

Abstract
Transient expression systems are valuable tools in molecular biology. Agrobacterial infiltration of leaves is well-established in tobacco, but has led to limited success in the model plant Arabidopsis thaliana. An efficient expression system combining the advantages of Arabidopsis (well-characterised) and the simplicity of leaf infiltration is desirable. Here, I describe Agrobacterium tumefaciens-mediated transformation of Tropaeolum majus (nasturtium, order Brassicales) as a remarkably simple, cheap and highly efficient transient expression system. It provides the Arabidopsis community with a tool to study subcellular localisation, protein–protein interactions and reporter gene activities (e.g. luciferase, β-glucuronidase) in a genetic background that is closely related to their primary model organism. Unlike Arabidopsis , Tropaeolum is capable of engaging in endomycorrhizal associations and is therefore relevant also to symbiosis research. RNAi-based approaches are more likely to succeed than in the distantly-related Nicotiana transformation system. Tropaeolum majus was voted the “medicinal plant of the year 2013”. Conquering this plant for genetic manipulations harbours potential for biotechnological and pharmacological applications.
Introduction
The soil-born Agrobacterium tumefaciens is the only organism capable of interkingdom gene transfer. It has been employed intensively for genetic manipulation of plant cells. Transformation is accomplished through the action of both bacterial and host proteins, many of which have been identified and functionally characterised [1]. Yet, it is still impossible to predict which plant species are easily accessible and which are recalcitrant to Agrobacterium -mediated transformation.
Arabidopsis thaliana is the best-characterised plant, whose properties (small and fully sequenced genome, short regeneration time, modest growth requirements) have facilitated research fundamentally [2,3]. Its close relatedness to several agriculturally relevant plants, such as rapes, cabbage and mustard, contribute to the popularity and relevance of Arabidopsis as model organism in plant science. In Arabidopsis , Agrobacterium -mediated T-DNA transfer is primarily employed to generate stable transgenic plants. Transient transformation efficiency is low, it strictly depends on growth conditions and has yielded satisfactory results only in young seedlings [4]. In contrast, stable transformation is conveniently achieved through the “floral dip” method [5], in which plants at the flowering stage are submerged into a suspension of agrobacteria carrying the desired transformation plasmid. Primary transformants are obtained within few months. However, if one wishes to transfer multiple transgenes, different antibiotic markers are needed to secure reliable selection. Alternatively, characterised transgenic individuals can be crossed, and individuals carrying both transgenes identified by genotyping. Although these approaches have their merits, substantial time and space are needed. For more rapid answers as to the subcellular localisation of a protein of interest, trans-activation capacity of transcription factors or protein–protein interactions transient expression systems are the method of choice. The most commonly used approaches are bombardment, PEG-mediated protoplast transformation and Agrobacterium -mediated transformation [6,7]. Transgenes can be introduced into protoplasts obtained from suspension cultures or from freshly isolated leaf mesophyll cells. The recently reported tape- Arabidopsis -sandwich strategy offers a further simplification of this technique [8]. Similarly easy transformation, allowing transfer of multiple transgenes at a time, has been achieved in Poinsettia [9]. However, only comparatively few species are amenable to protoplast transformation, and optimisation of isolation/transformation protocols are time-consuming. In addition, data obtained from studies in protoplasts, which are deprived of their cell wall and tissue context, need to be interpreted with caution.
An alternative approach for multiple transgene expression is Agrobacterium -mediated transformation. Again, this method has its merits and limitations. It does allow analysis in intact tissue, is easy to perform and a low-cost method. On the other hand, by definition, it strictly relies on the presence of the microbial pathogen. Agrobacteria carrying the construct(s) of interest are infiltrated into leaves of intact plants, and transgene expression can be studied within 3-14 days. Nicotiana tabacum [10] and Nicotiana benthamiana [11] are the favoured species for this approach. They are easy to propagate, and agrobacterial infiltration is normally performed in 6-8 week-old plants. For regular supply and growing of plants significant space is required. Also, in general a maximum of 4-6 leaves can be infiltrated per plant. Within a given plant, transformation efficiencies can differ substantially depending on the type of leaf (upper, lower) and leaf area (margin; distance to petiole etc.) being infiltrated [12]. Plants tend to flower early (<10 weeks) under the conditions optimised for their regular supply; and are normally not used at this stage anymore. However, most importantly, for the study of Arabidopsis -derived transgenes, Nicotiana is a sub-optimal system, since it is only distantly related to Arabidopsis .
The motivation of this study was to identify a plant species that is a) related to Arabidopsis , b) modest in its growth requirements (light/humidity), c) less space-occupying than Nicotiana , d) easy to infiltrate with Agrobacteria and e) transformable at high efficiency. Tropaeolum majus (nasturtium) fulfils all above requirements.
