their enzymes require salt, retaining activity in salt upto saturating concentration. Some halophilic proteases have been purified, characterized, showed maximum activity at neutral pH at a temperature ranging from 55-66ºC (15,31). Halophilic proteases have wide application in the processing of food, leather and detergents (1,13,24). Given the potential applications of halophilic proteases and the need to search for new halophilic organisms producing maximum enzyme production is a continuous process. We have reported the optimum culture conditions for the production of halophilic protease from Chromohalobacter sp. TVSP101 (33). In the present study we report the purification and characterization of an extremely halophilic, thermophilic protease from a newly isolated Chromohalobacter sp. TVSP101.
MATERIALS AND METHODS
Organism
Chromohalobacter sp. TVSP101 was isolated from solar saltern samples of Tuticorin, Tamilnadu, India. This extreme halophilic bacterium has been deposited in the National Centre for Industrial Microorganisms, Pune, India with accession number NCIM: 5237.
Growth conditions and Media composition
The bacterial strain TVSP101 was aerobically cultured at 40ºC in a tris-based medium containing (g/L): NaCl 233.6; MgCl2 10; KC15; Trisodium citrate 3; and peptone 10; skim milk powder 10 (14). The pH was adjusted to 7.2. The strain TVSP101 was grown in 250 ml conical flasks containing 50 ml of above mentioned media. The media was inoculated with 1 ml of a week old culture of Chromohalobacter sp. TVSP101 and incubated at 40ºC in an incubator shaker at 200 rpm for 96 h.
16S r DNA sequencing
The culture was allowed to grow for 4 days on 4 M NaCl containing LB agar at 40ºC. Single colony was resuspended in 20 μl of tris-HCl–EDTA saline (pH 7.2). The bacterial suspension was incubated for 10 min at 95ºC and centrifuged at 18,600 X g for 2 min. The supernatant was transferred to fresh tubes and used as total genomic DNA samples. The bacterial 16S rRNA gene was amplified from the total genomic DNA using universal eubacteria specific primers viz: 16F27 (51 CCA GAG TTT GAT CMT GGC TCA G 31) and 16R1 525XP (51 TTC TGC AGT CTA GAA GGA GGT GWT CCA GCC 31), which yielded a product of approximately 1500 base pairs. The PCR conditions was: 35 cycles of 95ºC denaturation for 1 min, annealing at 55ºC for 1 min and extension at 72ºC for 1 min, and final extension at 72ºC for 10 min. The PCR product was purified by PEG-NaCl precipitation as described earlier (29). Briefly, the PCR product was mixed with 0.6 volumes of PEG-NaCl solution (20% PEG 6000, 2.5 M NaCl) and incubated for 10 min. The pellet was washed twice with 70% ethanol and dried under vacuum which was then resuspended in glass distilled water at concentration of >0.1pmol/ml. The purified product was directly sequenced using Big Dye terminator kit (Applied Biosytems, Inc., Foster City, CA) as described earlier (25). The sequencing reactions were run on AB-PRISM automated sequencer (ABI-3730 DNA analyzer). The nucleotide sequence analysis of the sequence was done at BLAST-n site at NCBI server (www.Ncbi.nlm.nih.gov/BLAST). The alignment of the sequences were done using CLUSTAL W program V1.82 at European Bioinformatics site (www.ebi.ac.uk/clustalw). The analysis of 16S rRNA gene sequence was done at Ribosomal Database project (RDP) II (http//:rdp.cme.msu.edu). The sequence was refined manually after crosschecking with the raw data to remove ambiguities and was submitted to the Gene Bank with accession number DQ375807. The phylogenetic tree was constructed using the aligned sequences by the neighbor joining method using Kimura-2 parameter distances in MEGA2.1 software (18).
Assay of protease
Protease activity was determined by the method described by Brock et al. (4). Azocasein (Sigma) (0.8%, 1ml) dissolved in 50 mM Tris buffer (pH 8.0) contained 4.5 M NaCl was pipetted into eppendorf tubes (capacity 2.0 ml) and preincubated at 75ºC. The reaction was initiated by addition of 50 μl of enzyme solution, and performed at 75ºC for 15 min. An equal volume of 10% (w/v) trichloroacetic acid was added to terminate the reaction and the mixture allowed to stand at room temperature for 30 min and then centrifuged at 13000 x g for 10 min. The absorbance of the supernatant was determined at 450 nm and corrected by subtracting the absorbance of a blank sample lacking added substrate. One unit (U) of azocaseinase activity was defined as the amount of enzyme that produced a change in absorbance (0.01 per min) at 450 nm at 75ºC under the standard assay condition.
