

Abstract
We describe the simultaneous production of Bacillus subtilis based proteases and alpha amylase using a computer controlled laboratory scale 7.5 L batch bioreactor. The present strain is the first to be reported that concomitantly produces these two industrially important enzymes. The growth and sporulation of Bacillus subtilis was monitored and maximum production of alkaline protease and alpha amylase was found to coincide with maximum sporulation. Two types of proteases were detected in the fermentation broth; a neutral and an alkaline protease most active in a pH range of 7.0–8.0 and 8.0–10, respectively. Maximum production of proteases was observed at an incubation temperature of 37°C while that of alpha amylase was observed at 40°C. The optimum aeration and agitation levels for protease production were 0.6 L/L/min and 200rpm, respectively, and for alpha amylase were 0.6 L/L/min and 150 rpm. The kinetic parameters Yp/x and qp were also found to be significant at the given fermentation conditions.
Keywords: Simultaneous, sporulation, kinetics, fermentation, agitation
INTRODUCTION
Proteases and alpha amylases are the two most important industrial enzymes, together representing more than 70% of the total worldwide enzyme market (23). Proteases catalyze the cleavage of peptide bonds and cause the total hydrolysis of proteins while alpha amylases degrade α 1–4 linkage of starch and other substrates in an endo fashion producing maltose and glucose (1, 22). Since both these enzymes are physiologically necessary for microorganisms they are widely distributed in a number of microorganisms where they are produced in large quantities by through the fermentation process.
Both fungi and bacteria are employed for the production of proteases and alpha amylases but the choice of organism mainly depends upon the application of the enzyme produced. For example, proteases to be used in the food industry are mainly produced by fungi, whereas the proteases used in leather or detergent industries are mainly produced by bacteria. Similarly, the alpha amylases produced by fungi are more stable than those from bacteria due to high thermostability and heat sensitivity (6). The most commonly used microorganisms for protease production include Bacillus spp., Steptomyces spp., Aspergillus spp., Rhizopus spp. and Penicillium spp. (2, 9, 17) and for alpha amylase production include Bacillus spp., Steptomyces spp., Themomyces lanuginosus, Rhizopus spp., Trichoderma spp., Penicillium spp. Fusarium spp. and Aspergillus spp. (7, 8, 19, 1).
Proteases and alpha amylases occupy a pivotal position with respect to their applications in industrial fields and hence have acquired great significance in present day biotechnology. Although the enzymes can be derived from several sources such as plants, animals and microorganisms, industrial demands are generally met by enzymes from microbial sources. . Proteases are widely used in brewing, baking, tenderization of meat, dairy industry, synthesis of aspartame, developing effective therapeutic agents, treatment of wounds, detergent and leather industries (11, 12, 13, 23). The alpha amylases are extensively used in many industries including paper, textile, pharmaceuticals (8, 19), starch liquefaction, brewing, food and sugar industries (3, 4, 20).
Both enzymes can be produced economically and profitably on a large scale through fermentation process using a specific and potent strain. Submerged as well as solid-state fermentation techniques are used for the production of proteases and alpha amylases; however, bacteria usually give higher yields through submerged fermentation. A new trend in fermentation technology has arrived which is the simultaneous production of two or more products. In this technique, more than one product is produced by a single microorganism in a single fermentation batch under same or different conditions. Several workers have used this technique and have successfully produced more than one product simultaneously in a single batch operation (27, 10, 5, 25). This is a successful fermentation technique by which time, labor and energy can be saved and fermentation batches can be economized for maximum outputs from minimum inputs.
Keeping in mind the economics of a fermentation batch, the present work was undertaken to produce these two most important commercial enzymes simultaneously in a single strain of Bacillus subtilis using submerged fermentation technique with slight alteration of fermentation conditions.
MATERIALS AND METHODS
Organism, Growth and Maintenance
A highly productive strain of Bacillus subtilis IIB-26 was taken from the culture bank of Institute of Industrial Biotechnology, GC University, Lahore, which was originally isolated from the field soil. The culture was grown and maintained on nutrient agar medium. The bacterial slopes were incubated at 37°C for 48 h and then stored at 4°C. However, the strain was maintained by weekly transfers onto fresh slopes.