Here, I describe Agrobacterium -mediated transformation of T . majus as a convenient, cheap and efficient transient expression system. It facilitates studies in a genetic background that is closely related to the model plant Arabidopsis . In addition, it offers an alternative and complementary method to Nicotiana leaf infiltration. The accessibility of Tropaeolum to simple and fast genetic manipulation potentially drives progress in several fields of plant research, including those aimed at biotechnological and pharmacological applications. Unlike Arabidopsis , Tropaeolum is capable of engaging in endomycorrhizal associations, and is therefore also of interest to symbiosis researchers. Besides, it was voted the “medicinal plant of the year 2013”, and the here-described properties may contribute to its “fame” and popularity in the scientific community.
Materials and Methods
Plant growth and transformation
Tropaeolum majus L. seeds were directly placed into pots and covered with 1 cm soil. Pots were kept moist and placed at room temperature (20-24 °C, no special growth chamber, range of air humidity/temperature/light intensity tested). Plants that had formed at least 4-6 leaves were used for infiltration. Agrobacterium tumefaciens (GV3101) carrying the Ti helper plasmid pSOUP and a pGreen-derived [13] construct of interest were streaked from glycerol cultures onto a LB agar plate supplemented with rifampicin, tetracycline and kanamycin. After 2 days at 28 °C, loops of bacterial cells were transferred into 1.5 ml reaction tubes and resuspended in 1 ml infiltration liquid (IF): 10 mM MES, pH5.7; 10 mM MgCl2 and 100 µM acetosyringone. Samples were centrifuged, the supernatant fluid discarded, and the bacterial pellet was resuspended in IF, adjusted to an OD (600 nm) of 0.2. The bacterial suspensions were incubated at least 3 hours (dark, RT, no shaking) before infiltration.
Plasmids
pGreen derivatives carrying the CaMV35S promoter and transgenes for YFP, MKK4-SPYCE, MPK3-SPYNE or GUS have been described previously [14,15]. The coding region of luciferase was isolated as NcoI-XbaI fragment from pGL3 (Promega) and used to replace a YFP-encoding fragment in plasmid CaMV35S::YFP. Arabidopsis gene accession numbers are: MPK3: At3g45640, and MKK4: At1g51660.
UV microscopy
YFP expression and protein–protein interactions studies were conducted at a UV microscope (Leica DM5500B), equipped with excitation/emission filters: BP450–450 nm/LP515 nm as described previously [15].
Protein extraction and analysis
Arabidopsis thaliana (Col-O), Nicotiana benthamiana and Tropaeolum majus leaves were snap-frozen in liquid nitrogen. Proteins were extracted as previously described [15] separated by SDS-PAGE (12%) and visualised by Coomassie blue staining.
Additional technical comments
Tropaeolum majus growth is not seemingly impaired by the presence of agrobacteria, and the plant will form new leaves. An individual plant can be infiltrated multiple times. Transformation efficiency and level of transgene expression of primary and secondary infiltrations (1 week gap) are similar. When all leaves, present at the time of infiltration were used, plants tended to display chlorophyll bleaching and general disease symptoms. As a rule of thumb, a maximum of 70% leaf area can be infiltrated.
Results
For successful A. tumefaciens mediated transformation via leaf infiltration two obstacles need to be overcome; one is a “mechanic” issue, the other refers to a plant’s “willingness” or susceptibility to accept foreign DNA from a pathogen. Mechanically, infiltration means the introduction of liquid into leaf tissue. Leaf development involves cell formation that defines the leaf structure, and cell expansion. Rapidly expanding cells have a rather loose cell wall and may therefore be infiltrated more easily. The principal requirements for a candidate plant to offer improvement and/or an alternative to Nicotiana , include:
Easy to grow (space, time, light/humidity)
Related to Arabidopsis
Multiple leaves of similar width, ideally >5 qcm for easy handling
Late-flowering, offering a wider time window for infiltrations
Syringe-introduced liquid should spread readily within the leaf to give large transformation areas but few sites of wounding (point of injection).
Nicotiana and Arabidopsis belong to the order of Solanales and Brassicales, respectively, and thus are only distantly related (Figure 1). Several members of the Brassicales were critically examined with respect to the above-listed criteria. Nasturtium turned out to be by far the most suitable plant. It belongs to the family of Tropaeolaceae, one of the 17 families within the order Brassicales (Figure 1, right).
Figure 1. Angiosperm phylogeny and phylogenetic relation within the order Brassicacales.