Enzyme Purification
Cells were harvested by centrifugation at 10,000 x g for 10 min at 4ºC when the culture reached the stationary growth phase and this supernatant was used for purification of extracellular protease. One liter of culture supernatant was concentrated to 50 ml through ultrafiltration unit (MWCO 50-kDa, Nalgene). The concentrated enzyme was slowly mixed with 100 ml chilled absolute ethanol and allowed to stay for 2 h at -10ºC. The precipitate was recovered in 10 ml of 50 mM Tris buffer (pH 8) containing 4.5 M NaCl and dialysed against the same buffer at 4-ºC overnight. Hydrophobic interaction column chromatography was performed in a Pharmacia glass column (2X15 cm) containing 20 ml of phenyl sepharose 6B column, equilibrated with 50 mM Tris buffer pH 8 saturated with 2 M NaCl. The dialysed sample was loaded onto the column at a flow rate of 0.5 ml/min. The column was washed with 40 ml of 50 mM of Tris buffer pH 8 containing 10% NaCl until no absorbance was detected in the elute at 280 nm. The adsorbed proteins were then eluted with 40 ml of 10% sucrose and collected fractions each of 2 ml. The five fractions containing maximum protease activity were pooled, concentrated and dialysed overnight against 50 mM Tris buffer pH 8 containing 10% sucrose.
Gel filtration chromatography was performed using G-100 (1X65 cm) equilibrated with 50 mM Tris buffer (pH 8, 10% sucrose). The above concentrated sample was loaded on to the column followed by elution with the same buffer at a flow rate 0.1ml/min and fraction size of 1ml. The eluted active fractions were pooled and added required amount of NaCl to attain 4 M NaCl and dialyzed overnight against 50 mM Tris buffer of pH 8 containing 4 M NaCl. This purified enzyme was used for further biochemical characterization. Protein content was determined by method of Lowry et al. (21) with crystalline bovine serum albumin (Sigma Chemical) as the standard.
Gel Electrophoresis and activity staining
Polyacrylamide gel electrophoresis (PAGE) was done essentially as described by Laemmli (19) with 8% acrylamide. Protein bands were visualized by staining with 0.2% Coomassie brilliant blue R-250. Activity staining of protease in slab gels was done with agar sheets containing 1% casein (Hammerstein) with 4.5 M NaCl as replica plates. The purified enzyme sample after overnight dialysis against buffer containing 10% sucrose, the purified enzyme sample was loaded on non-denaturing electrophoresis gels prepared with same concentration of osmoprotectant sucrose (5). The bands of protein that were associated with protease activity were seen as clear zones on a dark background on the casein-agar replica sheet.
The effect of NaCl, pH and temperature on the purified protease
The azocaseinase activity of the purified enzyme was measured in different concentrations of NaCl (0 to 5 M) at 75ºC in 50 mM tris buffer of pH 8.0. For optimal pH, the purified enzyme was incubated at different buffers: 50 mM of citrate-phosphate for pHs from 5-6, 50 mM Tris buffer for pHs from 6 to 8, and 50 mM glycine-NaOH for pHs from 8 to 12 and assayed the protease activity at 75ºC in presence of 4.5 M NaCl. The effect of temperature on protease activity was measured from 30 to 90ºC in 50 mM Tris buffer of pH 8.0 containing 4.5 M. NaCl using azocasein as a substrate.
The effect of metal ions, inhibitors and osmolytes on purified protease
The effect of various metal ions, detergents and inhibitors was tested on the activity of purified enzyme at 75ºC in Tris buffer (pH 8) containing 4.5 M NaCl using azocasein as a substrate. The protease activity of the purified enzyme was measured at various concentrations (w/v) of osmolytes: betaine, sucrose, glycerol and mannitol in the absence of NaCl using azocasein as a substrate at 75ºC and pH 8 of 50 mM Tris buffer.