Inoculum preparation
Inoculum of Bacillus subtilis was prepared in 500 mL Erlenmeyer flasks containing 100 mL of nutrient broth. The flasks were sterilized in an autoclave at 121°C (15 Ib/in2 pressure) for 15 min. After cooling, the medium was aseptically inoculated with bacteria from a 48 h old slope. The flasks after inoculation were incubated for 24 h on a rotary incubator shaker (Gallenkamp, UK) at 37°C and 200 rpm.
Fermentation Experiments
The fermentation experiments for the simultaneous production of alkaline protease and alpha amylase from Bacillus subtilis were carried out in a laboratory scale 7.5 L batch bioreactor (New Bruinswick Scientific, USA) with a working volume of 5.0 L. The bioreactor was equipped with monitors, which were used to measure and control foam, temperature, pH, stirring rate and dissolved oxygen. The vessel of the bioreactor was equipped with a four-blade turbine.
Peristaltic pumps were attached to control the foam and pH by automatic addition of an antifoam silicon agent or an acid/base respectively. Five liters of the fermentation medium containing (g/L) soybean meal, 20; starch, 10; glucose, 5.0; polypeptone, 10; KH2PO4, 1.0; (NH4)2SO4, 1.0 and Na2CO3, 5.0 (pH 8.0) was added to a fermentor vessel and sterilized in an autoclave. After cooling, the medium was inoculated with already-prepared 200 mL of the inoculum containing 3.5 × 108 CFU/mL. The fermentor was run for a batch operation for the said interval of time. After the fixed interval of incubation, the fermented broth was centrifuged at 3830 × g for 10 min and the supernatant was assayed for protease and alpha amylase activities.
During all the experiments chemicals of analytical grade and calibrated labware were used.
Analytical Methods
Assay of Protease: The method of McDonald and Chen (16) was used for the assay of protease. Casein (4 mL of 1% solution in Phosphate buffer of pH 7.0 for the determination of neutral protease and pH 8.5 for the determination of alkaline protease) was incubated with 1 mL of enzyme sample at 30°C for one hour. The reaction was arrested by the addition of 5 mL of 5% trichloroacetic acid solution. The mixture was centrifuged at 2660 × g for 10 min and 1 mL of supernatant was mixed with 5 mL of alkaline reagent (made by mixing 1 mL of 1% CuSO4, 1 mL of 2.7% Sodium potassium tartarate with 100 mL of 2% Na2CO3). To this mixture 1 mL of 1N NaOH was added to make the contents of the tube alkaline. After 10 min., 0.5 mL of Folin and Ciocalteau reagent was added to the test tubes and mixed. The optical density of the solution was measured with UV/VIS spectrophotometer (CECIL, CE 7200, Cambridge, England) at 700 nm after 30 min.
One unit of protease activity is defined as the amount of enzyme required to produce an increase of 0.1 in optical density at 700 nm under the defined conditions.
Assay of alpha amylase: Alpha amylase activity was estimated according to the method of Rick and Stegbauer (24). One milliliter of enzyme sample was incubated with 1 mL of 1% soluble starch solution (in 0.1 M phosphate buffer pH 7.0) at 40°C for 10 min. The reducing sugars were measured by adding 3,5-dinitro salicylic acid reagent to the reaction mixture. The tubes were boiled for 5 min, cooled and measured for O.D at 546nm in the spectrophotometer.
One unit activity is defined as the amount of enzyme that releases one µmole of maltose per min.
Dry cell mass: After fermentation, the fermented broth was centrifuged in preweighed glass centrifuge tubes at 3830 × g for 15 min. The supernatant was removed and the pellet was washed with water three times, then dried at 105°C overnight in a hot air oven. After drying, the tubes were again weighed to determine the dry cell mass (DCM) of the bacteria.
Kinetic analysis: Kinetic parameters for batch fermentation experiments were determined according to the methods described by Pirt (21) and Lawford and Rouseau (14). The following parameters of kinetics were studied:
Maximum specific growth rate (µ)max per h - The value of (µ)max was calculated from plot of lnx vs. time of fermentation, where ‘x’ is the amount of dry cell mass.
-
Product yield coefficient (Yp/x) U/mL/mg - The value of Yp/x was determined by the equation:
Where ‘p’ is the amount of product (U/mL) and ‘x’ is the amount of dry cell mass (mg).
