

Abstract
The recognition of bacterial components on the intestinal epithelial cells occurs through the toll-like receptors and is followed by the induction of an effective innate immune response. We analyzed receptor expression and signaling pathways involved in activation of human colon adenocarcinoma cells after stimulation with porins and LPS of Shigella flexneri. We also analyzed the expression and production of some cytokines, of intercellular adhesion molecule-1, of antimicrobial peptides human β-defensins, and of the inducible form of nitric oxide synthase. Our data demonstrate that TLR2 is involved in porin recognition, whereas TLR4 with MD2, is required for LPS recognition.
Keywords: Porins, TLRs, PAMPs, Shigella flexneri
INTRODUCTION
The intestinal epithelium plays a crucial role in limiting direct contact of potential pathogenic bacteria with the mucosal epithelium and, in response to pathogenic bacteria, produces several cytokines and chemokines (10) Pathogenic bacteria deposit their toxic and proinflammatory surface components, such as LPS and porins, directly at the intestinal epithelial apical surface. Porins are trimeric hydrophobic proteins, resistant to proteolysis and form trans-membrane channels that allow the passive diffusion of solutes. These proteins may be released either during cell growth or during bacteriolysis (12) and cooperate with LPS (14, 16) to interfere with the cell function (15, 22). After infection with several strains of enteroinvasive bacteria (Salmonella, E.coli, Listeria, Shigella, etc.), the intestinal epithelial cells rapidly upregulate the expression of a program of host genes to activate a mucosal inflammatory and immune response and alter epithelial cell functions (17, 19, 37). Shigella flexneri is a Gram-negative pathogen responsible for bacillary dysentery in humans, the key step in its pathogenesis is the invasion of colonic epithelial cells. It was demonstrated that exposure to soluble Shigella products induced polymorphonuclear leukocytes transepithelial migration, thus indicating that the presence of extracellular bacterial products, in the absence of bacterial contact or invasion of the epithelium, was sufficient to initiate a proinflammatory response (7). Many of the genes that are activated in intestinal epithelial cells after bacterial invasion are target genes of the transcription factor NF-кB. NF-кB is a ubiquitous transcription factor involved in the inducible expression of a number of genes whose products, including many cytokines/chemokines and cell adhesion molecules, are involved in the inflammatory response (33). NFкB activation in intestinal epithelial cells is mediated by acknowledgment between bacterial products and a family of transmembrane receptors termed Toll-like receptors (TLRs). These receptors are called pattern-recognition receptors because they recognize repetitive patterns, i.e., pathogen-associated molecular patterns (PAMPs), present in several microorganisms including Gram-positive and Gram-negative bacteria, fungi and mycobacteria (11, 18). The interaction of TLRs with PAMPs results in the activation of multiple intracellular signaling events such as the activation of NF-кB and the production of cytokines (8, 24, 28). TLR4 is required for the recognition of LPS, whereas TLR2 is required for the recognition of bacterial lipoproteins and usually of Gram-positive and mycobacterial PAMPs (32, 36). Each TLR is a type 1 membrane protein, which, upon activation, results in the recruitment of myeloid differentiation factor 88 (MyD88) an adaptor protein that interacts with the TLRs through its own C-terminal Toll/IL-1R (TIR) domain (2, 4, 23, 31). The aim of this study was to evaluate the role of porins in directly stimulating the intestinal epithelium and, in particular, the exposure of the intestinal epithelium to Shigella flexneri porins. We analyzed the expression and production of some proinflammatory and chemoattractant cytokines such as IL-8, TNFα, and IL1-β, of intercellular adhesion molecule ICAM-1, of antimicrobial peptides human defensins HBD1 and HBD2, and of the inducible form of nitric oxide synthase (iNOS).
The possible differences in the signal transduction between porins and LPS and the functional importance of these molecules in activating NF-кB dependent proinflammatory genes in human colon adenocarcinoma cells were also investigated.
MATERIALS AND METHODS
Cell culture and stimuli
Caco-2 cells were cultured in Dulbecco’s Minimum Essential Medium (DMEM; GIBCO) supplemented with 2 mM glutamine, 100 U/ml penicillin, 0.1 mg/ml streptomycin and 10% fetal calf serum (FCS), in a 5% CO2 incubator. Caco-2 cells were grown as monolayers at 37°C on 6-well tissue culture clusters (Costar). In order to obtain polarization, the Caco-2 cells were cultured on transwell membranes. In these conditions, the cells undergo enterocytic differentiation and form a polarized monolayer, closely resembling, both morphologically and functionally, the human small intestine epithelium. Cell polarization was evaluated by measuring transepithelial electrical resistance: high values of electrical resistance, approximately 1000 W/cm2 (800-1200 W/cm2), were considered evidence of polarization, as well as of the integrity of the Caco-2 cell monolayers.
The cells were stimulated for 3, 6 and 24 h with 10μg/ml of LPS and 10 μg/ml of porins and incubated at 37°C in modified atmosphere at 5% CO2. The concentrations of 10 μg/ml were chosen as the best in dose-response preliminary experiments (data not shown).