The effect of organic solvents on purified protease
The azocaseinase activity of the purified enzyme was measured at various concentrations of organic solvents (v/v) at 75ºC in 50 mM Tris buffer (pH 8) containing 4.5 M NaCl. Ethanol, acetone, DMF and DMSO were used as organic solvents from 5-20% (v/v) in the reaction mixture.
RESULTS
Isolation and Identification of Bacterial culture
A halophilic bacterial strain TVSP101 producing extreme halophilic, thermophilic extracellular protease was isolated from salt sample of solar evaporation pond. The bacterium was Gram negative, rod shaped and pigmented light pinkish. No growth was observed in 0.5 M NaCl and required minimum 1M of NaCl for its growth. Optimum growth was observed at 4 M NaCl concentration. The strain TVSP101 was able to hydrolyze gelatin, casein, starch and utilized urea and citrate.
A total of 1491 bp product of PCR amplified 16S rRNA gene was determined for this isolate, which corresponds to more than 99% of the gene sequence. In the analyses at NCBI and RDP sites, it showed homology to 16S rRNA gene sequences from Chromohalobacter sp. In the phylogenetic analysis, the sequence formed a cluster within Chromohalobacter israelensis with 100% bootstrap value, confirming the identity of the isolate as strain of this species (Fig. 1). The halophilic strain TVSP101 was identified, as Chromohalobacter sp. TVSP101 and the NCBI accession number of the 16S r RNA gene sequences determined in this study is DQ375807.
Figure 1.

Phylogenetic tree of the strain TVSP101 and related organisms were aligned based on 16s r DNA sequences (neighbor-joining tree). Scale bar: number of nucleotide changes per sequence position. The number at nodes show the bootstrap values obtained with 1,000 resampling.
The proteolytic activity was noticed in the fermentation broth after 60 h and the activity reached the maximum level in the stationary growth phase (96 h) (14). The protease was unquestionably produced extracellularly; removal of cells by centrifugation, extensive washing of the cell paste, and lysing the cells resulted in minimal proteolytic activity.
Purification of extracellular protease
The extracellular protease from Chromohalobacter sp. TVSP101 was purified to 180 fold with a yield of 22% (Table 1), using a series of purification steps that included ultrafiltration (MWCO 50 kDa; Nalgene), ethanol precipitation, hydrophobic interaction column chromatography and gel permeation chromatography. The purified enzyme was homogenous showing a single protein band on SDS-PAGE with a molecular mass of 66 kDa when compared to authentic standards (Fig. 2a). Activity staining of crude and purified enzyme showed that only one extra cellular protease is secreted (Fig. 2b).
Table 1.
Purification of protease from Chromohalobacter sp. TVSP101.
| Purification step | Volume (ml) | Total Protein (mg) | Total activity(U) | Specific activity (U/mg) | Purification (fold) | Yield (%) |
|---|---|---|---|---|---|---|
| Culture mediumSupernatant | 1000 | 3300 | 96,000 | 29 | 1 | 100 |
| Protein concentratorNalgene (50kDa) | 50 | 212 | 74200 | 350 | 12 | 77 |
| Ethanol precipitate | 10 | 126 | 57280 | 455 | 16 | 60 |
| Phenylsepharose 6B column | 10ml | 62 | 52900 | 853 | 30 | 55 |
| Gel permeation G-100 | 8ml | 4 | 21424 | 5225 | 180 | 22 |
Figure 2.

(a) The CBB stained 8% SDS-PAGE of purified protease. Lane 1. Relative molecular mass standards. Myosin, Rabbit Muscle M W 205 kDa; Phosphorylase b 97 kDa; Bovine serum albumin-66 kDa; Ovalbumin-43 kDa; Carbonic anhydrase 29 kDa; Lane 2. Purified protease. (b) Activity staining in casein agar replica plate containing 4.5 M NaCl. Details are given in the materials and methods.