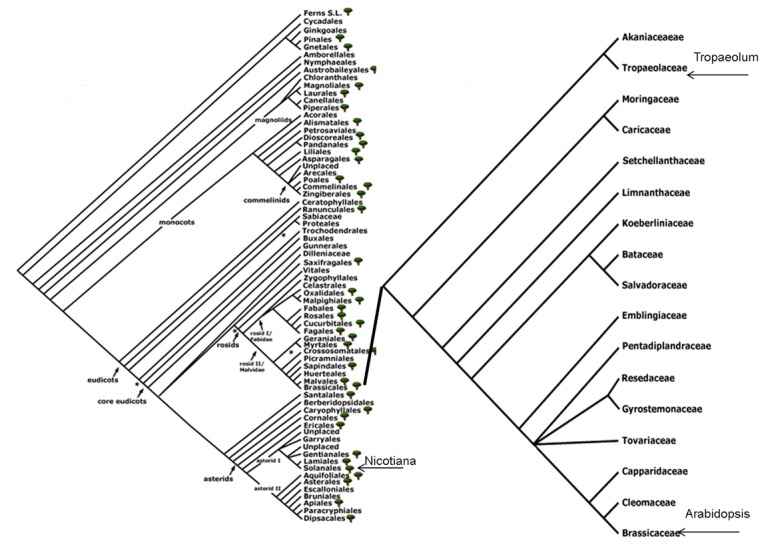
Positions of plant species mentioned in the manuscript are indicated. The image was modified from the „Angiosperm Phylogeny Website – Missouri Botanical Garden“. Reprinted from http://www.mobot.org/MOBOT/research/APweb/ under a CC BY license, with permission from Peter F. Stevens, original copyright 2013. (http://www.mobot.org/MOBOT/research/APweb/).
Tropaeolum majus seeds were sown directly on soil. Germination was observed within 3 days. Two-week-old plants had formed 4-6 leaves. Plants were tested for their accessibility to agrobacterial infiltration. To this end, a simplified and empirically optimised technique, based on the published transformation procedure of N . benthamiana [16] was used. An overview of material and steps involved is shown in Figure 2. Infiltration was performed by gently releasing the content of a syringe containing agrobacteria suspension into the abaxial side of leaves. Larger leaves were easier to infiltrate. The uptake of infiltration liquid was detectable as successive spreading of a dark-green zone (Figure 3A). In initial experiments, only single gene transfers were tested: At 5 dpi, leaves transformed with a CaMV35S::YFP construct (for constitutive expression of yellow fluorescent protein) were analysed by UV microscopy. Stunningly, transformation efficiency was >80%, as revealed by the strong fluorescence signal emitted from YFP-expressing cells (Figures 4 and S1). In leaves infiltrated with a control construct (luciferase) there was no detectable fluorescent signal, excluding the possibility that the signal seen in YFP samples derived from autofluorescence. Reciprocally, when leaves were assayed for luciferase activity, only those transformed with CaMV35S::luciferase displayed chemiluminescence upon treatment with the luciferin substrate (Figure 5).
Figure 2. Outline of the Tropaeolum transformation procedure.

Seeds are sown on soil and grown until at least four leaves have developed (app. 2 weeks). Agrobacteria carrying the construct of interest are streaked from glycerol cultures onto LB (incl. Antibiotics) plates for 2 days. Loops of cells are briefly washed and resuspended in infiltration liquid, adjusted to OD600nM 0.2, incubated for 3 hours and subsequently infiltrated into the leaves.
Figure 3. Leaf appearance after infiltration.

A) symptom-free leaf at 7 days post-infiltration. B) necrosis in leaves 21 days post-infiltration. Note that symptoms occur only around the infiltration site (syringe release) but not in the remaining infiltrated area.
Figure 4. Transient expression of Yellow Fluorescent Protein.

Tropaeolum majus leaves were infiltrated with agrobacteria carrying a construct for constitutive expression of YFP. Transgene expression was analysed 5 dpi by UV microscopy, using a low (left) and high-magnification objective. Top: fluorescence; bottom: bright-field image. No fluorescence was observed in leaves infiltrated with Agrobacteria carrying a control construct (luciferase) (right). For additional images see figure S1.
Figure 5. Luciferase activity in transgenic Tropaeolum majus tissue.
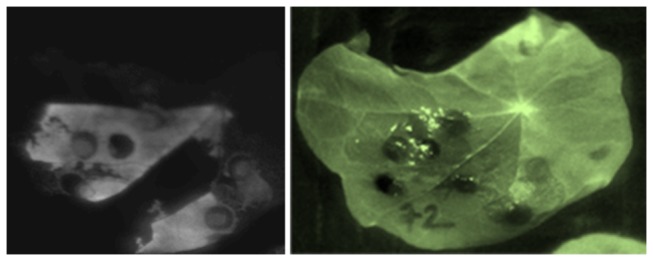
T . majus leaves were infiltrated with agrobacteria carrying a construct for constitutive expression of the luciferase gene. 5 days post-infiltration, an area of the leaf (top left) was infiltrated with substrate solution (10 mM MgCl2, 10 mM MES pH 5.7, 0.5% DMSO, 0.1 mM luciferin), followed by chemiluminescence detection. Left: chemiluminescence (Chemdoc Biorad, exposure 7 sec.). Right: brightfield image of leaves, showing the luciferin-infiltrated area. Note that some areas were not infiltrated with substrate solution only, but not with agrobacteria and vice versa to document that the signal is solely due to expression of the reporter transgene.