LPS from Shigella flexneri serotype 1A was purchased from Sigma (St.Louis, MO, USA). Porins were extracted and purified from Shigella flexneri according to the method of Nurminen et al., 1978. The purified porins had a mass of about 37-39 kDa (Fig.1 lane A). The purified porin preparations contained only traces (10 pg/ml) of LPS determined by the Limulus amoebocyte lysate assay (Associates of Cape Cod, Inc.; distributed by PBI International, Milan, Italy) as described by Yin et al., (38) and sodium dodecyl sulfate (SDS)-polyacrylamide gel electrophoresis (34). To demonstrate that the results obtained with porin stimulation were due to the effect of the porins and not to LPS contamination, we used LPS at 10 ng/ml as an inner control. This concentration was higher than that found in our porin preparation and lower than that chosen as the best for all the experiments.
Figure 1.
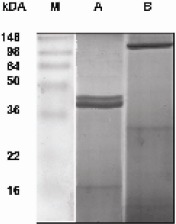
Electrophoretic pattern of the proteins present in a purified sample of Shigella flexneri porins : lane A purified preparation heat treated for 5 min, lane B purified preparation heat untreated. The SDS-polyacrylamide gel (12%) was stained with Coomassie blue. M: Prestained molecular mass markers (Invitrogen corporation)
IL-8, IL1-β, ICAM-1, iNOS, TNF-α, HBD1, HBD2, TLR2, TLR4 and MyD88 expression by Real-Time PCR
Total RNA isolated by High Pure RNA Isolation Kit (Roche Diagnostics, Milan, Italy) from Caco-2 cells before and after treatment with LPS and porins for 3, 6 and 24 h was transcribed by reverse transcriptase (Expand Reverse Transcriptase, Roche) at 42°C for 45 min according to the manufacturer’s protocol. The resulting cDNA was subjected to real-time polymerase chain reaction (PCR) analysis by rapid cycling in glass capillaries with a thermocycler (Light-Cycler; Roche Molecular Biochemicals, Milano, Italy). The reaction was performed in a final volume of 20μl with Light-Cycler-DNA Master SYBR Green I (Roche Diagnostics, Milano, Italy), which contains nucleotides, Taq DNA polymerase, buffer, SYBR Green I dye and 10mM MgCl2; cDNA (2 μl), primers and sterile H2O were added. The sequences of the PCR primers were showed in Table1. The amplification protocol consisted of an initial denaturation step at 95°C for 10 min. The following cycles for the primers were used: IL8: 95°C 5 sec, 55°C 6 sec, 72°C 12 sec, for 35 cycles; IL-1β: 95°C 5 sec, 58°C 14 sec, 72°C 28 sec, for 45 cycles; ICAM-1: 94°C 5 sec, 61°C 9 sec, 72°C 18 sec, for 35 cycles; iNOS: 94°C 5 sec, 60°C 5 sec, 72°C 10 sec, for 40 cycles; TNF-α: 94°C 5 sec, 60°C 13 sec, 72°C 27 sec, for 35 cycles; HBD1: 94°C 5 sec, 60°C 4 sec, 72°C 8 sec, for 35 cycles; HBD2: 94°C 5 sec, 63°C 6 sec, 72°C 10 sec, for 33 cycles; TLR2: 94°C 5 sec, 55°C 5 sec, 72°C 10 sec, for 30 cycles; TLR4: 94°C 5 sec, 55°C 13 sec, 72°C 27 sec, for 35 cycles; MyD88: 94°C 5 sec, 60°C 4 sec, 72°C 8 sec, for 35 cycles.
Table 1.
Primers used in the study
| Primars | Basa pairs | |
|---|---|---|
| IL-8 | 5’ oligo -5’-ATG ACT TCC AAG CTG GCC GTG-3’ | 297 hp |
| 3’ oligo -5’-TGA ATT CTC AGC CCT CTT CAA AAA CTT CTC-3’ | ||
| TWF-α | 5’oligo -5’-GAG CAC TGA AAG CAT GAT CCG-3’ | 632 bp |
| 3’ollgo -5’-AAA GTA GAC CTG CCC AG A CTC GG-3’ | ||
| ICAM 1 | 5’oligo -5’-CGT GCC GCA CTG ACC TGG AC -3’ | 446 bp |
| 3’oligo -5’-CCT CAC ACT ACT TCA CTG TCA CCT-3’ | ||
| iNOS | 5’oligo -5’-CGG TGT ATT TCC TTA CGA GGC GAC GAA GG-3’ | 259 bp |
| 3’ollgo -5’-GGT GCT GCT TGT TAG GAG GTC AAGTAA AGGGC3’ | ||
| IL-1β | 5’oligo -5’-GCA TCC AGC TAC GAA TCT CC-3’ | 708 bp |
| 3’oligD -5’-CCA CAT TCA GCA CAG GAC TC-3r | ||
| HED1 | 5’ollgo -5’-TTG TCT GAG ATG GCC TCA GGT GGT AAC-3’ | 2D0 bp |
| 3’ollgo -5’-TTT CAC TTC TGC GTC ATT TCT TCT GG-3’ | ||
| IHBD2 | 5’ollgo -5’-CCA GCC ATC AGC CAT GAG GGT-3’ | 254 bp |
| 3’ollgo -5’-GG A GCC CTT TCT GAA TCC GCA-3’ | ||
| TLR2 | 5’oligo -5’-CAA GGG CAG CTC AGG ATC-3’ | 234 bp |