The effect of NaCl, pH, temperature and thermal stability on purified protease
The azocaseinase activity was measured at different NaCl concentrations (Fig. 3). The protease was active in broad range of NaCl concentrations from 1 to 5 M with optimum activity at 4.5 M NaCl. In the absence of NaCl, the enzyme lost the activity completely and irreversibly. The enzyme was active in broad pH range of 7-10 with optimum activity at pH 8 (Fig. 4). The enzyme retained 82% of activity at pH 10 showing alkali tolerance. The protease was thermoactive in the temperature range of 60-80ºC with optimum at 75ºC (Fig. 5). Temperature kinetics of the protease suggests that the enzyme activity increases sharply from 60 to 75ºC, followed by a decline after 80ºC. The protease was stable in 4.5 M NaCl at 60 and 70ºC for 2hrs. The enzyme at 75ºC was stable up to 60 min and after 120 min it retained 68% of the activity. The purified enzyme in presence of 4.5 M NaCl at room temperature was stable for 20 days and stable for 45 days when stored at - 4º C. The purified enzyme was stable at minimum of 2 M NaCl at 4ºC for 12 hours but 50% activity was lost after two days (data not shown).
Figure 3.
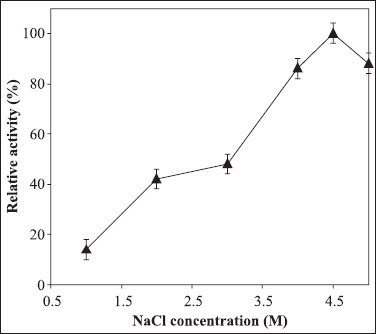
The effect of NaCl on azocaseinase activity of purified protease from Chromohalobacter sp. TVSP101 in 50 mM Tris buffer of pH 8 at 75°C. Results represent the means of three separate experiments, and indicate ±1 standard deviation.
Figure 4.
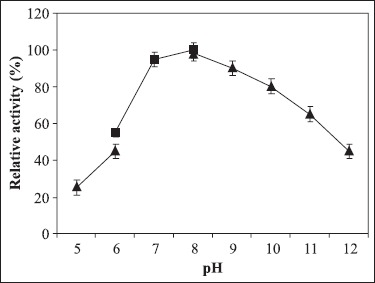
The effect of pH on azocaseinase activity of purified protease from Chromohalobacter sp. TVSP101 in 4.5 M NaCl, at 75ºC. Citrate-phosphate Buffer (50 mM) for pH values between 5 and 6, Tris-HCl buffer (50 mM) for pH values between 6 and 8, and glycine-NaOH buffer (50 mM) for pH values between 8 and 12. Results represent the means of three separate experiments, and bars indicate ±1 standard deviation.
Figure 5.
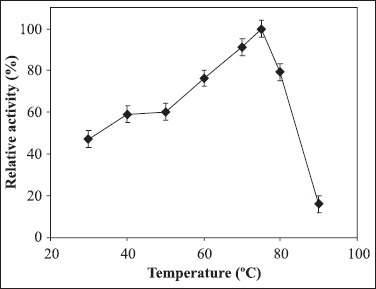
The effect of temperature on azocaseinase activity of purified protease from Chromohalobacter sp. TVSP101 in 50 mM Tris buffer pH 8 containing 4.5 M NaCl. Results represent the means of three separate experiments, and bars indicate ± standard deviation.
The effect of osmolytes
The effect of osmolytes in different concentrations in absence of NaCl was tested (Fig. 6). The purified enzyme retained 100% activity in 10 % (w/v) of glycine and sucrose but it required 20% of glycerol to retain complete activity. Betaine at 10% was not effective as other osmolytes but at 25% it retained 80% activity.
Figure 6.

The effect of osmolytes on azocaseinase activity of purified protease from Chromohalobacter sp. TVSP101 at 75°C in 50 mM Tris buffer of pH 8, in the absence of NaCl. (▄) Betaine, (▲) Glycerol, (▴) Glycine and (∆) Sucrose (w/v). Results represent the means of three separate experiments, and bars indicate ±1 standard deviation.
The effect of inhibitors and metal ions
Zinc inactivated the enzyme completely at 2 mM concentration. Nearly 60 % activity was lost in the presence of 2 mM of Mn2+, Hg2+ and Fe2+(Table 2). None of the metal ions stimulated the proteolytic activity. PMSF, a potent inhibitor of serine proteases, inactivated the purified protease completely and nearly instantaneously. At 0.1% SDS concentration complete activity was lost, hence it was much more effective denaturant. The EDTA and 1, 10- phenathroline did not inhibit the enzyme (Table 3).