To monitor the duration of “transient” expression, YFP-transformed leaves were examined over a 3-week-period. Strong YFP-derived fluorescence was still seen after two weeks, but hardly detectable after three weeks. The appearance of infiltrated leaves indicated that the presence of agrobacteria did not largely affect tissue vitality. At 7 dpi, leaves were virtually symptom-free (Figure 3A). At 21 dpi, necrosis was observed in the narrow area surrounding the infection site (Figure 3B).
Having established the transformation procedure, the suitability of nasturtium leaf infiltration as tool in popular aspects of molecular biology – such as reporter gene activity and protein–protein interaction studies was evaluated. Leaves were infiltrated with agrobacteria carrying a CaMV35S promoter-driven β-glucuronidase (GUS) gene. The GUS gene contains an intron to ensure that GUS activity solely derives from the plant, not from the pathogen. GUS staining in tissue 5 dpi revealed large areas with the typical blue, intensive colour indicating strong GUS activity (Figure 6). Concludingly, nasturtium is compatible with GUS reporter gene activity assays. The intron was recognised and spliced correctly.
Figure 6. β-glucuronidase activity in transgenic Tropaeolum majus tissue.
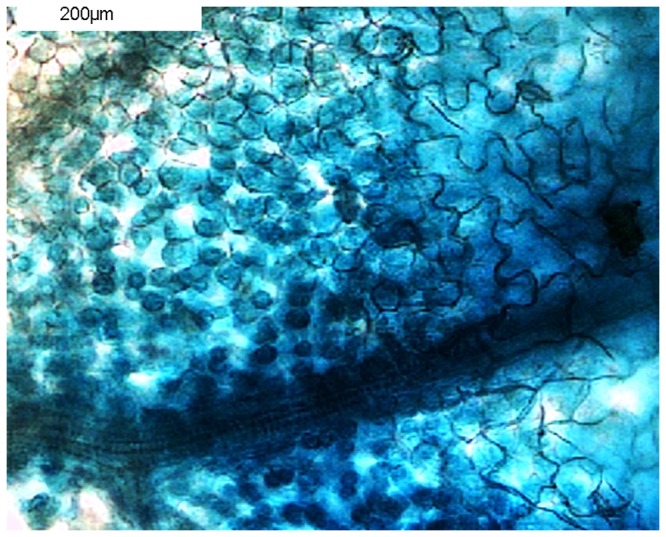
Leaves were infiltrated with Agrobacteria carrying a construct for constitutive expression of the GUS reporter gene (including an intron). Enzyme activity was assessed 5 dpi by infiltrating the leaf with substrate solution. The typical blue colour arises from β-glucuronidase-catalysed conversion of the substrate.
Next, co-delivery of multiple transgenes for protein–protein interaction studies was tested using the bimolecular fluorescence complementation (BiFC) approach [17]. I chose two well-characterised interacting proteins from Arabidopsis , representing components of a mitogen-activated protein kinase cascade [15,18]: MKK4, fused to the C-terminal YFP fragment and MPK3, fused to the N-terminal YFP fragment. UV microscopy revealed positive fluorescence in leaves 5 days after co-infiltration. Signals were observed in cytoplasm and nuclei (Figures 7 and S1), which is in line with the subcellular localisation of MKK4-MPK3 complexes in Arabidopsis protoplasts and Nicotiana leaves [15].
Figure 7. Protein-protein interaction studies in Tropaeolum majus tissue.