- Specific product yield coefficient (qp) U/mL/h - The value of qp was determined by the equation:
RESULTS AND DISCUSSION
Relationship of growth and sporulation with enzyme production
The correlation of growth and sporulation of Bacillus subtilis with enzyme production was studied and a set of typical enzyme production curves is presented in the figure 1 along with growth and sporulation curves. Figure 1 shows that bacterial growth reached its maximum value after 24 h of cultivation while maximum sporulation occurred after 42 h of incubation. It is also clear from the results that the production rate of alpha amylase is linear up to about 36 h and maximum alpha amylase production coincides with maximum sporulation. In fact, the curves for alpha amylase production and sporulation in B. subtilis are almost identical. During the present studies, it was found that two types of proteases i,e; alkaline and neutral protease were secreted in the fermentation broth by the microorganism. It is also known from the previous studies that Bacillus subtilis produces different types of enzymes during fermentation (28). The maximum production of alkaline protease also corresponded with the point of maximum sporulation, but the maximum production of neutral protease occurred somewhat earlier at about 36 h of cultivation. Alkaline protease has also been named “sporulation protease” and is an absolute requirement for sporulation (15, 29) in B. subtilis hence given the name.
Figure 1.
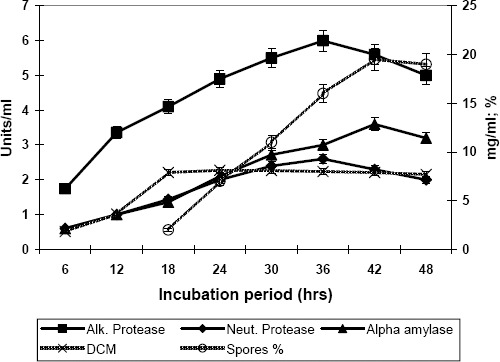
Relationship of growth and sporulation with the production of proteases and alpha amylase by Bacillus subtilis. Inc. temperature: 35°C; Agitation: 200rpm; Aeration: 0.6 L/L/min. Values are the mean of three replicates.
Under most growth conditions, Bacillus species produce extracellular protease during the post-exponential growth phase (26). Mandelstam (15) attributed this behavior to an increased need for turnover of cell proteins at the slower growth rate. Similarly the role of alpha amylase in sporulation may be energy liberation for bacteria by hydrolysis of materials present in the substrate. Overproduction of some extracellular enzymes is a reflection of the very high demands of energy and nitrogen compounds for sporulation.
Effect of incubation temperature on enzyme production
The effect of incubation temperature (30–50°C) on enzyme production, cell growth and sporulation was also studied and the results are presented in Figure 2. The cell mass was increased as the temperature was increased from 25°C, reached maximum (8.5 mg/mL) at 40°C, and then started decreasing which shows the inhibiting effect of higher temperature on the growth of the organism. Sporulation shows a progressive increase from 25 to 35°C (3–17 %) and between 35 and 50°C, it was fairly constant (17–19 %), in which temperature range, the yield of alpha amylase was also found to be maximum. However, production of proteases appeared to be maximum at a cultivation temperature of 37°C.
Figure 2.
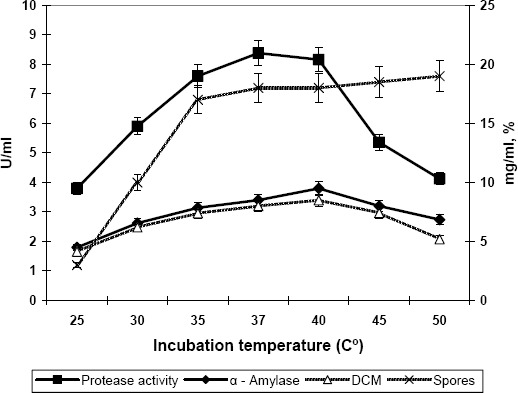
Effect of incubation temperature on the production of protease and alpha amylase by Bacillus subtilis. Inc. period: 42hrs; Agitation: 200rpm; Aeration: 0.6 L/L/min. Values are the mean of three replicates.
For alpha amylase production, it appears that a temperature of 40°C is optimal whilst for protease the optimum was around 37°C. There was very sharp maximum for proteolytic activity near 37°C, whilst the maximum for alpha amylase was observed at 40°C and was much less sharp. The kinetic parameters also revealed the same results as depicted in the experiments. The product yield coefficient (Yp/x) and specific product yield coefficient (qp) for protease production were maximum at 37°C showing the optimum temperature for protease production while in case of alpha amylase production the kinetic parameters showed maximum values at an incubation temperature of 40°C (Table 1).
Table 1.