| 3’ollgo -5’-GAA AGG GGC TTG AAC CAG GA-3’ | ||
| TLR4 | 5’ oligo -5’ -CAA GAA CCT GGA CCT GAG CT 3’ | 679 bp |
| 3’oligo -5’-ATT GCA CAG G CC CTC TAG AG -3’ | ||
| MD-2 | 5’oligo -5’-GCA CAT TTT CTA CAT TCC-3’ | 157 bp |
| 3’oligo -5’-CCT TAC ATC TTT TAC ACG-3’ | ||
| MyD88 | 5’oligo -5’-TAA GAA GGA CCA GCA GAG CC-3’ | 200 bp |
| 3’ollgo -5’-CAT GTA GTC CAG CAA CAG CC-3’ |
Each real-time PCR was done three times for each sample. SYBR Green I fluorescence was monitored at the end of each cycle to assess the amount of PCR product formed. After completion of the amplification protocol, a melting curve analysis was performed to confirm the specificity of amplification by cooling the sample to 65°C at a rate of 20°C/s, maintaining a temperature of 65°C for 10 sec and then heating at a rate of 0.2°C/s to 95°C, with continuous measurement of fluorescence. Cycle to cycle fluorescence emission readings were monitored and analyzed by Light-Cycler Data Software (Roche Molecular Biochemicals, Milano, Italy). The specificity of the amplification products was verified by electrophoresis on a 1.4% agarose gel stained with ethidium bromide (1μg/ml).
Sequence-specific standard curves were generated using serial dilution of specific DNA standards. All quantifications were normalized to the housekeeping gene β-actin. The data are presented as the ratio between the target gene expression and the gene expression in unstimulated conditions (29)
Immunoprecipitation
Cells treated or not with LPS and porins after 1, 3 and 24 h of incubation were washed and lysed with ice-cold buffer (50 mM HEPES, pH 7.5, 150 mM NaCl, 1% glycerol, 1% Triton, 1.5 mM MgCl2, 5 mM EGTA) supplemented with 20 mM sodium pyrophosphate, 40 μg/ml aprotinin, 4mM PMSF, 10 mM sodium orthovanadate and 25 mM NaF. Total extracts were cleared by centrifugation for 30 min at 4°C at 14000 rpm and assayed for protein content by Bradford's method (9). Two hundred μg of total proteins were incubated with 4 μg/ml of anti-TLR4 (HTA 125 Santa Cruz) and rotated overnight at 4°C. After incubation, 20μl of protein A/G sepharose were added and the samples were incubated for 2.5h at 4°C and then washed three times with 500 μl of lysis buffer. The bound proteins were analysed by Western blot analysis. TLR4/MD-2 complex was detected with anti-MD2 (IMG 415 Santa Cruz) and stained using the ECL system (Amersham Pharmacia Biotech, Piscataway, NJ).
Electrophoretic Mobility Shift Assay (EMSA)
Caco-2 cells, unstimulated and stimulated with porins and LPS (10μg/ml) for 90 min, were centrifuged and washed once in cold PBS. Cells were pelleted and nuclear extracts were prepared from cells using the Andrews and Faller method (5) and incubated with 70,000 bpm of a 22 bp oligonucleotide containing NF-кB consensus sequence (5’-AGTTGAGGGGACTTTCCCAGG-3’) (Roche Diagnostics). NF-кB was detected using [γ-32 P] dATP labeled IgK oligo probe. The DNA binding reaction was performed with 20 ml of binding buffer (2 mg of poly (dI-dC)) (Roche Diagnostics) and incubation was performed for 20’ at 25°C in the presence of 10 mM Tris-HCl, 1 mM MgCl2, 50 mM NaCl, 0.5 mM EDTA, 4% glycerol and 0.5 mM dithiothreitol. For the competition experiments, a 100- to 200-fold molar excess of AP-1 and NF-кB cold consensus oligonucleotide was added to the binding reaction mixture prior to adding the probe and incubated for 10’ at 25°C.After incubation, the samples were loaded onto 4% polyacrylamide gel using 0.5X Tris-Borate-EDTA and electrophoresed for 2h at 200V. The gels were dried and exposed to X-ray film (Kodak) at −80 °C using an intensifying screen.
RESULTS
Cytokine modulation: IL-1β, IL-8, TNF-α
IL1-β, TNF-α and IL-8 mRNA expressions were analysed in Caco-2 cells treated with porins and LPS (10μg/ml), at 3, 6 and 24 h. As shown in Fig.2 the Caco-2 cells constitutively expressed mRNA for these cytokines.
Figure 2.