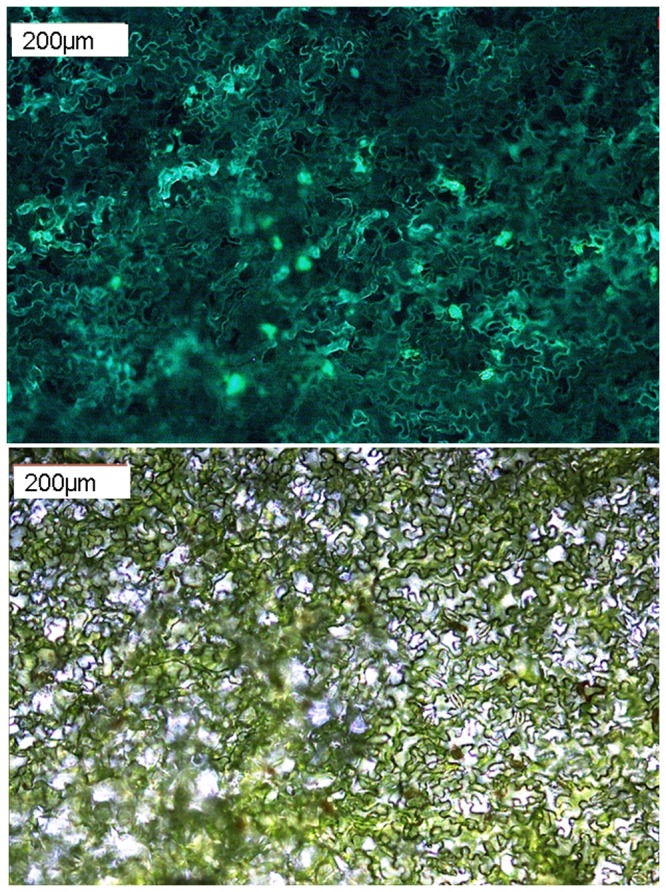
Leaves were infiltrated with Agrobacteria carrying constructs for the constitutive expression of MKK4 and MPK3, fused to the C- and N-terminal fragment of YFP, respectively. Bimolecular fluorescence complementation, documenting the interaction, was detected by UV microscopy, 5 dpi. top: UV image, bottom: bright-field image. For additional images see figure S1.
A number of downstream analyses in transient transformation experiments, such as immunoblotting, mass spectrometry and enzymatic assays involve the extraction of proteins. To compare Arabidopsis , N . benthamiana and nasturtium with respect to protein extractability, yield and profile, protein extracts from leaves of these species were prepared and separated by SDS-PAGE. Similar amounts of proteins were obtained (app. 15 µg/mg fresh weight). A dominant, approximately 55 kilo Dalton-sized band was contained in all samples. It most likely represents the large subunit of ribulose bisphosphate carboxylase. Expectedly, there were both similarities and differences in the protein profiles of the three species (Figure 8). As in Arabidopsis and N . benthamiana , nasturtium proteins are stable for at least 7 days (4°C storage and subsequent SDS-PAGE or catalase activity assay, not shown). In summary, functional T . majus . proteins can be isolated using standard plant protein extraction protocols.
Figure 8. Comparison of protein profiles in Arabidopsis , N . benthamiana and Tropaeolum majus .
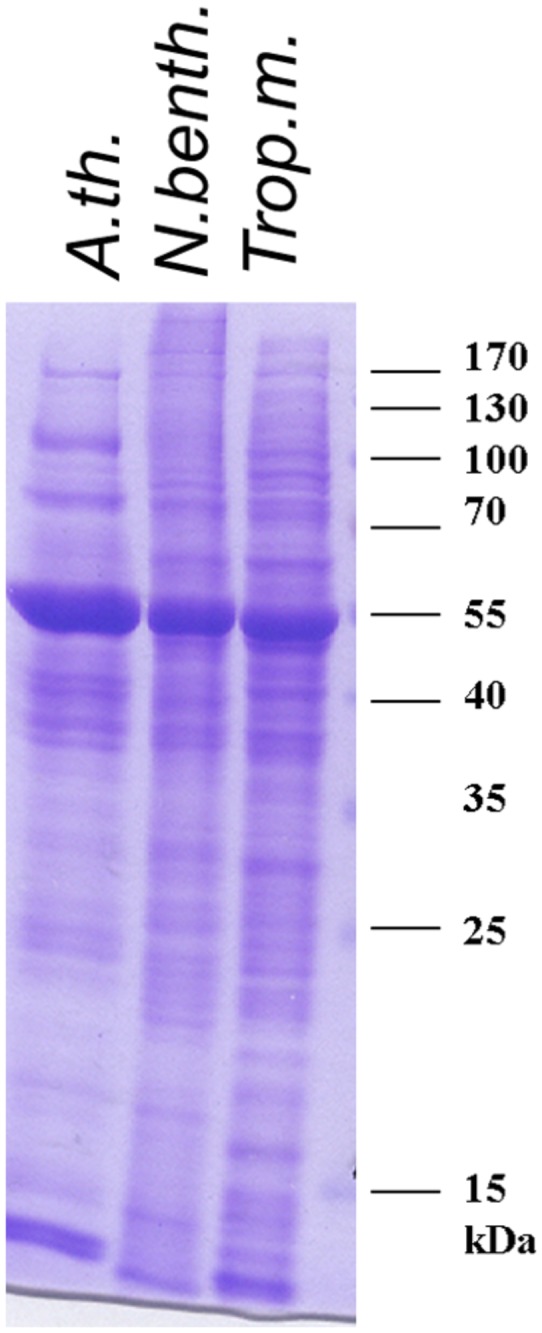
20 µg protein extracted from Arabidopsis rosette leaves, Nicotiana benthamiana or Tropaeolum majus leaves were separated by SDS-PAGE (12%) and visualised by Coomassie Blue staining.