Product yield coefficient (Yp/x) and specific rate of product formation (qp) of Bacillus subtilis at different incubation temperatures
| Incubation temperature (C°) | Yp/x for protease (U/mL/mg) | qp for protease (U/mL/h) | Yp/x for alpha amylase (U/mL/mg) | qp for alpha amylase (U/mL/h) |
|---|---|---|---|---|
| 25 | 0.917 | 0.121 | 0.434 | 0.057 |
| 30 | 0.951 | 0.126 | 0.422 | 0.056 |
| 35 | 1.02 | 0.135 | 0.437 | 0.058 |
| 37 | 1.047 | 0.139 | 0.425 | 0.056 |
| 40 | 0.96 | 0.127 | 0.45 | 0.059 |
| 45 | 0.722 | 0.096 | 0.431 | 0.057 |
| 50 | 0.686 | 0.091 | 0.430 | 0.057 |
Effect of aeration and agitation on enzyme production
In the experiments concerned with variation in aeration (0.2–1.0 L/L/min), the maximum cell mass was found at an aeration rate of 0.6 L/L/min, in conjunction with a sporulation rate of 18.5% (Figure 3). These properties of growth and sporulation corresponded to the maximum production of both the enzymes tested.
Figure 3.
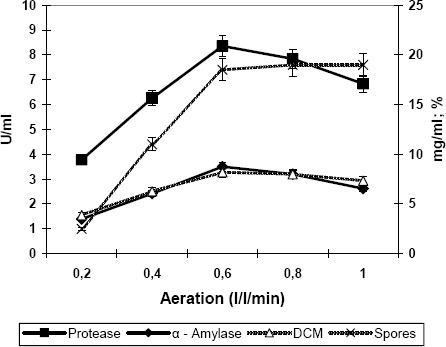
Effect of aeration on the production of proteases and alpha amylase by Bacillus subtilis. Inc. temperature: 35 C°; Agitation: 200rpm; Inc. period: 42hrs. Values are the mean of three replicates.
In the agitation experiments, variable stirring speeds ranging from 100 – 300 rpm were evaluated (Fig. 4). Stirring speeds lower than 200 rpm were not sufficient to keep the solids in the medium in homogeneous suspension; they also failed to break the clumps of cells. On the other hand, a speed higher than 250 rpm was found to cause excessive foaming thus necessitating the automatic addition of such large quantities of antifoam agent (silicon oil) that enzyme yields were impaired. In addition, the lower speed (150 rpm) was found to be slightly better for alpha amylase production, but for proteases the higher speed (200 rpm) produced significantly better results (18).
Figure 4.
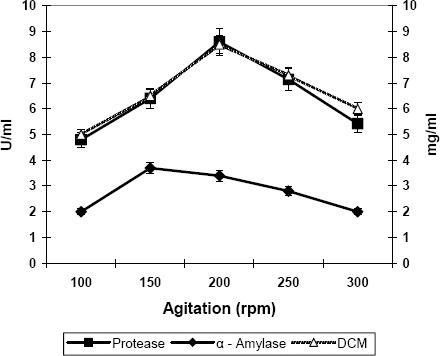
Effect of agitation on the production of proteases and alpha amylase by Bacillus subtilis. Inc. temperature: 35 C°; Inc. period: 42hrs; Aeration; 0.6 L/L/min. Values are the mean of three replicates.
The values of Yp/x and qp obtained after kinetic analysis of results revealed that the best aeration and agitation rates for protease production were 0.6 L/L/min and 250 rpm, respectively while for alpha amylase production the same aeration rate but a different agitation rate i,e; 150 rpm was found significant (Tables 2, 3).
Table 2.
Product yield coefficient (Yp/x) and specific rate of product formation (qp) of Bacillus subtilis at different aeration levels
| Aeration (L/L/min) | Yp/x for protease (U/mL/mg) | qp for protease (U/mL/h) | Yp/x for alpha amylase (U/mL/mg) | qp for alpha amylase (U/mL/h) |
|---|---|---|---|---|
| 0.2 | 0.97 | 0.129 | 0.36 | 0.047 |
| 0.4 | 0.9 | 0.131 | 0.38 | 0.050 |
| 0.6 | 1.02 | 0.135 | 0.42 | 0.055 |
| 0.8 | 0.98 | 0.130 | 0.40 | 0.053 |
| 1.0 | 0.93 | 0.123 | 0.35 | 0.046 |
Table 3.