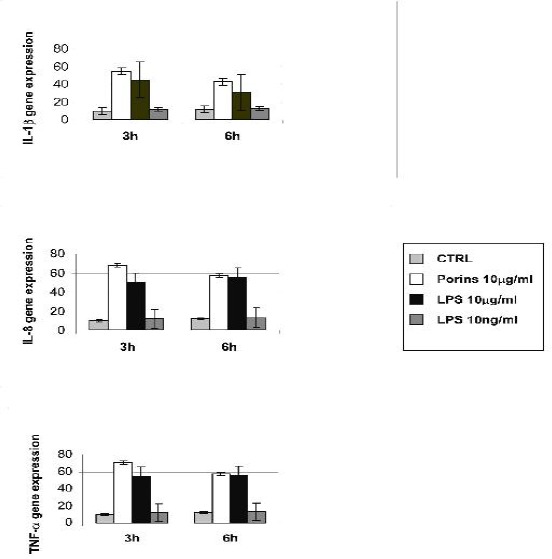
Real time for IL-8, IL1-β and TNF-α. Caco-2 cells treated or not with porins (10μg/ml), LPS (10μg/ml) and LPS (10ng/ml) at 3 and 6h. All quantifications were normalized to the housekeeping gene β-actin. The data shown are representative of three different experiments ± S.D. (P < 0.05).
The maximal expression of IL-8, IL1-β and TNF-α mRNA occurred after 3 h of porin treatment, and at this time, the mRNA upregulation was higher than that induced by LPS. At 6h the IL-1β mRNA expression retained the same pattern as that observed at 3 h. At the same time there was no detectable difference in the IL-8 and TNF-α mRNA expressions between porin and LPS stimulation. Treatment with 10 ng/ml LPS did not modify the basal level of the cytokines tested. At 24h there was no difference in the mRNA expression between the control and the treated cells (data not shown).
Expression of ICAM-1, iNOS, HBD1, HBD2
ICAM-1, iNOS, HBD1, HBD2 mRNA expressions were determined in Caco-2 cells treated with porins and LPS (10μg/ml), at 3, 6 and 24 h. As shown in Fig.3, Caco-2 constitutively expressed ICAM-1 and HBD1 mRNA, whereas there was no constitutive expression of iNOS and HBD-2 mRNA. ICAM-1 mRNA expression showed a marked increase compared to untreated cells, both at 3 and 6 h and the upregulation induced by porins was higher than the LPS upregulation at these times. At 3h and 6h iNOS and HBD2 mRNA showed the same pattern of expression, i.e., both were induced after stimulation of porins and LPS and there were no appreciable differences in the expression between the two stimuli. HBD1 mRNA expression showed only a slight upregulation after porin and LPS treatment at 3h. At 6h the expression overlapped the untreated control. Treatment with 10 ng/ml LPS did not modify the basal level of the cytokines tested. At 24h there was no difference in the mRNA expression between the control and the treated cells (data not shown).
Figure 3.
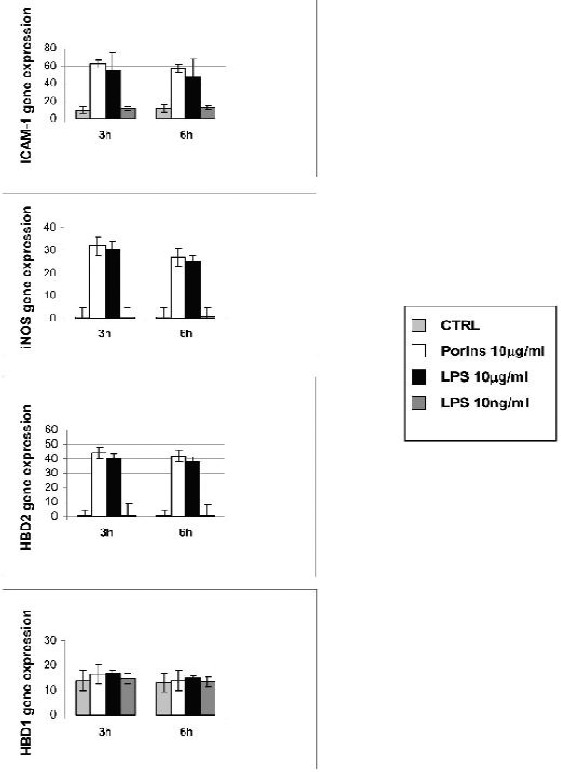
Real time for ICAM-1, iNOS, HBD1 and HBD2. Caco-2 cells treated or not with porins (10μg/ml), LPS (10μg/ml) and LPS (10ng/ml) at 3 and 6h. All quantifications were normalized to the housekeeping gene β-actin. The data shown are representative of three different experiments ± S.D. (P < 0.05).
Expression of TLR4, TLR2 and MyD88
Real-Time PCR assay showed that porins induced TLR2 expression on Caco-2 cells (Fig. 4). At 1 and 3h the increase in the expression of TLR2 was greater than for the untreated control. The effect of porins on TLR4 expression was found to be similar to the untreated control. There was also an increase in the mRNA expression of MyD88 after porin stimulation. After 1 and 3h of stimulation with LPS, the increase in the expression of TLR4 was greater compared to the untreated control, and the LPS TLR2 expression was similar to the untreated control. There was also an increase in the mRNA expression for MyD88 after LPS stimulation. At 6h there was no difference in mRNA expression between the control and the treated cells (data not shown).