Table 2.
Effect of metal ions on purified proteolytic enzyme from Chromohalobacter sp. TVSP101.
| Metal ions (2 mM) | Relative activity (%) |
|---|---|
| BaCl2 | 83 |
| CuSO4 | 80 |
| CaCl2 | 85 |
| CoCl2 | 61 |
| MgCl2 | 94 |
| MnCl2 | 40 |
| FeCl2 | 42 |
| HgCl2 | 43 |
| ZnCl2 | 00 |
| KCl | 100 |
Table 3.
Effect of inhibitors and detergents on purified proteolytic enzyme from Chromohalobacter sp. TVSP101.
| Inhibitors/Detergents | Relative activity (%) |
|---|---|
| SDS (0.1%) | 00 |
| Urea (8M) | 18 |
| PMSF (1mM) | 00 |
| 1,10 Phenanthroline (1mM) | 84 |
| EDTA (1mM) | 64 |
| EDTA (10 mM) | 62 |
| DTT (1mM) | 72 |
| Chymostatin (1mM) | 00 |
| Mercaptoethanol (0.1 %) | 82 |
| PCMB (1mM) | 84 |
The effect of organic solvents
High salt solutions are indirectly related to non-aqueous environments, as the salts act to reduce the water activity of the solution as well as to compete with the enzyme for available water. It was of interest, therefore, to investigate the effect of organic solvents on the function of the halophilic protease. The protease activity was determined in different concentrations of organic solvents acetone, ethanol, DMF and DMSO in presence of 4.5 M NaCl (Fig. 7). In presence of 10% (v/v) DMSO complete enzyme activity was retained compared with 25% loss in 10% (v/ v) DMF. Gradual loss of activity was observed with increase in concentrations of acetone and ethanol.
Figure 7.
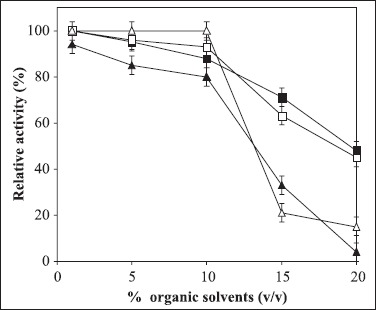
The effect of organic solvents on azocaseinase activity of purified protease from Chromohalobacter sp. TVSP101 at 75°C in 50 mM Tris buffer of pH 8 containing 4.5 M NaCl. (∆) DMSO, (▄) Acetone, (▴) ethanol and (▄) DMF (v/v). Results represent the means of three separate experiments, and bars indicate ±1 standard deviation.
DISCUSSION
Proteases from halophilic organisms have been characterized in a limited extent. Halophilic Chromohalobacter sp. TVSP101 grows and secretes the protease in the presence of high NaCl concentrations. Bacteria of genus Chromohalobacter grow on 0.9% NaCl with optimum 7.5 to 10% (w/v) NaCl (2), whereas Chromohalobacter sp. strain TVSP101 required minimum of 1 M NaCl and optimum of 4 M NaCl. In all cases, the halophilic enzymes require at least 4 M NaCl or KCl for optimal activity and stability (14,15). Activity staining of the crude enzyme indicates that only one extra-cellular protease is secreted during the stationary phase of Chromohalobacter sp. TVSP101. The protease exhibited halophilic characteristics with loss in activity below 1 M NaCl. Extracellular enzymes from microorganisms of the family Halobacteriaceae usually irreversibly lose activity at low ionic strength (20,28). This is, for instance, the case for amylase from Natronococcus sp. strain Ah-36 (17) and proteases from different strains of the genus Halobacterium (15,28). The protease was stable in 10% sucrose; the same was used during purification steps of hydrophobic interaction chromatography and gel permeation as the salt interferes during the purification process. Sucrose, glycerol, and other alcohols and amino acids or their derivatives have been used as osmolytes. These osmotically active substances maintain the halophilic cells in positive water balance and are compatible with the cellular metabolism (6,35). Glycerol and other osmolytes were reported to stabilize other halophilic enzymes (8,26). The purification procedure was very similar to those used for other halophilic enzymes (10,14,5,31,36) indicating the same halophilic behavior. The native molecular mass of Chromohalobacter sp. TVSP101 extracellular protease estimated by SDS -PAGE (66 kDa) is similar to that of protease from Halobacterium halobium (16) but higher than those determined for other halophilic proteases: 42 kDa for halolysins 172 P1 (15), 49 kDa for Natronobacterium sp (36), 60 kDa for Halobacterium halobium protease TuA 4 (30), and 45 kDa for Natrialba magadii extracellular serine protease (10); but it was Vidyasagar, M. et al. lower than of extracellular protease of Natronococcus occultus 130 kDa (31).