Product yield coefficient (Yp/x) and specific rate of product formation (qp) of Bacillus subtilis at different agitation rates
| Agitation (rpm) | Yp/x for protease (U/mL/mg) | qp for protease (U/mL/h) | Yp/x for alpha amylase (U/mL/mg) | qp for alpha amylase (U/mL/h) |
|---|---|---|---|---|
| 100 | 0.96 | 0.127 | 0.4 | 0.053 |
| 150 | 0.98 | 0.130 | 0.56 | 0.074 |
| 200 | 1.01 | 0.134 | 0.4 | 0.053 |
| 250 | 0.97 | 0.129 | 0.38 | 0.050 |
| 300 | 0.90 | 0.119 | 0.33 | 0.043 |
CONCLUSION
We conclude that the present strain of Bacillus subtilis can simultaneously produce alpha amylase and protease in high titers. The production was substantially enhanced by optimization of culture conditions. The strain can be proved very useful commercially for the production of proteases and amylase in a single batch operation.
ACKNOWLEDGEMENTS
We are thankful to Dr. Adrian Briggs, Department of Genetics, Harvard Medical School, USA for reviewing the manuscript.
REFERENCES
- 1.Abdullah R., Ashraf H., Haq I. Optimization and kinetic analysis of carbon sources on the production of alpha amylase by Saccharomyces cerevisiae. J. Food Technol. 2003;1(4):187–190. [Google Scholar]
- 2.Adinarayana K., Ellaiah P. Response surface optimization of the critical medium components for the production of alkaline protease by a newly isolated Bacillus sp. J. Pharm. Pharmaceut. Sci. 2002;5(3):272–278. [PubMed] [Google Scholar]
- 3.Batlle N., Carbonell J.V., Sendra J.M. Determination of depolymerization kinetics of amylose, amylopectin and soluble starch by Aspergillus oryzae ‘α-amylase’ using a fluorimetric 2-p-toluidine naphthalene-6-sulfonate /flow injection analysis system. Biotechnol. Bioeng. 2000;70(5):552–554. doi: 10.1002/1097-0290(20001205)70:5<544::aid-bit9>3.0.co;2-5. [DOI] [PubMed] [Google Scholar]
- 4.Carlsen M., Nielsen J., Villadsen J. Growth and alpha amylase production by Aspergillus oryzae during continuous cultivations. J. Biotechnol. 1996;45(1):81–93. [Google Scholar]
- 5.Chuanbin L., Yan L., Wei L., Zhiyou W., Dongmei W., Shulin C. Washington State University, Pullman: WA; 2005. Nisin and Lactic Acid Simultaneous Production from Cheese Industry Byproducts: Optimization of Fermentation Conditions Through Statistically Based Experimental Designs, Department of Biological Systems Engineering; pp. 99164–6120. [Google Scholar]
- 6.Duochuan L., Yijun Y., Youliang P., Chongyao S., Peijin Z., Yicum H. Purification and properties of a thermostable alpha amylase from the thermophilic fungus Thermomyces lanuginosus. Acta. Microbiol. SIN. 1997;37(2):107–114. [Google Scholar]
- 7.Fogarty W.M., Dooyle E.M., Kelly C.T. Comparison of the action pattern 2 high maltose forming alpha amylase on linear maltooligosaccharides. Enzyme Microbiol. Technol. 1999;25(3–5):330–335. [Google Scholar]
- 8.Haq I., Abdullah R., Ashraf H., Shah A.H. Isolation and screening of fungi for the biosynthesis of alpha amylase. J. Biotechnol. 2002;1(2–4):61–66. [Google Scholar]
- 9.Haq I., Mukhtar H. Biosynthesis of protease by Rhizopus oligosporous IHS13 by solid-state fermentation in a low cost medium. J. Basic Microbiol. 2004;44(4):280–287. doi: 10.1002/jobm.200410393. [DOI] [PubMed] [Google Scholar]
- 10.Hari K.S., Chowdary G.V. Optimization of simultaneous saccharification and fermentation for the production of ethanol from lignocellulosic biomass. J. Agric Food Chem. 2000;48(5):1971–6. doi: 10.1021/jf991296z. [DOI] [PubMed] [Google Scholar]
- 11.Inhs D.A., Schmidt W., Richter F.R. 1999. Jun, Proteolytic enzyme cleaner. US Pat. 5961366. [Google Scholar]
- 12.Kalisz M.H. Microbial Proteinases. Adv. Biochem. Eng. Biotechnol. 1988;36:17–55. doi: 10.1007/BFb0047944. [DOI] [PubMed] [Google Scholar]