Figure 4.
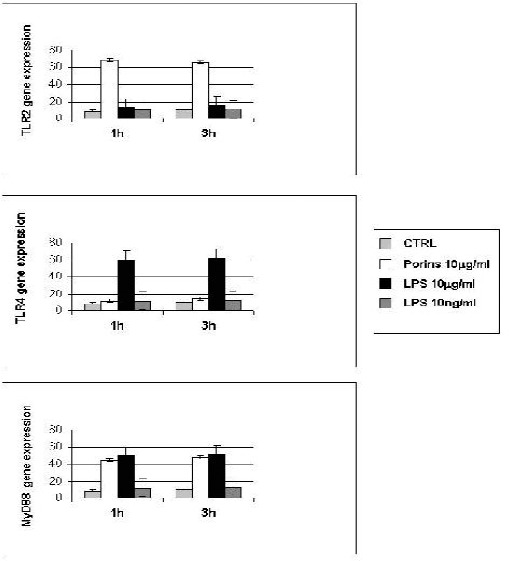
Real time for TLR-2, TLR-4 and MyD88. Caco-2 cells treated or not with porins (10μg/ml), LPS (10μg/ml) and LPS (10ng/ml) at 1 and 3h. All quantifications were normalized to the housekeeping gene β-actin. The data shown are representative of three different experiments ± S.D. (P < 0.05).
Immunoprecipitation for TLR4/MD2 complex: Caco-2 cells expressing TLR4 transcript were also positive for MD2 expression.
Analysis of immunoprecipitation experiments at 3 and 6h showed that only LPS stimulation induced a strong expression of the complex TLR4-MD2. Coprecipitation of MD2 with the HTA125 mAb therefore demonstrates physical anchoring of MD2 with TLR4 (Fig.5). There was little evidence of the TLR4-MD2 complex expression under porin stimulation. At 24h there was no difference between the control and the treated cells (data not shown).
Figure 5.
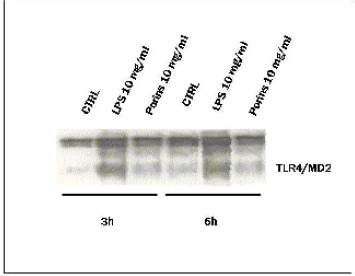
Immunoprecipitation for TLR4/MD2 complex. Cells treated or not with porins and LPS (10μg/ml) after 3, 6h of incubation were immunoprecipitated with mAb to TLR4 (HTA 125) and detected with anti-MD2. The results were the means of three independent experiments performed in duplicate.
NF-кB expression by Caco-2 cells in response to porins and LPS
To determine the involvement of the transcriptional factor NF-кB, we performed an EMSA assay with NF-кB consensus oligonucleotide in Caco-2 cells treated or not with porins and LPS for 90 min. Fig.6 shows the activation of the transcriptional factor NF-кB in response to both LPS and porins.
Figure 6.
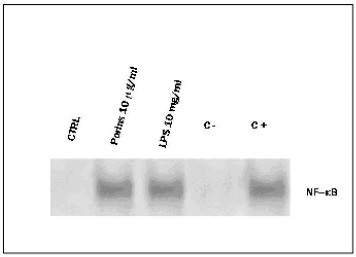
EMSA with NF-кB consensus oligonucleotide in Caco-2 cells treated or not with porins and LPS (10μg/ml) for 90 min. C-binding reaction with unlabeled excess of NF-кB consensus oligonucleotide. C+ binding reaction with unlabeled excess of AP-1 consensus oligonucleotide. The results were the means of three independent experiments performed in duplicate.
DISCUSSION
The intestinal epithelium has developed a sophisticated system in which an inflammatory response is induced to bacterial pathogens but not to non-pathogenic commensal organisms by utilizing signals from the Toll-like receptors (28). One of the most potent inflammatory mediators is LPS, a major structural component of Gram-negative bacteria, which induces the activation of NF-кB and the production of cytokines and other inflammatory molecules. Previous studies have shown NF-кB activation in epithelial cells infected by S. flexneri (13, 26) as well as by other Gram negative pathogens.
Porins play a role in bacterial virulence because of their ability to insert into the membrane of nucleated cells. Tufano et al., have shown in a previous papers that S. enteritidis serovar typhimurium porins stimulate platelet activating factor (PAF) biosynthesis by cultured human endothelial cells and that, in sepsis sustained by Gram-negative bacteria, the synthesis of PAF may be triggered either by bacterial endotoxins, or porins, or by endotoxin/porin induced cytokines. (35)
In the present study, we investigated the roles of porins, compared to LPS, in Shigella flexneri-induced inflammatory responses in human colon adenocarcinoma cells. Although the upstream stimuli of activation are distinct, both involve the activation of NF-кB and trigger an inflammatory cascade. We found that the cytokine expressions were different in response to porins or LPS; after 3h of porin stimulation all the cytokines tested (IL8, IL-1β, TNF-α) and ICAM-1 showed a stronger expression than LPS. We also observed, after 3 and 6h of treatment, the same induction pattern for HBD2 and iNOS, with no appreciable difference in the expression between the two stimuli. HBD2 is an inducible antimicrobial peptide synthesized by the epithelium that contributes together with other factors such as iNOS to counteract bacterial adherence and invasion.