Discussion and Conclusions
Tropaeolum and Arabidopsis are phylogenetically related. For unknown reasons, Arabidopsis “floral dipping” into suspensions of A. tumefaciens is an efficient and well-established method, while leaf infiltration yields poor transformation rates. Plants of the order Brassicales produce characteristic sulfur-rich metabolites, glucosinolates. These mustard-oil glycosides function in the defence against pests and pathogens [19], and might therefore be restrictive factor in Arabidopsis leaf transformation. However, in view of the data represented here, glucosinolates are not a barrier for agrobacterial infection. Likewise, this means that other species within the Brassicales may also be accessible to Agrobacterium -mediated leaf infiltration. Based on the above-listed characteristics Tropaeolum was studied here. Transgene expression in T . majus was transient, as it was clearly detectable 5-14 days post-infiltration, but not in leaves 3-weeks after infiltration. These expression kinetics are similar to those know from Nicotiana . To achieve stable transformation, resistance marker-based selection may be needed. It seems reasonable to assume that whole plants can be generated from transgenic cells via tissue-culturing of surface-sterilised leaves, a question that was not addressed here. I focussed on the advantages of Tropaeolum leaf infiltration as transient expression system, documenting its applicability for various experimental approaches in molecular biology. It offers a complimentary tool to established methods and combines some beneficial characteristics of Arabidopsis and the established Nicotiana transient expression systems (table 1). Tropaeolum and Arabidopsis genes are functionally replaceable, as had been shown for FAD2 encoding a fatty acid desaturase [20]. Research fields that may particularly benefit from the here-described expression system include those of xyloglucan metabolism [21], oleate biosynthesis [22] and endosymbiosis [23] and plant-insect interaction [24]. Given the health-promoting effects of Tropaeolum majus , including utinary tract protection [25,26], antidiuretic [27] and even anti-tumor activity [28], conquering this plant for genetic manipulations harbours potential for biotechnological and pharmacological applications. It may be employed as production platform of tailor-made glucosinolate compounds, similarly to sesquiterpenoid pharmaceutical production in tobacco [11].
Table 1. Overview on major features of Tropaeolum majus agroinfiltration as transient expression system.
| Taxonomic aspects | T . majus belongs in the order Brassicales, as does Arabidopsis . RNAi-based gene silencing with Arabidopsis -derived DNA constructs is more likely to succeed in Tropaeolum than in Nicotiana. |
| Experimental aspects | Plant growth |
| Modest space requirements | |
| Multiple plants grown in the same pot can be bound to the site (long flexible stems). | |
| Climate-controlled growth cabinet not required | |
| Transformation procedure | |
| simple and inexpensive | |
| Expanded leaves are equally suitable, no leaf-to-leaf variation observed. | |
| Infiltration requires some empirically acquired know-how. The infiltration liquid moves slowly, but gradually (30 sec/8 qcm). | |
| Transgene expression and kinetics | |
| Transformation efficiency is reproducibly high. > 80% of cells in the infiltrated area express the transgene (here tested for YFP, GUS, Luciferase). | |
| Expression detectable 4 days-post-infiltration (dpi), observed >10 dpi, (transient, kinetics comparable to Nicotiana ). | |
| Multiple gene transfer possible, e.g. for effector/reporter gene studies; interaction studies (here tested via BiFC assay). | |
| Limitations | |
| “Lotus effect”: Treatment with liquid effector substances (e.g. elicitors in stress research) may have to be applied by infiltration. | |
| Perspectives for basic and applied research | Relevant tool for basic research, e.g. |
| Endomycorrhizal interactions | |
| Biosynthesis and metabolism of glucosinolates and oleates | |
| Pharma industry and biotechnology | |
| May facilitate isolation of key components in biosynthesis of health-promoting substances (for transfer into other species; for in vitro synthesis etc.) | |
| Transgenic T . majus cell cultures may be established for large-scale production of desired substances and phyto-pharmaceuticals. |
Supporting Information
Microscopy images supporting the data shown in Figure 4 and Figure 7. T . majus leaves were infiltrated with agrobacterial suspensions to induce expression of luciferase (control, left), Yellow fluorescent protein (middle); or for protein–protein interaction studies (MKK4 and MPK3, fused to the C- and N-terminal fragment of YFP, respectively). Transgene expression was analysed 5 dpi by UV microscopy. Top: fluorescence; bottom: bright-field image.