- 13.Kumar C.G., Tagaki H. Microbial alkaline proteases: from a bio-industrial viewpoint. Biotechnol. Adv. 1999;17:561–594. doi: 10.1016/s0734-9750(99)00027-0. [DOI] [PubMed] [Google Scholar]
- 14.Lawford H.G, Roseau J.D. Mannose fermentation by ethanologenic recombinants of E. coli and kinetical aspects. Biotechnol. Lett. 1993;15:615–620. [Google Scholar]
- 15.Mandelstam J. Turnover of protein in growing and nongrowing population of Escherichia coli. Biochem J. 1958;69:110–119. doi: 10.1042/bj0690110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.McDonald C.E., Chen L.L. Lowry modification of the Folin reagent for determination of proteinase activity. Annals Biochem. 1965;10:175. doi: 10.1016/0003-2697(65)90255-1. [DOI] [PubMed] [Google Scholar]
- 17.Mohamed A., Abdel N., Ismail A.M.S., Ahmed S.A., Ahmed F., Fattah A. Production and Immobilization of alkaline protease from Bacillus Mycoides. Bioresource Technol. 1998;64:205–210. [Google Scholar]
- 18.Mukhtar H., Haq I. Production of alkaline protease by Bacillus subtilis and its application as a depilating agent in leather processing. Pak. J. Bot. 2008;40(4):1673–1679. [Google Scholar]
- 19.Nascimento W.C.A., Martins M.L.L. Production and properties of an extracellular protease from thermophilic Bacillus sp. Braz. J. Microbiol. 2004;35(1–2):91–96. [Google Scholar]
- 20.Nigam P., Singh D. Enzymes and microbial system involved in starch processing enzyme. Microb. Technol. 1995;17:770–778. [Google Scholar]
- 21.Ohdan K., Kuriki T., Takata H., Okada S. Introduction of raw starch-binding domains in to Bacillus subtilis alpha amylase by fusion with the starch-binding domain of Bacillus cyclomaltodextrin glucanotransferase. Appl. Environ. Microbiol. 2000;66(7):3058–3064. doi: 10.1128/aem.66.7.3058-3064.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Pirt S.J. Principles of cell and microbe cultivation. USA: Blackwells Scientific; 1975. [Google Scholar]
- 23.Ramachandran S., Patel A.K., Nampoothiri K.M., Chandran S., Szakacs G., Sccol C.R., Pandey A. Alpha amylase from a fungal culture grown on oil cakes and its properties. Braz. Arch. Biol. Technol. 2004;47(2):309–317. [Google Scholar]
- 24.Rao M.B., Tanksale A.M., Ghatge M.S., Deshpande V.V. Molecular and biotechnological aspects of microbial proteases. Microbiol. Mol. Biol. Rev. 1998;62:597–635. doi: 10.1128/mmbr.62.3.597-635.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Rick W., Stegbauer H.P. Alpha amylase of reducing groups. In: Bergmeyer H. V., editor. Methods of Enzymatic Analysis. Vol. 2. New York: Academic Press; 1974. pp. 885–890. [Google Scholar]
- 26.Sangeetha R, Geetha A, Arulpandi I. Concomitant production of protease and lipase by Bacillus licheniformis VSG1: production, purification and characterization. Braz. J. Microbiol. 2010:41. doi: 10.1590/S1517-838220100001000026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Schaeffer P. Sporulation and production of antibiotics, exoenzymes and exotoxins. Bacteriol. Rev. 1969;33:48–71. doi: 10.1128/br.33.1.48-71.1969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Toropova E.G., Maksimov V.N., Tkhaker V., Egorov N.S. Selection of the medium for the simultaneous synthesis of an antibiotic, protease and pigment by a Nocardia fructiferi culture. Nauchnye Doki. Vyss. Shkoly Biol. Nauki. 1984;10:96–101. [PubMed] [Google Scholar]
- 29.Yoneda Y., Maruo B. Mutation of Bacillus subtilis causing hyperproduction of alpha amylase and protease and its synergistic effect. J Bacteriol. 1975;124(1):48–54. doi: 10.1128/jb.124.1.48-54.1975. [DOI] [PMC free article] [PubMed] [Google Scholar]