Extracellular protease from Chromohalobacter sp. TVSP101 is a serine protease as it is inhibited by PMSF and chymostatin. It is rather thermophilic, showing maximal activity at 75ºC in 4.5 M NaCl. Most of the halophilic proteases reported have their temperature optimum in the range of 37 to 60ºC; 37ºC for protease from Halobacterium halobium (14), 40ºC for protease of Halobacterium sp. PB407 (34), and 60ºC for extracellular protease from N. occultus (31). The solvent effect is not universal and several solvents destabilize the enzyme. Loss of activity in DMF was higher as compared to other organic solvents. Solvents such as 1, 4 - dioxane and DMF also destabilize the enzyme relative to aqueous buffer at all concentrations of NaCl as reported by Kim and Dordick (16). Halophilic enzymes display optimal activity and stability in solutions containing high salt concentration, meaning low water activity (27). For this reason, they are excellent candidates as biocatalyst in aqueous-organic and organic solvent reactions. Particularly, halophilic proteases may have applications in protease-catalyzed peptide synthesis. These results contribute to fundamental and applied aspects of extremozymes and show the potential of halophilic proteins as biocatalysts in aqueous-organic media.
In general, all detergent compatible enzymes should be alkaline and thermostable in nature because of high pH (8-12) and temperatures (50-70ºC) in the laundry environments. Though the present protease is optimally active at 75ºC, but it retains more than 75% activity in the temperature range of 60-80ºC, thus could have potential use as detergent additive in presence of high salt concentrations. The protease of the strain TVSP101 also showed high activity at high salt concentration of 4 - 4.5 M NaCl. These results lead to the consideration for utilization of this bacterial enzyme in fish sauce production. To the best of our knowledge, no halophilic protease have been purified and characterized from Chromohalobacter sp. This is the first report for isolation of pure extracellular extreme halothermophilic serine protease from Chromohalobacter sp.
ACKNOWLEDGEMENTS
We wishes to thank the Department of Science and Technology (DST), Government of India, New Delhi, India for financial assistance.
REFERENCES
- 1.Alqueres S.M.C., Almeida R.V., Clementino M.M., Vieira R.P., Almeida W.I., Cardoso A.M., Martins O.B. Exploring the biotechnological applications in the archaeal domain. Braz. J. Microbiol. 2007;38:398–405. [Google Scholar]
- 2.Arahal D.R., Garcia M.T., Vargas C., Canovas D., Nieto J.J., Ventosa A. Chromohalobacter salexigens sp. nov., a moderately halophilic species that includes Halomonas elongata DSM 3043 and ATCC 33174. Int. J. Sys. Evol. Microbiol. 2001;51:1457–1462. doi: 10.1099/00207713-51-4-1457. [DOI] [PubMed] [Google Scholar]
- 3.Beddow C.G., Ardeshir A.G. The production of soluble fish protein solution for use in fish sauce manufacture. I. The use of added enzymes. J. Food. Technol. 1979;14:603–612. [Google Scholar]
- 4.Brock F.M., Frosberg C.W., Buchanan-Smith J.G. Proteolytic activity of rumen microorganisms and effect of proteinase inhibitors. Appl. Environ. Microbiol. 1982;44:561–569. doi: 10.1128/aem.44.3.561-569.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Cadenas Q., Engel P.C. Activity staining of halophilic enzymes: Substitution of salt with zwitterions in non-denaturing electrophoresis. Biochem Mol. Biol. Int. 1994;33:785–792. [PubMed] [Google Scholar]