(TIF)
Acknowledgments
I thank Prof. Dr. Lukas Mach for various support; and Helene Persak for technical assistance.
Funding Statement
AP gratefully acknowledges the Austrian Science Foundation for funding (Elise-Richter- Grant V167-B09). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Pitzschke A, Hirt H (2010) New insights into an old story: Agrobacterium-induced tumour formation in plants by plant transformation. EMBO J 29: 1021-1032. doi:10.1038/emboj.2010.8. PubMed: 20150897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Parinov S, Sundaresan V (2000) Functional genomics in Arabidopsis: large-scale insertional mutagenesis complements the genome sequencing project. Curr Opin Biotechnol 11: 157-161. doi:10.1016/S0958-1669(00)00075-6. PubMed: 10753770. [DOI] [PubMed] [Google Scholar]
- 3. Initiative AG (2000) Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 408: 796-815 [Google Scholar]
- 4. Li JF, Park E, von Arnim AG, Nebenführ A (2009) The FAST technique: a simplified Agrobacterium-based transformation method for transient gene expression analysis in seedlings of Arabidopsis and other plant species. Plants Methods 5: 6. doi:10.1186/1746-4811-5-6. PubMed: 19457242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Clough SJ, Bent AF (1998) Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J 16: 735-743. doi:10.1046/j.1365-313x.1998.00343.x. PubMed: 10069079. [DOI] [PubMed] [Google Scholar]
- 6. Rivera AL, Gómez-Lim M, Fernández F, Loske AM (2012) Physical methods for genetic plant transformation. Phys Life Rev 9: 308-345. doi:10.1016/j.plrev.2012.06.002. PubMed: 22704230. [DOI] [PubMed] [Google Scholar]
- 7. Rakoczy-Trojanowska M (2002) Alternative methods of plant transformation--a short review. Cell Mol Biol Lett 7: 849-858. PubMed: 12378268. [PubMed] [Google Scholar]
- 8. Wu FH, Shen SC, Lee LY, Lee SH, Chan MT et al. (2009) Tape-Arabidopsis Sandwich - a simpler Arabidopsis protoplast isolation method. Plants Methods 5: 16. doi:10.1186/1746-4811-5-16. PubMed: 19930690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Pitzschke A, Persak H (2012) Poinsettia protoplasts - a simple, robust and efficient system for transient gene expression studies. Plants Methods 8: 14. doi:10.1186/1746-4811-8-14. PubMed: 22559320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Andrews LB, Curtis WR (2005) Comparison of transient protein expression in tobacco leaves and plant suspension culture. Biotechnol Prog 21: 946-952. PubMed: 15932278. [DOI] [PubMed] [Google Scholar]
- 11. van Herpen TW, Cankar K, Nogueira M, Bosch D, Bouwmeester HJ et al. (2010) Nicotiana benthamiana as a production platform for artemisinin precursors. PLOS ONE 5: e14222. doi:10.1371/journal.pone.0014222. PubMed: 21151979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Piotrzkowski N, Schillberg S, Rasche S (2012) Tackling heterogeneity: a leaf disc-based assay for the high-throughput screening of transient gene expression in tobacco. PLOS ONE 7: e45803. doi:10.1371/journal.pone.0045803. PubMed: 23029251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Hellens RP, Edwards EA, Leyland NR, Bean S, Mullineaux PM (2000) pGreen: a versatile and flexible binary Ti vector for Agrobacterium-mediated plant transformation. Plant Mol Biol 42: 819-832. doi:10.1023/A:1006496308160. PubMed: 10890530. [DOI] [PubMed] [Google Scholar]
- 14. Djamei A, Pitzschke A, Nakagami H, Rajh I, Hirt H (2007) Trojan horse strategy in Agrobacterium transformation: abusing MAPK defense signaling. Science 318: 453-456. doi:10.1126/science.1148110. PubMed: 17947581. [DOI] [PubMed] [Google Scholar]
- 15. Persak H, Pitzschke A (2013) Tight interconnection and multi-level control of Arabidopsis MYB44 in MAPK cascade signalling. PLOS ONE 8: e57547. doi:10.1371/journal.pone.0057547. PubMed: 23437396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Voinnet O, Rivas S, Mestre P, Baulcombe D (2003) An enhanced transient expression system in plants based on suppression of gene silencing by the p19 protein of tomato bushy stunt virus. Plant J 33: 949-956. doi:10.1046/j.1365-313X.2003.01676.x. PubMed: 12609035. [DOI] [PubMed] [Google Scholar]