Our data confirm the immunomodulatory role of porins and LPS and indicate that, in our experimental model, the activation of NF-kB occurs via two different pathways: the increase in the expression of TLR2 and MyD88 under porin stimulation, and the formation of TLR4/MD2 complex and MyD88 activation under LPS stimulation. These results suggests that the presence of effector molecule MyD88 is involved in the activity of the porins, and that TLR2, but not TLR4, has a central role in the porin-induced immune response. TLR4, instead, is required for the recognition of LPS. Previous studies have reported that mutations or deletions in TLR4 genes make animals LPS-unresponsive (6, 27, 30). Some studies also found that an additional molecule is required for TLR4-mediated LPS signaling; this was subsequently identified as the secreted molecule MD2 (1, 3). To confirm the association between TLR4 and MD2 in Caco-2 cells, immunoprecipitation experiments were conducted and our results are consistent with the membrane anchoring of MD-2 via physical association with TLR4. After LPS stimulation, the mRNA expression for MyD88 also showed a marked increase. This suggests that MyD88 is involved in the pathway induced by LPS, and that TLR4, but not TLR2, has a central role in the LPS-induced immune response.
Recent work indicates the existence of signaling pathways from TLRs that do not depend on MyD88 (20, 21) thus suggesting that multiple pathways originate from TLRs, with some convergent on NF-кB and others generating different endpoint effectors.
The ability of porins to stimulate the production and release of proinflammatory cytokines and adhesion molecules, and the production of vasodilators, such as NO, suggests a possible role of these bacterial proteins in endothelial injury, which is highly involved in the physiopathological alterations occurring during Shigella infection. Porins and LPS of Shigella flexneri are clearly involved in the signaling pathway for the activation of NF-кB via MyD88. These two outer membrane components trigger the activation of the cytokine pathway and other factors involved in the inflammatory response and co-operate in Shigella invasion. Although the transduction pathways are different in part, porins and LPS might both be necessary for producing and releasing colon inflammatory mediators during Shigella invasion in intestinal epithelial cells.
A better understanding of the innate immunity in the gastrointestinal mucosa may improve current knowledge of the pathogenesis of inflammatory bowel disease.
ACKNOWLEDGEMENT
This work was supported by grants from MIUR for the Scientific Research Program of National Interest (PRIN) 2005.
RESUMO
Via de transdução de sinal pró-inflamatória induzida por porinas de Shigella flexneri em células caco-2
O reconhecimento de componentes bacterianos nas células epiteliais intestinais ocorre através de receptores toll-like e é seguido de indução de uma resposta imune inata efetiva. Neste estudo foram analisadas as vias de expressão do receptor e sinalização envolvidas na ativação de células humanas de adenocarcinoma do colon após a estimulação com porinas e LPS de Shigella flexneri. Foram também analisadas a expressão e produção de algumas citoquinas, da molécula -1 de adesão intercelular, de □-defensinas humanas a peptídios antimicrobianos e da forma indutível de oxido nítrico sintase. Os resultados demonstraram que TLR-2 está envolvido no reconhecimento de porinas, enquanto TLR4 com MD2 é necessário para o reconhecimento de LPS.
Palavras-chave: porinas, TLR, PAMP, Shigella flexneri
REFERENCES
- 1.Abreu M.T., Arnold E.T., Thomas L.S., Gonsky R., Zhou Y., Hu B., Arditi M. TLR4 and MD-2 expression is regulated by immune mediated signals in human intestinal epithelial cells. J. Biol. Chem. 2002;277:20431–20437. doi: 10.1074/jbc.M110333200. [DOI] [PubMed] [Google Scholar]
- 2.Akira S., Hoshino K., Kaisho T. The role of Toll like receptors and MyD88 in innate immune responses. J. Endotoxin. Res. 2000;6:383–387. [PubMed] [Google Scholar]
- 3.Akira S., Takeda K., Kaisho T. Toll like receptors: critical proteins linking innate and acquired immunity. Nat. Immunol. 2001;2:675–680. doi: 10.1038/90609. [DOI] [PubMed] [Google Scholar]
- 4.Akira S., Uematsu S., Takeuchi O. Pathogen recognition and innate immunity. Cell. 2006;124:783–801. doi: 10.1016/j.cell.2006.02.015. [DOI] [PubMed] [Google Scholar]