- 6.Da Costa M.S., Santos H., Galinski E.A. An overview of the role and diversity of compatible solutes in bacteria and archaea. In: Antranikian G, editor. Biotechnology of Extremophiles. Berlin Heidelberg New York: Advances in Biochemical engineering/Biotechnology, Vol 61. Springer; 1998. pp. 117–153. [DOI] [PubMed] [Google Scholar]
- 7.Dodia M.S., Joshi R.H., Patel R.K., Singh S.P. Characterization and stability of extracellular alkaline protease from halophilic and alkaliphilic bacteria isolated from saline habitat of costal Gujarat, India. Braz. J. Microbiol. 2006;37:276–282. [Google Scholar]
- 8.Galinski E.A. Compatible solute of halophilic eubacteria: molecular principles, water-solute interaction, stress protection. Experientia. 1993;49:487–490. [Google Scholar]
- 9.Gildberge A. Accelerated fish sauce fermentation by initial alkalification at low salt concentration. In: Miyachi S., Karube I., Ishida Y., et al., editors. Tokyo: Current Topic in Marine Biotechnology. Fuji Technol Press; 1989. pp. 101–104. [Google Scholar]
- 10.Gimenez M.I., Studdert C.A., Sanchez J.J., Decastro R.E. Extracellular protease of Natrialba magadii; Purification and biochemical characterization. Extremophiles. 2000;4(3):181–188. doi: 10.1007/s007920070033. [DOI] [PubMed] [Google Scholar]
- 11.Gomes J., Steiner W. The biocatalytic potential of extremophiles and extremozymes. Food Technol. Biotechnol. 2004;42:223–235. [Google Scholar]
- 12.Gupta A., Roy I., Patel R.K., Singh S.P., Khare S.K., Gupta M.N. One step purification and characterization of an alkaliphilic protease from haloalkaliphilic Bacillus sp. J. Chromatogr A. 2005;1075(1-2):103–8. doi: 10.1016/j.chroma.2005.03.127. [DOI] [PubMed] [Google Scholar]
- 13.Hough W.D., Danson J.M. Extremozymes. Curr. Opinion. Chem. Biol. 1999;3:39–46. doi: 10.1016/s1367-5931(99)80008-8. [DOI] [PubMed] [Google Scholar]
- 14.Izotova L.S., Strongin A.Y., Chekulaeva L.W., Sterkin V.E., Ostoslavskaya V.I., Lyublinskaya E.A., Timokhina E.A., Stepanov V.M. Purification and properties of serine protease from Halobacterium halobium. J. Bacteriol. 1993;155:826–830. doi: 10.1128/jb.155.2.826-830.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kamekura M., Seno Y.A. Halophilic extracellular protease from a halophilic archaebacterium strain172 p1. Biochem. Cell Biol. 1990;68:352–359. doi: 10.1139/o90-048. [DOI] [PubMed] [Google Scholar]
- 16.Kim J., Dordick J.S. Unusual salt and solvent dependence of a protease from an extreme halophile. Biotechnol. Bioeng. 1997;55:471–479. doi: 10.1002/(SICI)1097-0290(19970805)55:3<471::AID-BIT2>3.0.CO;2-9. [DOI] [PubMed] [Google Scholar]
- 17.Kobayashi T., Kanai H., Hayashi T., Akiba T., Akaboshi R., Horikoshi K. Haloalkaliphilic maltotriose-forming alpha-amylase from the archaebacterium Natronococcus sp. strain Ah-36. J. Bacteriol. 1992;174(11):3439–3444. doi: 10.1128/jb.174.11.3439-3444.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kumar S., Tamura K., Jakobsen I.B., Nei M. MEGA2: Molecular Evolutionary Genetic Analysis Software. Bioinformatics. 2001;17:1244–1245. doi: 10.1093/bioinformatics/17.12.1244. [DOI] [PubMed] [Google Scholar]
- 19.Laemmli U.K. Cleavage of structural proteins during assembly of the head of bacteriophage T 4. Nature. 1970;277:680–685. doi: 10.1038/227680a0. [DOI] [PubMed] [Google Scholar]
- 20.Larsen H. Biochemical aspects of extreme halophilism. Adv. Microbial. Physiol. 1967;1:97–132. [Google Scholar]