- 17. Walter M, Chaban C, Schütze K, Batistic O, Weckermann K et al. (2004) Visualization of protein interactions in living plant cells using bimolecular fluorescence complementation. Plant J 40: 428-438. doi:10.1111/j.1365-313X.2004.02219.x. PubMed: 15469500. [DOI] [PubMed] [Google Scholar]
- 18. Pitzschke A, Schikora A, Hirt H (2009) MAPK cascade signalling networks in plant defence. Curr Opin Plant Biol 12: 421-426. doi:10.1016/j.pbi.2009.06.008. PubMed: 19608449. [DOI] [PubMed] [Google Scholar]
- 19. Falk KL, Tokuhisa JG, Gershenzon J (2007) The effect of sulfur nutrition on plant glucosinolate content: physiology and molecular mechanisms. Plant Biol Stuttg 9: 573-581. doi:10.1055/s-2007-965431. PubMed: 17853357. [DOI] [PubMed] [Google Scholar]
- 20. Mietkiewska E, Brost JM, Giblin EM, Francis T, Wang S et al. (2006) A Tropaeolum majus FAD2 cDNA complements the fad2 mutation in transgenic Arabidopsis plants. Plant Sci 171: 187-193. doi:10.1016/j.plantsci.2006.03.006. [Google Scholar]
- 21. Crombie HJ, Chengappa S, Hellyer A, Reid JS (1998) A xyloglucan oligosaccharide-active, transglycosylating beta-D-glucosidase from the cotyledons of nasturtium (Tropaeolum majus L) seedlings--purification, properties and characterization of a cDNA clone. Plant J 15: 27-38. doi:10.1046/j.1365-313X.1998.00182.x. PubMed: 9744092. [DOI] [PubMed] [Google Scholar]
- 22. Xu J, Francis T, Mietkiewska E, Giblin EM, Barton DL et al. (2008) Cloning and characterization of an acyl-CoA-dependent diacylglycerol acyltransferase 1 (DGAT1) gene from Tropaeolum majus, and a study of the functional motifs of the DGAT protein using site-directed mutagenesis to modify enzyme activity and oil content. Plant Biotechnol J 6: 799-818. doi:10.1111/j.1467-7652.2008.00358.x. PubMed: 18631243. [DOI] [PubMed] [Google Scholar]
- 23. Markmann K, Giczey G, Parniske M (2008) Functional Adaptation of a Plant Receptor- Kinase Paved the Way for the Evolution of Intracellular Root Symbioses with Bacteria. PLOS Biol 6: e68. doi:10.1371/journal.pbio.0060068. PubMed: 18318603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Silva ME, Mussury RM, Vieira Mdo C, Alves Junior VV, Pereira ZV et al. (2011) Floral biology of Tropaeolum majus L. and its relation with Astylus variegatus activity (Germar 1824). An Acad Bras Cienc 83: 1251-1258. doi:10.1590/S0001-37652011005000046. PubMed: 22011767. [DOI] [PubMed] [Google Scholar]
- 25. Kleinwächter M, Schnug E, Selmar D (2008) The glucosinolate-myrosinase system in nasturtium (Tropaeolum majus L.): variability of biochemical parameters and screening for clones feasible for pharmaceutical utilization. J Agric Food Chem 56: 11165-11170. doi:10.1021/jf802053n. PubMed: 18986152. [DOI] [PubMed] [Google Scholar]
- 26. Lourenço EL, Muller JC, Boareto AC, Gomes C, Lourenço AC et al. (2012) Screening for in vivo (anti)estrogenic and (anti)androgenic activities of Tropaeolum majus L. and its effect on uterine contractility. J Ethnopharmacol 141: 418-423. doi:10.1016/j.jep.2012.03.004. PubMed: 22433534. [DOI] [PubMed] [Google Scholar]
- 27. Gasparotto A Jr., Boffo MA, Lourenço EL, Stefanello ME, Kassuya CA et al. (2009) Natriuretic and diuretic effects of Tropaeolum majus (Tropaeolaceae) in rats. J Ethnopharmacol 122: 517-522. doi:10.1016/j.jep.2009.01.021. PubMed: 19429322. [DOI] [PubMed] [Google Scholar]
- 28. Pintão AM, Pais MS, Coley H, Kelland LR, Judson IR (1995) In vitro and in vivo antitumor activity of benzyl isothiocyanate: a natural product from Tropaeolum majus. Planta Med 61: 233-236. doi:10.1055/s-2006-958062. PubMed: 7617765. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Microscopy images supporting the data shown in Figure 4 and Figure 7. T . majus leaves were infiltrated with agrobacterial suspensions to induce expression of luciferase (control, left), Yellow fluorescent protein (middle); or for protein–protein interaction studies (MKK4 and MPK3, fused to the C- and N-terminal fragment of YFP, respectively). Transgene expression was analysed 5 dpi by UV microscopy. Top: fluorescence; bottom: bright-field image.
(TIF)