- 5.Andrew N.C., Faller D.V. A rapid micro preparation technique for extraction of DNA-binding proteins from limiting number of mammalian cells. Nucleic Acid Res. 1991;19:2499–2506. doi: 10.1093/nar/19.9.2499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Arbour N.C., Lorenz E., Schutte B.C., Zabner J., Kline J.N., Jones M., Frees K., Watt J.L., Schwartz D.A. TLR4 mutations are associated with endotoxin hyporesponsiveness in humans. Nat. Genet. 2000;25:187–191. doi: 10.1038/76048. [DOI] [PubMed] [Google Scholar]
- 7.Beatty W.L., Meresse S., Gounon P., Davoust J., Mounier J., Sansonetti P.J., Gorvel J.P. Trafficking of Shigella lipolysaccharide in polarized intestinal epithelial cells. J. Cell. Biol. 1999;145:689–698. doi: 10.1083/jcb.145.4.689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Biswas A., Banerjee P., Mukherjee G., Biswas T. Porin of Shigella dysenteriae activates mouse peritoneal macrophage through Toll like receptors 2 and 6 to induce polarized type I response. Mol. Immunol. 2007;44:812–820. doi: 10.1016/j.molimm.2006.04.007. [DOI] [PubMed] [Google Scholar]
- 9.Bradford M. A rapid and sensitive method for the quantification of microgram-quantities of protein utilising the principle of dye binding. Anal. Biochem. 1976;72:248–256. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- 10.Cario E., Rosenberg I.M., Brandwein S.L., Beck P.L., Reinecker H.C., Podolsky D.K. Lipopolysaccharide activates distinct signalling pathways in intestinal epithelial cell lines expressing Toll like receptors. J. Immunol. 2000;164:966–972. doi: 10.4049/jimmunol.164.2.966. [DOI] [PubMed] [Google Scholar]
- 11.Cario E., Brown D., McKee M., Lynch-Devaney K., Gerken G., Podolsky D.K. Commensal-associated molecular patterns induce selective toll like receptor-trafficking from apical membrane to cytoplasmic compartments in polarized intestinal epithelium. Am. J. Pathol. 2002;160:165–173. doi: 10.1016/S0002-9440(10)64360-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Cusumano V., Tufano M.A., Mancuso G., Carbone M., Rossano F., Fera M.T., Ciliberti F.A., Ruocco E., Merendino R.A., Teti G. Porins of Pseudomonas aeruginosa induce release of tumor necrosis factor alpha and interleukin-6 by human leukocytes. Infect. Immun. 1997;65:1683–1692. doi: 10.1128/iai.65.5.1683-1687.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Elewaut D., Di Donato J.A., Kim J.M., Truong F., Eckmann L., Kagnoff M.F. NF-kappa B is a central regulator of the intestinal epithelial cell innate immune response induced by infection with enteroinvasive bacteria. J. Immunol. 1999;163:1457–1466. [PubMed] [Google Scholar]
- 14.Galdiero F., de L’ero Cipollaro G., Benedetto N., Galdiero M., Tufano M.A. Release of cytokines induced by Salmonella typhimurium porins. Infect. Immun. 1993;61:155–161. doi: 10.1128/iai.61.1.155-161.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Galdiero F., Gorga F., Bentivoglio C., Mancuso R., Galdiero E., Tufano M.A. The action of LPS porins and peptidoglycan fragments on human spermatozoa. Infection. 1988;16:349–353. doi: 10.1007/BF01644545. [DOI] [PubMed] [Google Scholar]
- 16.Galdiero M., Vitiello M., Sanzari E., D’Isanto M., Tortora A., Longanella A., Galdiero S. Porins from Salmonella entericaserovar Typhimurium activate the transcription factors activating protein1 and NF-kappa B through the Raf-1-mitogen activated protein kinase cascade. Infect. Immun. 2002;70:558–568. doi: 10.1128/IAI.70.2.558-568.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ireton K., Cossart P. Interaction of invasive bacteria with host signaling pathways. Curr. Opin. Cell Biol. 1998;10:276–283. doi: 10.1016/s0955-0674(98)80151-8. [DOI] [PubMed] [Google Scholar]
- 18.Janssens S., Beyaert R. Role of Toll like receptors in pathogen recognition. Clin. Microbiol. Rev. 2003;16:637–646. doi: 10.1128/CMR.16.4.637-646.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kagnoff M.F., Eckmann L. Epithelial cells as sensors for microbial infection. J. Clin. Invest. 1997;100:6–16. doi: 10.1172/JCI119522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kaisho T., Akira S. Dendritic-cell function in Toll like receptor and MyD88-knockout mice. Trends Immunol. 2001;22:78–83. doi: 10.1016/s1471-4906(00)01811-1. [DOI] [PubMed] [Google Scholar]
- 21.Kawai T., Akira S. Signaling to NF-kappa B by Toll like receptors. Trends Mol. Med. 2007;13:460–469. doi: 10.1016/j.molmed.2007.09.002. [DOI] [PubMed] [Google Scholar]