- 21.Lowry O.H., Rosebrough N.J., Farr A.L., Randall J.R. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951;193:265–275. [PubMed] [Google Scholar]
- 22.Margesin R., Schiner F. Potential of halotolerant and halophilic microorganisms for biotechnology. Extremophiles. 2001;5:73–83. doi: 10.1007/s007920100184. [DOI] [PubMed] [Google Scholar]
- 23.Namwong S., Hiroga T., Takada K., Tsunemi M., Tanasupawat S., Oda K. Halophilic serine protease from Halobacillus sp. SR5-3 isolated from fish sauce: Purification and characterization. Biosci. Biotechnol. Biochem. 2006;70(6):1395–1401. doi: 10.1271/bbb.50658. [DOI] [PubMed] [Google Scholar]
- 24.Nascimento W.C.A.D., Martins M.L.L. Studies on the stability of proteases from Bacillus sp and its compatibility with commercial detergents. Braz. J. Microbiol. 2006;37:307–311. [Google Scholar]
- 25.Pidiyar V.J., Jangid K., Patole M.S., Shouche Y.S. Studies on cultured and uncultered microbiota of wild Culex quinquefasciatus mosquito midgut based on 16s ribosomal RNA gene analysis. Am. J. Trop. Med. Hyg. 2004;70:597–603. [PubMed] [Google Scholar]
- 26.Reddy C.S., Jayalakshmi S.K., Sreeramulu K. Protease from Halobacterium salinarum. Indian J. Microbiol. 2003;43(4):253–256. [Google Scholar]
- 27.Ruiz D.M., De Castro R.E. Effect of organic solvents on the activity and stability of an extracellular protease secreted by the haloalkaliphilic archaeon Natrialba magadii. J. Ind. Microbiol. Biotechnol. 2007;34:111–115. doi: 10.1007/s10295-006-0174-4. [DOI] [PubMed] [Google Scholar]
- 28.Ryu K., Kim J., Dordick J.S. Catalytic properties and potential of an extracellular protease from an extreme halophile. Enzyme Microbiol. Technol. 1994;16:266–275. doi: 10.1016/0141-0229(94)90165-1. [DOI] [PubMed] [Google Scholar]
- 29.Sambrook J., Fritsch E.F., Maniatis T. 2nd edn. New York: Cold Spring Harbor Laboratory, Cold Spring Harbor; 1989. Molecular Cloning: A laboratory manual. [Google Scholar]
- 30.Schmitt W., Rdest U., Goebel W. Efficient high performance liquid chromatographic system for the purification of Halobacterial serine protease. J. Chromatgr. 1990;521:211–220. [Google Scholar]
- 31.Studdert C.A., Herrera M.K., Gil M.P., Sanchez J.J., De Castro R.E. Purification, biochemical characterization of the haloalkaliphilic archeon Natronococcus occultus extracellular serine protease. J. Gen. Microbiol. 2001;41(6):375–383. doi: 10.1002/1521-4028(200112)41:6<375::AID-JOBM375>3.0.CO;2-0. [DOI] [PubMed] [Google Scholar]
- 32.Ventosa A.A., Nieto J.J., Oren A. Biotechnological applications and potentialities of halophilic microorganisms. World. J. Microbiol. Biotechnol. 1995;11:85–94. doi: 10.1007/BF00339138. [DOI] [PubMed] [Google Scholar]
- 33.Vidyasagar M., Prakash S., Jayalakshmi S.K., Sreeramulu K. Optimization of culture conditions for the production of halothermophilic protease from halophilic bacterium Chromohalobacter sp. TVSP101. World. J. Microbiol. Biotechnol. 2007;23:655–662. [Google Scholar]
- 34.Werasit K., Preeyanuch B., Takuji O., Masatoshi G. Production and characterization of protease from an extremely halophilic Halobacterium sp. PB407. Kasetsart. J (Nat.Sci). 2004;38:15–20. [Google Scholar]
- 35.Yancey P.H. Organic osmolytes as compatible, metabolic and counteracting cryoprotectants in high osmolarity and other stresses. The J. Exptl. Biol. 2005;208:2819–2830. doi: 10.1242/jeb.01730. [DOI] [PubMed] [Google Scholar]
- 36.Yu X.T. Protease of haloalkaliphiles. In Superbugs. In: Horikoshi K, Grant WD, editors. New York: Springer Verlag; 1991. pp. 76–83. [Google Scholar]