- 22.Latz E., Visintin A., Lien E., Fitzgerald K.A., Monks B.G., Kurt Jones E.A., Golenbock D.T., Espevik T. Lipopolysaccharide rapidly traffics to and from Golgi apparatus with the Toll like receptor 4 MD-2-CD14 complex in a process that is distinct from the initiation of signal transduction. J. Biol. Chem. 2002;277:47834–47843. doi: 10.1074/jbc.M207873200. [DOI] [PubMed] [Google Scholar]
- 23.Massari P., Henneke P., Ho Y., Latz E., Golenbock D.T., Wetzler L.M. Cutting edge: Immune stimulation by neisseria porins is toll like receptor 2 and MyD88 dependent. J. Immunol. 2002;168:1533–1537. doi: 10.4049/jimmunol.168.4.1533. [DOI] [PubMed] [Google Scholar]
- 24.Massari P., Visintin A., Gunawardana J., Halmen K.A., King C.A., Golenbock D.T., Wetzler L.M. Meningococcal porin PorB binds to TLR2 and requires TLR1 for signaling. J. Immunol. 2006;176:2373–2380. doi: 10.4049/jimmunol.176.4.2373. [DOI] [PubMed] [Google Scholar]
- 25.Nurminen M. A mild procedure to isolate the 34K, 35K and 36K porins of the outer membrane of Salmonella typhimurium. FEMS Microbiol. Lett. 1978;3:331–334. [Google Scholar]
- 26.Philpott D.J., Yamaoka S., Israël A., Sansonetti P.J. Invasive Shigella flexneri activates NF-kappa B through a lipopolysaccharide dependent innate intracellular response and leads to IL 8 expression in epithelial cells. J. Immunol. 2000;165:903–914. doi: 10.4049/jimmunol.165.2.903. [DOI] [PubMed] [Google Scholar]
- 27.Poltorak A.X., Smirnova He I., Liu M.Y., Huffel C.V., Du X., Birdwell, D., Alejos E., Silva, M., Galanos C. Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: mutations in TLR4 gene. Science. 1998;282:2085–2088. doi: 10.1126/science.282.5396.2085. [DOI] [PubMed] [Google Scholar]
- 28.Sansonetti P.J. War and peace at mucosal surfaces. Nature Reviews. 2004;4:953–964. doi: 10.1038/nri1499. [DOI] [PubMed] [Google Scholar]
- 29.Schmittgen T.D., Zakrajsek B.A., Mills A.G., Gorn V., Singer M.J., Reed M.W. Quantitative reverse transcription-polymerase chain reaction to study mRNA decay: comparison of endpoint and real-time methods. Anal. Biochem. 2000;285:194–204. doi: 10.1006/abio.2000.4753. [DOI] [PubMed] [Google Scholar]
- 30.Shimazu R., Akashi S., Ogata H., Nagai Y., Fukudome K., Miyake K., Kimoto M. MD-2, a molecule that confers lipopolysaccharide responsiveness on Toll-like receptor 4. J. Exp. Med. 1999;189:1777–1782. doi: 10.1084/jem.189.11.1777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Singleton T.E., Massari P., Wetzler L.M. Neisserial porin induced dendritic cell activation is MyD88 and TLR2 dependent. J. Immunol. 2005;174:3545–3550. doi: 10.4049/jimmunol.174.6.3545. [DOI] [PubMed] [Google Scholar]
- 32.Tapping R.I., Akashi S., Miyake K., Godowski P.J., Tobias P.S. Toll-like receptor 4, but not toll-like receptor 2, is a signaling receptor for Escherichia and Salmonella lipopolysaccharides. J Immunol. 2000;165:5780–5787. doi: 10.4049/jimmunol.165.10.5780. [DOI] [PubMed] [Google Scholar]
- 33.Tato C.M., Hunter C.A. Host-pathogens interaction: subversion and utilization of the NF-kB pathway during infection. Infec. Immun. 2002;70:3311–3317. doi: 10.1128/IAI.70.7.3311-3317.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Tsai C.M., Frasch C.E. A sensitive silver stain for detecting lipopolysaccharides in polyacrylamide gels. Anal. Biochem. 1982;179:115–119. doi: 10.1016/0003-2697(82)90673-x. [DOI] [PubMed] [Google Scholar]
- 35.Tufano M.A., Biancone L., Rossano F., Capasso C., Baroni A., De Martino A., Iorio E.L., Silvestro L., Camussi G. Outer membrane porins from gram-negative bacteria stimulate platelet-activating-factor biosynthesis by cultured human endothelial cells. Eur. J. Biochem. 1993;214(3):685–693. doi: 10.1111/j.1432-1033.1993.tb17969.x. [DOI] [PubMed] [Google Scholar]
- 36.Vasselon T., Detmers P.A., Charron D., Haziot A. TLR2 recognizes a bacterial lipopeptide through direct binding. J. Immunol. 2004;173:7401–7405. doi: 10.4049/jimmunol.173.12.7401. [DOI] [PubMed] [Google Scholar]
- 37.Witthoft T., Eckmann L., Kim J.M., Kagnoff M.F. Enteroinvasive bacteria directly activate expression of iNOS and NO production in human colon epithelial cells. Am. J. Physiol. 1998;275:564–571. doi: 10.1152/ajpgi.1998.275.3.G564. [DOI] [PubMed] [Google Scholar]
- 38.Yin E.T., Galanos C., Kinsky S., Bradshow R.A., Wessler S., Luderitz ., Sarmiento M. F. Picogram-sensitive assay for endotoxin: gelatin of Limulus polyphemus blood cell lysate induced by purified lipopolysaccharide and lipid A from gram-negative bacteria. Biochim. Biophys. Acta. 1972;261:284–289. doi: 10.1016/0304-4165(72)90340-6. [DOI] [PubMed] [Google Scholar]